
Abstract
Natural Killer (NK) cells have been implicated in recurrent pregnancy loss (RPL). The p.Val176Phe (or Val158Phe) Single Nucleotide Polymorphism (SNP) in the FCGR3A gene encoding the FcγRIIIA or CD16a receptor has been associated with an enhanced affinity for IgG and stronger NK-mediated antibody-dependent cellular cytotoxicity. We hypothesized that the presence of at least one p.176Val variant associates with RPL and increased CD16a expression and alloantibodies e.g., against paternal human leukocyte antigen (HLA). In 50 women with RPL, we studied frequencies of the p.Val176Phe FCGR3A polymorphisms. Additionally, CD16a expression and anti-HLA antibody status were analyzed by flowcytometry and Luminex Single Antigens. In woman with RPL, frequencies were: 20% (VV), 42% (VF) and 38% (FF). This was comparable to frequencies from the European population in the NCBI SNP database and in an independent Dutch cohort of healthy women. NK cells from RPL women with a VV (22,575 [18731-24607]) and VF (24,294 [20157-26637]) polymorphism showed a higher expression of the CD16a receptor than NK cells from RPL women with FF (17,367 [13257-19730]). No difference in frequencies of the FCGR3A-p.176 SNP were detected when comparing women with or without class I and class II anti-HLA antibodies. Our study does not provide strong evidence for an association between the p.Val176Phe FCGR3A SNP and RPL.
Similar content being viewed by others

Introduction
Recurrent Pregnancy Loss (RPL) is a devastating problem for couples trying to conceive. Despite observed clinical associations, the current knowledge on etiological factors in RPL is still incomplete and more than 50% of cases remains unexplained1. As the fetus is genetically semi-allogenic to the mother, tight regulation of immune reactivity is essential for the proper establishment and continuation of successful pregnancy2. Emerging evidence now suggests that dysregulation of Natural Killer (NK) cell function could be responsible for several cases of unexplained RPL3.
NK cells are innate lymphocytes and peripheral blood NK cells (pNK) are primarily known as potent killers of virally-infected or tumor cells. NK cells kill through the release of granzyme and perforin containing granules or via death receptors4. Moreover, they can produce cytokines contributing to Th1 polarization and CD8 T cell function5. Two important lineages of NK cells have been described that can be classified according to the expression level of CD56: CD56dim NK cells predominantly express the FcγRIIIA receptor also referred to as CD16a and constitute around 90% of NK cells in peripheral blood and mediate cytotoxicity. Conversely, CD56bright NK cells lack expression of CD16a and while they are mostly known for their capacity to produce cytokines, they are poorly cytotoxic and form merely 5–10% of NK cells in peripheral blood6. In the secretory phase of the menstrual cycle and during early pregnancy, CD56brightCD16anegative NK cells constitute the predominant lymphocyte subset present in the non-pregnant endometrial or pregnant decidual layer of the uterus7. In contrast to their killing function in peripheral blood, uterine NK (uNK) cells regulate trophoblast invasion and promote placental vasculature hence contributing to adequate placentation8,9. The considerable interest in the role of uNK cells in pregnancy complications, particularly in RPL, indicates the perceived importance of these cells in early pregnancy. Although some controversy still exists, overall results indicate that RPL is associated with several abnormalities in NK cell number and activation10.
NK cell activation is controlled by a broad panel of inhibitory- and activating receptors, and the balance between signaling via these receptors determines the level of NK cell activation11. CD16a is one of the most powerful activating receptors on NK cells and it enables NK cells to respond to antibody-coated target cells and to exert antibody-dependent cellular cytotoxicity (ADCC)12. The process of ADCC starts with the binding of an antibody to its cognate antigen expressed on the target cell surface. The Fc domain of these antibodies is then bound by CD16a expressed on NK cells, triggering the release of cytotoxic granules and subsequent lysis and death of the target cell13. The strength of NK cell mediated ADCC is determined by factors like the isotype- or fucosylation status of the antibody14,15. Moreover, the rs396991 c.526G > T single nucleotide polymorphism (SNP) in the FCGR3A gene, results in expression of either valine or phenylalanine at amino acid position 176 (also known as position 158 in the mature protein excluding signal peptides) and has been reported to influence human IgG1 binding and ADCC activity16. Functional studies have shown that the presence of a homozygous p.176-valine (VV) results in a CD16a receptor with a higher affinity for IgG1 and IgG3 antibodies compared to the homozygous p.176-phenylalanine genotype (FF) and the triggering of NK cell activation or ADCC at a lower antibody concentration17,18,19,20. Moreover, two of these studies also suggested a gene dose effect since NK cells from heterozygous individuals showed an intermediate response17,18. ADCC is a major mechanism of action of therapeutic monoclonal antibodies (mAbs) such as rituximab and a higher ADCC response to rituximab as well as increased CD16a expression have been reported to correlate with the genotypes encoding a p.176-valine in the FCGR3A gene18. Nonetheless, another study did not find a relation between the p.176 polymorphism and CD16a expression on NK cells21. Multiple clinical studies in patients with various types of cancers, showed that homozygosity (VV), or in some studies the presence of at least one high affinity allele (Vx), was associated with an improved response to treatment with therapeutic antibodies like rituximab, trastuzumab or cetuximab16.
The percentage of uNK cells that express CD16a under physiological conditions is low, however, an elevated percentage of CD16a positive cells has been observed in woman with pregnancy complications19,20. In a mouse study, uNK cells have been shown to mediate ADCC against invading trophoblast cells of the fetus in an CD16a dependent manner23. Furthermore, 30% of women develop antibodies against paternal human leukocyte antigen (HLA) alloantigens expressed by the fetus24. These alloantibodies are considered a harmless phenomenon during most pregnancies primarily because the fetal cells most closely in contact with the immune system almost completely lack expression of polymorphic HLA class I and class II molecules25. Nevertheless, several studies suggested that these anti-HLA antibodies were associated with RPL as a higher incidence of anti-HLA antibodies has been observed in women with RPL compared to women without26. Although the potential underlying mechanism is not very well explored, these alloantibodies may contribute to RPL via complement fixation and/or ADCC resulting in damage of the invading trophoblast cells showing aberrant expression of polymorphic HLA molecules.
Given the possible impact of FCGR3A polymorphism on the strength of the ADCC response, we hypothesized that in women with RPL having at least one FCGR3A-p.176Val variant enhances the ADCC response, see Fig. 1. As such, we identified FCGR3A-p.176 polymorphisms in women with RPL. We compared these frequencies to the frequencies in the NCBI single nucleotide polymorphisms database and an independent cohort of healthy women. In addition, we sought to delineate differences in NK cell surface CD16a expression in women with RPL with a VV, VF and FF SNP. Since we expect the effect of FCGR3A polymorphism to be most relevant in women with antibodies against fetal alloantigens, we also determined HLA antibody status in women with RPL and studied distribution of the polymorphic subgroups.

Illustrative representation of antibody dependent cellular cytotoxicity (ADCC) of Natural Killer (NK) cells in the uterus with a CD16a receptor having two phenylalanine alleles (FF) depicted in brown and a CD16a receptor having at least one valine allele (VV/VF) depicted in purple. We hypothesize that a receptor having at least one valine allele induces a stronger ADCC response and an increased CD16a-mediated degranulation capacity of NK cells against the trophoblast in women with recurrent pregnancy loss (RPL).
Results
Baseline characteristics of women with RPL
Women with RPL were on average 31 years old, had on average 5 pregnancies, 4 pregnancy losses and no live birth (Table 1).
Frequencies of FCGR3A-p.176 genotypes in women with RPL are similar to frequencies in a European cohort and an independent cohort of healthy women
To study frequencies of the p.Val176Phe FCGR3A genotype in women with RPL, single-nucleotide changes in position 526 (G → T) were detected by sanger sequencing. Based on position 176, the following subgroups of FCGR3A genotypes were identified: valine/valine (VV), valine/phenylalanine (VF) and phenylalanine/phenylalanine (FF). Genotypes were subsequently identified as (VV/VF and FF) subgroups. The frequencies were as follows: 31 out of 50; 62% (VV/VF) and 19 out of 50; 38% (FF) for women with RPL (Table 2). When genotypes were identified as (VF/FF) and (VV) subgroups the frequency was 40 out of 50; 80% for (VF/FF) and 10 out of 50; 20% for (VV). No significant deviation from the Hardy–Weinberg expectation was observed.
Since no matched control group was available, the frequencies of a general European cohort were obtained from the NCBI single nucleotide polymorphisms database (dbSNP) and they were as follows: 52.4% (VV/VF) and 47.6% (FF) and additionally 90.4% (VF/FF) and 9.6% (VV) (Table 2). In addition, frequencies were used from a Dutch cohort of healthy females that was genotyped by MLPA for a previously published study. The frequencies for this cohort were comparable to the European cohort: 87 out of 164; 53% (VV/VF), 77 out of 164; 47% (FF) and 142 out of 164; 86.6% (VF/FF), 22 out of 164; 13.4% (VV) (Table 2). No significant deviation from the Hardy–Weinberg expectation was observed.
To enable statistical comparison of both cohorts, we first validated that the MLPA method used for typing of the healthy female cohort and the SBT method used for typing of the RPL cohort gave similar results. To do this, 5 women with RPL were genotyped for FCGR3A p.V176F by MLPA which gave identical genotypes as was obtained when the samples were analyzed by SBT. A statistical comparison of the RPL and the healthy control cohort subsequently showed that there was no significant difference between women with RPL and healthy women (P = 0.265) when comparing (VV/VF) and (FF) subgroups. Furthermore, there was no significant difference between women with RPL and the healthy control cohort when comparing (VF/FF) versus (VV) (P = 0.253). In addition, there was no significant difference (P = 0.374) when frequencies were compared for VV (20%; 10 out of 50), VF (42%; 21 out of 50) and FF (38%; 19 out of 50) in women with RPL, versus respectively 13.4%; 22 out of 164 (VV), 39.6%; 65 out of 164 (VF) and 47%; 77 out of 164 (FF) in healthy women (Table 2).
Analysis of the influence of the p.V176F polymorphism on CD16a positive NK cells
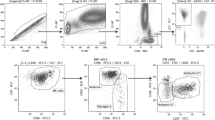
In addition to qualitative differences, the p.V176F SNP has been shown to result in quantitative differences in the level of receptor expression on NK cells. To study if these quantitative differences also occurred in our cohort of RPL women, the percentage of CD16a positive NK cells and the level of expression were determined by flowcytometry (gating strategy supplementary file) and compared between VV/VF and FF subgroups and additionally between VF/FF and VV subgroups of women with RPL. First, we compared distribution of total NK cells, defined as CD3negativeCD56positive and of the two main NK cell subsets; the CD56brightCD16anegative mostly known as cytokine producers and the cytotoxic CD56dimCD16apositive. No significant differences were found between the VV/VF and the FF subgroup when comparing percentage of total CD3negativeCD56positive NK cells (VV/VF 10.8 [7.5-17.4] versus FF 10.0 [8.0-19.6] P = 0.708), the percentage of CD56dimCD16apositive NK cells (VV/VF 88.1 [85.3-92.4] versus FF 87.8 [81.5-90.8] P = 0.612), or the percentage of CD56brightCD16anegative NK cells (VV/VF 4.1 [2.6-5.9] versus FF 4.0 [3.3-7.4] P = 0.470) (Fig. 2A–C).

Frequencies of NK cell subsets in women with RPL genotyped for CD16a. Percentage of total CD3negativeCD56positive NK cells from lymphocytes (A), CD56dimCD16apositive NK cells (B) and CD56brightCD16anegative NK cells (C) as measured by flow cytometry in women whose genotyping demonstrated FCGR3A-p.176 at least one valine allele (VV/VF) versus (FF) (A/B/C) or at least one phenylalanine allele (VF/FF) versus (VV) (D/E/F) or VV, VF and FF (G/H/I) respectively. Dots depict individuals, lines depict median and inter quartile range per group.
Second, no significant differences were found between VF/FF and VV subgroups when comparing percentage of total CD3negativeCD56positive NK cells (VF/FF 12,5 [8.3-18.6] versus VV 8,6 [5.2-10.8] P = 0.080), the percentage of CD56dimCD16apositive NK cells (VF/FF 88.1 [85.9-92.2] versus VV 83.0 [79.0-88.5] P = 0.096), or the percentage of CD56brightCD16anegative NK cells (VF/FF 3.9 [2.7-5.9] versus VV 6.1 [4.1-6.6] P = 0.099) (Fig. 2D–F).
Additionally, there was no significant difference in percentage of total NK cells (P = 0.120) when percentages were compared between VV (8.6 [5.2-10.8]), VF (14.3 [9.5-18.3]) and FF (10.0 [8.0-19.6]). Neither when percentage of CD56dimCD16apositive NK cells (P = 0.123); VV (83.0 [79.0-88.5]), VF (89.3 [86.2-92.7]), FF (87.8 [81.5-90.8), nor when percentage of CD56brightCD16anegative NK cells (P = 0.094); VV (6.1 [4.1-6.6]), VF (3.9 [2.3-4.9), FF (4.0 [3.3-7.4]) was compared between VV, VF and FF (Fig. 2G–I).
Next, the percentage of CD16a on all NK cells, irrespective of the expression level of CD56, was compared between the VV/VF and FF groups among women with RPL. No significant difference (P = 0.329) was observed between both groups (VV/VF 94.8 [93.0-96.7]) versus FF: 94.6 [90.7-95.5]) (Fig. 3A). Similarly, there was no significant difference between polymorphic subgroups when percentages of CD16a on total CD3negativeCD56positive NK cells (P = 0.176) were compared between (VF/FF): 95.0 [92.5-96.6] and (VV): 93.1 [92.6-94.9] (Fig. 3B). Moreover, there was no significant difference between subgroups when percentages of CD16a on total CD3negativeCD56positive NK cells (P = 0.116) were compared for VV (93.1% ([92.6-94.9]), VF (95.0% [94.3-97.0]) and FF (94.6% [90.7-95.5]) (Fig. 3C).

CD16a expression on NK cells in women with RPL genotyped for CD16a. Percentage of CD16a positive cells of total CD3negativeCD56positive NK cells (A/B/C) and MFI (D/E/F) normalized by FMO of CD16a expression measured by flow cytometry in women with at least one valine (VV/VF) versus (FF) FCGR3A-p.176 genotypes (A and D) or at least one phenylalanine allele (VF/FF) and (VV) genotypes (B and E) or VV, VF and FF genotypes (C and F). Dots depict individuals, lines depict median and interquartile range per group, *P < 0.05. Representative histogram of CD16a expression on NK cells (G) depicting FMO (no anti-CD16a) in grey and CD16a expression for the (VV) genotype in purple and the (FF) genotype in brown.
However, we observed a higher MFI for CD16a in RPL women with the VV/VF genotype (23575 [19782-26220]) versus the FF genotype (17367 [13257-19730]) (P = 0.001, Fig. 3D) but not in women in the (VF/FF) group (19924 [14971-25454]) versus the (VV) group (22575 [18731-24607]) (P = 0.603), Fig. 3E). In line with those data, the CD16a MFI was higher for RPL women with the VV or VF genotype than for RPL women with the FF genotype: MFI for VV was 22575 [18731-24607], for VF was 24294 [20157-26637] and for FF was 17367 [13257-19730] (P = 0.003) (Fig. 3F). All MFI values were normalized with a fluorescent minus one (FMO) (Fig. 3G).
No obvious association between HLA antibody status and frequencies of FCGR3A-p.176 genotypes
The impact of the FCGR3A-p.176 polymorphism will primarily be relevant in combination with maternal IgG alloantibodies with the potential to trigger NK cell mediated ADCC against fetal cells expressing paternal alloantigens. In the transplantation setting, the highly polymorphic family of HLA molecules (HLA-A/-B/C and HLA-DR/-DQ/-DP) represents the most important alloantigens and anti-HLA antibodies can provoke complement- and/or cell-mediated cytotoxicity against cells of the allograft27. Even in normal pregnancies, approximately 30% of women develop anti-HLA antibodies24. To obtain initial evidence for a potentially synergistic role of anti-HLA antibodies and FCGR3A-p.176 polymorphism, we determined, as a small pilot, the presence and specificity of anti-HLA antibodies by Luminex Single Antigens in 32 women from our RPL cohort. Anti-HLA antibodies were present in 26 out of 32 women (Fig. 4). Antibodies were directed against HLA class I in 11/32 women, against HLA class II in 3/32 women or against both, class I and class II in 12/32 women and we further defined specificity of the HLA class I and -class II antibodies found in the 32 tested women with RPL (Fig. 4). Out of 23 women who had antibodies against class I, 7 had antibodies against HLA-B, 3 against HLA-A, 2 against HLA-C, 1 against HLA-A + C, 2 against HLA-B + C, 3 against HLA-A + B and 5 against HLA-A + B + C. Out of 15 women who had antibodies against class II, 1 had antibodies against HLA-DP, 4 against HLA-DQ, 1 against HLA-DQ + DRB1, 1 against HLA-DQ + DRB3/4/5, 4 against HLA-DQ + DRB1 + DRB3/4/5, 1 against HLA-DRB1 + DRB3/4/5, 1 against HLA-DP + DRB3/4/5 and 2 against HLA-DP + DQ + DRB1 + DRB3/4/5 (Fig. 4).

Specificity of anti-HLA antibodies. Anti-HLA antibodies, including HLA-A, B, C, DP, DQ, and DR, were detected in sera of 32 women with RPL. Specificity and number of individuals that were positive for anti-HLA antibodies are indicated in a Venn diagram representation.
Next, we determined the frequencies of (VV/VF) and (FF) polymorphic subgroups of RPL women and compared them between women with and without antibodies. Frequencies of VV/VF in women with antibodies were 57,7% (class I and/or class II Ab positive), 60,9% (class I Ab positive), 46,7% (class II Ab positive) compared to 66,7%% in women who completely lacked anti-HLA antibodies, 55,5% in women without class I antibodies and 70,6% in women without HLA class II antibodies (Table 3). In this small group of patients, we did not observe a significant difference in frequencies of the p.Val176Phe genotypes (VV/VF versus FF) based on antibody status (class I and/or class II (P = 0.687), class I (P = 0.783) and class II (P = 0.169)). Additionally, we determined the frequencies of (VF/FF) and (VV) polymorphic subgroups of RPL women and compared them between women with and without antibodies in Table 3, showing that there was no significant difference in frequencies of the VF/FF versus VV genotypes based on antibody status (class I and/or class II (P = 0.063), class I (P = 0.186) and class II (P = 0.100)). Likewise, we did not observe a difference between women with- or without anti-HLA antibodies in frequencies of the VV, VF and FF genotypes (class I and/or class II (P = 0.083), class I (P = 0.285) and class II (P = 0.190)) (Table 3). Due to the low power, we did not evaluate CD16a in groups defined based on specificity of the anti-HLA antibodies.
The detrimental effect of the presence of donor specific antibodies (DSA) against HLA has clearly been shown in the kidney transplantation setting, where in the case of a living donor especially DSA against both HLA class I and class II, and in the case of a deceased donor DSA against HLA class I or against both HLA class I and II has been associated with reduced graft survival28. Hence, we compared the frequencies of VV/VF versus FF polymorphic subgroups, of VF/FF versus VV subgroups and of VV, VF and FF genotypes between women without antibodies, women with only class I antibodies, women with only class II antibodies and women with both class I and class II antibodies. No significant differences were observed between the different groups (P = 0.527 for VV/VF versus FF, P = 0.145 for VF/FF versus VV and P = 0.310 for VV versus VF versus FF) and having no, only class I, only class II and both class I plus II antibodies (Table 4).
Discussion
The present study was conducted to investigate occurrence of the p.V176F polymorphism in the FCGR3A gene in women with RPL. Moreover, we studied the association with NK cell surface CD16a expression and performed a small pilot to explore a potential association of the p.V176F SNP and anti-HLA antibodies.
In our women with RPL the VF genotype is the most common (42%) followed by FF (38%) and VV (20%). These frequencies are in line with results from previous studies that investigated the presence of the FCGR3A-p.176 polymorphism and reported VF or FF as most frequent genotype in different populations including ethnic groups from the Netherlands, Great Britain, Austria, Australia, China, Africa29 Norway30, Singapore31 and Japan32 and with the calculated frequencies from the dbSNP for a European cohort. Moreover, there was no significant difference when frequencies of women with RPL were compared to a Dutch cohort of healthy females.
Though moderate in size (n = 50), our study included a well characterized and relatively homogenous cohort of women with RPL, as the risk for clinical confounders was kept to a minimum by pre-screening the women by means of the PCVS evaluation program. By doing so cases of pregnancy loss caused by abnormal parental karyotype, thrombophilia, aberrant endocrine factors and abnormalities in ultrasound examination were excluded. Unfortunately, obstetric history was not recorded in the healthy women and no additional blood samples were available from these women for analysis of CD16a expression and anti-HLA antibody status, making it impossible to attribute differences to previous successful or unsuccessful pregnancies.
We observed a higher MFI for CD16a on NK cells from RPL women with at least one valine allele (VV/VF) (median 23,575) than on NK cells from RPL women with the homozygous phenylalanine (FF) genotype (median 17,367). However, there was no significant difference in the percentage of NK cells expressing CD16a. In our study, we used the REA423 Miltenyi antibody that recognizes the same epitope as the frequently used anti-CD16a clone 3G8. By using multiple anti-CD16a clones binding to different epitopes, three studies concluded that differences in CD16a expression levels between the p.Val176Phe variants were the result of enhanced affinity of the 3G8 clone for FCGR3A-p.176V rather than a true difference in the number of molecules20,21,33. However, two other studies that used a 3G8 clone reported similar MFI’s for CD16a for both homozygous F and V genotypes17,34. Although we cannot completely rule out that the difference in CD16a MFI that we observed was caused by a difference in affinity of the antibody, our data are in line with a study that showed that the absolute number of CD16a receptors per NK cell was significantly higher, in addition to higher expression of CD16a both at mRNA expression and at cell surface expression level, in individuals who expressed VV at FCGR3A-p.176 versus FF18. Additionally, it would be relevant to take copy number variation (CNV) in the FCGR3A gene into account in future analysis as it has been described to correlate with CD16a expression levels35.
In our study and previous studies assessing the impact of the p.176 SNP, CD16a expression levels were measured on pNK cells. To study the impact on RPL, it would be relevant to also determine CD16a expression on uNK cells, since these are in close proximity of the invading trophoblast and can directly interact with these cells during early pregnancy36. Although feasible protocols exist, uNK cells are more difficult to obtain and require menstrual blood or endometrium biopsy sampling22,37. The link between FCGR3A-p.176 polymorphisms and uNK cell CD16a expression levels has not been studied yet. However, previous studies showed that uNK cells predominantly lack CD16a expression and have a CD56brightCD16anegative phenotype which is in contrast to pNK cells where 85–95% of NK cells is CD56dimCD16apositive38,39,40. Despite their low frequencies, CD16apositive NK cells have been described to be present in decidua of 81.4% of women with a history of antiphospholipid antibody syndrome experiencing RPL41. Also, a significantly higher absolute cell count of CD16apostive endometrial NK cells has been described in infertile women when compared with fertile women42. This suggests that CD16a may be an important marker of failed immunological adaptation during the implantation phase of early pregnancy. Furthermore, CD16a expression on uNK cells could be increased by local factors. For example, decidual NK cells have been shown to acquire CD16a expression and a more cytotoxic effector function upon interaction with cytomegalovirus (CMV) infected fibroblast43, and CMV seropositivity has been described to be more frequently present in women with RPL as compared to control women with a healthy pregnancy44. To further investigate the potential impact of FCGR3A-p.176 polymorphism, genotyping including CNV and analysis of CD16a expression levels on uNK cells could be combined with analysis of viral status.
Since we anticipated that the hypothesized effect of FCGR3A polymorphism would be most pronounced in women with alloantibodies, anti-HLA antibody status was determined. In our small cohort, we did not observe an association between HLA antibody status and frequencies of the receptor genotypes. HLA-antibodies are known to play an important role in organ transplantation as the presence of pre-transplantation donor-specific HLA-antibodies is associated with rejection and impaired organ survival45. In pregnancy, the presence of HLA antibodies is presumably largely a harmless phenomenon since they occur in relatively high numbers in normal pregnancy46. However, their role could be debated as also harmful effects of anti-HLA antibodies on pregnancy outcome have been described47. A higher incidence of anti-HLA antibodies has been observed in women with RPL compared to women without RPL26. The exact mechanism behind increased HLA antibody formation in women with RPL is currently unclear but increasing gravidity24,48 and the fetal and maternal HLA phenotype combination49 may be important determinants responsible for the higher incidence of anti-HLA antibodies in women with RPL.
In our group of women with RPL, anti-HLA antibodies were present in 81% (26 out of 32). This is higher than seen in the previous mentioned study where anti-HLA antibodies were detected in 32% of RPL cases26. Since none of our women had reported a previous blood transfusion or transplant in their medical history, antibodies present were most likely from a previous pregnancy. Unfortunately, HLA-typing of the partner was unknown so it was not entirely certain that the antibodies present were directed against paternal antigens of the current- or a previous conceptus. In our Luminex assays, we assigned a MFI signal of > 1000 as anti-HLA antibody positive which was based on our experience with these assays in routine diagnostics for kidney- and stem cell transplantation. However, it remains to be elucidated whether this is a clinically relevant cut off for RPL. Our cohort was too small to draw strong conclusions based on the difference between the presence of antibodies against HLA. For future analysis it would be relevant to study the presence of class I and class II antibodies in more detail as it has been shown in a transplantation setting that having HLA class I and II donor specific antibodies is a clear risk factor for graft survival in especially deceased donor transplants28. Having class I and II antibodies might possibly also be a risk factor for pregnancy loss in women with RPL. To study this hypothesis more thoroughly, a larger cohort of women would be necessary and follow-up of these women in necessary to see if they indeed endure more losses over a longer period of time.
In addition, there are other ADCC-inducing (allo-)antibodies, aside from HLA, that might play an important role during pregnancy. Anti-platelet antigen antibodies have been associated with pregnancy loss, as an increased maternal ADCC immune response to fetal platelet antigens has shown to cause pregnancy loss in a murine model23. However, we did not determine anti-platelet antigen antibodies in our study. In addition, anti-phospholipid antibodies have been highly associated with recurrent pregnancy loss50. The women in our cohort did not have anti-phospholipid antibodies as they were tested for during the PCVS thrombophilia screening.
To study the potential impact of humoral rejection, including ADCC, in RPL in more detail several additional factors could be included in future analysis. Antibodies can only trigger ADCC if their cognate (paternal) antigens are sufficiently expressed on fetal cells. Under physiological conditions, trophoblast cells that are in close proximity with the maternal immune system lack expression of classical HLA class Ia and class II antigens while they do express HLA class Ib antigens which are known to dampen immune responses at the feto-maternal interface25. The almost complete lack of expression of paternal polymorphic HLA molecules would normally protect trophoblast cells from binding to anti-HLA antibodies and thus from complement- or cell mediated cytotoxicity. Extravillous trophoblast cells do, however, express polymorphic HLA-C molecules and mismatches in highly immunogenic HLA-C*07 and -C*17 have been associated with RPL51. Interestingly, a higher incidence of antibodies specific for HLA-C was previously found in women with recurrent pregnancy loss illustrating that more in-depth analysis of antibody specificity may help to unravel their role in the pathophysiology of unexplained recurrent pregnancy loss. Another factor to consider is inflammatory status of the uterus, as in early pregnancy a disturbed balance between tolerogenic- and inflammatory immune reactivity may lead to aberrant expression of paternal HLA molecules contributing to humoral rejection. Although this has not been studied in the uterus yet, inflammation-induced expression of HLA has been associated with humoral rejection in the transplantation setting52. In addition, human islets do not express HLA class II under normal conditions, but under inflammatory conditions there is induced expression of HLA class II53. Increased expression of paternal alloantigens like HLA together with increased CD16a expression and a more cytotoxic profile of uNK cells could then possibly lead to ADCC in early pregnancy, while under normal conditions no ADCC would occur, and having at least one valine allele for FCGR3A-p.176 may aggravate the response.
In summary, with the current set of experiments, we did not observe an association between the p.Val176Phe FCGR3A SNP and RPL. To further investigate the role of FCGR3A-p.176 polymorphisms and their possible association to functional uterine NK cell ADCC activity in RPL pathophysiology, genomic analysis can be combined with more in-depth analysis of (allo)antibody profiles and viral status.
Materials and methods
Sample collection
Women with RPL: Blood samples of 50 women with RPL were available for genotyping the p.V176F region of FCGR3A. Of these 50 samples, 41 were available for additional flowcytometric analysis and 32 sera samples were available for anti-HLA antibody testing. All women gave informed consent and participated in the preconceptional cardiovascular assessment program (PCVS). The PCVS evaluation is performed according to Dutch national guidelines (www.nvog.nl) of RPL at least 3 months after pregnancy loss and consists of parental karyotypic screening, thrombophilia screening, endocrine screening and gynecological ultrasound. Women with 2 or more reported pregnancy losses before 24 weeks of gestation, were included. Women were excluded if outcomes from the PCVS evaluation indicated abnormal parental karyotype, thrombophilia (for example, presence of anti-phospholipid antibodies/Factor V Leiden mutation/Prothrombin mutation/Lupus anticoagulant or deficiency of Protein C, Protein S, antithrombin), endocrine abnormality (e.g., thyroid dysfunction) or uterine anomalies. Leukocytes were isolated from ethylene diamine tetra acetic acid (EDTA) blood samples and directly used for flowcytometry and additionally for DNA extraction using the QIAamp DNA blood mini kit (Qiagen, Hilden, Germany). This study was approved by the Medical Ethical Committee of the Maastricht University Medical Centre (MUMC +) (14-4-118) and in accordance with the Declaration of Helsinki. Information on baseline characteristics and blood samples were obtained upon written informed consent.
Healthy women: a cohort of 164 Dutch healthy female donors from Sanquin Amsterdam, that gave written consent and were genotyped for the FCGR3A polymorphism in the context of a previous study54.
Genotyping the p.V176F region of FCGR3A
Genotyping of the 50 women with RPL for FCGR3A (p.V176F) was performed by sanger-based typing (SBT) using an FCGR3A gene-specific forward primer and a generic reverse primer, producing a 9654 bases long polymerase chain reaction (PCR) product, following a previously described protocol55. In addition, amplicons obtained were purified by ExoSAP-IT (Affymetrix, Santa Clara, California) and then sequenced using ABI BigDye Terminator Chemistry (Life Technologies) and an ABI 3730 sequencer (Life Technologies) with a specific forward and reverse sequencing primer. Data were analyzed using DNASTAR Lasergene SeqMan Pro (DNASTAR Lasergene, Madison, Wisconsin). Data were analyzed using Genemarker version 1.40 (Soft Genetics LLC, State College, PA).
The group of 164 healthy females, was genotyped in a previous study by a FCGR-specific multiplex ligation-dependent probe amplification (MLPA) assay (MRC-Holland, Amsterdam, The Netherlands)54. To ensure that both typing methods yielded comparable genotypic results, 5 women with of the current RPL cohort were genotyped by SBT and also by MLPA and this resulted in the same typing. Genetic data was tested for Hardy–Weinberg equilibrium.
Flowcytometry
Freshly isolated leukocytes were stained with conjugated antibodies of 2uL anti-CD3 (VioBlue, REA613, Miltenyi Biotec, Germany), 2uL anti-CD56 (APC-Vio770, REA196, Miltenyi Biotec, Germany) and 2uL anti-CD16a (FITC, REA423, Miltenyi Biotec, Germany) for 30 min at 4 °C in the dark. After two wash cycles, samples were measured on a FACS Canto II (BD Biosciences, San Jose, CA) and analyzed with the BD FACSDiva Software v8.0.2 (BD Biosciences, San Jose, CA) see supplementary file for gating strategy. In order to reduce inter-experimental variations, all samples were measured at a standardized sample rate of 5000 NK cells and with application settings to standardize voltage and compensation settings. Variation from the background signal was abrogated by normalizing MFI values with a FMO, see Fig. 3G for representative histogram.
Anti-HLA antibody testing
Serum anti‐HLA antibodies were tested using single antigen beads (LABScreen, One Lambda Inc/ThermoFisher, Canoga Park, CA), according to the manufacturer’s instructions. Anti‐HLA antibody profiles were analyzed using the HLA Fusion software v4.2 (One Lambda Inc/ThermoFisher, Canoga Park, CA). Antibodies with a mean channel fluorescence intensity (MFI) lower than 1000 were considered negative and antibodies with an MFI higher than 1000 positive.
Statistical analysis
Data was tested for normality with Shapiro–Wilk. Dichotomous data was analyzed with Chi Square and was compared between women with RPL and an independent cohort of healthy women (percentages of frequencies of polymorphisms) and between women with RPL categorized as either being positive or negative for class I, class II and class I and/or class II anti-HLA antibodies and between women with RPL categorized as only having class I, class II, class I and II or no anti-HLA antibodies (percentages of frequencies of polymorphisms), respectively. Continuous data among women with RPL was analyzed with Mann Whitney U or Kruskal–Wallis (percentages of NK cells (CD3negativeCD56positive), and of the CD56dimCD16apositive and CD56brightCD16anegative NK cell subsets and percentages plus MFI of CD16a expression on NK cells). Overall, a P-value below 0.05 was considered statically significant and all statistical analyses were conducted with IBM SPSS statistics version 25 (IBM Corp, Los Angeles, USA).
Data availability
The datasets used and/or analyzed during the current study available from the corresponding author on reasonable request.
References
El Hachem, H. et al. Recurrent pregnancy loss: Current perspectives. Int. J. Womens Health 9, 331–345 (2017).
Wang, N. F., Kolte, A. M., Larsen, E. C., Nielsen, H. S. & Christiansen, O. B. Immunologic abnormalities, treatments, and recurrent pregnancy loss: What is real and what is not? Clin. Obstet. Gynecol. 59, 509–523 (2016).
Moffett, A. & Colucci, F. Uterine NK Cells: Active regulators at the maternal-fetal interface. J. Clin. Invest. 124, 1872–1879 (2014).
Paul, S. & Lal, G. The molecular mechanism of natural killer cells function and its importance in cancer immunotherapy. Front. Immunol. 8, 1124 (2017).
Pallmer, K. & Oxenius, A. Recognition and regulation of T cells by NK cells. Front. Immunol. 7, 251 (2016).
Campbell, K. S. & Hasegawa, J. Natural killer cell biology: An update and future directions. J. Allergy Clin. Immunol. 132, 536–544 (2013).
Santoni, A., Carlino, C. & Gismondi, A. Uterine NK cell development, migration and function. Reprod. Biomed. Online 16, 202–210 (2008).
Smith, S. D., Dunk, C. E., Aplin, J. D., Harris, L. K. & Jones, R. L. Evidence for immune cell involvement in decidual spiral arteriole remodeling in early human pregnancy. Am. J. Pathol. 174, 1959–1971 (2009).
Helige, C. et al. Distribution of decidual natural killer cells and macrophages in the neighbourhood of the trophoblast invasion front: A quantitative evaluation. Hum. Reprod. 29, 8–17 (2014).
Ticconi, C., Pietropolli, A., Di Simone, N., Piccione, E. & Fazleabas, A. Endometrial immune dysfunction in recurrent pregnancy loss. Int. J. Mol. Sci. 20(21), 5332 (2019).
Long, E. O., Kim, H. S., Liu, D., Peterson, M. E. & Rajagopalan, S. Controlling natural killer cell responses: Integration of signals for activation and inhibition. Annu. Rev. Immunol. 31, 227–258 (2013).
Vivier, E., Tomasello, E., Baratin, M., Walzer, T. & Ugolini, S. Functions of natural killer cells. Nat. Immunol. 9, 503–510 (2008).
Lo Nigro, C. et al. NK-mediated antibody-dependent cell-mediated cytotoxicity in solid tumors: Biological evidence and clinical perspectives. Ann. Transl. Med. 7(5), 105 (2019).
Wang, W., Erbe, A. K., Hank, J. A., Morris, Z. S. & Sondel, P. M. NK cell-mediated antibody-dependent cellular cytotoxicity in cancer immunotherapy. Front. Immunol. 6, 368 (2015).
Temming, R. et al. Functional attributes of antibodies, effector cells, and target cells affecting NK cell-mediated antibody-dependent cellular cytotoxicity. J. Immunol. 203(12), 3126–3135 (2019).
Mellor, J. D., Brown, M. P., Irving, H. R., Zalcberg, J. R. & Dobrovic, A. A critical review of the role of Fc gamma receptor polymorphisms in the response to monoclonal antibodies in cancer. J. Hematol. Oncol. 6, 1 (2013).
Koene, H. R. et al. Fc gammaRIIIa-158V/F polymorphism influences the binding of IgG by natural killer cell Fc gammaRIIIa, independently of the Fc gammaRIIIa-48L/R/H phenotype. Blood 90, 1109–1114 (1997).
Hatjiharissi, E. et al. Increased natural killer cell expression of CD16, augmented binding and ADCC activity to rituximab among individuals expressing the Fc{gamma}RIIIa-158 V/V and V/F polymorphism. Blood 110(7), 2561–2564 (2007).
Bowles, J. A. et al. Anti-CD20 monoclonal antibody with enhanced affinity for CD16 activates NK cells at lower concentrations and more effectively than rituximab. Blood 108(8), 2648–2654 (2006).
Dall’Ozzo, S. et al. Rituximab-dependent cytotoxicity by natural killer cells: Influence of FCGR3A polymorphism on the concentration-effect relationship. Cancer Res. 64(13), 4664–4669 (2004).
Congy-Jolivet, N. et al. Fc gamma RIIIa expression is not increased on natural killer cells expressing the Fc gamma RIIIa-158V allotype. Cancer Res. 68(4), 976–980 (2008).
Giuliani, E., Parkin, K. L., Lessey, B. A., Young, S. L. & Fazleabas, A. T. Characterization of uterine NK cells in women with infertility or recurrent pregnancy loss and associated endometriosis. Am. J. Reprod. Immunol. 72, 262–269 (2014).
Yougbaré, I. et al. Activated NK cells cause placental dysfunction and miscarriages in fetal alloimmune thrombocytopenia. Nat. Commun. 8, 224 (2017).
Regan, L., Braude, P. R. & Hill, D. P. A prospective study of the incidence, time of appearance and significance of anti-paternal lymphocytotoxic antibodies in human pregnancy. Hum. Reprod. 6, 294–298 (1991).
van Nieuwenhoven, A. L., Heineman, M. J. & Faas, M. M. The immunology of successful pregnancy. Hum. Reprod. 9(4), 347–357 (2003).
Meuleman, T. et al. HLA-C antibodies in women with recurrent miscarriage suggests that antibody mediated rejection is one of the mechanisms leading to recurrent miscarriage. J. Reprod. Immunol. 116, 28–34 (2016).
Petersdorf, E. W. Role of major histocompatibility complex variation in graft-versus-host disease after hematopoietic cell transplantation. F1000Res 6, 617 (2017).
Kamburova, E. G. et al. Differential effects of donor-specific HLA antibodies in living versus deceased donor transplant. Am. J. Transpl. 18(9), 2274–2284 (2018).
Nagelkerke, S. Q. et al. Extensive ethnic variation and linkage disequilibrium at the FCGR2/3 locus: Different genetic associations revealed in kawasaki disease. Front. Immunol. 10, 185 (2019).
Van Sorge, N. M. et al. Severity of Guillain-Barré syndrome is associated with Fcγ Receptor III polymorphisms. J. Neuroimmunol. 162, 157–164 (2005).
Chong, K. T., Ho, W. F., Koo, S. H., Thompson, P. & Lee, E. J. D. Distribution of the FcgammaRIIIa 176 F/V polymorphism amongst healthy Chinese, Malays and Asian Indians in Singapore. Br. J. Clin. Pharmacol. 63(3), 328–332 (2007).
Van der Pol, W. L. et al. Evidence for non-random distribution of Fcγ receptor genotype combinations. Immunogenetics 55, 240–246 (2003).
Vance, B. A., Huizinga, T. W., Wardwell, K. & Guyre, P. M. Binding of monomeric human IgG defines an expression polymorphism of Fc gamma RIII on large granular lymphocyte/natural killer cells. J. Immunol. 151(11), 6429–6439 (1993).
Wu, J. et al. A novel polymorphism of FcgammaRIIIa (CD16) alters receptor function and predisposes to autoimmune disease. J. Clin. Invest. 100(5), 1059–1070 (1997).
Breunis, W. B. et al. Copy number variation at the FCGR locus includes FCGR3A, FCGR2C and FCGR3B but not FCGR2A and FCGR2B. Hum. Mutat. 30(5), E640–E650 (2009).
Wallace, A. E., Fraser, R. & Cartwright, J. E. Extravillous trophoblast and decidual natural killer cells: A remodelling partnership. Hum. Reprod. Update 18(4), 458–471 (2012).
van der Molen, R. G. et al. Menstrual blood closely resembles the uterine immune micro-environment and is clearly distinct from peripheral blood. Hum. Reprod. 29(2), 303–314 (2014).
Saito, S., Nakashima, A., Myojo-Higuma, S. & Shiozaki, A. The balance between cytotoxic NK cells and regulatory NK cells in human pregnancy. J. Reprod. Immunol. 77(1), 14–22 (2008).
Manaster, I. & Mandelboim, O. The unique properties of human NK cells in the uterine mucosa. Placenta 29, 60–66 (2008).
Tabiasco, J. et al. Human decidual NK cells: Unique phenotype and functional properties—A review. Placenta 27, 34–39 (2006).
Gomaa, M. F., Elkhouly, A. G., Farghly, M. M., Farid, L. A. & Awad, N. M. Uterine CD56dim and CD16+ cells in refractory antiphospholipid antibody-related pregnancy loss and chromosomally intact abortuses: A case-control study. J. Hum. Reprod. Sci. 10(1), 18–23 (2017).
Junovich, G. et al. Endometrial CD16+ and CD16- NK cell count in fertility and unexplained infertility. Am. J. Reprod. Immunol. 70, 182 (2013).
Siewiera, J. et al. Human cytomegalovirus infection elicits new decidual natural killer cell effect functions. PLOS Pathog. https://doi.org/10.1371/annotation/94476def-de2e-4a6e-b0c2-2f2d2ccae784 (2013).
Sherkat, R., Meidani, M., Zarabian, H., Rezaei, A. & Gholamrezaei, A. Seropositivity of cytomegalovirus in patients with recurrent pregnancy loss. J. Res. Med. Sci. 19, S22–S25 (2014).
Lefaucheur, C. et al. Clinical relevance of preformed HLA donor-specific antibodies in kidney transplantation. Am. J. Transpl. 8, 324–331 (2008).
Geneugelijk, K. et al. A previous miscarriage and a previous successful pregnancy have a different impact on HLA antibody formation during a subsequent successful pregnancy. Front. Immunol. 7, 571 (2016).
Lashley, E. E., Meuleman, T. & Claas, F. H. Beneficial or harmful effect of antipaternal human leukocyte antibodies on pregnancy outcome? A systematic review and meta-analysis. Am. J. Reprod. Immunol. 70, 87–103 (2013).
Triulzi, D. J. et al. The effect of previous pregnancy and transfusion on HLA alloimmunization in blood donors: Implications for a transfusion-related acute lung injury risk reduction strategy. Transfusion 49, 1825–1835 (2009).
Dankers, M. K. et al. Differential immunogenicity of paternal HLA class I antigens in pregnant women. Hum. Immunol. 64, 600–606 (2003).
Larsen, E. C., Christiansen, O. B., Kolte, A. M. & Macklon, N. New insights into mechanisms behind miscarriage. BMC Med. 11, 154 (2013).
Meuleman, T. et al. Paternal HLA-C is a risk factor in unexplained recurrent miscarriage. Am. J. Reprod. Immunol. 79(2), e12797 (2018).
Thomas, K. A., Valenzuela, N. M. & Reed, E. F. The perfect storm: HLA antibodies, complement, FcyRs and endothelium in transplant rejection. Trends Mol. Med. 21(5), 319–329 (2015).
Jackson, A. M. et al. Evidence for induced expression of HLA class II on human islets: Possible mechanism for HLA sensitization in transplant recipients. Transplantation 87(4), 500–506 (2009).
Nagelkerke, S. Q. et al. The association and functional relevance of genetic variation in low-to-medium-affinity Fc-gamma receptors with clinical platelet transfusion refractoriness. J. Thromb. Haemost. 18(8), 2047–2053 (2020).
Mahaweni, N. M. et al. A comprehensive overview of FCGR3A gene variability by full-length gene sequencing including the identification of V158F polymorphism. Sci. Rep. 8, 15983 (2018).
Acknowledgements
We gratefully acknowledge Stefan Molenbroeck for helping with genotyping the p.V176F region of the FCGR3A gene in the women with RPL. In addition, we sincerely thank Jaqueline Frijns, Dorien Logister and Coline van Groesen for helping with anti-HLA antibody screening in the women with RPL. Last, Fig. 1 of this manuscript was created with the aid of BioRender.
Funding
This work was supported by Academic Incentive Maastricht 2016.
Author information
Authors and Affiliations
Contributions
Conceptualization: D.H.J.H., L.W. Methodology: D.H.J.H., S.Q.N., C.E. M.V., L.W. Formal analysis and investigation: D.H.J.H., S.Q.N., L.W. Writing: original draft preparation: D.H.J.H., L.W. Writing: review and editing: S.A.N., S.Q.N., C.E.M.V., T.W.K., M.E.A.S., L.W. Funding acquisition: S.A.N. Supervision: L.W. All authors have read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Habets, D.H.J., Al-Nasiry, S., Nagelkerke, S.Q. et al. Analysis of FCGR3A-p.176Val variants in women with recurrent pregnancy loss and the association with CD16a expression and anti-HLA antibody status. Sci Rep 13, 5232 (2023). https://doi.org/10.1038/s41598-023-32156-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-023-32156-9
- Springer Nature Limited