
Abstract
Developing high-yielding rice varieties that possess favorable agronomic characteristics and enhanced grain Zn content is crucial in ensuring food security and addressing nutritional needs. This research employed ICIM, IM, and multi-parent population QTL mapping methods to identify important genetic regions associated with traits such as DF, PH, NT, NP, PL, YLD, TGW, GL, GW, Zn, and Fe. Two populations of recombinant inbred lines consisting of 373 lines were phenotyped for agronomic, yield and grain micronutrient traits for three seasons at IRRI, and genotyped by sequencing. Most of the traits demonstrated moderate to high broad-sense heritability. There was a positive relationship between Zn and Fe contents. The principal components and correlation results revealed a significant negative association between YLD and Zn/Fe. ICIM identified 81 QTLs, while IM detected 36 QTLs across populations. The multi-parent population analysis detected 27 QTLs with six of them consistently detected across seasons. We shortlisted eight candidate genes associated with yield QTLs, 19 genes with QTLs for agronomic traits, and 26 genes with Zn and Fe QTLs. Notable candidate genes included CL4 and d35 for YLD, dh1 for DF, OsIRX10, HDT702, sd1 for PH, OsD27 for NP, whereas WFP and OsIPI1 were associated with PL, OsRSR1 and OsMTP1 were associated to TGW. The OsNAS1, OsRZFP34, OsHMP5, OsMTP7, OsC3H33, and OsHMA1 were associated with Fe and Zn QTLs. We identified promising RILs with acceptable yield potential and high grain Zn content from each population. The major effect QTLs, genes and high Zn RILs identified in our study are useful for efficient Zn biofortification of rice.
Similar content being viewed by others

Introduction
The global need for food and nutritional security to end hunger and hidden hunger has been widely recognized1,2,3,4. Rice is the main source of calories in many South and Southeast Asian developing countries; however, milled rice is less nutritious5,6. Developing high-yielding and nutrient-dense rice varieties is crucial in achieving food and nutritional security7,8,9. Biofortified food crops can help alleviate malnutrition10,11,12,13. Recently, a concerted effort has been done to biofortify food crops with various vitamins and minerals14. Zinc (Zn), iron (Fe), and vitamin A deficiencies affect more than three billion people and can cause anemia, stunting, weakened immunity, blindness, diarrhea, and other health problems. Therefore, these nutrients have been prioritized for biofortification of food crops10,15,16.
The availability of genetic variation for grain Zn content in rice germplasm, particularly in landraces, pigmented rice, wild rice, and aus subgroup, has led to a significant focus on developing Zn-biofortified rice varieties7,17,18. The HarvestPlus and IRRI biofortification teams, along with national partners, are leading genetic biofortification efforts for rice. The first wave of high-Zn rice varieties has been successfully developed and introduced in South and Southeast Asia, and several promising high-Zn rice lines have been evaluated in different Asian and African countries2.
Developing Zn-biofortified rice varieties with higher yield and other desirable traits is challenging for rice breeders because of the genetic complexity of grain yield and grain Zn content, which are greatly influenced by the environmental factors19,20. Although there is often a negative correlation between grain Zn content and yield, a few specific germplasm accessions and populations have shown either no association or a negligible correlation between these two traits paving the way for successful Zn biofortification21,22,23.
Numerous Quantitative Trait Loci (QTL) mapping studies have been conducted in rice using different mapping populations8,17,18,20,24,25,26,27,28,29. Recent developments in high-throughput sequencing and robust statistical analysis have enabled the breeders to uncover more detailed information about the genetic basis of complex traits. Molecular markers have been successfully used by the breeders to develop high-yielding and nutritious rice varieties30,31,32,33. Breeding programs are transitioning from trait-based breeding approaches to modernized product development pipelines to accelerate varietal replacement23. It effectively integrates all the traits necessary for market acceptance into breeding products. As a result, the selection for high grain Zn content is being integrated into elite breeding pool to ensure the constant delivery of sufficient Zn to the diets of nutrition-vulnerable populations.
Multi-parent populations (MPPs) are important resources for exploring plant genetic architecture and identifying QTL because of their superior control over population structure, ability to capture greater genetic diversity, and decreased likelihood of false-positive QTL discovery34,35. The QTL effects in MPPs may exhibit varying degrees of allelic diversity, which affects QTL identification, but larger crossing/breeding combinations can boost QTL detection power despite having fewer parents36. Overall, MPPs have become increasingly popular in QTL mapping.
In this study, we analyzed phenotypic variation, correlations, and principal components of yield, agronomic traits, and grain Zn and Fe contents in two recombinant inbred line (RILs) populations of rice. We identified QTLs for grain yield, agronomic, and nutritional traits using multi-parent population analysis, interval mapping (IM) and inclusive composite interval mapping (ICIM). We also identified RILs with high grain Zn content and acceptable yield potential that can serve as parental lines in rice biofortification programs.
Results
Phenotypic performance
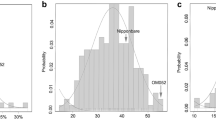
Most traits showed differences among parents (Table 1). Kaliboro had higher values for PH, NT, NP, TGW, GL, GW, and Zn than the other parents across the seasons. Similarly, wider variations and normal distributions were observed for all traits in both populations across the seasons (Fig. 1). DF, PH, TGW, GL, and Zn showed higher heritability, whereas Fe consistently showed lower heritability in both populations. The RILs exhibit transgressive segregation on both sides of the bell curve. The analysis of variance revealed significant genotype by seasonal effects for all traits in both populations, except for DF, PH, NT, Fe and Zn in P1 (Table 1).

Frequency distributions of agronomic, yield, Fe, and Zn in the P1 and P2 RIL populations of rice evaluated during the 2017 DS, 2017 WS, and 2018 DS. DF days to flowering (days), PH plant height (cm), NT number of tillers, NP number of panicles, PL panicle length (cm), YLD yield (kgha-1), TGW thousand grain weight (g), GL grain length (mm), GW grain width (mm), Fe iron (ppm), Zn zinc (ppm).
Correlation and principal component analysis
Out of 55 possible correlations in each population per season, the number of significant positive correlations was higher than the number of significant negative correlations (Fig. 2). The grain Zn content was consistently and positively correlated with PH, GW, and Fe, while it was negatively correlated with DF, GL, and YLD. Similarly, NT and NP, PL and PH, GW and TGW, GL and PL, and PL and YLD were positively correlated, whereas GL and GW were negatively correlated across the seasons and populations.

Correlations among agronomic, yield, Fe, and Zn in the P1 and P2 RIL populations of rice during the 2017 DS, 2017 WS, and 2018 DS. Positive correlations are indicated by dark purple color, while negative correlations are indicated by light purple color. Bigger circles correspond to stronger correlation coefficients. DF days to flowering (days), PH plant height (cm), NT number of tillers, NP number of panicles, PL panicle length (cm), YLD yield (kgha-1), TGW thousand grain weight (g), GL grain length (mm), GW grain width (mm), Fe iron (ppm), Zn zinc (ppm), *p < 0.05, **p < 0.01, ***p < 0.001.
The first two principal components (PC1 and PC2) explained the maximum total variance of 48.31% and 46.96% across seasons in P1 and P2, respectively (Table S1; Fig. 3). DF, NT, NP, GL, GW YLD, and Zn were the major contributors to the total PC1 variance (21–26%) in P1. Similarly, NT, NP, PL, GL, GW, YLD, Fe, and Zn contributed to the PC1 variance (25–29%) in P2. NT, NP, and YLD consistently showed high factor loading (0.4–0.6) across seasons in both populations.

Principal component analysis using the key variance components (PC1 and PC2) for yield, agronomic traits, Fe, and Zn of P1 and P2 RIL populations grown in 2017 DS, 2017 WS and 2018 DS. Purple dots inside the box correspond to the RILs of P1 and P2. A small angle between traits (colored in red) implies positive correlation, a large one suggests negative correlation, and a 90-degree angle indicates no correlation between traits. Traits with a high contribution to the population variance are in longer vector lines whereas traits with low contribution are in shorter lines. DF days to flowering (days), PH plant height (cm), NT number of tillers, NP number of panicles, PL panicle length (cm), YLD yield (kgha-1), TGW thousand grain weight (g), GL grain length (mm), GW grain width (mm), Fe iron (ppm), Zn zinc (ppm).
Identification of RILs with higher YLD and Zn
We identified the top five RILs in each population with higher YLD and Zn content (Table 2). In P1, among the top five RILs, Zn ranged from 12.9 to 27.3 ppm. IR120447-B-2-4-2 had the highest YLD of 8511 kgha−1 in the 2017 DS, with Zn of 21.8 ppm in 17DS, while IR120447-B-3-6-4 had the highest Zn of 27.3 and 26.8 ppm in the 2017 and 2018 DS, respectively, with a YLD of 5747.0 kgha−1 recorded in 17DS comparable with check variety IRRI 156. IR120447-B-2-5-2 had the DF of 80 days with a YLD of 5625.0 kgha−1 obtained in 18DS and Zn content of 20.3 ppm in 17DS. Meanwhile, IR120447-B-6-6-1 had the PH of 83.6 cm average across season and a Zn content of 21.6 ppm observed in 17DS. All of the top RILs in P1 possessed alleles that increased YLD, Zn, and Fe, with IR 120447-B-3–6-4 having the highest number of YLD (qYLD4.2, qYLD4.3, qYLD4.4, qYLD6.1, qYLD6.2, qYLD7.1) and Zn (qZn2.1, qZn5.1, qZn5.2, qZn10.1) QTLs but only one for Fe (qFe6.2), whereas the others carried two Fe QTLs. In P2, among selected lines, Zn ranged from 16.5–28.6 ppm and YLD from 3398.3–6541.4 kgha−1. IR120455-B-19-1-1 had the highest YLD average of 5361 kgha–1 across season with Zn from 18.4 ppm in 17WS to 22.6 ppm in 17DS, while IR120455-B-1-6-1 had the highest Zn content of 28.6 ppm in 17DS and YLD of 6388.1 kgha-1 obtained in 17DS. Moreover, IR 120455-B-17-5-1 had the DF of 81.4 average across season with high Zn of 22.3 ppm in 17DS. The YLD of all RILs in P1 like IR 120447-B-2-2-1, IR 120447-B-2-4-2, and IR 120447-B-2-5-2 exhibited higher YLD values compared to IRRI 156 the high yielding check in the 17DS and 18DS. However, in P2, the YLD of the 5 RILs was more consistent across seasons, with IRRI 156 performing relatively well. Three of the RILs in P2 carried combined alleles for YLD, Zn, and Fe with IR 120455-B-1-6-1 containing the greatest number of QTLs (qYLD4.4, qYLD6.2, qZn1.1, qZn2.1, qZn4.1, qZn4.2, qZn5.1, qZn6.3, qFe1.1). These RILs can be a good genetic resource for biofortification breeding programs.
Construction of linkage map
We processed the 11,609 SNPs data by removing redundant SNPs and those with a high missing rate (> 30%). In all 2018 and 2213 SNPs were obtained for P1 and P2, respectively (Table S2). The highest and lowest numbers of SNPs were on chromosomes 2 (274) and 9 (89) for P1 and on chromosomes 4 (328) and 10 (70) for P2, respectively. The total lengths of the linkage maps were 1232.2 cM for P1 and 1071.5 cM for P2. The average marker intervals were 0.61 cM and 0.48 cM in P1 and P2, respectively.
QTLs identified using ICIM and IM analysis
Seventy-five QTLs were identified for yield and agronomic traits using either IM or ICIM, and 22 of these QTLs were detected in both the analyses (Table 3; Fig. 4). The PVE of the QTL varied from 5% to 16.6% across the populations. There were nine QTLs for DF on chromosomes 2, 4, 6, 10, and 12, which explained a PV of 5.2–13.6%. The qDF2.2, contributed by IR95040, had the highest PVE (13.6%). Among the 15 QTLs detected for PH on chromosomes 1, 2, 4, 6, 7, 8, and 10, qPH1.2, was consistently identified on chromosome 1 (39.6–40.1 Mb), and the Kaliboro allele at this locus increased PH by 13.2–15.9 cm. Seven QTLs were mapped for NT on chromosomes 3, 4, 5, and 7. The qNT5.1 detected at 5.5–5.9 Mb on chromosome 5 explained the highest PV (16.6%) in P1. Likewise, eight QTLs were uncovered for NP on chromosomes 2, 3, 4, and 7, which explained a PV of 5–9.3%. The qNP7.3, explained 9.3% of the PV in P1. Eleven QTLs were identified for PL on chromosomes 2, 3, 4, 6, 7, and 8, explaining 5.8–13.5% PV. The qPL3.1, had the highest PVE and was contributed by IR95097. qPL4.2, was consistently mapped on chromosome 4 (20.6–20.8 Mb) during both the DS and by both analyses (ICIM and IM). Five QTLs were identified for YLD on chromosomes 4, 6, and 7, with a PVE ranging from 5% to 14.8%. It is interesting to note that a large effect QTL for yield, qYLD4.1 (PVE > 10%) was detected on chromosome 4 (12.6–13.3 Mb) in P2. Four QTLs were detected for TGW on chromosomes 1, 4, 6, and 12, explaining a PV of 5.1–10.7%. The qTGW1.1 had the highest effect (PVE, 10.7%). Likewise, five QTLs responsible for GL located on chromosomes 2, 4, and 10 explained a PVE of 7.9 to 10.6%. Among them, qGL2.2 and qGL10.1 explained PV > 10%. A total of 11 QTLs were found for GW on chromosomes 2, 3, 4, 6, 7, and 10, with PVE that varied from 5.2 to 13.8%. Two large effect QTLs (qGW4.2 and qGW4.5) with PVE > 10% were detected on chromosome 4.

Genomic visualization of QTLs identified for yield, agronomic traits, Fe, and Zn in RIL populations using ICIM and IM analyses. The outer circle represents the rice linkage map. The middle circle represents DF, PH, PL, NT, NP, and YLD QTLs. The inner circle represents the QTLs for GL, GW, TGW, Fe, and Zn.
Five QTLs (qFe4.1, qFe6.1, qFe6.2, qFe9.1 and qFe11.1) were detected for grain Fe content on chromosomes 4, 6, 9, and 11 in P1. The Kaliboro allele at qFe4.1 and qFe6.1 increased Fe content. Thirteen QTLs were associated with Zn content on chromosomes 3, 4, 5, 6, 7, 8, 10, and 12, accounted for 5.2–18.3% of PVE. Interestingly, four QTLs were consistently detected on chromosomes 3, 7, and 10 in P1 and on chromosomes 4 and 8 in P2. The qZn3.1 located at 11.5–11.6 Mb on chromosome 3 explained 7.5% and 6% PVE during 17DS and 17WS respectively. While qZn7.3 was located between 28.9–29.1 Mb on chromosome 7 had an additive effect of 2.6 ppm in 17DS and 2.8 ppm in 18DS. The qZn10.1, detected between 19.13–19.14 Mb on chromosome 10 had the highest PVE (> 10%) and additive effect (3.1 ppm), and the trait-enhancing allele was contributed by Kaliboro. Similarly, qZn4.1 had an additive effect of > 2.4 ppm across the three seasons.
QTLs identified using multi-parent population analysis
The MPP QTL analyses detected twenty-two QTLs for yield and agronomic traits, and five QTLs for grain Zn and Fe contents (Table 4; Fig. 5). The qPH1.2 was consistently detected on chromosome 1 with a PVE > 53.5% and a P-value of 37.7. There were five QTLs for DF on chromosomes 2, 5, 7, and 9 (PVE—4.4 to 8.9%; P value -3.7 to 7). NT had one QTL on chromosome 11, whereas NP had two QTLs on chromosomes 2 and 11. Three QTLs were identified for PL on chromosomes 1, 2, and 4, their P-values ranged from 3.7 to 8.4 and PVE from 5.7 to 13%. There were two QTLs for TGW on chromosomes 3 and 5; qTGW3.1, had the highest PVE (6.6%) and P value (6.0). Among the three QTLs detected for GL, qGL3.1 explained 16.5% of the PV. Similarly, three QTL were detected for GW on chromosomes 3 and 4, and their PVE varied from 4.6 to 9.2%. Two QTLs detected for YLD on chromosomes 4 and 6 had PVE of < 10%. For grain Zn content, four QTLs were identified on chromosomes 1, 2, and 5, and the large effect QTL qZn5.2 explained 10.8% PV. There was only one QTL for Fe content on chromosome 1 with a PVE of 5.1%.

Genomic visualization of QTLs identified for yield, agronomic traits, Fe, and Zn in RIL populations using multiparent population analysis. The outer circle represents the rice linkage map. The middle circle represents yield and agronomic, Fe, and Zn QTLs.
Candidate genes underlying consistent QTLs
We shortlisted 53 candidate genes underlying the QTLs detected for agronomic, yield, and nutritional traits (Table 5). Eight candidate genes were associated with two YLD QTLs: OsD11, OsFCP1, DAO, DWA1, Rf17, and OsCRC1 co-located with qYLD4.4, while dwarf35 and OsMIOX were co-located with qYLD6.2. Candidate gene dh1 was associated with qDF2.3, whereas seven genes that known to be associated with plant height (SDG714, OsSSI2, OsIRX10, HDT702, lhd2, OsCKX4, sd1) were identified with qPH1.2. Six candidate genes were co-located with four QTLs such as qNT11.1 (OsARF1), qNP11.1 (OsD27), qPL1.1 (OsFBP1, OsAAP6), and qPL8.2 (WFP, OsIPI1). Similarly, OsRSR1, OsMTP1, OsVPS22, OsLOX2 (L-2), and OsERO1 were co-located with the QTLs for TWG, GL, and GW. We analyzed the expression profiles of the 12 candidate genes using RiceXPro. The 12 agronomic and yield candidate genes included OsIRX10, HDT702, lhd2, and sd1 for PH; OsARF1 for NT; OsFBP1, OsAAP6, WFP, and OsIPI1 for PL; OsRSR1, and OsMTP1 for TGW; and OsD11 for yield (Fig. 7).

Gene networks showing the relationship between the (A) sd1 gene and other putative QTLs on chromosome 1, (B) OsNAS1 gene and other putative QTLs on chromosome 3, (C) OsSAP13 gene and other putative QTLs on chromosome 1; (D) OsRZFP34 gene and other putative QTLs on chromosome 1, (E) OsC3H33 gene and other putative QTLs on chromosome 5, (F) OsC3H37 gene and other putative QTLs on chromosome 5, (G) OsHMA1 gene and other putative QTLs on chromosome 6.
Twenty-six candidate genes were associated with QTLs detected for Zn and Fe contents. Three candidate genes (OsSAP13, OsRZFP34, and OsHMP5) were co-located with qZn1.1. The OsNAS1 and OsSET16 co-localized with qZn3.1, whereas qZn4.1 harbored OsDLN109 and OsMTP7. On chromosome 5, OsC3H33 was co-located with qZn5.1, OsGATA1, and OsC3H37, OsSET22, OsSIRP1, and OsAIR1 were co-located with qZn5.2. Likewise, Osferroportin, OsHMP32, OsHMA1, OsHMA7, OsRFPHC-6, and OsC3H53 were linked to QTLs detected for Zn on chromosomes 6 and 7. The OsHMP7 and OsRFP were co-located with qFe1.1. To comprehend the expression patterns of these genes during various developmental stages, the RiceXPro database was used to examine the expression profiles of several genes. The OsRZFP34, OsHMA1, OsMTP7, OsRFPHC-6 and OsC3H37 were over expressed in the embryo (Fig. 8), while OsNAS1, OsC3H37 and OsRFP over expressed in the endosperm. Furthermore, the ovary exhibited over-expression of several genes that included OsHMP5, OsFerroportin, OsHMA1, OsNas1, OsMTP7, OsRFPC-6, and OsC3H53.

Graphical representation of the expression profiles of 12 candidate genes for agronomic and yield traits throughout rice development, based on spatiotemporal profiling across various tissues and organs, with data extracted from RiceXPro (http://ricexpro.dna.affrc.go.jp/).
Discussion
The biofortification of rice with improved grain Zn content is a priority nutritional improvement program that helps tackling Zn malnutrition37,38. The investigation of molecular mechanisms involved in Zn metal homeostasis in rice contributes to the enhancement of Zn biofortification of rice23,27,31. Recent advancements in genotyping techniques, as well as statistical and bioinformatics tools have facilitated accurate and precise QTL mapping in diverse set of populations34,39. In this study, we employed ICIM mapping and MPP analysis to conduct QTL analysis on two biparental populations with shared parentage, and identified robust QTLs associated with Zn content that can be effectively utilized for genomic-assisted breeding34,40.
We observed significant variations in the majority of traits across the three seasons in both RILs populations, with most of the traits showing a normal distribution. The use of high Zn elite breeding lines as recipient parents generates effective variations that can be directly used in pre-breeding, as opposed to utilizing highly contrasting parents, which may result in reduced agronomic performance and lead to the inclusion of undesirable alleles. These variations also imply the quantitative or polygenic inheritance of the traits8,17,31. Our study revealed moderate to high heritability values (H2) for traits such as DF, PH, PL, YLD, TGW, GL, and Zn, which is consistent with results of previous studies conducted using different populations17,18,20,31. These traits can easily be transferred to the next generation through conventional method and permits effective phenotypic selection for their improvement16,41,42. Meanwhile, traits with lower to intermediate heritability can integrate conventional and molecular techniques in improving these traits.
In both RILs populations across seasons, DF, PL, and GL showed a positive correlation with YLD which is expected since these traits are considered as yield components that directly influence the trait. In contrast, Zn exhibited a negative correlation with YLD consistently across populations and seasons that conforms with previous studies8,17,20,31,43. The unfavorable correlation between YLD and Zn presents a challenge for breeders in developing high-yielding Zn-biofortified rice varieties, as it may indicate a different mechanism at the genetic level that controls both traits, having opposite effects on their improvement. To overcome this challenge, appropriate breeding techniques and selection procedures must be applied to identify stable high-Zn donor lines with a greater or acceptable YLD7,23,38. It is essential to properly combine YLD and Zn to develop high-yielding Zn-biofortified rice varieties that address food and nutritional security in developing countries. On the other hand, the present study confirms previous findings that grain Zn exhibited both positive and negative relationships with TGW and GL, respectively, whereas Fe exhibited a negative relationship with both TGW and GL24,31, these can be attributed to the dilution effect where Fe and Zn concentration tend to be diluted as grain related traits increase. However, we also found substantial positive correlations between Zn and Fe in both populations across seasons, consistent with previous findings44,45,46,47,48. These results suggest that common molecular mechanisms, pathways, and gene networks likely underlie the uptake, translocation, and loading of these two elements in rice grains7. However, there were some differences in the deposition of Fe and Zn within the rice grains. While most of the Fe is deposited in the aleurone layer, a significant portion of Zn is loaded into the endosperm49. This variation may be useful in breeding high-Zn rice through biofortification breeding, whereas high-Fe rice is being developed using genetic engineering or genome editing approaches2,23.
Principal component analysis (PCA) is a powerful tool for summarizing multivariate data and achieving the best representation in a lower-dimensional space50. Generally, PCs with vector co-efficients greater than 0.3 are considered important regardless of their direction of influence51. Component loadings for each principal component revealed that YLD, PL, and GL were among the phenotypic traits contributing positive projections, whereas Fe and Zn contributed negatively to PCs in P1. The results revealed that these traits significantly contribute to the overall variability in the population52. The PL and GL can be considered in the selection criteria in selecting high-yielding lines. Moreover, the positive significant contributions of YLD further indicates the higher likelihood of identifying high-yielding lines in the populations. In P2, YLD showed positive projections while Fe and Zn showed negative projections in 2017DS, whereas YLD showed negative projections while Fe and Zn showed positive projections in the two PCs during 2017WS and 2018DS. These opposite projections of YLD and Zn on PCA corroborated the results of the correlation analysis. The results of PCA can be utilized to identify the traits with the greatest impact on the phenotype of RILs for line selection. Interestingly, we observed that some correlated variables, such as NT and NP, Fe and Zn, and GW, had similar PC effects (positive or negative) for each season and population.
In our study, we found more QTLs in P1 compared to its counterpart during WS. The donor parent Kaliboro contributed 71% of the positive alleles. The biparental and MPP analyses identified 93 and 27 QTLs, respectively (Tables 3, 4). The MPP analysis identified only 27 QTLs for 11 traits, three of these QTLs (qPH1.2, qPL4.2, and qZn5.1) were consistently detected across populations and seasons. It also detected two YLD QTLs (qYLD4.4 and qYLD6.2) on chromosomes 4 and 6. Where as, IM identified two QTLs on chromosomes 4 and 7 while ICIM identified five QTLs on chromosomes 4, 6, and 7 that were associated with YLD. Previous studies reported yield QTLs at the same chromosomal regions on chromosomes 4 and 7. Khahani et al. (2021) identified MQTL-YLD4 at 29.15–30.85 Mb on chromosome 4 that harbored multiple genes associated with grain yield67. In another study, Zhang et al. (2021) using a high-density genetic map of hybrid rice, revealed the presence of a consistent QTL for YLD (qGYP-6b) on chromosome 6 across different environments68. These findings highlight the need for further exploration of these specific genomic regions on chromosomes 4 and 6 to identify key regulatory genes influencing YLD in rice.
Our analysis revealed QTLs for Fe on chromosomes 1, 4, 6, 9, and 11 using various methods, which is consistent with previous studies that identified common regions on chromosomes 1, 3, and 7 in rice16,17,18,21,31,69,70,71. Specifically, we identified QTLs for Fe content on chromosome 1 using MPP, on chromosomes 4 and 6 using ICIM, and on chromosomes 9 and 11 using IM. Moreover, a recent review by Swamy et al. (2021) reported several QTL for Fe on chromosomes 1, 3, and 7, and also shortlisted candidate genes for Fe homeostasis38. These results suggest that multiple genomic regions and genes are associated with Fe accumulation in rice, which may provide valuable targets for future breeding efforts to develop Fe-biofortified rice varieties.
In MPP analysis, a consistent QTL for Zn16 was found on chromosome 5 similar to the report of Palanog et al. (2023). Moreover, numerous studies have also reported similar genomic regions making it potential loci for genomics assisted breeding for Zn biofortification24,55,59,64. Zn QTLs were also detected on chromosomes 3, 4, 5, 6, 7, 8, and 10 using ICIM and IM. Previous QTL mapping studies have identified Zn QTLs on all rice chromosomes8,17,18,19,20,27,31,43,44,55,60,61,65,72,73. However, it is important to note that these QTL mapping experiments used different parental genotypes, mapping populations, field sites, and environmental conditions. Hence, further validation is required prior to integrating the Zn QTLs into the biofortification program.
Our candidate gene analysis that were co-localized with QTLs for different agronomic traits such as DH1 for DF74, sd1, OsIRX10, and HDT702 for PH75,76,77, OsD27 for NP78, Wealthy Farmer's Panicle (WFP) and OsIPI1 for PL79,80, OsRSR1 and OsMTP1 for TGW81, and OsVPS22 for GL with proven biological and physiological functions and directly involved with the traits. With the help of KnetMiner, we were able to identify the relationship of the candidate genes with other genes proximately connected with them and regions where they are largely expressed. It appears that there are associations between the sd1 gene and 39 QTLs in the KnetMiner, as well as another gene GA20OX5, which is responsible for shortening plant height (Fig. 6A). We also identified CL4 and d35 gene co-located with YLD QTLs known for their importance in panicle development and regulation of plant height that have contributed to crop productivity82,83. These genes are potential candidates for functional validation and for introgression through MAB to develop high yielding nutritious rice varieties84,85. For instance, OsRSR1 and OsMTP1 were co-located with TGW QTLs that play an essential role in producing higher grain YLD, these genes were also involved in metal transportation within the rice plant86,87.

Graphical representation of the expression profiles of 12 candidate genes for Zn and Fe throughout rice development, based on spatiotemporal profiling across various tissues and organs, with data extracted from RiceXPro (http://ricexpro.dna.affrc.go.jp/).
One of the highlights of our study is the identification of putative genes co-located with major Zn QTLs such as OsNAS1, OsSAP13, OsMTP7 OsC3H33, OsC3H37, OsSDG728/SET22, OsSIRP1, OsAIR1, OsHMA1, OsFerroportin, and OsRFP (Fig. 8). These genes are deeply involved in Zn binding and heavy metals homeostasis and have been connected to delivering Zn to developing rice seeds88 (Fig. 6B–G). They are also expressed and upregulated in various parts of rice such as roots, embryo, ovary, stem, anther, pistil, lemma, and palea at various growth stages of rice development (Fig. 8, RiceXPro). The identification of these potential genes offers valuable insights into the molecular processes involved in Zn uptake and transport within rice plants. This emphasizes the significance of understanding the genetic basis of Zn absorption and movement in rice plants, which carries implications for improving both the nutritional quality and productivity of this important staple crop.
Our study has identified promising lines with superior YLD and Zn in both populations. We selected IR 120447-B-2-4-2 with an average YLD of 5222.5 kgha-1 and Zn content of 18.9 ppm in P1 and IR 120455-B-19-1-1 with an average YLD of 5361.4 kgha-1 and Zn content of 20.3 ppm in P2. These promising lines highlight the increased chances of achieving the primary goal of biofortification breeding–to develop high-yielding Zn-biofortified varieties. These lines will be further evaluated and can be nominated for varietal release trials, these lines can also serve as parents in rice biofortification.
Conclusion
Phenotypic evaluation of RILs populations across seasons revealed wider variations for agronomic, yield and nutritional traits. QTL analyses (MPP, ICIM and IM) of multiple populations evaluated for three seasons identified consistent QTLs for agronomic, yield and biofortification traits. They were co-located with candidate genes important in the expression and regulation of these traits. We identified the top five RILs in each population with higher YLD (≥ 5 tons ha-1) and Zn content (12.9–28.6 ppm). Our findings provide breeders with valuable information on the genetic basis of agronomic, yield and Zn traits as well as SNP markers linked to candidate genes of important QTLs. Moreover, the identification of high-YLD and high-Zn rice lines will aid the rapid development of Zn biofortified rice varieties.
Materials and methods
Plant materials and phenotyping
These experiments were conducted at the Zeigler Experiment Station, International Rice Research Institute (IRRI), Los Baños, Laguna, the Philippines. The two RILs populations were derived from crosses of IR95040 x Kaliboro (IRGC 77201-1) (P1) and IR95097 × Kaliboro (IRGC 77201-1) (P2). The parental lines IR95040 and IR95097 are high-Zn breeding lines developed by the Biofortification group of IRRI, while Kaliboro is an Aus-type rice accession from IRRI Genebank with high grain Zn and Fe contents. The two populations, P1 and P2, were composed of 178 and 195 RILs, respectively. The seeds of the two populations and parental lines used in this study were obtained from the Biofortification group of IRRI. Lines were grown in an alpha-lattice design with two replications each during the 2017 dry season (DS) and wet season (WS) and the 2018 DS. Seeds were sown in a seedbed, and 21-days-old seedlings were transplanted. Standard agronomic practices and plant protection measures were applied to ensure good crop growth and complete grain development.
Data on days to 50% flowering (DF) were recorded when 50% of the plants in the plot started flowering. At maturity, three randomly selected plants from the middle of the plot were sampled to measure plant height (PH, cm) and panicle length (PL, cm). Grains were harvested from each plot, dried, weighed, and corrected to 14% moisture content for grain yield (YLD, kg ha-1). Post-harvest data, thousand-grain weight (TGW, g), grain length (GL, mm), and grain width (GW, mm), were also measured following the standard evaluation system of rice95.
Grain Zn and Fe contents were measured using an XRF-Bruker S2 Ranger. Rice samples were obtained from the bulked seeds of each plot. Samples were dehulled, and approximately 5 g of seeds were prepared for grain Zn and Fe content measurements. The samples were analyzed twice, with measurements expressed in parts per million (ppm). We identified ten promising RILs based on agronomic traits, grain yield, and Zn content for Zn biofortification breeding.
Data analysis
The means of the traits were used for statistical analysis. All basic statistical parameters were generated using STAR v. 2.0.1, whereas ANOVA and broad-sense heritability (H2) were analyzed using PBTools v. 1.4 (http://bbi.irri.org/). Histograms, principal component analysis, and Pearson’s correlation were estimated using R-Studio v. 1.2.5 (https://www.rstudio.com/).
DNA isolation and sequencing
Leaf samples were collected from 21-days-old seedlings from each of the 373 RILs and their parents (IR95040, IR95097, and Kaliboro IRGC 77201-1). DNA was isolated using a modified CTAB method115. DNA quality was assessed by electrophoresis on 1% agarose gel. Using this platform, high-quality DNA was obtained for SNP genotyping (Tea Break Bioinformatics, Limited). The ApeKI restriction enzyme was used for library preparation following a protocol modified by Elshire et al. (2011)116. For GBS, a type II restriction endonuclease (ApeKI) was used for DNA digestion, the digested DNAs was ligated to the adapter, and a 96-plex library was constructed according to the GBS protocol. High-density genotyping of all individuals was performed using genotyping-by-sequencing (GBS) with Tea Break Bioinformatics (http://tenformatics.com/index.html).
Sequence data analysis and SNP identification
The genotype sequence SNP data was analyzed using the Tassel 5 GBS pipeline117. SNP calling and filtering were performed using the Tassel pipeline117. The variant call format (VCF) was filtered using a minor allele frequency (MAF) > 0.01, call rate ≥ 95%, and the Hardy–Weinberg equilibrium (HWE) test with a threshold value of P < 0.001. Finally, 221,419 high-quality SNPs were selected for downstream analyses. The SNPs retained after filtering using the Tassel pipeline were manually filtered. SNPs with monomorphic alleles between the two parental lines, high heterozygosity, and more than 10% of missing SNP calls were excluded from further analysis. A total of 11,609 SNP markers were retained for linkage map construction in both populations.
Linkage mapping and QTL analysis
Linkage map construction and QTL analysis were performed using QTL ICIMapping v. 4.1118. The grouping of SNPs was based on their physical map locations on chromosomes. The ordering of the markers within each chromosome was based on recombination events between SNPs. The recombination distance was calculated using the Kosambi function119.
We performed QTL analyses for agronomic, yield and grain micronutrient (Zn and Fe) traits. The critical threshold value for QTL detection was calculated based on Churchill and Doerge (1994)120. SNPs associated with QTL identified by linkage mapping were anchored to the physical map. Information regarding the physical location of SNPs flanking the QTL was used to determine the position of the QTL. All QTL were visualized using the RCircos R package111. To increase the power of QTL detection and compare the effects of QTLs detected in cross-specific models, multi-parent population QTL mapping was performed using the mppR package40.
Candidate gene analysis
Candidate genes associated with agronomic traits, grain yield, grain Zn and Fe contents were analyzed across all detected QTLs. Identification of candidate genes within the QTL regions was performed using available online database: the “Rice Genome Annotation Project” website (http://rice.plantbiology.msu.edu/cgi-bin/ORFinfopage.), the “Gramene QTL Database” (http://archive.gramene.org/qtl/), and Oryzabase (https://shigen.nig.ac.jp/rice/oryzabase/quicksearch/list). Subsequently, locus IDs for these candidate genes were leveraged for trait association and gene ontology annotation. This facilitated the identification of GO terms relevant to the traits of interest. Leveraging literature-based evidence and GO, we identified top candidate genes. Additionally, we utilized Knetminer (http://knetminer.rothamsted.ac.uk/Oryza_sativa/) to construct a network mapping the relationships between candidate genes and related QTLs. The tissue-specific and developmental stage-specific expression patterns of these candidate genes were further elucidated using data from The Rice Expression Profile Database (http://ricexpro.dna.affrc.go.jp), which provided insights into gene expression throughout the rice plant’s life cycle.
This research did not involve any human participants or animals. The study adhered to applicable institutional, national, and international guidelines and legislation, ensuring that all experiments on rice (Oryza sativa L.) plants were conducted in accordance with relevant regulations. Additionally, this research was not conducted with endangered species and was carried out in accordance with the Declaration of the IUCN Policy on Research Involving Endangered Species.
Data availability
All data generated and analyzed supporting the findings of this study are available within the paper and within its supporting information published online.
Abbreviations
- DF:
-
Days to flowering
- Fe:
-
Iron/grain iron (ppm)
- GBS:
-
Genotyping-by-sequencing
- GL:
-
Grain length (mm)
- GW:
-
Grain width (mm)
- H2 :
-
Broad-sense heritability
- HWE:
-
Hardy weinberg equilibrium
- ICIM:
-
Inclusive composite interval mapping
- IM:
-
Interval mapping
- MAF:
-
Minor allele frequency
- NP:
-
Number of panicles
- NT:
-
Number of tillers
- PCA:
-
Principal component analysis
- PH:
-
Plant height (cm)
- PL:
-
Panicle length (cm)
- QTL:
-
Quantitative trait loci
- SNPs:
-
Single nucleotide polymorphisms
- TGW:
-
Thousand grain weight (g)
- VCF:
-
Variant call format
- YLD:
-
Grain yield (kgha−1)
- Zn:
-
Zinc/grain zinc (ppm)
References
Garcia-Oliveira, A. L., Chander, S., Ortiz, R., Menkir, A. & Gedil, M. Genetic basis and breeding perspectives of grain iron and zinc enrichment in cereals. Front. Plant Sci. 9, 937 (2018).
Tsakirpaloglou, N., Swamy, B. P. M., Acuin, C. & Slamet-Loedin, I. H. Biofortified Zn and Fe rice: potential contribution for dietary mineral and human health. In Nutritional Quality Improvement in Plants (eds Jaiwal, P. K. et al.) (Cham, 2019).
McCann, S., Perapoch, A. M. & Moore, S. The role of iron in brain development: A systematic review. Nutrients https://doi.org/10.3390/nu12072001(2020) (2001).
Basheer, S., Ashique, V. V. & Grover, A. The food and nutrition status in India: A systematic review. In Sustainable Health Through Food, Nutrition, and Lifestyle (eds Grover, A., Singh, A. & Singh, R. B.), 143–159 (2023).
Reinke, R. et al. Rice biofortification—Progress and challenges in improving the nutritional value of rice. Proceedings 36(1), 69. https://doi.org/10.3390/proceedings2019036069 (2020).
Tan, B. L. & Norhaizan, M. E. Rice by-Products: Phytochemicals and Food Products Application (Springer International Publishing, 2020).
Swamy, B. P. M. et al. Advances in breeding for high grain zinc in rice. Rice 9, 1–16 (2016).
Calayugan, M. I. C. et al. Genetic analysis of agronomic traits and grain iron and zinc contents in a doubled haploid population of rice (Oryza sativa L.). Sci. Rep. https://doi.org/10.1038/s41598-020-59184-z (2020).
Rao, S. D. et al. Zinc biofortified rice varieties: Challenges, possibilities, and progress in India. Front. Nutr. 7, 26 (2020).
Palanog, A. D. et al. Zinc and iron nutrition status in the Philippines population and local soils. Front. Nutr. 6, 81 (2019).
Bashaasha, B. et al. Association between bio-fortification and child nutrition among smallholder households in Uganda. J. Agric. Econ. Rural Dev. 6(2), 752–759 (2020).
Athar, T. et al. Biofortification and the involved modern approaches. J. Elem. 25(2), 717–731. https://doi.org/10.5601/jelem.2020.25.1.1911 (2020).
Yadav, S., Sandhu, N., Singh, V., Catolos, M. & Kumar, A. Genotyping-by-sequencing based QTL mapping for rice grain yield under reproductive stage drought stress tolerance. Sci. Rep. https://doi.org/10.1038/s41598-019-50880-z (2019).
Dwivedi, S. L., Garcia-Oliveira, A. L., Govindaraj, M. & Ortiz, R. Biofortification to avoid malnutrition in humans in a changing climate: Enhancing micronutrient bioavailability in seed, tuber, and storage roots. Front. Plant Sci. 14, 1119148. https://doi.org/10.3389/fpls.2023.1119148 (2023).
Garg, M. et al. Biofortified crops generated by breeding, agronomy, and transgenic approaches are improving lives of millions of people around the world. Front. Nutr. 5, 12. https://doi.org/10.3389/fnut.2018.00012 (2018).
Palanog, A. D. et al. Molecular dissection of connected rice populations revealed important genomic regions for agronomic and biofortification traits. Front. Plant Sci. 14, 1157507 (2023).
Swamy, B. P. M. et al. QTL analysis for grain iron and zinc contents in two O. Nivara-derived backcross populations. Rice Sci. 25(4), 197–207. https://doi.org/10.1016/j.rsci.2018.06.003 (2018).
Swamy, B. P. M. et al. Identification of genomic regions associated with agronomic and biofortification traits in DH populations of rice. PLoS ONE 13(8), 1–20. https://doi.org/10.1371/journal.pone.0201756 (2018).
Zaw, H. et al. Exploring genetic architecture of grain yield and quality traits in a 16-way indica by japonica rice MAGIC global population. Sci. Rep. 9(1), 1–11. https://doi.org/10.1038/s41598-019-55357-7 (2019).
Descalsota-Empleo, G. I. et al. Genetic mapping of QTL for agronomic traits and grain mineral elements in rice. Crop J. 7(4), 560–572. https://doi.org/10.1016/j.cj.2019.03.002 (2019).
Gregorio, G. B., Senadhira, D., Htut, T. & Graham, R. D. Breeding for trace mineral density in rice. Food Nutr. Bull. 21, 382–386 (2000).
Norton, G. J. et al. Genetic mapping of the rice ionome in leaves and grain: Identification of QTL for 17 elements including arsenic, cadmium, iron and selenium. Plant Soil 329, 139–153 (2010).
Calayugan, M. I. C. et al. Zinc-biofortified rice: a sustainable food-based product for fighting zinc malnutrition. In Rice Improvement: Physiological, Molecular Breeding and Genetic Perspectives (eds Ali, J. & Wani, S. H.) (Springer International Publishing, 2021).
Descalsota-Empleo, G. I. et al. Genetic dissection of grain nutritional traits and leaf blight resistance in rice. Genes 10(1), 30. https://doi.org/10.3390/genes10010030 (2019).
Pradhan, S. et al. Linkage disequilibrium mapping for grain Fe and Zn enhancing QTLs useful for nutrient dense rice breeding. BMC Plant Biol. https://doi.org/10.1186/s12870-020-2262-4 (2020).
Lee, S. M. et al. QTL analysis for Fe and Zn contents in rice grains using a doubled haploid population derived from a cross between rice (Oryza sativa) cultivar 93–11 and milyang 352. Plant Breed. Biotech. 8(1), 69–76 (2020).
Joshi, G. et al. Meta-QTL s and haplotypes for efficient zinc biofortification of rice. Plant Genome 16(4), 20315 (2023).
Wallace, J. & Mitchell, S. Genotyping-by-sequencing. Curr. Protoc. Plant Biol. 2(1), 64–77 (2017).
de Ronne, M., Légaré, G., Belzile, F., Boyle, B. & Torkamaneh, D. 3D-GBS: A universal genotyping-by-sequencing approach for genomic selection and other high-throughput low-cost applications in species with small to medium-sized genomes. Plant Methods 19(1), 1–10 (2023).
Collard, B. C. & Mackill, D. J. Marker-assisted selection: An approach for precision plant breeding in the twenty-first century. Phil. Trans. R. Soc. B Biol. Sci. 363(1491), 557–572 (2008).
Descalsota, G. I. L. et al. Genome-wide association mapping in a rice MAGIC plus population detects QTLs and genes useful for biofortification. Front. Plant Sci. https://doi.org/10.3389/fpls.2018.01347 (2018).
Cruet-Burgos, C. et al. Advancing provitamin A biofortification in sorghum: Genome-wide association studies of grain carotenoids in global germplasm. Plant Genome 13, e20013. https://doi.org/10.1002/tpg2.20013 (2020).
Wu, X. et al. Genome-wide SNP identification and association mapping for seed mineral content in mung bean (Vigna radiata L.). Front. Genet. 11, 656. https://doi.org/10.3389/fgene.2020.00656 (2020).
Scott, M. F. et al. Multi-parent populations in crops: A toolbox integrating genomics and genetic mapping with breeding. Heredity 125(6), 396–416 (2020).
Garin, V., Malosetti, M. & van Eeuwijk, F. Multi-parent multi-environment QTL analysis: An illustration with the EU-NAM flint population. Theor. Appl. Genet. 133, 2627–2638 (2020).
Garin, V., Wimmer, V., Borchardt, D., Malosetti, M. & van Eeuwijk, F. The influence of QTL allelic diversity on QTL detection in multi-parent populations: A simulation study in sugar beet. BMC Genom. Data 22(1), 1–12 (2021).
Mwanga, R. O. et al. Breeding progress for vitamin A, Iron and Zinc biofortification, drought tolerance, and sweetpotato virus disease resistance in sweetpotato. Front. Sustain. Food Syst. 5, 616674 (2021).
Swamy, B. M., Marathi, B., Ribeiro-Barros, A. I., Calayugan, M. I. C. & Ricachenevsky, F. K. Iron biofortification in rice: An update on quantitative trait loci and candidate genes. Front. Plant Sci. 12, 647341 (2021).
Paccapelo, M. V., Kelly, A. M., Christopher, J. T. & Verbyla, A. P. WGNAM: Whole-genome nested association mapping. Theor. AppL. Genet. 135(7), 2213–2232 (2022).
Garin, V., Wimmer, V., Mezmouk, S., Malosetti, M. & van Eeuwijk, F. How do the type of QTL effect and the form of the residual term influence QTL detection in multi-parent populations? A case study in the maize EU-NAM population. Theor. Appl. Genet. 130, 1753–1764 (2017).
Holland, J. B. Implementation of molecular markers for quantitative traits in breeding programs—challenges and opportunities. New directions for a diverse planet: proceedings for the 4th international crop science congress. Regional Institute, Gosford, Australia (2004).
Cobb, J. N., DeClerck, G., Greenberg, A., Clark, R. & McCouch, S. Next-generation phenotyping: Requirements and strategies for enhancing our understanding of genotype–phenotype relationships and its relevance to crop improvement. Theor. Appl. Genet. 126, 867–887 (2013).
Dixit, S. et al. Identification of genomic region(s) responsible for high iron and zinc content in rice. Sci. Rep. 9, 8136. https://doi.org/10.1038/s41598-019-43888-y (2019).
Stangoulis, J. C., Huynh, B. L., Welch, R. M., Choi, E. Y. & Graham, R. D. Quantitative trait loci for phytate in rice grain and their relationship with grain micronutrient content. Euphytica 154, 289–294 (2007).
Paltridge, N. G., Palmer, L. J., Milham, P. J., Guild, G. E. & Stangoulis, J. C. Energy-dispersive X-ray fluorescence analysis of zinc and iron concentration in rice and pearl millet grain. Plant Soil 361, 251–260 (2012).
Gande, N. K. et al. Identification of putative candidate gene markers for grain zinc content using recombinant inbred lines (RIL) population of IRRI38 X Jeerigesanna. Afr. J. Biotechnol. 13(5), 657–663 (2014).
Norton, G. J. et al. Genome wide association mapping of grain arsenic, copper, molybdenum and zinc in rice (Oryza sativa L.) grown at four international field sites. PLoS ONE 9(2), e89685. https://doi.org/10.1371/journal.pone.0089685 (2014).
Rao, D. S. et al. Assessment of grain zinc and iron variability in rice germplasm using energy dispersive X-ray fluorescence spectrophotometer. J. Rice Res. 7(1), 45–52 (2014).
Diaz-Benito, S. et al. Iron and zinc in the embryo and endosperm of rice (Oryza sativa L.) seeds in contrasting 2’-deoxymugeneic acid/nicotiamine scenarios. Front. Plant Sci. 9, 1190. https://doi.org/10.3389/fpls.2018.01190 (2018).
Josse, J. & Husson, F. Selecting the number of components in principal component analysis using cross-validation approximations. Comput. Stat. Data Anal. 56(6), 1869–1879 (2012).
Raji, A. A. Assessment of genetic diversity and heterotic relationships in African improved and local cassava (Manihot esculenta Crantz) germplasm. PhD thesis. University of Ibadan, Nigeria (2002).
Thakur, K. & Sarma, M. K. Genetic diversity and principal component analysis in cultivated rice (Oryza sativa) varieties in Assam. Indian J. Agric. Sci. https://doi.org/10.56093/ijas.v931i2.132052 (2023).
Wang, X. et al. Identification and application of major quantitative trait loci for panicle length in rice (Oryza sativa) through single-segment substitution lines. Plant Breed. 138(3), 299–308 (2019).
Jia, B. et al. Quantitative trait loci mapping of panicle traits in rice. Mol. Biol. Res. Commun. 8(1), 9 (2019).
Hu, B. L. et al. Mapping QTLs for mineral element contents in brown and milled rice using an Oryza sativa× O. rufipogon backcross inbred line population. Cereal Res. Commun. 44, 57–68 (2016).
Mohiuddin, S. J., Haque, M. A., Haque, M. M., Islam, M. T. & Biswas, P. S. Genetic analysis reveals a major effect QTL associated with high grain zinc content in rice (Oryza sativa L.). Plant Breed, Biotechnol. 8(4), 327–340 (2020).
Nawaz, Z. et al. Genome-wide association mapping of quantitative trait loci (QTLs) for contents of eight elements in brown rice (Oryza sativa L.). J. Agric. Food Chem. 63(36), 8008–8016 (2015).
Anuradha, K. et al. Evaluating rice germplasm for iron and zinc content in brown rice and seed dimensions. J. Geophys. Res. 4, 19–25 (2012).
Islam, M. Z. et al. Mapping QTLs underpin nutrition components in aromatic rice germplasm. PLoS ONE 15(6), e0234395 (2020).
Yu, Y. H. et al. Mapping of quantitative trait loci for contents of macro-and microelements in milled rice (Oryza sativa L.). J. Agric. Food Chem. 63(35), 7813–7818 (2015).
Bollinedi, H. et al. Genome-wide association study reveals novel marker-trait associations (MTAs) governing the localization of Fe and Zn in the rice grain. Front. Genet. 11, 497543 (2020).
Zhang, M. et al. Mapping and validation of quantitative trait loci associated with concentrations of 16 elements in unmilled rice grain. Theor. Appl. Genet. 127, 137–165 (2014).
Sahu, P. K. et al. Genome-wide association mapping revealed numerous novel genomic loci for grain nutritional and yield-related traits in rice (Oryza sativa L.) landraces. 3 Biotech. 10, 1–22 (2020).
Garcia-Oliveira, A. L., Tan, L., Fu, Y. & Sun, C. Genetic identification of quantitative trait loci for contents of mineral nutrients in rice grain. J. Integr. Plant Biol. 51, 84–92 (2009).
Xu, Q. et al. Examining two sets of introgression lines in rice (Oryza sativa L.) reveals favorable alleles that improve grain Zn and Fe concentrations. PLoS ONE 10(7), e0131846 (2015).
Ishikawa, R. et al. Detection of quantitative trait loci controlling grain zinc content using Australian wild rice, Oryza meridionalis, a potential genetic resource for biofortification of rice. PLoS ONE 12(10), e0187224. https://doi.org/10.1371/journal.pone.0187224 (2017).
Khahani, B., Tavakol, E., Shariati, V. & Rossini, L. Meta-QTL and ortho-MQTL analyses identified genomic regions controlling rice yield, yield-related traits and root architecture under water deficit conditions. Sci. Rep. 11(1), 1–18 (2021).
Zhang, M. et al. Finding new addictive QTL for yield traits based on a high-density genetic map in hybrid rice. Plant Growth Regul. 93(1), 105–115 (2021).
Lu, K. et al. Quantitative trait loci controlling Cu, Ca, Zn, Mn and Fe content in rice grains. J. Genet. 87(3), 305 (2008).
Nagesh Ravindrababu, V., Usharani, G. & Reddy, T. D. Grain iron and zinc association studies in rice (Oryza sativa L.) F1 progenies. Arch. Appl. Sci. Res. 4(1), 696–702 (2012).
Jeong, O. Y. et al. Analysis of QTL responsible for grain iron and zinc content in doubled haploid lines of rice (Oryza sativa) derived from an intra-japonica cross. Plant Breed. https://doi.org/10.1111/pbr.12787 (2020).
Lalasa, L. M., Radhika, K., Neeraja, C. N., Babu, V. R. & Usharani, G. Molecular mapping of the chromosomal regions associated with zinc content in grains of rice (Oryza sativa L.) using microsatellite markers. J. Microbiol. Biotechnol. Res. 2(6), 900–905 (2012).
Bekele, B. D., Naveen, G. K., Rakhi, S. & Shashidhar, H. E. Genetic evaluation of recombinant inbred lines of rice (Oryza sativa L.) for grain zinc concentrations, yield-related traits and identification of associated SSR markers. Pak. J. Biol. Sci. 16(23), 1714–1721 (2013).
Li, A. et al. DH1, a LOB domain-like protein required for glume formation in rice. Plant Mol. Biol. 66(5), 491–502. https://doi.org/10.1007/s11103-007-9283-3 (2008).
Chen, X. et al. Inactivation of OsIRX10 leads to decreased Xylan content in rice culm cell walls and improved biomass saccharification. Mol. Plant 6(2), 570–573. https://doi.org/10.1093/mp/sss135 (2013).
Gao, S. et al. CYTOKININ OXIDASE/DEHYDROGENASE4 integrates cytokinin and auxin signaling to control rice crown root formation. Plant Physiol. 165(3), 1035–1046. https://doi.org/10.1104/pp.114.238584 (2014).
Sasaki, A. et al. A mutant gibberellin-synthesis gene in rice. Nature 416(6882), 701–702. https://doi.org/10.1038/416701a (2002).
Lin, H. et al. DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell 21(5), 1512–1525. https://doi.org/10.1105/tpc.109.065987 (2009).
Miura, K. et al. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nature Genet. 42(6), 545–549. https://doi.org/10.1038/ng.592 (2010).
Jiao, Y. et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat. Genet. 42(6), 541–544. https://doi.org/10.1038/ng.591 (2010).
Li, R., Li, M., Ashraf, U., Liu, S. & Zhang, J. Exploring the relationships between yield and yield-related traits for rice varieties released in China from 1978 to 2017. Front. Plant Sci. https://doi.org/10.3389/fpls.2019.00543 (2019).
Itoh, H. et al. A rice semi-dwarf gene, tan-ginbozu (D35), encodes the gibberellin biosynthesis enzyme, ent-Kaurene oxidase. Plant Mol. Biol. 54(4), 533–547. https://doi.org/10.1023/b:plan.0000038261.21060.47 (2004).
Guo, M. et al. Clustered spikelets 4, encoding a putative cytochrome P450 protein CYP724B1, is essential for rice panicle development. Ch. Sci. Bull. 59(31), 4050–4059. https://doi.org/10.1007/s11434-014-0568-z (2014).
Aswidinnoor, H., Listiyanto, R. & Suwarno, W. B. Genetic architecture of new plant type rice (Oryza sativa L.) lines based on a 12-years multi-experiment. SABRAO J. Breed. Genet. 55(4), 1025–1037. https://doi.org/10.54910/sabrao2023.55.4.2 (2023).
Aswidinnoor, H. et al. Stability analysis, agronomic performance, and grain quality of elite new plant type rice lines (Oryza sativa L.) developed for tropical lowland ecosystem. Front. Sustain. Food Syst. https://doi.org/10.3389/fsufs.2023.1147611 (2023).
Fu, F. & Xue, H. Coexpression analysis identifies rice starch regulator1, a rice AP2/EREBP family transcription factor, as a novel rice starch biosynthesis regulator. Plant Physiol. 154(2), 927–938. https://doi.org/10.1104/pp.110.159517 (2010).
Yuan, L., Yang, S., Liu, B., Zhang, M. & Wu, K. Molecular characterization of a rice metal tolerance protein, OsMTP1. Plant Cell Rep. 31(1), 67–79. https://doi.org/10.1007/s00299-011-1140-9 (2011).
Takahashi, R., Bashir, K., Ishimaru, Y., Nishizawa, N. & Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal. Behavior. 7(12), 1605–1607 (2012).
Yin, M. et al. Dissecting the genetic basis of yield traits and validation of a novel quantitative trait locus for grain width and weight in rice. Plants 13(6), 770 (2024).
Tao, Y. et al. Exploitation of heterosis loci for yield and yield components in rice using chromosome segment substitution lines. Sci. Rep. 6(1), 36802 (2016).
Chen, X. et al. Association mapping of thousand grain weight using SSR and SNP markers in rice (Oryza sativa L.) across six environments. Trop. Plant Biol. 14, 143–155 (2021).
Sun, L. et al. Identification of quantitative trait loci for grain size and the contributions of major grain-size QTLs to grain weight in rice. Mol. Breed. 31, 451–461 (2013).
Sun, P. et al. Dissection and validation of quantitative trait loci (QTLs) conferring grain size and grain weight in rice. Euphytica 220(3), 43 (2024).
Zhou, Y. et al. Fine mapping of five grain size QTLs which affect grain yield and quality in rice. Int. J. Mol. Sci. 25(8), 4149 (2024).
IRRI. Standard evaluation system for rice (SES). International Rice Research Institute, Manila, Philippines (2002).
Ding, Y. et al. SDG714, a histone H3K9 methyltransferase, is involved in Tos17 DNA methylation and transposition in rice. Plant Cell 19(1), 9–22 (2007).
Jiang, C. J. et al. Suppression of the rice fatty-acid desaturase gene OsSSI2 enhances resistance to blast and leaf blight diseases in rice. Mol. Plant Microbe Interact. 22(7), 820–829 (2009).
Hu, Y. et al. Rice histone deacetylase genes display specific expression patterns and developmental functions. Biochem. Biophys. Res. Commun. 388(2), 266–271 (2009).
Xiong, G. S. et al. LEAFY HEAD2, which encodes a putative RNA-binding protein, regulates shoot development of rice. Cell Res. 16(3), 267–276 (2006).
Kawakatsu, T. et al. PLASTOCHRON2 regulates leaf initiation and maturation in rice. Plant cell 18, 612–625 (2006).
Song, Y., Wang, L. & Xiong, L. Comprehensive expression profiling analysis of OsIAA gene family in developmental processes and in response to phytohormone and stress treatments. Planta 229, 577–591 (2009).
Lee, S. K. et al. Loss of cytosolic fructose-1, 6-bisphosphatase limits photosynthetic sucrose synthesis and causes severe growth retardations in rice (Oryza sativa). Plant Cell Environ. 31(12), 1851–1863 (2008).
Peng, B. et al. OsAAP6 functions as an important regulator of grain protein content and nutritional quality in rice. Nat. Commun. 5(1), 4847 (2014).
Wu, Y. et al. CLUSTERED PRIMARY BRANCH 1, a new allele of DWARF11, controls panicle architecture and seed size in rice. Plant Biotechnol. J. 14, 377–386 (2016).
Ohmori, Y.WOX4 is required for meristem function to maintain the undifferentiated state and is negatively regulated by the CLE gene FCP1 in rice. Plant Cell 25(1), 229–241 (2013).
Zhao, Z. et al. A role for a dioxygenase in auxin metabolism and reproductive development in rice. Dev. Cell 27(1), 113–122 (2013).
Zhu, X. & Xiong, L. Putative megaenzyme DWA1 plays essential roles in drought resistance by regulating stress-induced wax deposition in rice. Proc. Natl. Acad. Sci. 110(44), 17790–17795 (2013).
Fujii, S. & Toriyama, K. Suppressed expression of RETROGRADE-REGULATED MALE STERILITY restores pollen fertility in cytoplasmic male sterile rice plants. Proc. Natl. Acad. Sci. 106(23), 9513–9518 (2009).
Miao, C. et al. Central region component1, a novel synaptonemal complex component, is essential for meiotic recombination initiation in rice. Plant Cell 25(8), 2998–3009 (2013).
Duan, J. et al. OsMIOX, a myo-inositol oxygenase gene, improves drought tolerance through scavenging of reactive oxygen species in rice (Oryza sativa L.). Plant Sci. 196, 143–151 (2012).
Zhang, H., Meltzer, P. & Davis, S. RCircos: An R package for circos 2D track plots. BMC Bioinform. 14(1), 244. https://doi.org/10.1186/1471-2105-14-244 (2013).
Huang, J. et al. OsLOX2, a rice type I lipoxygenase, confers opposite effects on seed germination and longevity. Transgenic Res. 23, 643–655 (2014).
Onda, Y., Kumamaru, T. & Kawagoe, Y. ER membrane-localized oxidoreductase Ero1 is required for disulfide bond formation in the rice endosperm. Proc. Natl. Acad. Sci. 106(33), 14156–14161 (2009).
Kappara, S., Neelamraju, S. & Ramanan, R. Down regulation of a heavy metal transporter gene influences several domestication traits and grain Fe–Zn content in rice. Plant Sci. 276, 208–219 (2018).
Murray, M. G. & Thompson, W. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 8(19), 4321–4326 (1980).
Elshire, R. J. et al. A robust, simple genotyping-by-sequencing (GBS) approach for high diversity species. PLoS ONE 6(5), e19379 (2011).
Glaubitz, J. C. et al. TASSEL-GBS: A high capacity genotyping by sequencing analysis pipeline. PLoS ONE 9(2), 0090346. https://doi.org/10.1371/journal.pone (2014).
Meng, L., Li, H., Zhang, L. & Wang, J. QTL IciMapping: Integrated software for genetic linkage map construction and quantitative trait locus mapping in biparental populations. Crop J. 3(3), 269–283 (2015).
Kosambi, D. D. The estimation of map distances from recombination values. Ann. Eugen. 12, 172–175 (1944).
Churchill, G. A. & Doerge, R. Empirical threshold values for quantitative trait mapping. Genetics 138(3), 963–971 (1994).
Acknowledgements
The authors are grateful to the Biofortification group of IRRI for its invaluable help in the conduct of the study. MICC wishes to thank ADB-JSP-IRRI and ICropS, UPLB, and ADP thanks PhilRice for the scholarship grant. TKH would like to thank NATP project for providing scholarship for his PhD.
Author information
Authors and Affiliations
Contributions
MICC led the conduct of the experiment, analyzed the data, and prepared the draft manuscript. ADP, AA, and MAIA were involved in conducting field experiments and gathering phenotypic and genotypic data. ADP, TKH, GJ, BC thoroughly edited and contributed significantly to the improvement of the manuscript. BPMS conceptualized the study, designed the experiments, and thoroughly edited the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Calayugan, M.I.C., Hore, T.K., Palanog, A.D. et al. Deciphering the genetic basis of agronomic, yield, and nutritional traits in rice (Oryza sativa L.) using a saturated GBS-based SNP linkage map. Sci Rep 14, 18024 (2024). https://doi.org/10.1038/s41598-024-67543-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-67543-3
- Springer Nature Limited