
Abstract
Plant-based mulch has been proposed as a sustainable way of maintaining soil fertility. However, the role of mulch diversity, quality, and size in decomposition dynamics, and their effect on crop yield, has not been fully explored. We investigated how mulch quality, proxied by the constituent plant species diversity, and residue size drive mulch decomposition, nutrient release, crop nutrition, and yield. A rhizotron experiment was set up with barley as a model crop, with the addition of mulch of two particle sizes (1.5 and 30 cm) and four different plant residue mixes of differing biodiversity (17, 12, 6, and 1 species) in a fully factorial design. Soil nutrient dynamics were measured at advanced decomposition stages, together with residue quality, arbuscular mycorrhizal fungal (AMF) root colonisation, and crop yield. Residue mass loss was significantly affected by its chemical composition. Initial NDF content was more restricted factor in C and N mineralisation than C:N or lignin. Long residues retained significantly higher C and N content, than short residues. Crop yield was not affected by residue type or size. Residue size significantly affected barley growth rate, influencing seed protein content. Soil available K was significantly increased by residues with a higher initial C:N ratio. Short residues resulted in higher soil Zn. Residues of higher diversity resulted inhigher AMF root colonisationof the barley plants. Generally, long residue mulches maintain higher fertilisation capacity at advanced stage of decomposition than short ones, without a deleterious effect on crop yield. Further investigation should evaluate the effect of continuous application of long residue mulches on soil fertility and microbial symbiosis.
Similar content being viewed by others


Introduction
Plant-based mulch can significantly affect the physical, chemical, and biological properties of agricultural soils1,2,3,4. Mulches can provide physical protection from soil erosion and enhance and maintain soil fertility, particularly in conservation tillage and organic farming systems5,6,7. Sustainability in agroecosystems is typically defined by the reliance on local resources and diversity conservation8. Therefore, mulch from cover crops and crop residues can contribute to sustainability by promoting soil fertility and the diversity of soil-dwelling organisms9. However, in contrast to mineral fertilisers, the release of nitrogen from organic sourcescan be unpredictable and much more susceptible to variations due to environmental conditions10. In particular, the effect of mulching on soil carbon (C) and nitrogen (N) as indicators of soil qualityis not well understood11. Generally, the greater the mass of residues, the higher the soil C content, yet the mineralisation of N is not so straightforward and is affected by many soil conditions12.
The timing of nutrient release in relation to crop demand during the growing season is an important consideration when using mulch. Su et al.4 found that plants had increased nitrogen uptake and growth during the early stages of development after mulching. However, the benefits diminished over time, possibly due to increased ammonia volatilisation. It's important to note that soil nitrogen availability does not always increase immediately after mulch application, particularly during the vegetative stages of the crop when soil nitrate content is low13. Additionally, rapid decomposition of mulch may not always be beneficial as it can result in nutrient release too early in the growing season, leading to limited nutrient supply in the later stages of crop development9.
Concentrations of available nutrients can also depend on the number of applications of plant mulch. Pavlu et al.14 observed increasing concentrations of available P and K in ascending order from one to three annual applications. P concentrations are typically higher in mulched topsoil due to rapid fixation by soil particles and to immobilisation by microorganisms15. These factors above can lead to nutritional benefits in crops yet not necessarily reflect in yield. For example, higher seed protein content in soybean was shown with mulch application (compared to no mulch), yet there were no measurable effects on yield13.
The importance of residue diversity in stimulating soil ecosystem services is not well understood, particularly the types of plant traits and quantity of species required16,17. When plant residues come from a mixture of plant species, the decomposition process can be faster (synergistic effect), slower (antagonistic effect), or proportional to constituents (additive), depending on the individual plants involved18. Mixtures of different plant species may increase plant residue-derived C assimilation by soil microbes in the early stages of decomposition, compared to residues of single plant species, due to functional complementarity which reduces competition between microbial communities19. Different groups of microbes use different enzymes to decompose substrates of varying chemical composition20, so residues with high plant diversity can promote microbial21 and arbuscular mycorrhizal fungal (AMF) diversity22.
The decomposition of plant residues can be affected by the contact between soil particles and plant material and the size of plant residue per unit mass. Shredding mulch residues to small particle sizes breaks up the continuity of recalcitrant plant tissues, potentially aiding decomposition by increasing accessibility and surface area available to soil microbes23. Residue size interacts with the C:N ratio and the availability of N in the soil. For example, the decomposition of short vs long residues may not be accelerated, despite higher contact with soil, when residues are rich in N24. The particle size of straw residues (high C:N ratio) did not play an essential role in N immobilisation and soil microbial biomass in field conditions with low soil available inorganic N25. The immobilisation, mineralisation, and denitrification of N were higher in shredded residues with high C:N ratio in N rich soil, but only in the short term23,24. This suggests that residue particle size does not affect the final amount of nitrogen that is released in the soil but only the timing of the release and availability to plants.
Consequently, this study aims to improve our understanding of the optimal size of mulch fragments (whole plant versus shredded) and the importance of plant residue (bio)diversity. We hypothesise that (i) long residues maintain higher fertilisation capacity than short residues at the end of the growing season, (ii) soil nutrient availability at the later stages of decomposition are affected by both residue size and residue quality (residue C:N ratio, N, and recalcitrant substances), (iii) AMF root colonisation increases with increasing residue species richness, (iv) and crop quality is affected by both residue size and residue quality but with no deleterious effect on crop yield. Results are discussed in the context of soil health and fertility, with the view of optimising long-term mulch use.
Methodology
Rhizotron setup
The experiment was conducted at the Crop and Environment Laboratory of the University of Reading in the UK, between June and November 2018. Thirty-six minirhizotrons were constructed using 0.5 cm thick PVC sheeting, with each representing an independent replicate (as shown in Fig. 1). One side of each rhizotron was made of clear acrylic to allow for observation of root growth and was removable to facilitate soil sampling. The clear acrylic was covered with thermawrap silver foil between observations and sampling to prevent light penetration and reduce temperature fluctuations. Each rhizotron was 1 m tall to enable root system expansion at depth and was 30 × 5 cm wide, providing a soil volume of 0.015 m3. A layer of gravel approximately 1.5 cm thick was placed at the bottom of each rhizotron to facilitate drainage, and the remainder was filled with approximately 20 kg of commercially supplied, well-homogenised loamy sand topsoil (bulk soil) sieved to 8 mm and with a pH of 7.3 ± 0.032 SD. The rhizotrons were positioned at a 70-degree angle throughout the experiment, with the transparent side facing downward to encourage root growth along the transparent side for better observation. They were initially placed outside and then moved into a greenhouse in October to aid plant senescence.

Rhizotron and soil sampling design. Large circles indicate root extraction samples, except for the 10–20 cm depth which was used to measure soil solution pH and bulk density. The large circles at 0–10 cm depth were used to detect AMF colonisation in barley roots. Yellow highlighted samples were taken on day 70, blue highlighted on day 137 after mulch application.
Experimental design
Spring barley (Hordeum vulgare L., var. Laureate) seeds (56 mg to 66 mg weight) were sown on 11th June 2018 into a germination tray. Plant emergence occurred after 15 days, vigorous plants between 15 and 22 cm height were then transplanted into rhizotrons 17 days later. Two plants per rhizotron were planted 15 cm apart and 7.5 cm from each edge (Fig. S1). An automated irrigation system with four drippers per rhizotron was installed to maintain moisture at 55–60% Water Holding Capacity (WHC). Irrigation stopped 106 days after plant emergence to promote seed maturity. Growth stages of the barley plant were observed every 1 to 2 weeks, and the plants were harvested 121 days after mulch application.
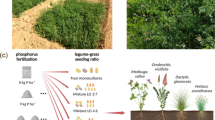
Eight biomass residue treatments were established in a fully factorial design (n = 4), comprising four plant diversity mixtures (types) and two plant residue particle sizes (short of 1.5 cm (S), and long of 30 cm (L) so long as the length of the soil surface in a rhizotron). A control (C)with no mulch application was also established. The residue types were: Perennial ryegrass (P) (1 plant species), Smart Grass™ (S) (6 species), Biomix™ (B) (12 species), and Herbal™ (H) (17 species) (Table S1).No threatened species were used in this research, following IUCN policy statement on research involving species at risk of extinction. Experimental research, including the collection of plant material, was conducted on cultivated plants as cover crop plants, in compliance with relevant institutional, national, and international guidelines and legislation. Plant material was sourced from the University of Reading farm, specifically the DiverseForages grassland field sites. These residue types weresuitable to address our hypothesis because they included residue mixtures with different characteristics. Residue characteristics include both residue diversity and functional traits. Residue diversity may concern either the species composition or the species richness which is the number of species participating in a residue mixture. The functional traits of residues may concern either the chemical composition, which determines the residue quality, or the morphological features of residues26,27. Fresh plant residue was collected from field plots planted with the aforementioned forage mixtures on 29th June 2018, cut to specified sizes, and placed on the soil surface in rhizotrons randomly allocated to each treatment 4 days later. Plant mass representative of 23 g dry mass was used in each treatment. Thus, the nine treatments were: HL, HS, BL, BS, SL, SS, PL, PS, and C. No additional fertilisation or pesticide treatment was applied to the rhizotrons during the experiment. The encoding P(1), S(6), B(12), and H(17) as well as PL(1), PS(1), SL(6), SS(6), BL(12), BS(12), HL(17), and HS(17) has been adopted where it was considered necessary to indicate the number of species participating in the different residue types or treatments, respectively.
Soil and plant material sampling and analyses
Soil samples were collected from the rhizotrons on two occasions: 70 and 137 days after mulch application (Fig. 1). In total, 21 soil samples were taken from each rhizotron (4 in the first sampling period and 17 in the second), resulting in a total of 756 samples from all rhizotrons (Fig. S2). Soil samples for root extraction and measurement of soil pH and bulk density were collected on day 137 only to avoid early soil disturbance. In addition, soil samples from a depth of 70–75 cm were collected on day 137 only to detect any differences in soil available nitrogen at that depth, which would indicate that the decomposition process was still ongoing at the end of the growing season. All samples, except those containing roots, were air-dried, sieved to < 2 mm, and stored at room temperature. Samples containing roots were stored at 4 °C until root extraction. Roots were extracted by submerging soil samples in tap water over a 1 mm sieve and collecting all floating roots..
Soil samples for Mehlich 3 extraction to detect available nutrients other than N were received from 0–5 to 20–25 cm depth, while samples to detect available N were received from relatively higher depths (5–10, 50–55, and 70–75 cm) because N is more prone to leaching. Root samples were collected from the depths of 0–10, 25–35, and 55–65 cm to include roots from a wide range of the soil profile, while the 10–20 cm depth was used for samples to measure soil solution pH and soil bulk density at the nearest to the soil surface available depth, considering the collection of all other samples.
After the final harvest, mulch residue, aboveground and belowground barley plant biomass, and soil samples were analysed to establish key physical and chemical properties of investigated plant-soil systems. Mulch residue and barley biomass were dried for 48 to 72 h at 80 °C until constant weight and then at 105 °C for 24 h to establish dry weight. Total carbon, nitrogen, and protein content of mulch, plant tissue, and seeds and total carbon and nitrogen of soil sub-samples were determined through combustion (LECO CHN 628 analyser, LECO Corporation)28. An ANKOM 200 Fibre Analyser (ANKOM Technology)29 was used to measure percentage content of neutral detergent fibre (NDF) (Hemicellulose, Cellulose, and Lignin), acid detergent fibre (ADF) (Cellulose and Lignin), and acid detergent lignin (ADL) (Lignin) of mulch residues according to ANKOM Technology protocols. All three fractions NDF, ADF, or ADL potentially can be used as a measure of residue recalcitrance. We then estimated the % Cellulose by subtracting the % ADL from % ADF, and the % Hemicellulose by subtracting the % ADF from the% NDF. Samples were dried at 80 °C to constant weight and ground in a Fritsch grinder (Glen Creston Ltd) with a 1 mm sieve. Arbuscular mycorrhizal fungi (AMF) colonisation and abundance was estimated by black ink staining according to Vierheilig et al.30.
Air-dried 10 g soil samples sieved to < 2 mm were suspended in centrifuge tubes in 25 mL of ultra-pure water and shaken for 15 min on an end-over-end shaker to measuresoil solution pH31. Available N was estimated by KCl extraction method according to Great Britain M.A.F.F.32 standard protocol using 40 g of air-dry soil in 200 ml 1 M potassium chloride solution, measured colourimetrically by a San Continuous Flow Injection Analyzer (SCALAR Instruments)33. The Mehlich 3 method34 was used to evaluate the availability of P, K, Mg, Mn, Fe, Cu and Zn nutrients by Perkin Elmer-Optima 7300 DV ICP-OES analyser(PerkinElmer, Inc.)35,36,37. Air dry soil samples of 2 g and 20 ml of Mehlich 3 extracting solution were used. Other soil samples were air-dried and sieved to < 2 mm to establish soil texture, which was measured with a hydrometerby adding 50 ml sodium hexametaphosphate solution (SHMP) at 50 g/l to 40 g soil subsamples38. Soil water holding capacity (WHC) was estimated gravimetrically using 50 g fresh soil samples with the method described by Harding and Ross39. Three samples were taken from the middle of three rhizotrons in random from the depth of 12–16 cm to measure average bulk density40. The height of the main stem of barley plants from the soil surface to the base of the flag leaf was used as a proxy of growth rate (cm/d).
Statistical analysis
All statistical analyses were carried out in Minitab 19 (Minitab, LLC)41 except Principal Component Analysis which was conducted in R-Studio (RStudio, PBC)42. All measurements from a rhizotron were averaged to obtain a single mean value, the unit of replication of this study being the rhizotron (n = 4), and ɑ < 0.05 was used to denote significance. One-way ANOVA and a General Linear Model were used to detect differences between treatments for one factor (treatments, residue type, or residue size) and two factors (type and size of residues), respectively. Measurements repeated in time (day 70, and day 137) or position (middle and right side of rhizotrons) or soil depth were analysed by Mixed Effects Model with rhizotrons as a random factor and type of residues, size of residues, time, depth, and position as fixed factors. All data subjected to analyses of variance were tested for normality and homogeneity, using Darling-Anderson and Levene tests, respectively. When these conditions were not satisfied, data were transformed by log10 or Box-Cox transformation with optimal or rounded λ. Kruskal–Wallis test was conducted instead of one-way ANOVA if transformations did not normalise data variance sufficiently. When a significant treatment effect was observed, a Tukey post-hoc test was also conducted. Comparisons with the control treatment were made using the Dunnett test. Data were separated and analysed with ANOVA for a specific sampling time or depth, when necessary, where normality or equality of variances were not satisfied even after data transformation. Descriptive statistics included means and standard deviations rather than standard errors unless it was otherwise stated.
Simple regression analysis was conducted in order to examine the significance and the degree of the effect of a single independent variable to a response variable in cases where there was an apparent influence (residue dry mass loss vs initial residue NDF or N content or initial residue C:N ratio, soil K vs initial residue C:N ratio, AMF root colonisation vs residue species richness, seed protein vs main stem elongation rate as continuous predictor variable and residue size as categorical predictor variable). Homogeneity of variance of the data was tested with Levene test, and normality of data distribution with Anderson–Darling test. The Spearman correlation was used when normality or equality of variances of data were not satisfied. Pearson correlation was used to assess the significant correlations between variables.
Multivariate analysis was also conducted on data collected from barley plants and soils using Principal Component Analyses (PCAs) to assess differences between treatments in several variables simultaneously, concerning measurements on barley plants or soil nutrients, and the relations of the treatments with those variables.
Results
Residue dry mass loss, and initial and final quality
Residue (mulch) dry mass loss due to decomposition was significantly different between treatments (N = 4, F = 2.57, p-value = 0.040) and specifically between BS(12) (71.71 ± 7.21) and PL(1) (47.78 ± 6.44) (T-value = −3.40, p-value = 0.041, Fig. 2).

Final residue dry mass loss (121 days after mulch application). Mean values are depicted with x (N = 4, F = 2.57, p-value = 0.040). The residue types were: Perennial ryegrass (P) (1 plant species), Smart Grass (S) (6 species), Biomix (B) (12 species), and Herbal (H) (17 species). Residue treatments including fourplant diversity mixtures (types) and two plant residue fibre sizes (short of 1.5 cm (S), and long of 30 cm (L)). Treatments that do not share a common letter are significantly different (p < 0.05).
Further analysis showed a significant effect of residue diversity (residue types) on residue dry mass loss (N = 8, F = 5.40, p-value = 0.006). There were significant differences between P(1) (50.26 ± 13.33) and B(12) (70.02 ± 5.39) (T-value = -3.98, adjusted p-value = 0.003), but no significant between long and short residue size (N = 16, F = 0.05, p-value = 0.823).
The P(1) residue type had the lowest initial C:N ratio (24.2 ± 2.268) while S(6) had the highest (33.5 ± 1.65), B(12) had 30.4 ± 3.64 and H(17) 28.1 ± 1.801 (N = 8, F = 20.24, p-value < 0.001, Tables 1, S2 and S3). Type P(1) had the highest mean value (41.644 ± 0.160) of initial C content of residues, significantly different from B(12) (41.229 ± 0.256), and H(17) (41.207 ± 0.195) (N = 8, p-value = 0.001, Tables S2, and S3). S(6) type had the lowest initial Ν content (1.238 ± 0.055) while P(1) had the highest (1.734 ± 0.170), and there were significant differences between types (N = 8, F = 19.55, p-value < 0.001, Tables S2 and S3). The same pattern was observed in the initial protein content of residues (N = 8, F = 19.55, p-value < 0.001). There were no significant differences in initial ADL (lignin) content between residue types (N = 4, Kruskal–Wallis test p-value = 0.277, Fig. S3, Table S4). There was a significant positive correlation between NDF and ADF (r = 0.938, p-value < 0.001) and no significant correlation between NDF and lignin (r = −0.021, p-value = 0.937) or between ADF and lignin (r = −0.058, p-value = 0.832). Moreover, differences between residue types in ADF were statistically more significant (N = 4, F = 5.76, p-value = 0.011) than in cellulose alone (Kruskal Wallis test, N = 4, H-value = 6.60, p-value = 0.086), while it was of equal significance in both NDF (N = 4, F = 16.14, p-value < 0.001) and in hemicellulose alone (N = 4, F = 23.04, p-value = < 0.001) (Fig. S3, Tables S4 and S5).
Residues harvested at the end of the experiment showed significant differences in C:N ratio between treatments (N = 4, F = 4.27, p-value = 0.003). PL(1) treatment which had the highest mean value was significantly higher than HS(17) and BS(12) (Fig. 3a, TablesS6 and S7). Further analysis showed significant differences in final C:N ratio between types (N = 8, F = 7.57, p-value = 0.001) as well as between short (15.029 ± 2.099) and long (16.744 ± 2.595) residues (N = 16, F = 6.66, p-value = 0.016). The C:N ratio was significantly higher in P(1) (17.959 ± 2.407) than in B(12) type (13.883 ± 1.797) (T-value = 4.33, adjusted p-value = 0.001), in P(1) than in H(17) type (14.922 ± 2.000) (T-value = 3.23, adjusted p-value = 0.018), and in S(6) (16.786 ± 1.655) than in B(12) type (T-value = 3.09, adjusted p-value = 0.024). Final C content of residues was also significantly higher (59.20%) in long residues (30.95 ± 4.68) than short residues (19.44 ± 5.89) (N = 16, F = 36.74, p-value < 0.001). PL(1), SL(6), and HL(17) were significantly higher than BS(12), HS(17), and PS(1) (N = 4, F = 6.02, p-value < 0.001, Fig. 3b, Tables S6 and S7). There were significantly higher final N contents in HL(17), BL(12), SL(6) treatments than in HS(17), and PS(12) (N = 4, F = 5.68, p-value = 0.001, Fig. 3c,Tables S6 and S7). In addition, final N content of residues was significantly higher (44.06%) in long residues (1.854 ± 0.154) than short ones (1.287 ± 0.303) (N = 16, F = 38.05, p-value < 0.001).

Box and whiskers plots of plant residue initial and final (a) C:N, (b) % C, and (c) % N ratio for the different types (B, H, P, and S) (initial residues) and Treatments (BL, BS, HL, HS, PL, PS, SL, and SS) (final residues). The residue types were: Perennial ryegrass (P) (1 plant species), Smart Grass (S) (6 species), Biomix (B) (12 species), and Herbal (H) (17 species). Residue treatments included four plant diversity mixtures (types) and two plant residue fibre sizes (short of 1.5 cm (S), and long of 30 cm (L)). Lower case letters refer to comparisons between initial residue types. Upper case letters refer to comparisons between final residue treatments. Mean values that do not share a common letter are significantly different (p < 0.05).
Simple linear regression analysis was conducted using the residue dry mass loss as response variable (y) and either the initial NDF content or the initial N content or the initial C:N ratio of residues (x) as predictor variable (N = 8). The analysis showed that the regression model could explain only r2 = 21.12% or 24.50% or 21.36%, respectively of the variation of the response variable. However, the influence of the predictor was statistically significant in all cases (p-value = 0.008 for NDF, 0.004 for N, and 0.008 for C:N ratio). The regression equations were y = 113.1 − 1.190 × for NDF, y = 106.2 − 30.99 × for N, and y = 16.2 + 1.548 × for C:N ratio.
Soil nutrient content
Soil samples taken at harvest time on day 137 from 10 to 20 cm depth from the middle of the rhizotrons showed no significant differences in soil solution pH between treatments.
Soil NH4+ content was highly significantly lower on day 70 (1.302 ± 0.356) than day 137 (1.678 ± 0.379) (N = 64, F = 46.65, p-value < 0.001) and significantly higher at the top 5–10 cm (1.689 ± 0.280) than at 50–55 cm (1.292 ± 0.428) depth (N = 64, F = 52.13, p-value < 0.001, Tables S8, S9, and Fig. S4).
The effect of residue diversity and of residue size on soil NO3- content was not significant on day 70 at 50–55 cm depth. However, mean value of all treatments (1.407 ± 0.852) was significantly lower than that of bulk soil (initial soil, prior to its use in rhizotrons) (11.334 ± 1.533, N = 4, F = 40.80, p-value < 0.001).
Depth significantly affected all soil nutrient contents (P, K, Mg, Fe, Mn, Zn, and Cu), and time significantly affected all nutrients except Fe and Mn (Tables S10 and S11). Macronutrients P and Mg had higher mean values at 20–25 cm depth than at 0–5 cm, but the opposite was true for K. Macronutrient concentrations were higher on day 70 than on day 137 at both 0–5 and 20–25 cm depths. Further analysis showed significant differencesin soil K content between diverse residue types on day 137 at 0–5 cm depth (N = 8, F = 4.16, p-value = 0.017, Fig. 4). Tukey’s post-hoc testing showed significant differences between S(6) (mean = 102.60 ± 31.70) and P(1) (68.66 ± 13.11) (T-value = 2.92, p-value = 0.035), and between S(6) and B(12) (68.44 ± 24.12) (T-value = 3.17, p-value = 0.020) types. Spearman’s correlation confirmed there was a positive and statistically significant association between the soil K content and the initial C:N ratio of residues (Spearman ρ = 0.439, p = 0.007).

The effect of different types (S, B, H, P in descending order of their initial C:N ratio) of residues on soil K concentration (Mehlich 3 extraction), in comparison to unamended Control (C), 137 days after mulch application. Results include all types, and depth of 0–5 cm (N = 8). The residue types were: Perennial ryegrass (P) (1 plant species), Smart Grass (S) (6 species), Biomix (B) (12 species), and Herbal (H) (17 species). Means and bars of one standard error from the mean are depicted. Types that do not share a common letter are significantly different (p-value < 0.05).
All micronutrients (Fe, Mn, Zn, and Cu) had significantly higher mean values (p-values < 0.001) at 0–5 cm depth than at 20–25 cm. Further analysis revealed significantly lower soil Zn content (−38.30%) in long residues (21.73 ± 8.77) than short (35.22 ± 15.69) on day 137 at 0–5 cm depth (N = 16, F = 7.53, p-value = 0.011).
The influence of the different treatments on soil nutrients (P, K, Mg, Fe, Mn, Zn, and Cu) on a multivariate basis, at the depths of 0–5 cm and 20–25 cm, was tested in Principal Component Analyses on day 70 (Fig. S5), and on day 137 after mulch application (Fig. S6). Although treatments were overlapping in most cases, Control seemed to demonstrate the most negative relation with soil nutrients compared to the other treatments, most clearly at 0–5 cm depth on day 70 (Fig. S5a). Also, the contribution of S(6) type in the enrichment of soil K compared to the other treatments was obvious at 0–5 cm depth on day 137 (Fig. S6a).
Cultivation of barley resulted in statistically significant reduction in soil K (N = 4, Kruskal–Wallis p-value < 0.001) and Mn (N = 4, F = 2.50, p-value = 0.011) in comparison to the bulk soil.
Samples taken from the area of maximum plant root interaction,the middle of the rhizotrons, at 0-5 and 20–25 cm depth (day 137), showed significant differences in soil nutrient contents in comparison to samples from the poorly rooted (right) side of rhizotrons (N = 64, F = 16.27, p-value < 0.001 for soil K, F = 56.91, p-value < 0.001 for P, F = 6.60, p-value = 0.012 for soil Mn, F = 14.52, p-value < 0.001 for soil Zn, and F = 16.51, p-value < 0.001 for soil Cu).
Barley plants
Residue size significantly affected the length of ears (N = 32, F = 4.96 and p-value = 0.030). Treatments with short size residues had higher mean values (8.337 ± 1.453) than those with long size (7.778 ± 0.052). Total yield was not significantly affected by the different treatments. Likewise, barley seed protein content was not statistically significantly different between treatments of long and short residues (N = 4, F = 1.90, p-value = 0.182) or between treatments of different diversity types. However, in all types except P(1) all treatments with long size residues had higher values of protein content than treatments with short size residues, and Control had lower value than any treatment with long residues (Table S12). Spearman correlation showed a significant negative correlation (Spearman ρ = −0.529, p-value = 0.001) between length of ear and seed protein content (%).
Barley plant growth rate
All treatments followed the same pattern of stem elongation (Fig. 5). Main stem elongation rate (cm/d) was significantly higher in long residues (0.871 ± 0.230) than short residues (0.749 ± 0.257) 25 days after planting (42 days after plant emergence) (N = 16, F = 4.00, p-value = 0.050). However, 99 days after planting (116 days after plant emergence) main stem elongation in long residue treatments were significantly lower (0.509 ± 0.061) than in short ones (0.540 ± 0.0521) (N = 16, F = 4.98, p-value = 0.003). In contrast, residue type did not affect significantly main stem elongation rate.

Mean rate of main stem elongation (cm/day) of barley plants from soil surface to the base of the flag leaf for the different treatments (BL, BS, C, HL, HS, PL, PS, SL, SS, and C), 11, 25, 37, 44, 51, and 99 days after planting (28, 42, 54, 61, 68, and 116 days after plant emergence). GS = growing stage58. The residue types were: Perennial ryegrass (P) (1 plant species), Smart Grass (S) (6 species), Biomix (B) (12 species), and Herbal (H) (17 species). Residue treatments including four plant diversity mixtures (types) and two plant residue fibre sizes (short of 1.5 cm (S), and long of 30 cm (L)). Control treatment (C) was with no residues.
Linear regression analysis of seed protein content (%) as the response variable, main stem elongation rate (MSER) as the continuous predictor variable, and residue size (long, short, and Control) as categorical predictor variable on day 25 after planting was conducted (Fig. S8). The analysis showed that main stem elongation rate could explain only r2 = 15.13% of the final seed protein content, but the influence was significant (N = 8, F = 4.62, p-value = 0.040).
The influence of the different treatments on several variables concerning barley plant biomass (variables v1 and v2), the relation of barley plants with symbiotic microbes (variable v3), and seed quality and yield (variables v4 to v6) on a multivariate basis, was tested in a Principal Component Analysis (Fig. 6). From this it is evident, HL(17) treatment had the strongest positive relationship with AMF colonisation (v3 variable) and the strongest negative with total seed dry mass (v4), opposite Control treatment. In the rest of variables differences between treatments were not clear confirming the lack of significant differences as treatments were highly overlapped.

PCA ordination of variables v1 = Barley root dry mass (g) (from combined samples at 25-35 cm and 55–65 cm depth), v2 = barley plant dry mass (g) per plant per rhizotron (without ears and roots), v3 = % AMF colonisation in barley roots, v4 = total seed dry mass (g) per plant per rhizotron, v5 = % barley seed protein content, v6 = % barley seed carbon content, for the treatments HL, HS, BL, BS, SL, SS, PL, PS, and Control (C) concerning measurements on barley plants. The residue types were: Perennial ryegrass (P) (1 plant species), Smart Grass (S) (6 species), Biomix (B) (12 species), and Herbal (H) (17 species). Residue treatments including fourplant diversity mixtures (types) and two plant residue fibre sizes (short of 1.5 cm (S), and long of 30 cm (L)). Control treatment (C) was with no residues. Large points depict mean values, while ellipses depict confidence intervals (ɑ = 0.05) of mean values for each treatment. The percentages represent data variation explained by the two first Principal Components (Dim1 and Dim2), bottom axis represents Dim1 normalised score, and left axis represents Dim2 normalised score.
AMF colonisation
The Arbuscular Mycorrhizal Fungi (AMF) root colonisation was assessed on day 137 and showed no significant differences due totreatments (HL, HS, BL, BS, SL, SS, PL, PS, C) or residue size. However, mean values indicated that residues of higher species richness had higher AMF colonisation. Control had the lowest mean value (Table 2).
Linear regression analysis showed that residue species richness could explain only r2 = 15% of the AMF root colonisation variation. However, the influence of the predictor was statistically significant (N = 8, p-value = 0.02). The regression equation was y = 30.60 + 0.365x, which means that for every plant species that is added in the residue mixture an increase of 0.365% in AMF root colonisation was expected (Fig. S7).
Discussion
Residue quality and degree of decomposition
The residue types' initial C:N ratios were allsignificantly different, ranging between 15 and 33. Soil decomposer microbes have a C:N ratio of around 8 in their body and assimilate about 1/3 of the decomposed C. Therefore, an initial residue C:N ratio ≤ 24 is considered to drive to net N mineralisation even at later stages of decomposition, contrary to residues with C:N ratio > 2443. However, this threshold may rise to more than 30 by the activity of soil fauna44,45. C:N ratio affects organic matter decomposition in N-poor soils, while its effect in N-rich soils can be insignificant46. Apart from C:N ratio, decomposition is also greatly affected by residue recalcitrance47. Interestingly, P(1) type had the lowest C:N ratio but the highest recalcitrance, not significantly different from that of S(6) type despite S(6) having the highest C:N ratio (Table 1).
In all cases, treatments with long residues resulted in higher final C:N ratio, C, and N content than those of the same type with short residues at the end of the growing season. Higher final C:N ratio in residues of the same type denotes lower decomposition rate because C:N ratio decreases during decomposition after an initial short increase48. Considering the dry mass loss of residues was not significantly affected by their size, this is clear evidence that long residues contributed to a higher accumulation of C and N on soil surface, in the form of C and N content retained in residues, at the end of the growing season than the short residues. This suggests possible positive effects on the mineralisation of C and N, and possibly of other nutrients, for subsequent crops with continuous application of long versus short residue mulches (higher fertilisation capacity).
Moreover, the final C:N ratio was strongly affected by residue quality. As decomposition was progressing the C:N ratios of the different residue types were decreasing with a tendency for the values to converge49. However, the lignin and other recalcitrant fractions of residues that were less affected during decomposition50 slowed down the decrease of the C:N ratios of the residues. Indeed, final C:N ratio was higher at P(1) and S(6) types and especially at PL(1) and SL(6) treatments (long residues of higher recalcitrance) than at B(12) and H(17) and especially at BS(12) and HS(17) (short residues of lower recalcitrance). NDF content was the best measure of residue recalcitrance as opposed to ADF or lignin alone since there were no significant differences in initial lignin content between the different residues. In addition, NDF contains all recalcitrant fractions (hemicellulose, cellulose, and lignin) and it was significantly correlated with ADF but not with lignin. Jensen et al.51 found that hemicellulose and cellulose were the factors that explained the greatest variability of C mineralisation rather than lignin. Therefore,C and N mineralisation of residues were affected mainly by residue size and initial NDF content rather than by initial C:N ratio and lignin content. It is possible that lower final N content in short residues was due to slightly increased N availability from more recalcitrant fractions caused by chopping and subsequent N uptake by plants or N losses by leaching. Chopped material is decomposed faster during the early days or weeks of decomposition because of larger exposure of the residue surface to direct contact with soil and soil microbes50. Assuming that the short residues are in better contact with the soil, the decomposition of short residuesare more likely to be dominated by bacteria and those of long residues by hyphae-producing fungi3,52. Bacteria preferentially decompose the readily available fractions leaving the more recalcitrant lignin fraction, while fungi decompose the more recalcitrant fractions53. Therefore, the final residue quality was affected by both residue size and residue quality.
Dry mass loss was not significantly influenced by residue size in the long term, consistent with previous studies24,54. However, it appears to be related to both residue recalcitrance and the C:N ratio or N content. This was confirmed by a simple linear regression analysis, where the effects of NDF, N, and C:N ratio on residue mass loss were statistically significant, although each variable could only explain a small part of the variation in dry mass loss (between 21.12 and 24.50%). Decomposition of lignin in residues with high recalcitrance and high N content is limited because the relevant decomposer microbes require a readily available carbon source, and excessive N makes these microbes more vulnerable to competition with other microbes55. Therefore, the higher the initial NDF content of residues, the lower the dry mass loss (primarily), and the higher the initial N content in residues of higher recalcitrance, the lower the final dry mass loss. Xu et al.56 also observed significant differences in biomass loss in a one-year experiment, with higher loss in species with a higher C:N ratio but low recalcitrance. The microorganisms that are adapted to the decomposition of recalcitrant substances are favoured by residues with a higher C:N ratio, as they can outcompete other microbes24. Alternatively, a higher diversity of residues may lead to higher specificity in enzymatic activity during decomposition, driving to higher residue mass loss. Higher microbial activity in residue mulch of higher diversity (mixed than single plant species) has also been reported in previous research but without being accompanied by higher microbial biomass or microbial community composition, attributed this effect to the activation of dormant microbial populations by residue mixtures56,57.
The fact that long residues resulted in higher final C and N content than short residues, while at the same time residue mass loss was not significantly affected by residue size, implies that long residues maintained higher fertilisation capacity than short residues at the end of the growing season, in line with our hypothesis.
Dynamics of soil nutrients
At and after barley growing stage GS6158, the N uptake requirement of barley plants declines as plant growth has almost been completed, and plants redistribute N to feed the developing grains59. The decomposition of the residues initially affected the availability of NH4+ and soluble organic N (SON).NH4+ is typically not prone to leaching60 but may be subjected to nitrification61, and the resulting NO2− and NO3− may leach downwards, a process detected in our samples from the 50–55 cm depth on day 70. Subsequently, the nitrification activity was restricted considerably on day 137, probably due to the cessation of irrigation to promote seed maturity62. Both short and long residues led to significantly higher values of soil NH4+on day 137 than on day 70 (Tables S8, S9, and Fig S4). The observed lower values of soil NH4+ on day 70 may be attributed to plant uptake because maximum nutrient uptake is up to GS70 (end of the flowering stage)63. Moreover, mean values of short residues were higher than those of long residues on day 70, and the opposite was true on day 137, although differences were not significant (Table S8). This shows that the rate of N mineralisation was initially higher in short residues, but it was reversed in favour of long residues sometimes up to day 137. Angers and Recous23 also observed higher decomposition rates at small particle size residues incorporated into the soil, followed by higher rates at long size (up to 10 cm) one. In contrast, experiments with fine and ground (usually < 1 cm) particle size residues mixed with soil showed effect of particle size on N dynamics only in early stage of decomposition24,54,64.
Mean values of soil NO3- were not significantly affected by the treatments, although they were far lower than the initial measurements in bulk soil. In addition, no significant effect of residue diversity or functional traits on soil NH4+ content was observed on both day 70 and day 137. Contrary to our experiment, N mineralisation was reported to be affected by residue chemical composition after 100 days of incubation, where high positive correlations had been found between N mineralisation and N to lignin content ratio of residues65. Any such effect in our experiment possibly occurred at earlier stages of decomposition and differences were soon compensated for, or it was negligible. Notably, Fox et al.66 concluded that when incorporating residues into soil, (lignin + polyphenol):N ratio could be used as a reliable predictor of N mineralisation rate of residues, but they did not find significant correlation between N mineralisation and residue N or lignin or polyphenol content alone. Our experimental data, supported by these previous observations, proffers a way forward where residue length may be used to time N release with crop demand.
Soil K was higher in all treatments with residues than in Control, in concordance with previous research67. The return of plant residues to the soil is a considerable source of soil K replenishment. K is readily released as K+ ions to soil solution during decomposition because it remains in plants in ionic form in the cell solution and contributes, like Mg, to the production of extracellular enzymes68. We hypothesised that the residue chemical composition (quality) and residue size would affect nutrient dynamics. Our data showed that K was the only nutrient significantly affected by residue chemistry. Soil K was higher in residues with higher C:N ratio, and this positive correlation was statistically significant as confirmed by Spearman’s correlation. Probably, the fact that K largely remains in the plant cells in ionic form makes it less prone to be bound in residue recalcitrant substances and therefore its release to soil solution during decomposition is dependent on initial residue C:N rather than NDF content. It is possible, residues of higher quality released higher amounts of K earlier due to higher initial rates of decomposition, while the opposite was true with lower-quality residues69.
Soil micronutrients Fe, Mn, Zn, and Cu had significantly higher mean values at 0–5 cm depth than at 20–25 cm. In the field their concentrations are higher at the surface (Ap) soil horizons70. Soil Zn was the only nutrient that was significantly affected by residue size, confirming our hypothesis, with higher values in short residues on day 137 at 0–5 cm depth. It is estimated that about 50% of soils cultivated with cereals worldwide suffer from low Zn content with negative impact in production and grain quality71. Increased release of Zn at later stage of decomposition by short residues may be deemed as a precursor of an increased Zn release by long residues in a successive cash crop. On the contrary, higher soil Zn content could result from phytosiderophore exudates by barley plants (graminaceous monocotyledonous species) in Fe or Zn deficient soil72. In this case long residues provide an advantage by releasing Zn slowly and steadily, avoiding Zn deficiency.
Principal Component Analysis showed that Control was generally a negatively related treatment with all soil nutrients in comparison to the other treatments. This indicates the value of residue mulch in enrichment of soil nutrients even at 20–25 cm depth on both day 70 and day 137.
The significant differences on soil nutrient contents that were observed between samples from the middle and the right side of rhizotrons on day 137 imply that the effect of plant root interaction should be considered in incubation experiments.
Impact on barley plants
Yield of barley plants was not affected by residue diversityor size. This is consistent with earlier studies, e. g. Reichert et al.73 found no differences in crop (cassava) yield between treatments with chopped mulch, although it concerned residues with very small particle sizes. On the contrary, Awopegba et al.74 noticed significant differences in crop (maise) yield between types of treatments with chopped material applied on soil with a traditional hoe. In our experiment, there were indications that long residues resulted in higher seed protein content than short residues. Long residues have slower rate of decomposition and are thus able to provide more N at later stage of decomposition (GS37)58 than the short residues,which is crucial to increase protein content of grains75. Indeed, long residues resulted in significantly higher rates of barley stem elongation 42 days after plant emergence (at GS31 to GS39—rapid stem elongation stage) compared to short residues. This indicates increased N supply by long residues at the stage of flag leaf emergence (GS37), which results in increased seed protein content because N uptake by plants at GS37 is transferred to ear and seed development at GS59 GS8775. The linear regression analysis confirmed the association of the main stem elongation rate on day 42 after plant emergence with the final seed protein content, which showed a small but significant influence. In addition, this relationship was further confirmed by a Spearman’s correlation which showed a significantly negative correlation between final seed protein content and length of ears. Therefore, it seems that long residues contributed to higher main stem elongation rate on day 42 after plant emergence resulting in shorter ear length and higher seed protein content opposite to the short residues. In contrast mulch diversity was not found to impact protein content. The effect of residue size on grains’ protein content should be further investigated as it is of great interest for both farmers and food processors.
In line with our hypothesis, it has already been shown that long residues had higher final C:N ratios, C, and N in comparison to short residues. In addition, it has been shown that there were no significant differences in residue dry mass loss or in crop yield between different residue size. Therefore, we conclude that long residues are characterised by an enhanced potential to provide nutrients to soil microbes and to the next crop at the end of the growing season than the short residues, without a yield penalty. At the same time, long residues provide better soil surface coverage with organic material. Considering results were derived from a one growing season mesocosm experiment, it is highly possible the iteration of the practice of using long residues as mulch in successive crops could result in continuous enrichment of soil with nutrients, increase of soil organic matter, better physical conditions on soil surface, improvement of soil microbial community, and higher cost effectiveness in comparison to short residues. Further long-term research is needed to confirm this supposition.
Influence on symbiotic relationships
The AMF root colonisation was not significantly different between treatments. However, mean values were increased with increasing number of residue plant species. Regression analysis, considering the species richness (17, 12, 6, 1 and 0) of the different residue types and Control, showed a statistically significant influence of species richness in AMF root colonisation. However, the predictor could explain only a small variation of the response. Nevertheless, an increase in plant residue species richness on the soil surface is responsible for even a small increase in AMF root colonisation of the crop plants is very important in terms of agricultural economic performance. This was consistent with the previous finding by Burrows and Pfleger22 who observed increasing AMF sporulation with an increasing number of species of cover crop plants, attributed to an increased number of AMF species triggered by the increased cover crop plant diversity. In our experiment, the potential increase in AMF colonisation in barley plant roots was triggered plainly by the deposition of cover crops cultivated elsewhere. This indicated that residue mulch diversity may influence AMF root colonisation, which is confirming previous research76, but further investigation is needed.
Conclusions
Dry mass loss of plant residue mulch was significantly affected by residue chemical composition. It was higher in residues of lower recalcitrance (NDF content) and with lower N content. Moreover, the final quality of residues was strongly affected by residue size, resulting in higher C and N content of long residues in comparison to short ones. This indicates a higher fertilization capacity of long residues than of short ones for subsequent crops with continuous application of mulches. Soil K and Zn were the only nutrients that were significantly affected by residue quality and size, respectively, at later decomposition stages. Treatments of higher initial C:N ratio were related with higher amounts of soil K, while short residues provided more Zn. Crop yield was not affected by residue quality or size. Long residues supported significantly higher rates of barley stem elongation than short residues at the stage of rapid stem elongation, where N availability is determined for high seed protein content. There were indications that mulches with long residues increased seed protein content, which is important for farmers and grain processors. Therefore, residue size may be used to regulate N release with crop demand. The Arbuscular Mycorrhizal Fungi root colonisation was higher but not significantly different in treatments with higher plant species richness, which indicates a possible effect of mulch diversity on symbiotic relationships. In summary, long residue mulches composed of diverse plant species mixtures have enhanced residual fertilisation capacity at the end of the growing season than short residues with no deleterious effect on crop yield. Further long-term research is needed to investigate the effect of continuous application of plant residue mulches on the enrichment of soil nutrient content, the increase of soil organic matter, and the interactions with symbiotic microbial communities.
References
Sharma, O.P., Bambawale, O.M., Gopali, J.B., Bhagat, S., Yelshetty, S., Singh, S.K., Anand, R., & Singh, O.M. Field Guide Mung Bean and Urd Bean. (Government of India, Department of Agricultural and Co-Operation, NCIPM, ICAR, 2011).
Mulumba, L. N. & Lal, R. Mulching effects on selected soil physical properties. Soil Tillage Res. 98(1), 106–111. https://doi.org/10.1016/j.still.2007.10.011 (2008).
Abiven, S. & Recous, S. Mineralisation of crop residues on the soil surface or incorporated in the soil under controlled conditions. Biol. Fertil. Soils 43(6), 849–852. https://doi.org/10.1007/s00374-007-0165-2 (2007).
Su, W. et al. Influence of rice straw mulching on seed yield and nitrogen use efficiency of winter oilseed rape (Brassica napus L.) in intensive rice–oil seed rape cropping system. Field Crops Res. 159(Supplement C), 53–61. https://doi.org/10.1016/j.fcr.2014.01.007 (2014).
Peterson, G. A. et al. Reduced tillage and increasing cropping intensity in the Great Plains conserves soil C. Soil Tillage Res. 47(3), 207–218. https://doi.org/10.1016/S0167-1987(98)00107-X (1998).
Blanco-Canqui, H., Schlegel, A. J. & Heer, W. F. Soil-profile distribution of carbon and associated properties in no-till along a precipitation gradient in the central Great Plains. Agric. Ecosyst. Environ. 144(1), 107–116. https://doi.org/10.1016/j.agee.2011.07.004 (2011).
Carr, M. P., Gramig, G. G. & Liebig, A. M. ‘Impacts of organic zero tillage systems on crops. Weeds Soil Qual. Sustain. https://doi.org/10.3390/su5073172 (2013).
Miglécz, T. et al. Establishment of three cover crop mixtures in vineyards. Sci. Horticult. 197(Supplement C), 117–123. https://doi.org/10.1016/j.scienta.2015.09.017 (2015).
Marinari, S., Mancinelli, R., Brunetti, P. & Campiglia, E. Soil quality, microbial functions and tomato yield under cover crop mulching in the Mediterranean environment. Soil Tillage Res. 145(Supplement C), 20–28. https://doi.org/10.1016/j.still.2014.08.002 (2015).
Rodrigues, M. Â. et al. Soil nitrogen availability in olive orchards after mulching legume cover crop residues. Sci. Horticult. 158(Supplement C), 45–51. https://doi.org/10.1016/j.scienta.2013.04.035 (2013).
Calderón, F. J., Nielsen, D., Acosta-Martínez, V., Vigil, M. F. & Lyon, D. Cover crop and irrigation effects on soil microbial communities and enzymes in semiarid agroecosystems. Pedosphere 26(2), 192–205. https://doi.org/10.1016/S1002-0160(15)60034-0 (2016).
Dietrich, G. et al. Sugarcane mulch C and N dynamics during decomposition under different rates of trash removal. Agricult. Ecosyst. Environ. 243(Supplement C), 123–131. https://doi.org/10.1016/j.agee.2017.04.013 (2017).
Siczek, A. & Lipiec, J. Soybean nodulation and nitrogen fixation in response to soil compaction and surface straw mulching. Soil Tillage Res. 114(1), 50–56. https://doi.org/10.1016/j.still.2011.04.001 (2011).
Pavlu, L., Gaisler, J., Hejcman, M. & Pavlu, V. V. What is the effect of long-term mulching and traditional cutting regimes on soil and biomass chemical properties, species richness and herbage production in Dactylis glomerata grassland?. Agricult. Ecosyst. Environ. 217(Supplement C), 13–21. https://doi.org/10.1016/j.agee.2015.10.026 (2016).
Lynch, J. P. & Brown, K. M. Topsoil foraging—An architectural adaptation of plants to low phosphorus availability. Plant Soil https://doi.org/10.1023/A:1013324727040 (2001).
Finney, D. M., White, C. M. & Kaye, J. P. ‘Biomass production and carbon/nitrogen ratio influence ecosystem services from cover crop mixtures. Agron. J. (Madison, WI: The American Society of Agronomy Inc) 108, 39–52. https://doi.org/10.2134/agronj15.0182 (2016).
White, C. M. et al. Managing the trade off between nitrogen supply and retention with cover crop mixtures. Agric. Ecosyst. Environ. 237, 121–133. https://doi.org/10.1016/j.agee.2016.12.016 (2017).
Redin, M. et al. How the chemical composition and heterogeneity of crop residue mixtures decomposing at the soil surface affects C and N mineralisation. Soil Biol. Biochem. 78, 65–75. https://doi.org/10.1016/j.soilbio.2014.07.014 (2014).
Shu, X. et al. Applying cover crop residues as diverse mixtures increases initial microbial assimilation of crop residue-derived carbon. Eur. J. Soil Sci. (John Wiley & Sons, Ltd) 73(2), e13232. https://doi.org/10.1111/ejss.13232 (2022).
Fontaine, S., Mariotti, A. & Abbadie, L. The priming effect of organic matter: A question of microbial competition?. Soil Biol. Biochem. 35(6), 837–843. https://doi.org/10.1016/S0038-0717(03)00123-8 (2003).
Wu, Q., Borkovec, M. & Sticker, H. On particle-size distributions in soils. Soil Sci. Soc. Am. J. 57, 883–890 (1993).
Burrows, R. L. & Pfleger, F. L. Arbuscular mycorrhizal fungi respond to increasing plant diversity. Can. J. Bot. 80, 120–130 (2002).
Angers, D. A. & Recous, S. Decomposition of wheat straw and rye residues as affected by particle size. Plant Soil 189(2), 197–203. https://doi.org/10.1023/A:1004207219678 (1997).
Bremer, E., van Houtum, W. & van Kessel, C. Carbon dioxide evolution from wheat and lentil residues as affected by grinding, added nitrogen, and the absence of soil. Biol. Fertil. Soils 11(3), 221–227. https://doi.org/10.1007/BF00335771 (1991).
Jensen, E.S. & Ambus, P. Effects of barley straw particle size and distribution in soil on soil N dynamics and plant growth. in Proceedings of the “Soil Nitrogen Immobilisation Workshop” at MLURI, Aberdeen 2–3 November, 1994 (1994).
Santonja, M. et al. Contrasting responses of bacterial and fungal communities to plant litter diversity in a Mediterranean oak forest. Soil Biol. Biochem. (Elsevier) 125(June), 27–36. https://doi.org/10.1016/j.soilbio.2018.06.020 (2018).
Hättenschwiler, S., Fromin, N. & Barantal, S. Functional diversity of terrestrial microbial decomposers and their substrates. C.R. Biol. 334(5), 393–402. https://doi.org/10.1016/j.crvi.2011.03.001 (2011).
LECO Corporation. https://www.leco.com/product/628series. Accessed 4 Mar 2018 (2018).
ANKOM Technology. https://www.ankom.com/product-catalog/ankom-200-fiber-analyzer. Accessed 19 Oct 2018 (2018).
Vierheilig, H., Coughlan, A. P., Wyss, U. & Piche, Y. Ink and Vinegar, a simple staining technique for arbuscular mycorrhizal fungi. Appl. Environ. Microbiol. 64, 5004–5007 (1998).
Blakemore, L. C., Searle, P. L. & Daly, B. K. Methods for Chemical Analysis of Soils (1987).
Great Britain. M.A.F.F. The Analysis of Agricultural Materials : A Manual of the Analytical Methods Used by the Agricultural Development and Advisory Service/Ministry of Agriculture, Fisheries and Food. (eds. G. B. A. D. and A. Service & F. and F. Great Britain). 427 (Ministry of Agriculture, H.M.S.O Reference book (Great Britain. Ministry of Agriculture, Fisheries and Food), 1986).
SCALAR Instruments. https://www.skalar.com/analyzers/automated-wet-chemistry-analyzers/. Accessed 23 Nov 2018 (2018).
Mehlich, A. ‘Mehlich 3 soil test extractant: A modification of Mehlich 2 extractant’. Commun. Soil Sci. Plant Anal. (Taylor & Francis) 15(12), 1409–1416. https://doi.org/10.1080/00103628409367568 (1984).
PerkinElmer, Inc. https://www.perkinelmer.com/. Accessed 11 Nov 2018 (2018).
Pierzynski, G. M. Methods of Phosphorus Analysis for Soils, Sediments, Residuals, and Waters (2000).
AgroEcoLab”UMD. Field and Lab Methods. Mehlich-3 Extraction for Labile Soil Phosphorus. https://www.agroecologylab.com/methods. Accessed 17 Oct 2018 (2018).
Bouyoucos, G. J. Hydrometer method improved for making particle size analyses of soils. Agron. J. 54(5), 464–465. https://doi.org/10.2134/agronj1962.00021962005400050028x (1962).
Harding, D. E. & Ross, D. J. ‘Some factors in low-temperature storage influencing the mineralisable-nitrogen of soils. J. Sci. Food Agricult. (John Wiley & Sons Ltd) 15(12), 829–834. https://doi.org/10.1002/jsfa.2740151203 (1964).
Gradwell, B. M. W. & Birrell, K. S. Methods for Physical Analysis of Soils (1979).
Minitab, LLC. https://www.minitab.com/en-us/company/. Accessed 14 Dec 2017 (2017).
RStudio, PBC. https://rstudio.com/. Accessed 14 Dec 2017 (2017).
Trinsoutrot, I. et al. ‘Biochemical quality of crop residues and carbon and nitrogen mineralization kinetics under nonlimiting nitrogen conditions. Soil Sci. Soc. Am. J. (John Wiley & Sons Ltd) 64(3), 918–926. https://doi.org/10.2136/sssaj2000.643918x (2000).
Frouz, J., Roubíčková, A., Heděnec, P. & Tajovský, K. ‘Do soil fauna really hasten litter decomposition? A meta-analysis of enclosure studies. Eur. J. Soil Biol. 68, 18–24. https://doi.org/10.1016/j.ejsobi.2015.03.002 (2015).
Brust, G. E. Chapter 9—Management Strategies for Organic Vegetable Fertility (Biswas, D., Micallef, S. A. B. T.-S., & P. for O. F. eds.). 193–212. https://doi.org/10.1016/B978-0-12-812060-6.00009-X (Academic Press, 2019).
Bonanomi, G. et al. Soil fertility promotes decomposition rate of nutrient poor, but not nutrient rich litter through nitrogen transfer. Plant Soil 412, 397–411 (2017).
Vahdat, E., Nourbakhsh, F. & Basiri, M. Lignin content of range plant residues controls N mineralisation in soil. Eur. J. Soil Biol. 47(4), 243–246. https://doi.org/10.1016/j.ejsobi.2011.05.001 (2011).
Spain, A. V. & Hodgen, M. J. Changes in the composition of sugarcane harvest residues during decomposition as a surface mulch. Biol. Fertil. Soils 17(3), 225–231. https://doi.org/10.1007/BF00336327 (1994).
Akratos, C. S., Tekerlekopoulou, A. G., Vasiliadou, I. A. & Vayenas, D. V. Chapter 8—Cocomposting of Olive Mill Waste for the Production of Soil Amendments. (Galanakis, C. M. B. T.-O. M. W. ed.). 161–182. https://doi.org/10.1016/B978-0-12-805314-0.00008-X (Academic Press, 2017).
Jensen, E. S. Plant Litter Particle Size—Effects on Decomposition and Nitrogen Dynamics in Soil (2014).
Jensen, L. S. et al. Influence of biochemical quality on C and N mineralisation from a broad variety of plant materials in soil. Plant Soil 273(1), 307–326. https://doi.org/10.1007/s11104-004-8128-y (2005).
Grosso, F., Bååth, E. & De Nicola, F. Bacterial and fungal growth on different plant litter in Mediterranean soils: Effects of C/N ratio and soil pH. Appl. Soil Ecol. 108(Supplement C), 1–7. https://doi.org/10.1016/j.apsoil.2016.07.020 (2016).
Strickland, M. S. & Rousk, J. Considering fungal: Bacterial dominance in soils—Methods, controls, and ecosystem implications. Soil Biol. Biochem. (Elsevier Ltd) 42(9), 1385–1395. https://doi.org/10.1016/j.soilbio.2010.05.007 (2010).
Ambus, P., Jensen, E. S. & Robertson, G. P. Nitrous oxide and N-leaching losses from agricultural soil: Influence of crop residue particle size, quality and placement. Phyton-Ann. Rei Bot. 41(3), 7–15 (2001).
Fog, K. The Effect of Added Nitrogen on the Rate of Decomposition of Organic Matter. 433–462 (1988).
Xu, Y. et al. Dominant effects of organic carbon chemistry on decomposition dynamics of crop residues in a Mollisol. Soil Biol. Biochem. 115, 221–232. https://doi.org/10.1016/j.soilbio.2017.08.029 (2017).
Shu, X. et al. Cover crop residue diversity enhances microbial activity and biomass with additive effects on microbial structure. Soil Res. https://doi.org/10.1071/SR21105 (2021).
Tottman, D. R., Makepeace, R. J. & Broad, H. The decimal code for the growth stages of cereals, with illustrations. Ann. Appl. Biol. (Wiley/Blackwell) 93(2), 221–234. https://doi.org/10.1111/j.1744-7348.1979.tb06534.x (1986).
YARA. Crop Nutrition—Barley, Optimal Rate and Timing of Nutrients in Barley. https://www.yaracanada.ca/crop-nutrition/barley/optimal-rate-and-timing-of-nutrients/. Accessed 6 June 2018 (2021).
Brady, C. N. & Weil, R. R. The Nature and Properties of Soils (Pearson Education Inc., 2002).
Justes, E., Beaudoin, N., Bertuzzi, P., Charles, R., Constantin, J., Dürr, C., Hermon, C., Joannon, A., Le Bas, C., Mary, B., Mignolet, C., Montfort, F., Ruiz, L., Sarthou, J. P., Souchère, V., Tournebize, J., Savini, I. & Réchauchère, O. The use of cover crops to reduce nitrate leaching: Effect on the water and nitrogen balance and other ecosystem services. in The International Fertiliser Society Meeting (2012).
Jalota, S. K., Vashisht, B. B., Sharma, S. & Kaur, S. Chapter 1—Emission of Greenhouse Gases and Their Warming Effect (Jalota, S. K. et al. eds.). 1–53. https://doi.org/10.1016/B978-0-12-809520-1.00001-X. (Academic Press, 2018).
Malhi, S. S., Johnston, A. M., Schoenau, J. J., Wang, Z. H. & Vera, C. L. Seasonal biomass accumulation and nutrient uptake of pea and lentil on a black chernozem soil in Saskatchewan. J. Plant Nutr. 30(5), 721–737. https://doi.org/10.1080/01904160701289578 (2007).
Ambus, P. & Jensen, E. S. Nitrogen mineralisation and denitrification as influenced by crop residue particle size. Plant Soil 197(2), 261–270. https://doi.org/10.1023/A:1004276631914 (1997).
Zhonglu, G., Shuhua, Z., Juan, J. & Chongfa, C. Nitrogen mineralisation controlled by N/P ratio of plant residues from riparian buffer strip. Eur. J. Soil Biol. 67, 5–11. https://doi.org/10.1016/j.ejsobi.2014.12.004 (2015).
Fox, R. H., Myers, R. J. K. & Vallis, I. The nitrogen mineralisation rate of legume residues in soil as influenced by their polyphenol, lignin, and nitrogen contents. Plant Soil 129(2), 251–259. https://doi.org/10.1007/BF00032420 (1990).
Singh, B. & Khind, S. C. Nutrient transformations in soils amended with green manures. Adv. Soil Sci. https://doi.org/10.1007/978-1-4612-2930-8_5 (1992).
Xie, K., Cakmak, I., Wang, S., Zhang, F. & Guo, S. Synergistic and antagonistic interactions between potassium and magnesium in higher plants. Crop J. (Crop Science Society of China and Institute of Crop Science) https://doi.org/10.1016/j.cj.2020.10.005 (2020).
Campbell, C. A., Biederbeck, V. O., Winkleman, G. E. & Lafond, G. P. Influence of legumes and fertilisation on deep distribution of available phosphorus (Olsen-P) in a thin Black Chernozemic soil. Can. J. Soil Sci. 73(4), 555–565. https://doi.org/10.4141/cjss93-055 (1993).
Fageria, N. K., Baligar, C. & Clark, R. B. Micronutrients in Crop Production, Advances in Agronomy. https://doi.org/10.1016/s0065-2113(02)77015-6 (Elsevier Inc., 2002).
Graham, R. D., & Welch, R. M. Breeding for staple food crops with high micronutrient density. In Working Papers on Agricultural Strategies for Micronutrients No. 3. (International Food Policy Research Institute, 1996).
Li, L., Tilman, D., Lambers, H. & Zhang, F. S. Plant diversity and overyielding: Insights from belowground facilitation of intercropping in agriculture. New Phytol. 203(1), 63–69. https://doi.org/10.1111/nph.12778 (2014).
Reichert, J. M. et al. Fragmentation, fiber separation, decomposition, and nutrient release of secondary-forest biomass, mechanically chopped-and-mulched, and cassava production in the Amazon. Agric. Ecosyst. Environ. 204(Supplement C), 8–16. https://doi.org/10.1016/j.agee.2015.02.005 (2015).
Awopegba, M., Oladele, S. & Awodun, M. Effect of mulch types on nutrient composition, maise (Zea mays L.) yield and soil properties of a tropical Alfisol in Southwestern Nigeria. Eur. J. Soil Sci. (EJSS) 6, 121. https://doi.org/10.18393/ejss.286546 (2017).
Boyle, G. The Spring Barley Guide Crops Environment & Land Use Programme. Teagasc. https://www.teagasc.ie/media/website/publications/2015/The-Spring-Barley-Guide.pdf. (2017).
Bainard, L. D., Klironomos, J. N. & Gordon, A. M. Arbuscular mycorrhizal fungi in tree-based intercropping systems: A review of their abundance and diversity. Pedobiologia 54(2), 57–61. https://doi.org/10.1016/j.pedobi.2010.11.001 (2011).
Gaitanis, D. Raw Data of an Experiment to Assess the Effect of Plant Residue Size, Quality, and Diversity, Applied as Mulch, on Soil Nutrient Dynamics, on Arbuscular Mycorrhizal Fungi Root Colonisation, and on Crop Plant Yield and Quality. https://doi.org/10.17864/1947.000395. (University of Reading, Dataset, 2022).
Acknowledgements
Authors would like to thank the contributors of the DiverseForages Project and Dr Anna Tomson in person who kindly provided plant residue material. Data supporting the results reported in this paper are openly available from the University of Reading Research Data Archive at https://doi.org/10.17864/1947.00039577.
Author information
Authors and Affiliations
Contributions
Conceptualisation, D.G.; Experimental design, D.G., M.T., M.L.; Experimental work, D.G.; Laboratory analyses, D.G.; Data analysis, D.G.; Data curation, D.G., M.L., and M.T.; Supervision, M.L. and M.T.; Writing-original draft, D.G.; Writing-review and editing, D.G., M.L., and M.T.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Gaitanis, D., Lukac, M. & Tibbett, M. Fragment size and diversity of mulches affect their decomposition, nutrient dynamics, and mycorrhizal root colonisation. Sci Rep 13, 9383 (2023). https://doi.org/10.1038/s41598-023-36457-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-36457-x
- Springer Nature Limited