
Abstract
The development of genotypes that can tolerate high levels of salt is crucial for the efficient use of salt-affected land and for enhancing crop productivity worldwide. Therefore, incorporating salinity tolerance is a critical trait that crops must possess. Salt resistance is a complex character, controlled by multiple genes both physiologically and genetically. To examine the genetic foundation of salt tolerance, we assessed 16 F1 hybrids and their eight parental lines under normal and salt stress (15 dS/m) conditions. Under salt stress conditions significant reduction was observed for plant height (PH), bolls/plant (NBP), boll weight (BW), seed cotton yield (SCY), lint% (LP), fiber length (FL), fiber strength (FS), potassium to sodium ratio (K+/Na+), potassium contents (K+), total soluble proteins (TSP), carotenoids (Car) and chlorophyll contents. Furthermore, the mean values for hydrogen peroxide (H2O2), sodium contents (Na+), catalase (CAT), superoxide dismutase (SOD), peroxidase (POD), and fiber fineness (FF) were increased under salt stress. Moderate to high heritability and genetic advancement was observed for NBP, BW, LP, SCY, K+/Na+, SOD, CAT, POD, Car, TSP, FL, and FS. Mean performance and multivariate analysis of 24 cotton genotypes based on various agro-physiological and biochemical parameters suggested that the genotypes FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, along with crosses FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333 achieved the maximum values for K+/Na+, K+, TSP, POD, Chlb, CAT, Car, LP, FS, FL, PH, NBP, BW, and SCY under salt stress and declared as salt resistant genotypes. The above-mentioned genotypes also showed relatively higher expression levels of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 at 15 dS/m and proved the role of these ERF genes in salt tolerance in cotton. These findings suggest that these genotypes have the potential for the development of salt-tolerant cotton varieties with desirable fiber quality traits.
Similar content being viewed by others
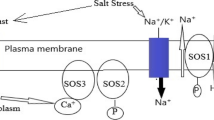


Introduction
Soil salinization is a significant obstacle to the sustainable growth of global agriculture and a major threat to the ecological health of the soil. Approximately 37% of all cultivated lands globally are categorized as sodic. It is estimated that almost 50% of the world’s (2.5 billion hectares) of irrigated lands are significantly impacted by salinization1. Additionally, every year nearly four million acres of cropland are abandoned due to severe salt damage2. Besides the existing salt-affected soil, ongoing secondary salinization in arable land is caused by several factors such as climate change, excessive irrigation, irrigation with saline water, and excessive use of mineral fertilizers3. When crop plants experience salt stress, several physiological and biochemical changes occur, including water imbalance, ionic toxicity, nutrient deficiency, oxidative stress and hormonal imbalance which ultimately reduce the crop yield4.
Cotton (Gossypium spp.), a main agro-industrial crop, is grown worldwide for its significant production of natural fibers and seed oil. India, China, USA, Brazil, Pakistan, and Australia are among the major cotton-growing countries. Despite being in the top 5 globally, Pakistan has a low yield compared to other leading countries due to various biotic and abiotic factors (drought, heat, and salinity)5. Among all these abiotic stresses salinity is a major threat to cotton production. In Pakistan due to competition between cereals and other cash crops, the cultivation of cotton is being moved to saline and alkaline soils. Besides these factors, the increasing demand of natural fiber for textiles, due to population growth and societal reasons, it’s essential to produce high-quality cotton fibers in abundance6.
Cotton is tolerant to moderate salinity (7.7 ds/m), but increased soil salinity negatively affects its growth and yield. At the plant level, salinity disrupts various physiological and biochemical processes like ionic toxicity, photosynthesis process, and oxidative injury. As a consequence of this phenomenon, there is inhibition of cell division and expansion in leaf surface area, leading to developmental changes, metabolic adaptations, and decreased stem growth and root cell proliferation. These factors directly contribute to a reduction in the NBP, BW, SCY and fiber quality parameters7,8. Salt stress triggers the production of higher levels of reactive oxygen species (ROS) such as superoxide radicals (O2−), H2O2, singlet oxygen (1O2) and hydroxyl radicals (OH−), which induce oxidative stress in the plant9. The production of ROS at higher level leads to oxidation and damage of plant cell organelles like mitochondria, peroxisomes and chloroplasts. In response, the cell produces antioxidant enzymes such as superoxide dismutase (SOD), peroxidases (POD), catalase (CAT), and non-enzymatic antioxdants10 to counter the detrimental effects of oxidation, safeguard the organelles, and uphold the normal functioning of the cell11.
Genetic switches also referred to as transcription factors (TFs), attach themselves to specific cis-acting elements, regulating downstream expression of the gene and aiding in the management of stress-induced impairment12. In various plant species, many TFs are either induced or suppressed after exposure to salinity stress, facilitating adaptation to saline environments13. The AP2/ERF gene family is a plant-specific group of TFs characterized by an AP2/ERF-type DNA-binding domain that consists of roughly 60–70 amino acids14. This family was initially discovered in the Arabidopsis homeotic gene APETALA2 (AP2). The AP2/ERF gene family is subdivided into four subfamilies, namely AP2, ERF, RAV, and DREB. The AP2 subfamily differs from the other three subfamilies in that it possesses double AP2/ERF domains, while the other three subfamilies have a single AP2/ERF domain15. In the past two decades, the ERF family of genes has garnered significant importance, as overexpression of ERF genes in various plants has been shown to enhance tolerance against abiotic stresses and pathogen resistance in genetically modified plants. In the previous study, based on transcriptome and qRT-PCR analysis, several potential candidate genes (Ghi-ERF-6D.1, Ghi-ERF-2D.6, Ghi-ERF-12D.13, Ghi-ERF-11D.5, and Ghi-ERF-7A.6) that promote salinity tolerance in upland cotton13.
Salinity poses a significant economic and environmental challenge to the global cotton industry, which highlights the need to enhance the salt tolerance of cotton and develop germplasm capable of producing higher yields under changing climatic situations16. To mitigate the impact of salinity stress, several strategies have been employed to enhance resilience in cotton plants. Among these, the development of salt-tolerant germplasm is regarded by cotton breeders as the most effective and sustainable solution16. Utilizing conventional breeding techniques to screen available germplasm for morphological, physiological, and biochemical traits can leverage natural variations and significantly enhance cotton plant tolerance to salt stress. Measuring the levels of antioxidant enzymes produced in response to salt stress can serve as an effective screening strategy for identifying germplasm with the potential to yield higher crops under saline conditions8. Plant’s ability to resist toxic effects of NaCl salinity depends on genetic make-up of plants or variations in physiological processes which enable the plants to cope with salt stress, which include degree of ion exclusion, tolerance to osmotic stress, and tissue tolerance. Moreover, the mechanism of salt tolerance varies with the type of species, type of cultivar of the same species, and plant developmental stage which makes it more complex17. Consequently, devising screening techniques and parameters for evaluating salt tolerance becomes a challenging endeavor18. Experts emphasize the need to transfer identified physiological and genetic components of salt tolerance into crop cultivars adapted to local environments. Identified salt-tolerant crop cultivars with physiological traits contributing in salt tolerance can be used as donors in breeding for salt tolerance. However, considerable genetic variability is a prerequisite for any specific physiological trait, based on which selection or screening can be made. In this study, we used pure lines which are acclimatized to various ecological zones of Pakistan. The present research aims to achieve three objectives: (i) identification of salt-tolerant cotton cultivars with novel source of salt tolerance from local germplasm, (ii) development of a selection criterion that incorporates agro-physiological and biochemical characters for the production of salt-resistant cotton cultivars, and (iii) evaluation of the expression levels of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 genes in salt-tolerant cotton genotypes.
Materials and methods
The experimental work was carried out on the research premises of FB Genetics, Four Brothers Group, Pakistan during the cropping year 2021. The experimental site is located at 74˚ east longitude and 31˚ north latitude. Under normal field conditions, the F0 seeds of eight genotypes were sown and then, in a Line × Tester manner, were crossed. Four genotypes (namely FBS-FALCON, FBS-SMART, FBG-222 and Barani-333) were used as lines and four genotypes (namely JSQ-White Hold, FBG-333, Barani-222 and Ghauri-3) were used as testers (Table 1). When the cotton genotypes were flowering, they were crossed in a line × tester way (4 × 4). To create selfed seeds, certain buds were also wrapped in glassine bags. The mature selfed and crossed bolls were harvested and preserved. Subsequently, the cotton fibers from each group were processed separately using a single roller ginning machine (Testex, Model: TB510C) after being picked. In the next growing season, 24 genotypes (including 8 parents and their 16F1 hybrids) were grown in containers with normal soil (1.9 dS/m) under RCBD with three replications. After the emergence of seedlings, healthy specimens were selected for each genotype in every replication, all of which were subsequently subjected to salt stress through irrigation water with a known EC. The Analysis of soil properties of the experimental site was also conducted (Table 2). The salt stress was induced following a well-established procedure8. This method involves gradually increasing the salinity to a target level of ~ 15 dS/m in two distinct phases to avert seedling injury and ensure survival. Sodium chloride (NaCl) was calculated and added to the groundwater using the U.S. Salinity Lab formula19.
In this equation, TSS (Total Soluble Salts) represents the difference in EC (desired EC—initial EC) multiplied by 12.66, and the saturation percentage of the soil was established at 42%.
During the initial phase, the 2-week-old seedlings were subjected to a salinity level of 7.5 dS/m, which necessitated the addition of approximately 17.74 g of NaCl to each 10 kg pot. Subsequently, in the second phase, the 4-week-old seedlings were subjected to an escalated salinity level of 15 dS/m, which required a further addition of about 23.72 g of NaCl per 10 kg pot. Thus, approximately 41.46 g (17.74 g + 23.72 g) of NaCl was added per 10 kg pot in the two phases to attain the desired salinity level. After reaching this level, no further salt was added. Following this precise modulation of salinity, standard agricultural procedures were then followed for irrigation. This careful approach enabled us to achieve gradual increments in salinity while preventing seedling injury and ensuring survival throughout the experiment.
All agronomic procedures were appropriately carried out from the time of sowing until harvesting. Figure S1 displays the environmental conditions, including temperature, humidity, and rainfall, during crop growth. Upon reaching maturity (i.e., after 140 days of growth and > 80% completion of boll formation), the following parameters were measured from five chosen plants from each replication of both normal and saline treatment: plant height, bolls/plant, boll weight, seed cotton yield/plant, lint%, H2O2, SOD, POD, CAT, TSP, chlorophyll contents (a & b), carotenoids, fiber strength, fiber length, fiber fineness, Na+, K+ and K+/Na+ ratio.
Ion analysis
The analysis of sodium and potassium ions involved grinding hot air-dried leaves using a pestle and mortar. Concentrated nitric acid and sulfuric acid in a 2:1 ratio were used for the digestion of ground leaves on the hot plate. Then digested samples were cooled down by adding distilled water at room temperature. A flame photometer (410 Flame Photometer) was used to take out readings and then the K+ /Na+ ratio was calculated.
Hydrogen peroxide (µmol/g-FW)
Bernt and Bergmeyer20 method was used to measure the hydrogen peroxide of treated and control samples. 0.5 g of leaf sample was homogenized using liquid nitrogen for each control and treated group. 1.5 ml of 100 mM potassium phosphate buffer (pH 6.8) was used for the suspension of ground leaves then the suspension was centrifuged at 18,000×g for 20 min at 40 °C. The supernatant (0.25 ml) was taken out and mixed with 1.25 ml of peroxidase reagent containing; 40 μg peroxidase/ml, 0.005% (w/v) O-dianizidine and 83 mM potassium phosphate buffer (pH 7.0) at 30 °C to initiate the reaction. After 10 min, the reaction was halted by adding 0.25 ml of 1 N perchloric acid and centrifuging the mixture at 5000×g for 5 min. A spectrophotometer (NanoDrop™ 8000 Spectrophotometer Thermo Fisher Scientific, Sweden) was used to measure the absorbance at 436 nm and the amount of H2O2 was determined based on an extinction coefficient of 39.4 mM−1/cm21. The SOD activity was measured in terms of enzyme units that inhibited the photochemical reduction of nitro blue tetrazolium (NBT).
Superoxide dismutase
To quantify SOD, a reaction mixture containing 100 μl of enzyme extract, potassium phosphate buffer (pH 5), 200 μl of methionine, 200 μl of Triton X, 100 μl of NBT, and 800 μl of distilled water was prepared. The mixture was exposed to ultraviolet light for 15 min, followed by the addition of 100 μl of Riboflavin. The absorbance at 560 nm was measured using a spectrophotometer.
Peroxidase (U/mg protein)
The same enzyme extracted for measuring SOD was used to measure POD values. It is calculated as the number of enzyme units that oxidized guaiacol. The reaction mixture was prepared by mixing 100 μl H2O2 (40 mM), 100 μl guaiacol (20 mM) with 100 μl of enzyme extract, and 800 μl potassium phosphate buffer (pH 5) in Eppendorf tube and a spectrophotometer was used to measure absorbance at 470 nm wavelength22.
Total Soluble Proteins (mg/g-FW)
To determine the Total Soluble Proteins (TSP), leaf tissue extraction was carried out using vortex and centrifugation in a phosphate buffer (pH 4). Furthermore, a 40 μl portion of the same enzyme extract was mixed with 160 μl of Bradford reagent, added to an ELISA plate, and subjected to spectrophotometer readings at a wavelength of 595 nm absorbance23.
Catalase (U/mg protein)
Catalase activity was determined by measuring the amount of H2O2 consumed and transformed into H2O and O2. Spectrophotometer values were recorded at 240 nm absorbance to calculate CAT activity22.
Chlorophyll contents and carotenoids assay
The chlorophyll and carotenoid contents were measured by following24. 0.5 g of cotton leaf sample underwent crushing in 8–10 ml of 80% acetone (volume/volume). Subsequently, homogenization occurred through filter paper, and the absorbance of the resulting solution was measured at wavelengths of 645 and 663 nm. The quantification of chlorophyll a, chlorophyll b, and carotenoids followed this procedure.
The chlorophyll a & b and carotenoids were estimated as follows,
where,
W = weight of leaf sample, V = volume of sample, Em = 2500.
qRT-PCR of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 in tolerant cotton genotypes
The newly emerged leaves of control and stressed (15 dS/m) genotypes at two leaves stage were sampled. The RNA was extracted using a kit protocol (RNAprep Pure Plant Kit by Tiangen, Beijing, China) and the quality and integrity of the RNA were checked using gel electrophoresis and NanoDrop 2000 spectrophotometer (Thermo Scientific, USA) respectively. The cDNA was prepared using (PrimeScript® RT Reagent Kit, Takara Biotechnology Co., China). Three repeats of both genes were used technically. The real-time differential expression of mRNA of both genes was measured using qRT-PCR (Maxima SYBR Green)/ROX qPCR Master mix (2X), cat#K0221, Thermo scientific, USA). Gene normalization was obtained using GAPDH as an internal control. The gene-specific primers were used for the expression of both genes (Table 3). In a PCR reaction gene-specific short-length primers for ERF2 and ERF7 gene were used. A total volume of PCR reaction mixture was prepared in a volume of 25 µl. General PCR conditions using short-length primers were as follows in this study: firstly, one cycle of initial denaturation at 95 °C for 3 min; secondly, 28 cycles of denaturation at 95 °C for 45 s, annealing at 66 °C for 45 s, and extension at 72 °C for 2 min; finally, one cycle of final extension at 72 °C for 10 min.
Fiber quality parameters
A seed cotton sample was weighed and subjected to a single roller ginning machine, to separate lint from the seeds. The percentage (%) of the lint was then calculated. This was achieved by dividing the weight of the lint by the weight of the seed cotton in the sample, the obtained value was then finally expressed as a percentage. Furthermore, employing a high-volume instrument (HVI-900, USTER, USA), the resulting lint was given an in-depth analysis to determine characteristics such as fiber strength, fiber fineness and length parameters.
Statistical analysis
The split-plot analysis of variance (ANOVA) was performed with two factors25. The statistical tools prcomp, ggplot2, and Hmisc from R 4.1.1 were used to conduct principal components, and correlation analyses respectively on the mean data. Falconer and Mackay’s method was followed to estimate heritability and genetic advance26,27.
Ethics approval and consent to participate
Study protocol complies with relevant institutional, national, and international guidelines and legislation.
Results
The results of the ANOVA suggested significant variations among parents and their F1 hybrids under salt stress conditions. These differences indicated the existence of genetic diversity in the studied germplasm for salinity tolerance, as shown in (Table 4). Moreover, the treatment and Genotypes × Treatment interaction for all characteristics were highly significant, indicating that all genotypes responded differently to salt stress (Table 4). All cotton genotypes experienced a negative impact on their agronomic and yield-related traits under salt stress. The reduction was observed in agronomic traits such as PH, NBP, BW, SCY, lint%, FL, and FS as shown in (Table 5). Interestingly, fiber fineness (FF) was increased under saline environments. Furthermore, the mean values for H2O2, Na+, CAT, POD, and SOD were increased under salt stress, while the mean values for K+/Na+, K+, TSP, Chla and b were decreased under saline conditions.
The GAM was classified into three categories: low (0–10%), moderate (10–20%), and high (20% or higher). In terms of heritability, values exceeding 80% were considered very high, values ranging from 60 to 79% were moderately high, values between 40 and 59% were medium, and values lower than 40% were regarded as low28. Under saline conditions, high genetic advance as a percentage of the mean (GAM) was observed for BW, CAT, Chla, Chlb, FF, NBP, POD, SCY, SOD and TSP whereas moderate GAM was observed for PH, LP, Na+, K+, FS, FL, and Car. The heritability estimates for BW, CAT, Car, Chlb, FL, FS, K+, K+/Na+, LP, NBP, PH, POD, SOD, and TSP, were very high whilst moderate heritability estimates were observed for SCY, Na+, H2O2, FF, and Chla under saline conditions (Table 5).
Mean performance of agronomic, and fiber quality traits
Under the influence of salt stress conditions (15 ds/m), the growth and agronomic characteristics of the examined genotypes were negatively impacted. Notably, the effects of salt stress were particularly evident in terms of plant height, the number of bolls per plant, and boll weight, lint percentage, fiber fineness, and fiber strength for all the genotypes studied along with crosses, as depicted in (Figs. 1 and 2). Under salt stress conditions (15 ds/m), certain genotypes and crosses demonstrated remarkable growth and yield performance compared to other studied genotypes. Genotypes FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, along with crosses FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333 displayed notable resilience and exhibited superior performance for following traits such as PH, NBP, BW, SCY, lint%, FL, FF, and FS. Conversely, the genotypes (FBS-Smart, FBG-222, FBG-333, and Barani-222), and crosses (FBS-FALCON × Ghauri-3, FBS-SMART × JSQ-White Hold, FBS-SMART × Barani-222, FBS-SMART × Ghauri-3, FBG-222 × JSQ-White Hold, FBG-222 × Ghauri-3, Barani-333 × JSQ-White Hold, and Barani-333 × Ghauri-3) demonstrated lower performance for agronomic and fiber quality traits under salt stress conditions. Notably, among the studied crosses (FBS-FALCON × FBG-333, FBS-FALCON × Barani-222, FBS-SMART × FBG-333, and Barani-333 × Barani-222), revealed moderate performance for PH, NBP, BW, SCY, lint%, FL, FF, and FS under salt stress conditions. These specific crosses demonstrated moderate resilience to the adverse effects of salt stress, indicating their potential for further exploration and breeding programs (Figs. 1 and 2).

Mean performance of cotton genotypes under control and salinity stress for (A) BW, Boll weight (g); (B) NBP, Number of bolls; (C) PH, Plant height (cm); (D) SCY, Seed cotton yield (g/plant).

Mean performance of cotton genotypes under control and salinity stress for (A) Lint (%) = Lint percentage; (B) FF , Fiber fineness (µg/inch); (C) FS, Fiber strength (g/tex); (D) FL, Fiber length (mm).
Biochemical traits
The biochemical traits such as chlorophyll a (Chla), chlorophyll b (Chlb), total soluble proteins (TSP), and carotenoid (CAR) exhibited a statistically significant decrease in all genotypes under salt stress (15 ds/m) (Fig. 3). The application of salt stress treatment led to a more significant reduction in Chla, Chlb, TSP, and CAR levels in specific genotypes and crosses, such as FBS-Smart, FBG-222, FBG-333, Barani-222, FBS-FALCON × Ghauri-3, FBS-SMART × JSQ-White Hold, FBS-SMART × Barani-222, FBS-SMART × Ghauri-3, FBG-222 × JSQ-White Hold, FBG-222 × Ghauri-3, Barani-333 × JSQ-White Hold, and Barani-333 × Ghauri-3. However, the genotypes FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333 exhibited the ability to maintain moderate level of Chla, Chlb, TSP, and CAR (Fig. 3). Furthermore, certain genotypes and crosses, such as FBS-FALCON × FBG-333, FBS-FALCON × Barani-222, FBS-SMART × FBG-333, and Barani-333 × Barani-222 exhibited less reduction in Chla, Chlb, TSP, and CAR level compared to susceptible genotypes (Fig. 3).

Mean performance of cotton genotypes under control and salinity stress for (A) Chla, Chlorophyll a (mg/g FW); (B) Chlb, Chlorophyll a (mg/g FW); (C) Car, Carotenoids (mg/g FW); (D) TSP, Total soluble proteins (mg/g FW).
The application of NaCl treatment had a significant impact on the ionic homeostasis of plants, particularly on Na+, K+, and the K+/Na+ ratio. Notably, the Na+ content in NaCl-treated plants was found to be significantly higher compared to the control group, indicating an increase in sodium accumulation as a result of the treatment. The mean graph of (Fig. 4) depicted that certain genotypes and crosses, including FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333 lower increase in sodium contents compared other genotypes. Compared to other genotypes, these genotypes also revealed less reduction in potassium contents. Consequently, these genotypes and crosses maintained a higher ratio of potassium to sodium content, similar to that of the control group and performed well for biochemical, agronomic and fiber quality characters under saline environments (Fig. 4). However, specific crosses such as FBS-FALCON × FBG-333, FBS-FALCON × Barani-222, FBS-SMART × FBG-333, and Barani-333 × Barani-222 demonstrated a moderate accumulation of Na+, and exhibited moderate potassium to sodium ratio. The mean graph reveals that the genotypes and crosses, including FBS-Smart, FBG-222, FBG-333, Barani-222, FBS-FALCON × Ghauri-3, FBS-SMART × JSQ-White Hold, FBS-SMART × Barani-222, FBS-SMART × Ghauri-3, FBG-222 × JSQ-White Hold, FBG-222 × Ghauri-3, Barani-333 × JSQ-White Hold, and Barani-333 × Ghauri-3 exhibited higher levels of Na+, along with a lower potassium to sodium ratio under salt stress conditions and declared as salt susceptible genotypes (Fig. 4).

Mean performance of cotton genotypes under control and salinity stress for (A) Na+, Sodium contents (B) K+, Potassium contents (C) K+/Na+, Potassium-to-sodium ratio (%).
Under salt treatment (15 ds/m), the cotton genotypes displayed a significant rise in the levels of reactive oxygen species (ROS). In contrast to the control group, all genotypes exhibited noticeable increases in the levels of H2O2, indicating the presence of oxidative stress induced by salt stress. However, it is noteworthy that susceptible genotypes and specific crosses, including FBS-Smart, FBG-222, FBG-333, Barani-222, FBS-FALCON × Ghauri-3, FBS-SMART × JSQ-White Hold, FBS-SMART × Barani-222, FBS-SMART × Ghauri-3, FBG-222 × JSQ-White Hold, FBG-222 × Ghauri-3, Barani-333 × JSQ-White Hold, and Barani-333 × Ghauri-3, exhibited greater levels of oxidative damage and showed higher value for H2O2 under saline conditions (Fig. 5A). This observation suggests that these particular cultivars possess lower tolerance to salt stress when compared to tolerant genotypes such as FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333. In contrast, moderate levels of H2O2 were observed in the following crosses such as FBS-FALCON × FBG-333, FBS-FALCON × Barani-222, FBS-SMART × FBG-333, and Barani-333 × Barani-222 (Fig. 5A). These genotypes also showed a moderate level of SOD, POD, and CAT activities under saline environmental conditions. Our findings indicate that under the salt stress condition of (15 ds/m), there was a higher level of SOD, CAT, and POD activity observed compared to the control group. The tolerant genotypes exhibited optimal levels of SOD, CAT, and POD activity under saline conditions. The tolerant genotypes FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333 revealed high performance for SOD, POD and CAT as compared to moderately tolerant and susceptible genotypes. The susceptible genotypes FBS-Smart, FBG-222, FBG-333, Barani-222, FBS-FALCON × Ghauri-3, FBS-SMART × JSQ-White Hold, FBS-SMART × Barani-222, FBS-SMART × Ghauri-3, FBG-222 × JSQ-White Hold, FBG-222 × Ghauri-3, Barani-333 × JSQ-White Hold, and Barani-333 × Ghauri-3 showed lesser increase in SOD, POD and CAT activities under saline conditions (Fig. 5B, C, D).

Mean performance of cotton genotypes under control and salinity stress for (A) H2O2, Hydrogen peroxide (µmol/g FW) (B) SOD, Superoxide dismutase (U/mg protein) (C) POD, Peroxidase (U/mg protein) (D) CAT, Catalase (U/mg protein).
Correlation analysis
The relationship between various morpho-physiological and biochemical traits is crucial in determining the most suitable genotype under both normal and saline environments. The Pearson correlation coefficients were computed separately for normal and saline conditions. Under normal conditions, FL, Na+, TSP, CAT, FS, PH, FF, POD, and SOD showed positive association with each other. Lint % revealed a significant positive relationship with FL, TSP, CAT, FS, FF, POD, Chla, Chlb, BW, and CAR contents under control conditions (Fig. 6). Under salt stress conditions, H2O2 and Na+ revealed a significant negative relationship with all morphological, agronomical (SCY, BW, NBP, and LP), fiber quality traits (FL, FS, & FF) and physiological characters (CAT, TSP, SOD, POD, Car, chlorophyll contents, K+/Na+, and K+ respectively (Fig. 2). Under salt stress environments all agronomic characters (SCY, BW, NBP, and LP) revealed positive relationship with physiological traits CAT, TSP, POD, Car, chlorophyll contents, K+/Na+, and K+ respectively. The fiber quality parameters showed a positive association with all antioxidant and biochemical traits under salt stress conditions (Fig. 7).

Corplot for various traits under control conditions. BW, Boll weight; NBP, Number of bolls; PH, Plant height; SCY, Seed cotton yield; Lint (%), Lint percentage; FF, Fiber fineness; FS, Fiber strength; FL, Fiber length; Chla, Chlorophyll a; Chlb, Chlorophyll a (mg/g FW); (C) Car, Carotenoids (mg/g FW); (D) TSP, Total soluble proteins; Na+, Sodium; K+, Potassium; K+/Na+, Sodium-to-potassium ratio; H2O2, Hydrogen peroxide; SOD, Superoxide dismutase; POD, Peroxidase; CAT, Catalase.

Corplot for various traits under salinity stress. BW, Boll weight; NBP, Number of bolls; PH, Plant height; SCY, Seed cotton yield; Lint (%), Lint percentage; FF, Fiber fineness; FS, Fiber strength; FL, Fiber length; Chla, Chlorophyll a; Chlb, Chlorophyll a (mg/g FW); (C) Car, Carotenoids (mg/g FW); (D) TSP, Total soluble proteins; Na+, Sodium; K+ , Potassium; K+/Na+, Sodium-to-potassium ratio; H2O2, Hydrogen peroxide; SOD, Superoxide dismutase; POD, Peroxidase; CAT, Catalase.
Principal component analysis (PCA) for agro-physiological, biochemical, and fiber quality traits
PCA is a statistical approach for analyzing and simplifying complex datasets. We used PCA to evaluate the correlation among variables and the genetic diversity in studied accessions and their association with studied characters under normal and saline conditions. The PC-1 and PC-2 explained 37.6% and 13% of the whole variation under control conditions, respectively (Fig. 8). The relative distance of variables from the origin in PC1 and PC2 showed how much each variable contributes to the total variation among the studied accessions. Under normal conditions, the biplot demonstrated that genotypes of quadrant-IV had high potential for SCY, LP, BW, chlorophyll contents, POD, FS, SOD, K+, and carotenoid contents. The genotypes of quadrant-III performed better for NBP, TSP, CAT, PH, Na+, and FL (Fig. 8). The genotypes of quadrant-I revealed good performance for potassium to sodium ratio whereas the genotypes of quadrant-II showed optimal performance for H2O2 under control conditions (Fig. 8).

Principal component analysis of cotton genotypes for various traits under control condition. BW, Boll weight; NBP, Number of bolls; PH, Plant height; SCY, Seed cotton yield; Lint (%), Lint percentage; FF, Fiber fineness; FS, Fiber strength; FL, Fiber length; Chla, Chlorophyll a; Chlb, Chlorophyll a (mg/g FW); (C) Car, Carotenoids (mg/g FW); (D) TSP, Total soluble proteins; Na+, Sodium; K+, Potassium; K+/Na+ , Sodium-to-potassium ratio; H2O2, Hydrogen peroxide; SOD, Superoxide dismutase; POD, Peroxidase; CAT, Catalase.
Under saline conditions, the biplot of PC-1 and PC-2, together account for 90.20% of the total variation. In the biplot, traits close to each other were positively associated with each other (Fig. 9). Under salt stress conditions, the biplot of PC1 and PC2, revealed a positive association between Na+ and H2O2 and the genotypes FBG-222, FBS-SMART, FBG-333, Barani-333 × JSQ-White Hold, FBS-SMART × Ghauri-3, Barani-333 × Ghauri-3, FBG-222 × JSQ-White Hold, FBS-SMART × JSQ-White Hold, FBS-SMART × Ghauri-3, FBS-FALCON × Ghauri-3, Barani-222, and FBS-SMART × Barani-222 showed a positive relationship with these traits and declared as salt susceptible. Whereas the genotypes FBS-FALCON × JSQ-White Hold, FBS-FALCON, JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, Barani-333, Ghauri-3, and Barani-333 × FBG-333 revealed a positive correlation with K+/Na+, K+, POD, SOD, CAT, TSP, Chla, Chlb, CAR, LP, FS, FL, FS, NBP, BW, and SCY and declared as salt tolerant (Fig. 9). The genotypes FBS-FALCON × FBG-333, FBS-FALCON × Barani-222, and FBS-SMART × FBG-333 showed significant association with FF under saline conditions. The insights obtained from PCA can be utilized to obtain valuable outcomes (Fig. 9).

Principal component analysis of cotton genotypes for various traits under salinity stress. BW, Boll weight; NBP, Number of bolls; PH, Plant height; SCY, Seed cotton yield; Lint (%), Lint percentage; FF, Fiber fineness; FS, Fiber strength; FL, Fiber length; Chla, Chlorophyll a; Chlb, Chlorophyll a (mg/g FW); (C) Car, Carotenoids (mg/g FW); (D) TSP, Total soluble proteins; Na+ , Sodium; K+, Potassium; K+/Na+, Sodium-to-potassium ratio; H2O2, Hydrogen peroxide; SOD, Superoxide dismutase; POD, Peroxidase; CAT, Catalase.
Examination of the expression of ERF genes (Ghi-ERF-2D.6 and Ghi-ERF-7A.6) by qPCR
The quantitative measurement of mRNA expression in the leaf of genotypes at salt stress levels of control and 15 ds/m was performed. As an internal control of gene normalization, the GAPDH was employed. In Fig. 10, the genes Ghi-ERF-2D.6 and Ghi-ERF-7A.6 showed relatively higher expression at 15 ds/m. In genotype JSQ-white Hold, the expression of Ghi-ERF-2D.6 is 1.5 folds higher at 15 ds/m than control whereas the expression of Ghi-ERF-7A.6 is 2.2 folds higher at 15 ds/m than control. The genotype Barani-333 × FBG-333 showed 2 folds higher expression of Ghi-ERF-2D.6 at 15 ds/m than control whereas the expression of Ghi-ERF-7A.6 is 2.7 folds higher at 15 ds/m than control. The expression of both Ghi-ERF-2D.6 and Ghi-ERF-7A.6 in genotype FBG-222 × Barani-222 was measured 2 folds higher at 15 ds/m than control. In genotype Ghauri, the expression of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 was 2.5 folds and 2.2 folds higher at 15 ds/m than control respectively. The genotype FBG-222 × FBG-333 showed 1.5 folds and 3 folds higher expression of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 at 15 ds/m than control respectively (Fig. 10). In genotype FBS-FALCON × JSQ-white Gold, the expression of Ghi-ERF-2D.6 is 2 folds higher whereas the expression of Ghi-ERF-7A.6 is 2.7 folds higher at 15 ds/m than control respectively. The genotype Barani-333 showed 2.5 folds and 3 folds higher expression of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 at 15 ds/m than control respectively. The expression of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 in genotype FBS-FALCON was measured 2.1 folds and 2.3 folds higher at 15 ds/m than control respectively. The expression level of both the genes was increased at 15 ds/m and showed significant tolerance against salt stress (Fig. 10). The PCR reaction using short length primers was run to detect the correct size of the gene, which corresponds 120 bp (Fig. 11).

Relative expression patterns of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 genes under normal and salinity stress (15 dS/m NaCl) treatment analyzed by qRT-PCR.

Detection of genes (ERF2 and ERF7) using short length primers; Lane M: shows molecular weight marker; Lane 1: shows positive control; lane 2, 3, 6, 7: shows negative control; lane 4: shows ERF2; Lane 5: shows ERF7 corresponding a size of 120 bp.
Discussion
Cotton is an economically important crop that is grown worldwide for its fiber and seed oil. However, cotton growth and productivity are often limited by various abiotic stresses, including salt stress4. Salt stress can have a significant impact on cotton physiology, leading to reduced growth, disrupted water and ionic balance, oxidative stress, and decrease in yield. Understanding these physiological effects is critical for developing strategies to mitigate the negative effects of salt stress and improve cotton productivity in salt-affected soils29,30.
A significant amount of work has been done so far to develop cotton cultivars that can tolerate stress. In this pursuit, plant breeders often depend on the genetic variability present in the available germplasm to select desirable traits31. To improve breeding programs aimed at producing salt-tolerant cotton genotypes, it is crucial to have current knowledge on genetic variability and heritability31. In this study, Line × Tester approach was used, where four lines and four testers were crossed, resulting in the production of 16 F1 hybrids. The success of improving crop plants genetically is dependent on the level of heritability of traits that are economically valuable32. Traits that exhibit high heritability and genetic advance are more likely to be transmitted to the next generation in greater proportions. When there is both a high h2b and high GAM, this can lead to genetic gains through the selection process33. In this study, traits such as NBP, BW, LP, SCY, K+/Na+, CAT, SOD, POD, TSP, Car, FL and FS demonstrated moderate to high heritability and GAM, suggesting the presence of additive gene action. These traits can be useful for early-stage selection of genotypes, which can then be utilized in breeding programs focused on improvement34. Under salt stress environment, significant reduction was observed in agronomic traits such as PH, NBP, BW, SCY, and lint%. Interestingly, fiber fineness (FF) was increased under saline environments. Furthermore, the mean values for H2O2, CAT, SOD, and POD, were increased under salt stress, while the mean values for TSP, Car, Chl a and b were decreased under saline conditions8. In a stress breeding program, measuring chlorophyll content is crucial in identifying salt-tolerant genotypes, as higher chlorophyll contents correlate with greater salt tolerance. In our study chlorophyll contents were reduced under salt stress conditions35. The reduction in chlorophyll contents, leads to increased production of H2O218. This creates oxidative stress due to the strong negative association between H2O2 and chlorophyll content under salt treatment36. One of the ways in which oxidative stress can reduce cotton yield under salt stress is by inhibiting photosynthesis. Salt stress-induced oxidative stress can damage the photosynthetic machinery, resulting in a decrease in the efficiency of photosynthesis. This, in turn, reduces the plant’s ability to produce energy and fix carbon dioxide, which are essential for growth and yield37,38,39. Sodium and potassium are essential ions that play critical roles in different biochemical processes in plants, including osmoregulation, ion homeostasis, and enzyme activation40. Salt stress can cause an influx of toxic ions such as sodium and chloride into the plant and decrease the potassium contents in plants, which disrupt ion homeostasis, leading to ion toxicity and osmotic stress3. This can impair the plant’s ability to maintain normal cellular function, resulting in reduced cotton yield and boll weight. In our study, the genotypes FBS-FALCON × Ghauri-3, FBS-SMART × JSQ-White Hold, FBS-SMART × Barani-222, FBS-SMART × Ghauri-3, FBG-222 × JSQ-White Hold, FBG-222 × Ghauri-3, Barani-333 × JSQ-White Hold, and Barani-333 × Ghauri-3 revealed higher Na+ contents and lower values for the K+ and K+/Na+ and performed poor for agronomic physiological and fiber quality traits. The excess Na+ accumulation can also reduce water uptake by the plant, as well as it also competes with other ions, such as potassium and calcium, for binding sites on the root surface and within the plant8. This can reduce the uptake of other vital nutrients and lead to nutrient deficiencies, which can further reduce yield of the seed cotton and boll weight8. In our study, negative association of Na+ and H2O2 with fiber quality traits and agronomic as well as antioxidant traits was observed under salt stress conditions. It was reported that tolerant plants accumulated a lower amount of Na+ ions, while sensitive plants accumulated a higher amount of Na+ ions within their cells41. Salt-tolerant plants maintain their Na+/K+ ratio by reducing the uptake of Na+ from the roots, sequestering the excess Na+ present in the cytosol into the vacuole, and promoting the efflux of Na+ from root cells41.
Antioxidants play a critical role in protecting cotton plants from oxidative stress under salt stress. Antioxidants such as catalase, peroxidase, superoxide dismutase and non-enzymatic antioxidants (carotenoids, ascorbate and flavonoids) can neutralize ROS by donating electrons to them, preventing the formation of more ROS and the damage they can cause4. In response to salt stress, plants activate protective enzymes such as SOD, POD, and CAT, to enhance their ability to eliminate ROS6,30. The current study observed an increase in the activities of SOD, POD, and CAT in leaves under salt stress. The increased SOD activity in leaves may aid in scavenging oxygen-free radicals, while the elevated CAT activity may facilitate the breakdown of H2O2 into water and oxygen, thus reducing its levels. This finding is consistent with previous reports6,8,10 who also noted an increase in SOD, POD, and CAT activities in cotton leaves under salinity stress. The POD activity was enhanced up to 53% in resistant cultivars37. The enhancement of POD activity contributes to the improvement of photosynthetic activity, thereby highlighting the important role of antioxidant defense mechanisms in mitigating salt stress37. The enhanced activity of antioxidants such as SOD, POD, and CAT were associated with salt tolerance during fiber development42. Under stressful conditions, carotenoids tend to increase as their primary function is to protect against singlet oxygen43,44,45. High-yielding cultivars often exhibit elevated levels of CAT, TSP, and POD as they play an active role in regulating H2O2 levels by scavenging it to maintain optimal levels. The genotypes FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, along with crosses FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333 revealed higher level of K+/Na+, K+, POD, SOD, CAT, TSP, Chla, Chlb, and CAR and declared as salt tolerant cultivars. The above-mentioned genotypes also showed relatively higher expression level of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 at 15 dS/m and proved the role of ERF genes in salt tolerance in cotton. Similar findings also reported in cotton46, maize47, quinoa48 and wheat49.
Conclusions
The genetic variability study of cotton germplasm for morphological, biochemical, and fiber quality traits under salt stress revealed that the genotypes the FBS-Falcon, Barani-333, JSQ-White Hold, Ghauri, along with crosses FBS-FALCON × JSQ-White Hold, FBG-222 × FBG-333, FBG-222 × Barani-222, and Barani-333 × FBG-333 performed optimally and exhibited salt tolerance based on their K+/Na+ ratio, K+ levels, and various enzyme activities such as POD, SOD, and CAT. Furthermore, these genotypes also exhibited higher levels of TSP, Chla, Chlb, and Car under salt stress, indicating their ability to survive under the effects of salt stress. These genotypes also showed relatively higher expression level of Ghi-ERF-2D.6 and Ghi-ERF-7A.6 at 15 dS/m. Importantly, these genotypes also demonstrated favorable fiber quality traits, making them potential candidates for the development of salt-tolerant cotton varieties with desirable fiber properties. Future studies focused on the molecular foundation of salt tolerance in these genotypes could facilitate the development of improved cotton varieties that can thrive in salt-affected regions and contribute to the sustainable production of cotton. Further research on the mechanisms underlying salt tolerance in cotton plants is warranted to develop a better grasp of the molecular foundation of salt tolerance and to facilitate the development of improved salt-tolerant cotton varieties.
Data availability
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Abbreviations
- PH:
-
Plant Height (cm)
- NBP:
-
Number of bolls/plant
- BW:
-
Boll weight (g)
- SCY:
-
Seed cotton yield (g)
- LP:
-
Lint (%)
- Na+ :
-
Sodium concentration
- K+ :
-
Potassium concentration
- K+/Na+ :
-
Potassium to sodium ratio
- H2O2 :
-
Hydrogen peroxide (μmol/g)
- CAT:
-
Catalase (U/mg protein)
- SOD:
-
Super-oxidase dismutase (U/mg protein)
- POD:
-
Peroxidase (U/mg protein)
- TSP:
-
Total soluble Protein (mg/g FW)
- Caro.:
-
Carotenoid (mg/g FW)
- Chl a&b:
-
Chlorophyll contents a & b (mg/g FW)
- FF:
-
Fiber fineness (µg/in)
- FL:
-
Fiber length (mm)
- FS:
-
Fiber strength (g/tex)
References
Suhani, I. et al. Restoration, construction, and conservation of degrading wetlands: A step toward sustainable management practices. In Restoration of Wetland Ecosystem: A Trajectory Towards a Sustainable Environment 1–16 (2020).
Mau, Y. & Porporato, A. A dynamical system approach to soil salinity and sodicity. Adv. Water Resour. 83, 68–76 (2015).
Zafar, M. M. et al. Genetic variation studies of ionic and within boll yield components in cotton (Gossypium hirsutum L.) Under salt stress. J. Nat. Fibers 19, 3063–3082 (2022).
Guo, H. et al. Growth, ionic homeostasis, and physiological responses of cotton under different salt and alkali stresses. Sci. Rep. 10, 21844 (2020).
Chaudhry U. F. et al. Genetic studies in different F2 segregating population for yield and fiber quality traits in cotton (Gossypium hirsutum L.), Int J Agri Biosci 11, 11 (2022).
Zafar, M. M. et al. Effects of salinity stress on some growth, physiological, and biochemical parameters in cotton (Gossypium hirsutum L.) germplasm. J. Nat. Fibers 19, 8854–8886 (2021).
Abdelraheem, A. et al. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crops Prod. 130, 118–129 (2019).
Farooq, M. A. et al. A study towards the development of salt tolerant upland cotton (Gossypium hirsutum L.). J. Nat. Fibers 19, 4115–4131 (2022).
Zafar, M. M. et al. Advancing heat tolerance in cotton through integration of multiple stress tolerance indices and multivariate analyses. Int. J. Agric. Biosci. 13, 65–75 (2024).
Farooq, M. A. et al. Roles of reactive oxygen species and mitochondria in seed germination. Front. Plant Sci. 12, 781734 (2021).
Munawar, W. et al. Differential morphophysiological and biochemical responses of cotton genotypes under various salinity stress levels during early growth stage. Front. Plant Sci. 12, 622309 (2021).
Hu, Y. & Schmidhalter, U. Opportunity and challenges of phenotyping plant salt tolerance. Trends Plant Sci. 28, 552–566 (2023).
Huang, H. et al. SLNL: A novel method for gene selection and phenotype classification. Int. J. Intell. Syst. 37, 6283–6304 (2022).
Zafar, M. M. et al. Genome-wide characterization and expression analysis of Erf gene family in cotton. BMC Plant Biol. 22, 134 (2022).
Long, L. et al. Transcriptome analysis reveals differentially expressed ERF transcription factors associated with salt response in cotton. Plant Sci. 281, 72–81 (2019).
Qiao, Z. X. et al. Molecular cloning and functional analysis of an ERF gene from cotton (Gossypium hirsutum). Biochim. Biophys. Acta Gene Regul. Mech. 1779, 122–127 (2008).
Zafar, M. M. et al. Biochemical and associated agronomic traits in Gossypium hirsutum L. under high temperature stress. Agron 12, 1310 (2022).
Maryum, Z. et al. An overview of salinity stress, mechanism of salinity tolerance and strategies for its management in cotton. Front. Plant Sci. 13, 907937 (2022).
Li, Y. et al. Deciphering the probiotic properties and safety assessment of a novel multi-stress-tolerant aromatic yeast Pichia kudriavzevii HJ2 from marine mangroves. Food Biosci. 56, 103248 (2023).
Richards, L. A. Diagnosis and improvement of saline and alkali soils (US Department of Agriculture, 1954).
Allen, R. et al. Effect of catalase inactivation on levels of inorganic peroxides, superoxide dismutase, glutathione, oxygen consumption and life span in adult houseflies (Musca domestica). Biochem. J. 216, 503–506 (1983).
Zhang, L. et al. Morphological and physiological responses of cotton (Gossypium hirsutum L.) plants to salinity. PLoS One 9, e112807 (2014).
Liu, D. et al. Uptake and accumulation and oxidative stress in garlic (Allium sativum L.) under lead phytotoxicity. Ecotoxicology 18(1), 134–143 (2009).
Bradford, M. M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254 (1976).
Arnon, D. I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 24, 1 (1949).
Steel, R. G. D. & Torrie, J. H. Principles and Procedures of Statistics, A Biometrical Approach (McGraw-Hill Kogakusha Ltd., 1980).
Falconer, D. S. Introduction to Quantitative Genetics (Pearson Education India, 1996).
Racine, J. S. RStudio: A platform-independent IDE for R and Sweave. In: JSTOR (2012).
Johnson, H. W. et al. Estimates of genetic and environmental variability in soybeans. Agron J. 47, 314–318 (1955).
Noman, M. U. & Azhar, S. Metabolomics, a potential way to improve abiotic stresses tolerance in cereal crops. Int. J. Agric. Biosci. 12, 47–55 (2023).
Wang, N. et al. Relative contribution of Na+/K+ homeostasis, photochemical efficiency and antioxidant defense system to differential salt tolerance in cotton (Gossypium hirsutum L.) cultivars. Plant Physiol. Biochem. 119, 121–131 (2017).
Zafar, M. M. et al. Unraveling heat tolerance in upland cotton (Gossypium hirsutum L.) using univariate and multivariate analysis. Front. Plant Sci. 12, 727835 (2021).
Kumar, C. P. S. et al. Studies on genetic variability, heritability and genetic advance in cotton (Gossypium hirsutum L.). Plant Arc 19, 934–937 (2019).
Manan, A. et al. Genetic analysis of biochemical, fiber yield and quality traits of upland cotton under high-temperature. Plant Prod. Sci. 25, 105–119 (2022).
Sahar, A. et al. Genetic variability for yield and fiber related traits in genetically modified cotton. J. Cotton Res. 4, 1–10 (2021).
Saleh, B. Effect of salt stress on growth and chlorophyll content of some cultivated cotton varieties grown in Syria. Commun. Soil Sci. Plant Anal. 43, 1976–1983 (2012).
Dong, Y. et al. Salt-tolerance diversity in diploid and polyploid cotton (Gossypium) species. Plant J. 101, 1135–1151 (2020).
Nadeem, M. et al. Expression analysis of fiber related genes in different staple length genotypes in cotton (G. hirsutum L.). Int. J. Agric. Biosci. 12, 180–187 (2023).
Nawaz, M. et al. Impact of salt stress on cotton. Int. J. Agric. Biosci. 12, 98–103 (2023).
Ibrahim, W. et al. Comparative physiological analysis in the tolerance to salinity and drought individual and combination in two cotton genotypes with contrasting salt tolerance. Physiol. Plant 165, 155–168 (2019).
Hamani, A. K. M. et al. Responses of leaf gas exchange attributes, photosynthetic pigments and antioxidant enzymes in NaCl-stressed cotton (Gossypium hirsutum L.) seedlings to exogenous glycine betaine and salicylic acid. BMC Plant Biol. 20, 1–14 (2020).
Ju, F. et al. Integrative transcriptomic, metabolomic and physiological analyses revealed the physiological and molecular mechanisms by which potassium regulates the salt tolerance of cotton (Gossypium hirsutum L.) roots. Ind. Crops Prod. 193, 116177 (2023).
Sharif, I. et al. Salinity stress in cotton: Effects, mechanism of tolerance and its management strategies. Physiol. Mol. Biol. Plants 25, 807–820 (2019).
Liaqat, K. et al. Assessment of tomato accessions for various seedling attributes under NaCl salt stress. Int. J. Agric. Biosci. 12, 116–121 (2023).
Zafar, M. M. et al. Drought and heat stress on cotton genotypes suggested agro-physiological and biochemical features for climate resilience. Front. Plant Sci. 14, 1265700 (2023).
Azeem, M. et al. Salinity stress improves antioxidant potential by modulating physio-biochemical responses in Moringa oleifera Lam. Sci. Rep. 13, 2895 (2023).
Zhang, J. et al. Comparative phenotypic and transcriptomic analysis reveals key responses of upland cotton to salinity stress during postgermination. Front. Plant Sci. 12, 639104 (2021).
Ashraf, M. A. et al. Phenological application of selenium differentially improves growth, oxidative defense and ion homeostasis in maize under salinity stress. Plant Physiol. Biochem. 123, 268–280 (2018).
Shabbir, A. et al. Biochar mitigates arsenic-induced human health risks and phytotoxicity in quinoa under saline conditions by modulating ionic and oxidative stress responses. Environ. Pollut. 287, 117348 (2021).
Acknowledgements
This research was supported by The Researchers Supporting Project number (RSPD2024R952), King Saud University, Riyadh, Saudi Arabia. The authors would like to thank the FB Genetics, Four Brothers Group, Lahore, Pakistan for providing space and facilities to conduct this experiment.
Funding
This research was supported by The Researchers Supporting Project number (RSPD2024R952), King Saud University, Riyadh, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
M.M.Z., A.R. and W.S.C. wrote the initial draft of the manuscript. A.A. provided the space and help to conduct the experiment. J.A., A.P., A.S., S.Z., A.B.S., K.M.E. and H.S. helped in data recording and statistical analyses. F.Q., M.M.Z. and A.S. review and edit the manuscript. The final approval was given by F.Q. and X.J.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zafar, M.M., Razzaq, A., Chattha, W.S. et al. Investigation of salt tolerance in cotton germplasm by analyzing agro-physiological traits and ERF genes expression. Sci Rep 14, 11809 (2024). https://doi.org/10.1038/s41598-024-60778-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-024-60778-0
- Springer Nature Limited