
Abstract
TFIIH is a general transcription factor with kinase and helicase activities. The kinase activity resides in the Kin28 subunit of TFIIH. The role of Kin28 kinase in the early steps of transcription is well established. Here we report a novel role of Kin28 in the termination of transcription. We show that RNAPII reads through a termination signal upon kinase inhibition. Furthermore, the recruitment of termination factors towards the 3′ end of a gene was compromised in the kinase mutant, thus confirming the termination defect. A concomitant decrease in crosslinking of termination factors near the 5′ end of genes was also observed in the kinase-defective mutant. Simultaneous presence of termination factors towards both the ends of a gene is indicative of gene looping; while the loss of termination factor occupancy from the distal ends suggest the abolition of a looped gene conformation. Accordingly, CCC analysis revealed that the looped architecture of genes was severely compromised in the Kin28 kinase mutant. In a looping defective sua7-1 mutant, even the enzymatically active Kin28 kinase could not rescue the termination defect. These results strongly suggest a crucial role of Kin28 kinase-dependent gene looping in the termination of transcription in budding yeast.
Similar content being viewed by others
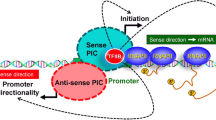


Introduction
The kinase subunit of TFIIH, Kin28, phosphorylates carboxy-terminal domain (CTD) of RNAPII at serine-5 and serine-7 residues1. The Kin28-mediated phosphorylation of serine-5 takes place immediately after initiation and is crucial for promoter clearance2,3,4,5. The serine-5 phosphorylation is also required for the recruitment of the capping enzyme for capping of mRNA at the 5′ end6,7,8. The function of serine-7 phosphorylation in transcription of protein coding genes is not clear yet1,9,10
Our initial understanding of the function of TFIIH kinase under physiological conditions has come from studies with the temperature-sensitive mutants of Kin28 in budding yeast. The shifting of Kin28 mutants to the elevated temperature adversely affected the recruitment of TFIIH complex at the promoter resulting in a dramatic decrease in the CTD-serine-5 phosphorylation and a concomitant decrease in the level of steady state mRNA level11,12. The temperature-sensitive mutation has been found to affect the catalytic activity of Kin28 kinase as well as its interaction with other subunits of TFIIH11,13,14. To determine the specific role of Kin28 kinase in transcription by RNAPII, the ATP-binding pocket of the enzyme was engineered to make it respond to the specific inhibitor NA-PP1, which is an analog of ATP. In the presence of NA-PP1, the kinase activity of analog-sensitive Kin28 mutant (Kin28-as) is almost completely inhibited without affecting its interaction with the subunits of TFIIH complex15. The studies using Kin28-as mutant revealed that the kinase activity is neither required for recruitment of TFIIH at the promoter region, nor it is essential for transcription5,12,14,16. A global decrease in the transcript level in the absence of Kin28 kinase activity, however, was observed12. This decrease in mRNA level in the Kin28-as mutant was attributed to the effect of serine-5 phosphorylation on the capping of mRNA at the 5′ end rather than a direct role of Kin28 kinase in transcription. This view has been challenged by recent studies, which demonstrated a direct role of Kin28 kinase in transcription4,5. These studies implicated serine-5 phosphorylation in release of Mediator complex from the promoter-proximal region following initiation of transcription, thereby facilitating promoter clearance.
A number of genes in budding yeast exhibit a physical interaction of the terminator region of the gene with its cognate promoter resulting in the formation of looped gene architecture17,18,19,20,21,22,23,24,25,26,27,28. Formation of such transcription-dependent gene loops has been observed in higher eukaryotes as well29,30,31,32,33. Gene looping has been implicated in transcription directionality, reinitiation of transcription, transcription memory, intron-mediated transcriptional enhancement and termination of transcription17,19,20,22,23,27,28,29,34. Kin28 has been shown to affect gene looping in budding yeast24. It was however not clear in this study if kinase activity of Kin28 is required for gene loop formation. Also, the physiological significance of Kin28-mediated gene looping remained unclear.
To investigate the precise role of Kin28 kinase in transcription by RNAPII, we examined the transcription of a number of inducible and constitutively expressed genes in Kin28-as mutant. Our results suggest that Kin28 kinase is not essential for transcription, but is necessary for the optimal transcription of both inducible and non-inducible genes. Kin28 crosslinked to both the 5′ and 3′ ends of transcriptionally engaged genes. In the absence of kinase activity, localization of Kin28 towards the 3′ end of genes exhibited a dramatic decline. The delocalization of Kin28 from the 3′ end coincided with the polymerase reading through the termination signal with a concomitant loss of looped gene conformation. These results implicate Kin28 kinase activity both in termination of transcription as well as gene looping. We propose that Kin28 kinase-mediated gene looping facilitates termination of transcription in at least a subset of genes in budding yeast.
Results
Kin28 kinase is required for optimal transcription of both inducible and constitutively expressed genes
Although CTD-kinase activity of Kin28 is not essential either for transcription or for the survival of yeast cells, growth of cells is severely inhibited in the kinase-defective mutant5,12,14,15. These results suggest that either Kin28 kinase is affecting the transcription cycle in a subtle way, or it is playing a role in a yet unknown aspect of cellular dynamic14. To further probe the function of Kin28 kinase in transcription by RNAPII, we used the analog-sensitive Kin28-as mutant described earlier14,15. In the presence of 5–10 μM NA-PP1 analog in the growth medium, the CTD kinase activity of Kin28-as mutant is almost completely inhibited in less than 60 minutes14.
To investigate the precise function of Kin28 kinase in transcription cycle, we examined transcription of five inducible (HXT1, MET16, CHA1, GAL10 and INO1) and six constitutively expressed genes (ACT1, ASC1, IMD4, MSN5, SPC1 and CMP1) in Kin28-as mutant in the presence and absence of NA-PP1. The transcription was monitored in terms of steady state mRNA level determined by RT-PCR approach. In the presence of NA-PP1, transcription of both inducible and constitutively expressed genes exhibited a decline by about 2–10 folds in the mutant (Supplemental Figs S1 and S2). No such decrease in RNA level was observed in the isogenic wild type strain in the presence of NA-PP1 or in the Kin28-as strain in the absence of NA-PP1. Our results are in disagreement with another study that did not find a significant decrease in the global mRNA level upon inhibition of Kin28 kinase using the same kin28-as mutant14. Using the identical chemical genetic approach, however, another group reported a drastic decrease in mRNA level of a majority of yeast genes upon inhibition of Kin28 kinase12. The discrepancy in the result was due to the use of a different normalization control and is discussed in detail in Hong et al.12. Our results are in agreement with the findings of Hong et al.12.
The Kin28 kinase activity is essential for capping of mRNA at the 5′ end of genes35. Since capping of mRNA has been shown to affect the stability of mRNA, it was possible that the observed decrease in mRNA level of genes in the absence of kinase activity was not due to the effect of kinase on transcription, but on the stability of transcripts. To clarify the issue, we checked the nascent transcription of two inducible genes CHA1 and HXT1, as well as two constitutively expressed genes ACT1 and ASC1, by transcription run-on (TRO) approach. TRO assay is a better indicator of transcriptional activity of a gene than RNAPII density ChIP as it measures nascent transcription of a gene. The Kin28-as cells were grown to the mid-log phase and transcription was induced in the presence and absence of NA-PP1 as described in Kanin et al.14. The strand-specific TRO analysis was carried out using Br-UTP by the modification of method described in Core et al., (2008)34. The results show that the nascent transcription of CHA1 and HXT1 decreased by about 2 fold in the presence of NA-PP1 (Fig. 1B,C). The nascent transcription of two constitutively expressed genes ACT1 and ASC1 also registered a steady 3-4 fold decline in the absence of Kin28 kinase activity (Fig. 1D,E). Thus, Kin28 kinase has a direct effect on the transcription of both inducible and non-inducible genes.

Kin28 kinase inhibition reduces nascent transcription and results in polymerase reading through the termination signal.
(A) Schematic depiction of a gene showing the positions of three primers 1, 2, 3, 4, 5, 6, 7 and 5′ primer used for cDNA synthesis following TRO procedure. (B–E) Quantification of RNA levels detected following TRO analysis in Kin28-as mutant in the absence (-NA-PP1) and presence (+NA-PP1) of NA-PP1. The transcript level of 18S was used as the control for normalization of results.
Kin28 kinase is required for termination of transcription
The strand-specific TRO analysis of genes using Br-UTP in the Kin28-as strain revealed a rather unexpected role of Kin28 in RNAPII transcription cycle. In all four genes that we tested, polymerase read through the termination signal in the absence of Kin28 kinase activity thereby implying a defect in transcription termination of these genes (Fig. 1). TRO analysis revealed a very weak or no detectable polymerase signal in the regions 3, 4, 5 and 6 located downstream of the poly(A)-site of CHA1 and HXT1 in the mutant in the absence of NA-PP1 (Fig. 1B,C, black bars). In the presence of NA-PP1, however, there was a dramatic increase in TRO signal in the downstream regions 3-6 of both CHA1 and HXT1 (Fig. 1B,C, white bars). Similar results were observed for the constitutively expressed genes ACT1 and ASC1 (Fig. 1D,E). Thus, RNAPII was not able to read the termination signal of CHA1, HXT1, ACT1 and ASC1 efficiently under kinase-defective condition and continued transcribing the downstream regions. The TRO approach using Br-UTP has been used previously to demonstrate the transcription termination defect in mammalian cells as well36,37. The downstream TRO signals observed here are not the consequence of transcription of non-coding RNAs like SUTs and CUTs initiating from the vicinity of the poly(A) site as the RNA reverse transcribed using primers 1-7 were PCR amplified using a forward 5′ primer located in the promoter proximal region as shown in Fig. 1A. To corroborate the terminator readthrough phenotype of Kin28-as mutant, we performed RNAPII-density ChIP for ACT1 and CHA1 in the presence and absence of NA-PP1. The polymerase readthrough the poly(A) signal of both genes under kinase defective condition thereby affirming the termination defect observed using TRO approach (Supplemental Fig. S3). These results strongly suggest a role for Kin28 kinase in termination of transcription.
To further probe the role of Kin28 kinase in termination, we checked the recruitment of CF1 and CPF termination complexes towards the 3′ end of genes in the kinase defective mutant. We expected that if Kin28 kinase activity is required for termination of transcription, the recruitment of either CF1 or CPF complexes may be adversely affected under the kinase defective condition. We used the ChIP approach to monitor the recruitment of CF1 complex towards the 3′ end of genes using its Rna15 subunit, while Ssu72 subunit was used to detect the recruitment of the CPF complex in the mutant strain. ChIP analysis revealed that both Rna15 and Ssu72 occupied the terminator region of all four genes in the mutant in the absence of NA-PP1 (Fig. 2, region T, black bars). In the presence of NA-PP1, however, crosslinking of Rna15 to the 3′ end decreased by about 50-80% (Fig. 2B, region T, white bars), while that of Ssu72 declined by more than 75% (Fig. 2C, region T, white bars). Thus, the recruitment of both CF1 and CPF complexes towards the terminator region of CHA1, HXT1, ACT1 and ASC1 was compromised under the kinase defective condition. Taken together these results strongly suggest a novel role for Kin28 kinase in termination of transcription for at least a subset of genes in budding yeast.

Rna15 and Ssu72 occupancy of the promoter and terminator regions is adversely affected in the absence of Kin28 kinase activity.
(A) Schematic depiction of a gene indicating the position of the promoter (P), coding (M) and terminator (T) regions amplified by PCR in the ChIP assay. (B,C) Quantification of ChIP signals showing crosslinking of Rna15-TAP or Ssu72-TAP to different regions in kin28-as mutant in the absence of NA-PP1 (-NA-PP1) or in the presence of NA-PP1 (+NA-PP1). Input represents DNA prior to immunoprecipitation. Error bars represent one unit of standard deviation.
Kin28 contacts and affects phosphorylation of CTD at the 3′ end of genes
The experiments described above firmly established the role of Kin28 kinase in termination of transcription for at least some yeast genes. To have an insight into the role of Kin28 kinase in termination of transcription, we checked if Kin28 brings about termination by physically interacting with the 3′ end of genes and if the kinase activity is required for this interaction. ChIP was performed in a strain bearing TAP-tagged version of Kin28-as allele, in the presence and absence of NA-PP1. As expected, Kin28 was recruited towards the 5′ end of CHA1, HXT1, ACT1 and ASC1 in the absence of NA-PP1 (Fig. 3B–E, region P, black bars). In the presence of NA-PP1, however, crosslinking of Kin28 to the promoter region registered a 50-80% decline (Fig. 3B–E, region P, white bars). Interestingly, Kin28 was also found localized near the 3′ end of genes (Fig. 3B–E, region T, black bars). A genomewide analysis has also found Kin28 crosslinked to the 3′ end of a number of transcriptionally active genes in yeast38. It was, however, not clear from this study if the recruitment of Kin28 near the terminator region required its kinase activity. Our results show that the crosslinking of Kin28 towards the terminator region of genes was significantly reduced in the absence of its kinase activity (Fig. 3B–E, region P, white bars). The Kin28 ChIP signal at the 3′ end of CHA1 and ACT1 decreased by nearly 50%, while at HXT1 and ASC1 a more than 75% decline was observed under kinase-defective condition (Fig. 3B–E). When Kin28 ChIP signal was normalized with the RNAPII signal, we found no net decrease in Kin28 near the 5′ end of genes in the absence of kinase activity. Kin28 signal at the 3′ end, however, still registered a significant decline under kinase-defective condition (Supplemental Fig. S4). These results suggest that the presence of Kin28 at the 3′ end of genes is essential for its termination function.

Kin28 occupancy of the promoter and terminator regions is compromised in the absence of Kin28 kinase activity.
(A) Schematic depiction of a gene indicating the position of the promoter (P), coding (M) and terminator (T) regions PCR amplified in the ChIP assay. (B–E) Quantification of ChIP signals showing crosslinking of Kin28 to the promoter, coding and terminator regions of genes in kin28-as mutant in the absence of NA-PP1 (-NA-PP1) or in the presence of NA-PP1 (+NA-PP1). Input represents DNA prior to immunoprecipitation.
To confirm the recruitment of Kin28 towards the 3′ end of genes, we examined the phosphorylation status of CTD-serine-5 and CTD-serine-7 near the 3′ end of CHA1 and ACT1 in the mutant strain. Although phosphorylation of serine-5 near the promoter-proximal region is well established, there are conflicting reports regarding its phosphorylation near the 3′ end of genes. A few recent studies demonstrated phosphorylation of serine-5 at the 3′ end of a subset of yeast genes39,40. Our results are in agreement with these reports. We found elevated levels of serine-5 phosphorylation at the 3′ end of both CHA1 and ACT1 (Supplemental Figs S5 and S6, region T, black bars). A similar elevated level of serine-7 phosphorylation was observed near the terminator region of both genes (Supplemental Figs S5 and S6, region T, black bars). In the presence of NA-PP1, however, a 2-4 fold decrease in phosphorylation of both serine-5 as well as serine-7 was observed for two genes in the analog-sensitive mutant (Supplemental Figs S5 and S6, region T, white bars). As expected, no appreciable decrease in the level of serine-2 phosphorylation was observed towards the 3′ end of either CHA1 or ACT1 in the absence of Kin28 kinase activity (Supplemental Figs S5 and S6, region T).
The overall conclusion of these results is that the recruitment of Kin28 towards the 3′ end of genes as well as Kin28-mediated termination of transcription are both dependent on its kinase activity in budding yeast.
Kin28 physically interacts with the CF1 and CPF complexes
The results described above ruled out the possibility of Kin28 kinase playing an indirect role in termination of transcription by affecting CTD-serine-2 phosphorylation. Since inactivation of Kin28 kinase resulted in lowering of both CTD-serine-5 and serine-7 phosphorylation near the 3′ end of genes, the possibility of serine-5 and serine-7 phosphorylation playing a role in transcription termination cannot be ruled out. These results, however, raised the possibility of a direct role of Kin28 in transcription termination by interacting with the CF1 and CPF complexes and facilitating their recruitment towards the 3′ end of genes. We therefore examined the physical interaction of Kin28 with the CF1 and CPF complexes by coimmunoprecipitation approach. The TAP-tagged Kin28 was pulled down using IgG-Sepharose beads and the presence of either Rna15 or Ssu72 was detected in the immunoprecipitated fraction. Our results show that Kin28 interacts with both CF1 subunit Rna15 and CPF subunit Ssu72 in the absence of NA-PP1 (Fig. 4A,B, lane 2). In the presence of NA-PP1, however, Kin28 interaction with both RNA15 and Ssu72 was completely abolished (Fig. 4A,B, lane 4). These results show that Kin28 interacts with both the CF1 and CPF complexes and this interaction is dependent on its kinase activity. Whether the Kin28-CF1 and Kin28-CPF interactions are facilitated by the Kin28-mediated phosphorylation of CTD or that of a subunit of CF1 or CPF complex needs further investigation. The possibility of Kin28 influencing termination both indirectly by affecting the CTD-serine-5 phosphorylation in the terminator region and directly by interacting with the termination factors near the 3′ of the gene also cannot be ruled out.

Kin28 physical interacts with Rna15 and Ssu72 in a manner dependent on its kinase activity.
(A) Western blot analysis showing coimmunoprecipitation of Rna15-Myc with TAP-tagged Kin28 in Kin28-as mutant in the absence (-NA-PP1) and presence (+NA-PP1). (B) Western blot analysis showing coimmunoprecipitation of Ssu72-Myc with TAP-tagged Kin28 in Kin28-as mutant in the absence (-NA-PP1) and presence (+NA-PP1). Kin28 was detected by anti-CBP antibodies, while Rna15 and Ssu72 were detected by anti-Myc antibodies. As a control, immunoprecipitation was performed from cells without TAP-tagged Kin28. IP, immunoprecipitation.
Kin28 kinase is required for gene looping
We have previously demonstrated simultaneous crosslinking of general transcription factors and termination factors to both the 5′ and 3′ ends of a transcriptionally active gene18,21,34,41. We also showed that the simultaneous terminal end occupancy of initiation and termination factors is accompanied by a physical interaction of the promoter and terminator regions of a gene resulting in the formation of looped gene architecture18,21,34,41. We found a similar crosslinking of Rna15, Ssu72 and Kin28 to both the ends of all four genes tested in this analysis in the absence of NA-PP1 (Fig. 2). In the presence of NA-PP1, however, in addition to loss of 3′ end occupancy, 5′ end occupancy of both Rna15 and Ssu72 registered a 30-90% decline. The crosslinking of Kin28 to both the ends of a gene similarly decreased in the absence of its kinase activity. These results strongly suggested that all six genes examined here assumed a looped conformation in a manner dependent on Kin28 kinase activity. We therefore carried out CCC analysis of CHA1, HXT1, ACT1 and ASC1 in Kin28-as cells in the presence and absence of NA-PP1. We have previously used this approach to demonstrate looping of genes in a transcription-dependent manner18. In CCC assay, the physical interaction of the promoter and terminator regions of a gene is converted into a PCR product obtained using primers flanking the promoter (P1 primer) and the terminator (T1) regions as shown in Fig. 5 (top panel)42. CCC analysis revealed that CHA1, HXT1, ACT1 and ASC1 assume a looped gene conformation during transcription in the Kin28-as mutant as indicated by a strong P1T1 PCR signal for all four genes (Fig. 5A-D, -NA-PP1 lane and black bars). In the presence of NA-PP1, however, the looped gene architecture of all four genes was abrogated as there was a 3–40 fold decline in P1T1 PCR signal for different genes (Fig. 5A-D, +NA-PP1 lane and white bars). A logical conclusion of these results is that the kinase activity of Kin28 is essential for gene looping.

Gene looping is compromised in the absence of Kin28 kinase activity.
Top panels in (A–D) represent a scaled schematic depiction of genes indicating the positions of AluI and NlaIV restriction sites (vertical lines) and the PCR primers (arrows) used for CCC analysis. (A – D), CCC analysis of CHA1, HXT1, ACT1 and ASC1 in kin28-as mutant in the absence of NA-PP1 (-NA-PP1) or in the presence of NA-PP1 (+NA-PP1). P1T1 PCR reflects the looping signal and the C1C2 PCR reflects the loading control indicating that an equal amount of template DNA was used in each CCC PCR reaction. The gel figures shown here are cropped to indicate the bands specific for P1T1 and C1C2 PCR signals used in quantification.
Kin28 kinase-dependent gene looping facilitates termination of transcription
The results described above clearly demonstrate that both termination of transcription and gene looping are dependent on the Kin28 kinase activity. The observation raises the possibility of Kin28 kinase bringing about termination of transcription through gene looping. If gene looping is indeed facilitating termination of transcription, then disruption of gene looping by mutating a factor other than Kin28 will also lead to a termination defect. We therefore used the looping defective TFIIB mutant sua7-1 to check our hypothesis. In the sua7-1 mutant, the recruitment of TFIIB at the promoter and the initiation of transcription remain unaffected. The 3′ end occupancy of TFIIB, however, is severely compromised in the mutant26. Consequently, the sua7-1 mutant is defective in gene looping26. We have previously used this strain to demonstrate that Srb5-dependent gene looping facilitates termination of transcription of a subset of genes23. To determine if Kin28 kinase-mediated gene looping is similarly affecting termination of transcription, we performed strand-specific TRO analysis of CHA1, HXT1, ACT1 and ASC1 in the looping defective sua7-1 strain. Our results show that the polymerase reads through the terminator region of all four genes in the looping defective strain (Supplemental Figs 7B, 7C, 7D and 7E, white bars). No such read-through was observed in the isogenic wild type strain (Supplemental Figs 7B, 7C, 7D and 7E, black bars). This termination defect occurred even in the presence of enzymatically active Kin28. Furthermore, crosslinking of Kin28 towards the 3′ end of genes was severely compromised in sua7-1 cells (Supplemental Fig. S8). On the basis of these results, we conclude that it is not Kin28 kinase activity per se, but the kinase-mediated gene looping that facilitates termination of transcription.
Discussion
Since its discovery, the function of CTD kinase activity of TFIIH in transcription has been the focus of intense scrutiny. Using the analog-sensitive mutant of Kin28, it has been demonstrated that Kin28 kinase is neither essential for transcription nor for survival of yeast cells12,14. Our results suggest that Kin28 kinase is not an absolute requirement for transcription, but is required for optimal transcription of genes. The prevailing view is that the general transcription factors are required for the transcription of a vast majority of RNAPII-dependent genes43,44,45. This may not be entirely true. A recent study revealed that the TFIIB is required for transcription of only a subset of genes in humans46. It is possible that the Kin28 kinase activity of TFIIH is also not necessary for transcription of all RNAPII-transcribed genes. If the TFIIH-kinase is required for transcription of a subset of non-essential genes, the cell may still be viable in the absence of the kinase activity, but the cell fitness may be adversely affected. This may explain why a defect in Kin28 kinase does not affect the cell viability and global poly(A)-mRNA level drastically, but is still necessary for normal growth of yeast cells. Alternatively, Kin28 kinase may be affecting transcription of protein coding genes on a genomewide scale, but compensatory mechanisms such as a parallel decrease in the mRNA degradation rate is buffering the global transcript level47,48,49,50.
The Kin28 kinase occupies both the 5′ and the 3′ ends of a number of genes51,52. Most studies have focused on the role of TFIIH kinase at the 5′ end of genes. Here we demonstrate a novel role of Kin28 kinase in termination of transcription. We provide several lines of evidence in support of our claim. First, RNAPII reads through the termination signal into the downstream region under the kinase-defective condition (Fig. 1). Second, recruitment of CF1 and CPF termination complexes towards the terminator end of gene is compromised in the absence of kinase activity (Fig. 2). Third, localization of Kin28 at the 3′ end of genes is dependent on the kinase activity of protein (Fig. 3). Genomewide RNAPII ChIP analysis has been performed in the presence and absence of Kin28 kinase activity in budding yeast5,53. These studies revealed the role of Kin28 kinase in the promoter escape step of transcription of a subset of genes. To corroborate the role Kin28 kinase in termination of transcription, we compared our results with the genomewide data published in these two studies. We extracted the RNAPII-ChIP data for four genes used in our study; CHA1, HXT1, ACT1 and ASC1, from Struhl’s published genomewide database (Supplemental Fig. S9). All four genes exhibit a decrease in polymerase density in the coding region in the absence of Kin28 (Supplemental Fig. S9). More importantly, there was about 1.5 to 2.5 fold increase in the polymerase density beyond the 3′ end of all four genes in the absence of Kin28 (Supplemental Fig. S9). These results are in broad agreement with the analysis presented in this study. Even CHA1, which is expressed at a very low level under the condition Struhl’s ChIP-Seq analysis was performed, exhibited an increase in polymerase signal towards the 3′ end of the gene in the absence of Kin28 (Supplemental Fig. S9 A). We, however, could not find a correlation between Kin28-dependent termination and the SAGA-dependence of genes. In Bentley’s study, we found a slightly higher RNAPII signal near the 3′ end of long genes in the absence of Kin28 kinase activity53. Whether the higher polymerase density beyond the 3′ end of long genes in the absence of Kin28 kinase reflects a termination defect, however, needs further investigation. Inhibition of Cdk7, which is the human homologue of Kin28, similarly affects termination of transcription of several genes54. Thus, the role of TFIIH kinase in termination could be an evolutionarily conserved feature of transcription by RNAPII in eukaryotes.
TFIIH is not the only general transcription factor that has been implicated in termination of transcription. A similar termination function has been found for TFIIB as well. Just like Kin28, TFIIB cross-links to the 3′ end of genes and facilitates recruitment of the termination factors there26. Although, a role of TFIIB in termination has yet to be established in budding yeast, it has already been demonstrated in mammals and flies55,56. We have recently demonstrated the role of another initiation factor, Mediator complex, in the termination of transcription23,41. The emerging view is that the initiation and termination factors do not have exclusive roles in the initiation and termination steps of transcription respectively. We have shown that at least some initiation factors participate in the termination of transcription, while additionally, some termination factors function in the initiation/reinitiation of transcription34.
We have previously demonstrated that gene looping facilitates interaction of the promoter-bound factors with the 3′ end of genes and of terminator-bound factors with the 5′ end of genes18,21,23,34. We hypothesize that it is gene looping that allows a transcription factor to function at both the ends of a gene. The termination function of TFIIB in flies and mammalian system is completely dependent on its interaction with the 3′ end of a gene55,56. Here we show that Kin28 crosslinking to the 3′ end of a gene and its consequent role in termination of transcription may also be dependent on gene looping. Previously, we have shown that Srb5-dependent gene looping helps in termination of transcription41. We have also shown that gene looping is lost in the mutants of termination factors that are defective in termination21,34. On the basis of these results we propose that gene looping is playing a general role in termination of transcription in budding yeast. Accordingly, we show that TFIIB-dependent gene looping similarly affects the termination of transcription of the same set of genes that are affected by Kin28 kinase. Although, gene looping has been shown for only a few genes in higher eukaryotes, TFIIB similarly crosslinks to the 3′ end of genes in mammalian cells as well and facilitates termination of transcription56. The crosslinking of TFIIB to both the ends of genes in mammalian systems strongly suggest that genes are in looped conformation. The possibility of gene looping playing a similar role in termination of transcription in higher eukaryotes cannot be ruled out.
Methods
Yeast Strains
The yeast strains used in this study are listed in supplemental Table S1. Strains BPM5, SAM89 and SAM90, which contain C-terminal TAP-tagged TFIIH subunits Tfb4, Kin28 and Ssl2 respectively, were constructed by transforming the wild type strain BY4733 with DNA that was PCR amplified from plasmid pBS1539 (URA marker). Strains SAM99, SAM107 and ZD3, which contain C-terminal TAP-tagged TFIIH subunits Kin28, Ssl2 and Tfb4 respectively, were constructed by transforming the Kin28as mutant yFR763 with DNA that was PCR amplified from plasmid pBS1479 (TRP marker). The C-terminal Myc-tagged strains SAM93, SAM51, SAM94, SAM103, SAM101, SAM102, SAM104 and SAM105 which contain Myc tagged Rna15 or Ssu72, respectively, were created by transforming either wild type or Kin28as cells with DNA that was PCR amplified from plasmid pFA6-13Myc-HIS3mx6. Strain SAM106, which contains a C-terminal Myc tagged Rpb4 was created by transforming Kin28as cells with DNA that was PCR amplified from plasmid pFA6-13Myc-HIS3mx6. Strain AS1 was derived from BY4733 as described in57.
Cell Culture and Kinase Inhibition with NA-PP1
Kin28-as mutant cells were grown in uracil-depleted media to an A600 0.6 as previously described21. Cell cultures were then divided evenly and transferred to 100 ml of fresh media that either contained 25 μl of DMSO (control) or DMSO + NAPP1 (Final Concentration 7 μM NA-PP1). The induction and inhibition of kinase activity occurred simultaneously for the inducible genes. The cells were grown under inducing and +/– NAPP1 conditions for either 30, 60 or 90 minutes during the time course experiments. In all other experiments, the cells were grown under inducing +/– NAPP1 conditions for one hour. In figures that depict a time 0 measurement, an equal aliquot of cells was removed prior to induction and kinase inhibition. For constitutively expressed genes, cells were grown to A600 0.6 and then grown in the presence and absence of NA-PP1 as described above.
Capture Chromosome Conformation Assay (CCC)
CCC experiments were performed as described previously42.
Chromatin Immunoprecipitation (ChIP)
ChIP experiments were performed as described previously21.
Transcription Analysis
Transcription analysis was performed by the RT-PCR approach as described previously18.
Strand-Specific ‘Transcription Run On’ (TRO) Assay
The strand specific ‘Transcription Run-On’ (TRO) assay was performed by the modification of method described in Core et al.58. The protocol can be divided into three steps:
Step I. Labeling and isolation of nascent RNA with Brd-UTP
Cells were grown as described above. The cell pellet was washed with 10 ml of ice cold TMN buffer (10 mM Tris-HCl pH 7.5, 5 mM MgCl2, 100 mM NaCl) and resuspended in 940 μl of DEPC (Diethylpyrocarbonate)-treated ice cold water. Cells were permeabilized by the incubating with 60 μl 10% sarkosyl at 4 °C for 25 minutes with gentle shaking. Permeabilized ells were spun down by centrifugation at 1.2×g for 6 minutes at 4 °C. The cells were resuspended in 150 μl of transcription elongation reaction buffer (50 mM Tris-HCl pH 7.5, 100 mM KCl, 10 mM MgCl2, 2 mM DTT, 0.75 mM each of ATP, CTP, GTP and Brd-UTP, 5 μl of RNAse inhibitor cocktail from NEB) and incubated at 30 °C for 5 minutes. Cells were immediately lysed by addition of 500 μl ice cold Trizol and 250 μl acid-washed glass beads followed by vigorous shaking at 4 °C for 20 minutes. The lysate was transferred to a new 1.5 ml microfuge tube containing 500 μl of Trizol and 200 μl of chloroform. The tubes were shaken vigorously and centrifuged at 14000 rpm at 25 °C for 20 minutes. The supernatant was extracted with phenol-chloroform three times. Total RNA was precipitated by incubating with NaCl overnight at −20 °C. The RNA pellet was washed once with ice cold 70% ethanol and resuspended in 100 μl of DEPC-treated water. RNA was further purified using the QIAGEN RNA Easy kit and eluted twice with 50 μl of DEPC treated water.
Step II. Purification of Brd-UTP-labeled RNA
About 25 μl of anti-BrdU-conjugated agarose beads were washed three times with 500 μl of binding buffer (0.25× SSPE buffer, 1 mM EDTA, 0.05% Tween20, 37.5 mM NaCl). Beads were blocked using 500 μl of blocking buffer (485 μl binding buffer containing 5 μl of 10% polyvinylpyrolidone, 10 μl of Ultrapure BSA, Sigma) by gently shaking for 1 hour at 4 °C. Beads were washed two times with 500 μl of binding buffer and then 400 μl of binding buffer was added to the beads and placed on ice.
The RNA from step I was incubated at 65 °C water bath for 5 minutes and immediately placed on ice for 2 minutes. The RNA was then incubated with the processed anti-BrdU beads with gentle shaking for 2 hours at 4 °C. Beads were then washed sequentially with 500 μl of binding buffer, 500 μl of low salt buffer (0.2× SSPE, 1 mM EDTA, 0.05% Tween20) 500 μl of high salt buffer (0.25× SSPE, 1 mM EDTA, 0.05% Tween-20, 100 mM NaCl) and 500 μl of TET buffer (1×TE buffer, 0.05% Tween20). RNA was eluted two times with 150 μl of elution buffer (20 mM DTT, 150 mM NaCl, 50 mM Tris-HCl pH 7.5, 1 mM EDTA, 0.1% SDS) by incubating at 42 °C for 5 minutes followed by a final elution with 200 μl of elution buffer at 42 °C for 5 minutes. After phenol-chloroform extraction, RNA was precipitated overnight at -20 °C in the presence of NaCl. RNA pellet was resuspended in 26 μl of DEPC-treated water and quantified using a Nanodrop spectrophotometer. RNA concentration was adjusted to 50 ng/μl.
Step III. Reverse transcription and PCR amplification of RNA
The cDNA synthesis was performed with Protoscript II (NEB) using the strand specific primers following to manufacturer’s recommendation. PCR amplification was performed using either Advantage polymerase or Taq polymerase depending on amplicon size. Quantification was done as described in Medler et al., (2011).
Coimmunoprecipitation (CO-IP)
CO-IP was performed as described previously21.
Western Blot Analysis
Western blotting was performed as described previously21. Anti-CBP and anti-Myc antibodies were from Upstate Biotechnology. Anti-CTD antibodies 3e10, 3e8 and 4e12 that detect CTD-serine-2-P, CTD-serine-5-P and CTD-serine-7-P respectively were purchased from Millipore.
Data Analysis
The data shown in figures is the result of at least four biological replicates. The data was quantified and statistical analysis was performed as described in42. Error bars represent one unit of standard deviation.
Additional Information
How to cite this article: Medler, S. and Ansari, A. Gene looping facilitates TFIIH kinase-mediated termination of transcription. Sci. Rep. 5, 12586; doi: 10.1038/srep12586 (2015).
References
Kim, M., Suh, H., Cho, E. J. & Buratowski, S. Phosphorylation of the yeast Rpb1 C-terminal domain at serines 2, 5 and 7. J Biol Chem 284, 26421–6 (2009).
Buratowski, S. Progression through the RNA polymerase II CTD cycle. Mol Cell 36, 541–6 (2009).
Luse, D. S. & Samkurashvili, I. The transition from initiation to elongation by RNA polymerase II. Cold Spring Harb Symp Quant Biol 63, 289–300 (1998).
Jeronimo, C. & Robert, F. Kin28 regulates the transient association of Mediator with core promoters. Nat Struct Mol Biol 21, 449–55 (2014).
Wong, K. H., Jin, Y. & Struhl, K. TFIIH Phosphorylation of the Pol II CTD Stimulates Mediator Dissociation from the Preinitiation Complex and Promoter Escape. Mol Cell 54, 601–12 (2014).
Cho, E. J., Takagi, T., Moore, C. R. & Buratowski, S. mRNA capping enzyme is recruited to the transcription complex by phosphorylation of the RNA polymerase II carboxy-terminal domain. Genes Dev 11, 3319–26 (1997).
McCracken, S. et al. 5′-Capping enzymes are targeted to pre-mRNA by binding to the phosphorylated carboxy-terminal domain of RNA polymerase II. Genes Dev 11, 3306–18 (1997).
Komarnitsky, P., Cho, E. J. & Buratowski, S. Different phosphorylated forms of RNA polymerase II and associated mRNA processing factors during transcription. Genes Dev 14, 2452–60 (2000).
Akhtar, M. S. et al. TFIIH kinase places bivalent marks on the carboxy-terminal domain of RNA polymerase II. Mol Cell 34, 387–93 (2009).
Egloff, S. et al. Serine-7 of the RNA polymerase II CTD is specifically required for snRNA gene expression. Science 318, 1777–9 (2007).
Valay, J. G. et al. The KIN28 gene is required both for RNA polymerase II mediated transcription and phosphorylation of the Rpb1p CTD. J Mol Biol 249, 535–44 (1995).
Hong, S. W. et al. Phosphorylation of the RNA polymerase II C-terminal domain by TFIIH kinase is not essential for transcription of Saccharomyces cerevisiae genome. Proc Natl Acad Sci USA 106, 14276–80 (2009).
Keogh, M. C., Cho, E. J., Podolny, V. & Buratowski, S. Kin28 is found within TFIIH and a Kin28-Ccl1-Tfb3 trimer complex with differential sensitivities to T-loop phosphorylation. Mol Cell Biol 22, 1288–97 (2002).
Kanin, E. I. et al. Chemical inhibition of the TFIIH-associated kinase Cdk7/Kin28 does not impair global mRNA synthesis. Proc Natl Acad Sci USA 104, 5812–7 (2007).
Liu, Y. et al. Two cyclin-dependent kinases promote RNA polymerase II transcription and formation of the scaffold complex. Mol Cell Biol 24, 1721–35 (2004).
Bataille, A. R. et al. A universal RNA polymerase II CTD cycle is orchestrated by complex interplays between kinase, phosphatase and isomerase enzymes along genes. Mol Cell 45, 158–70 (2012).
Ansari, A. & Hampsey, M. A role for the CPF 3′-end processing machinery in RNAP II-dependent gene looping. Genes Dev 19, 2969–78 (2005).
El Kaderi, B., Medler, S., Raghunayakula, S. & Ansari, A. Gene looping is conferred by activator-dependent interaction of transcription initiation and termination machineries. J Biol Chem 284, 25015–25 (2009).
Hampsey, M., Singh, B. N., Ansari, A., Laine, J. P. & Krishnamurthy, S. Control of eukaryotic gene expression: gene loops and transcriptional memory. Adv Enzyme Regul 51, 118–25 (2011).
Laine, J. P., Singh, B. N., Krishnamurthy, S. & Hampsey, M. A physiological role for gene loops in yeast. Genes Dev 23, 2604–9 (2009).
Medler, S. et al. Evidence for a complex of transcription factor IIB with poly(A) polymerase and cleavage factor 1 subunits required for gene looping. J Biol Chem 286, 33709–18 (2011).
Moabbi, A. M., Agarwal, N., El Kaderi, B. & Ansari, A. Role for gene looping in intron-mediated enhancement of transcription. Proc Natl Acad Sci USA 109, 8505–10 (2012).
Mukundan, B. & Ansari, A. Srb5/Med18-mediated termination of transcription is dependent on gene looping. J Biol Chem 288, 11384–94 (2013).
O’Sullivan, J. M. et al. Gene loops juxtapose promoters and terminators in yeast. Nat Genet 36, 1014–8 (2004).
Singh, B. N., Ansari, A. & Hampsey, M. Detection of gene loops by 3C in yeast. Methods 48, 361–7 (2009).
Singh, B. N. & Hampsey, M. A transcription-independent role for TFIIB in gene looping. Mol Cell 27, 806–16 (2007).
Tan-Wong, S. M., Wijayatilake, H. D. & Proudfoot, N. J. Gene loops function to maintain transcriptional memory through interaction with the nuclear pore complex. Genes Dev 23, 2610–24 (2009).
Tan-Wong, S. M. et al. Gene loops enhance transcriptional directionality. Science 338, 671–5 (2012).
Castelnuovo, M. & Stutz, F. Gene loops and HDACs to promote transcription directionality. Nucleus 4, 92–4 (2013).
Crevillen, P., Sonmez, C., Wu, Z. & Dean, C. A gene loop containing the floral repressor FLC is disrupted in the early phase of vernalization. EMBO J 32, 140–8 (2013).
O’Reilly, D. & Greaves, D. R. Cell-type-specific expression of the human CD68 gene is associated with changes in Pol II phosphorylation and short-range intrachromosomal gene looping. Genomics 90, 407–15 (2007).
Perkins, K. J., Lusic, M., Mitar, I., Giacca, M. & Proudfoot, N. J. Transcription-dependent gene looping of the HIV-1 provirus is dictated by recognition of pre-mRNA processing signals. Mol Cell 29, 56–68 (2008).
Yun, K., So, J. S., Jash, A. & Im, S. H. Lymphoid enhancer binding factor 1 regulates transcription through gene looping. J Immunol 183, 5129–37 (2009).
Al Husini, N., Kudla, P. & Ansari, A. A role for CF1A 3′ end processing complex in promoter-associated transcription. PLoS Genet 9, e1003722 (2013).
Ho, C. K. & Shuman, S. Distinct roles for CTD Ser-2 and Ser-5 phosphorylation in the recruitment and allosteric activation of mammalian mRNA capping enzyme. Mol Cell 3, 405–11 (1999).
Mapendano, C. K., Lykke-Andersen, S., Kjems, J., Bertrand, E. & Jensen, T. H. Crosstalk between mRNA 3′ end processing and transcription initiation. Mol Cell 40, 410–22 (2010).
Skourti-Stathaki, K., Kamieniarz-Gdula, K. & Proudfoot, N. J. R-loops induce repressive chromatin marks over mammalian gene terminators. Nature 516, 436–9 (2014).
Rhee, H. S. & Pugh, B. F. Genome-wide structure and organization of eukaryotic pre-initiation complexes. Nature 483, 295–301 (2012).
Tietjen, J. R. et al. Chemical-genomic dissection of the CTD code. Nat Struct Mol Biol 17, 1154–61 (2010).
Drogat, J. & Hermand, D. Gene-specific requirement of RNA polymerase II CTD phosphorylation. Mol Microbiol 84, 995–1004 (2012).
Mukundan, B. & Ansari, A. Novel role for mediator complex subunit Srb5/Med18 in termination of transcription. J Biol Chem 286, 37053–7 (2011).
El Kaderi, B., Medler, S. & Ansari, A. Analysis of interactions between genomic loci through Chromosome Conformation Capture (3C). Curr Protoc Cell Biol Chapter 22, Unit22 15 (2012).
Venters, B. J. & Pugh, B. F. Genomic organization of human transcription initiation complexes. Nature 502, 53–8 (2013).
Murakami, K. et al. Architecture of an RNA polymerase II transcription pre-initiation complex. Science 342, 1238724 (2013).
Luse, D. S. The RNA polymerase II preinitiation complex: Through what pathway is the complex assembled? Transcription 5(2013).
Gelev, V. et al. A new paradigm for transcription factor TFIIB functionality. Sci Rep 4, 3664 (2014).
Helenius, K. et al. Requirement of TFIIH kinase subunit Mat1 for RNA Pol II C-terminal domain Ser5 phosphorylation, transcription and mRNA turnover. Nucleic Acids Res 39, 5025–35 (2011).
Sun, M. et al. Comparative dynamic transcriptome analysis (cDTA) reveals mutual feedback between mRNA synthesis and degradation. Genome Res 22, 1350–9 (2012).
Schulz, D., Pirkl, N., Lehmann, E. & Cramer, P. Rpb4 subunit functions mainly in mRNA synthesis by RNA polymerase II. J Biol Chem 289, 17446–52 (2014).
Bonnet, J. et al. The SAGA coactivator complex acts on the whole transcribed genome and is required for RNA polymerase II transcription. Genes Dev 28, 1999–2012 (2014).
Mayer, A. et al. Uniform transitions of the general RNA polymerase II transcription complex. Nat Struct Mol Biol 17, 1272–8 (2010).
Zhang, J. et al. ChIA-PET analysis of transcriptional chromatin interactions. Methods 58, 289–99 (2012).
Kim, H. et al. Gene-specific RNA polymerase II phosphorylation and the CTD code. Nat Struct Mol Biol 17, 1279–86 (2010).
Glover-Cutter, K. et al. TFIIH-associated Cdk7 kinase functions in phosphorylation of C-terminal domain Ser7 residues, promoter-proximal pausing and termination by RNA polymerase II. Mol Cell Biol 29, 5455–64 (2009).
Henriques, T. et al. Transcription termination between polo and snap, two closely spaced tandem genes of D. melanogaster. Transcription 3, 198–212 (2012).
Wang, Y., Fairley, J. A. & Roberts, S. G. Phosphorylation of TFIIB links transcription initiation and termination. Curr Biol 20, 548–53 (2010).
Pinto, I., Wu, W. H., Na, J. G. & Hampsey, M. Characterization of sua7 mutations defines a domain of TFIIB involved in transcription start site selection in yeast. J Biol Chem 269, 30569–73 (1994).
Core, L. J., Waterfall, J. J. & Lis, J. T. Nascent RNA sequencing reveals widespread pausing and divergent initiation at human promoters. Science 322, 1845–8 (2008 ).
Acknowledgements
We are grateful to Dr. Lori Pile (Wayne State University) for critical reading of the manuscript. We thank Dr. Chhabi Govind (Oakland University) for kindly providing the Kin28-as mutant generated by the laboratories of Drs. Kevin Shokat, Steve Hahn and Aseem Ansari. We thank Neha Agarwal and Zuzer Dhoondia for helpful suggestions.
Author information
Authors and Affiliations
Contributions
A.A. conceived the idea and supervised research, A.A. and S.M. designed experiments, S.M. performed all experiments, A.A. and S.M. analyzed data, A.A. wrote paper.
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Electronic supplementary material
Rights and permissions
This work is licensed under a Creative Commons Attribution 4.0 International License. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in the credit line; if the material is not included under the Creative Commons license, users will need to obtain permission from the license holder to reproduce the material. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Medler, S., Ansari, A. Gene looping facilitates TFIIH kinase-mediated termination of transcription. Sci Rep 5, 12586 (2015). https://doi.org/10.1038/srep12586
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/srep12586
- Springer Nature Limited
This article is cited by
-
Beyond the canonical role of TFIIB in eukaryotic transcription
Current Genetics (2022)
-
Dissecting the Pol II transcription cycle and derailing cancer with CDK inhibitors
Nature Chemical Biology (2020)
-
RNA polymerase II phosphorylation and gene looping: new roles for the Rpb4/7 heterodimer in regulating gene expression
Current Genetics (2020)
-
Genomewide Analysis of Clp1 Function in Transcription in Budding Yeast
Scientific Reports (2017)