
Abstract
Endoglucanase (EC 3.2.1.4) catalysing the hydrolysis of β-1.4-glycosidic linkage of cellulose molecules is an enzyme of tremendous industrial importance. The present study describes a response surface methodology based predicted model to deduce a set of fermentation conditions for optimum growth and activity of recombinant endoglucanase in E. coli BL21 (DE3). Numerous significant parameters including fermentation media composition, temperature (Celsius), pH and agitation rate (rpm) were analysed systemically by employing central composite design. This effort reports highly efficient recombinant endoglucanase overproduction (6.9 gl−1 of biomass) with 30% expression by E. coli in modified M9NG media incubated at 37 °C and pH 7 agitated at 200 rpm. Addition of 3 mM glucose and 24 mM glycerol in the M9NG media has shown positive effect on the enzyme yield and activity. The CMCase activity experimentally estimated was found to be 1185 U/mg with the optimized parameters. The outcomes of both the responses by the predicted quadratic model were found in consensus with the obtained values. Our results well depicted the favourable conditions to further scale-up the volumetric yield of other relevant recombinant enzymes and proteins.
Similar content being viewed by others


Introduction
Cellulases with distinctive and manifold applications in a diverse array of fields, hold the third highest position in the global commercial enzymes market1. The demand of cellulases has been escalating day by day in a way that, to cope up with its mandate is a tangible challenge in recent years. High efficacy of recombinant microbes also results in higher productivity and expression of recombinant enzymes, ensuring their availability and utilization at an extreme level2. The production enzymes with desired characteristics is imperative to encounter the limitations of naturally produced enzymes3. Optimum bacterial growth, superlative production and top-notch expression of recombinant enzymes are significantly influenced by various fermentation parameters. Nature of production host organism and selection of plasmid vector is crucial to the expression of a particular enzyme4. Genetic manipulation for the production of heterologous proteins in E. coli has been progressively improved. Achieving a fast growth rate up to one generation per twenty minutes, under optimized conditions makes it an attractive procedure5,6. As compared to other hosts, competent cells preparation is relatively less complicated and transformation of foreign plasmid DNA in E. coli is eminently successful and efficient7,8. High yield of heterologous proteins can be achieved by amalgamation of simple and complex media components ranging from simple carbohydrates to complex peptones, tryptones, yeast extract and so forth. Optimization of media composition aids in attaining high productivity with cost reduction. Similarly, the selection of a media-microbe match can also enhance the yield9,10. A strategical approach for the induction of recombinant cellulase included the supplementation of attributed, economical and non-toxic inducers. Lactose fits best as an inducer according to the above mentioned properties11. Lactose being metabolized as a carbon source by the cells for optimal growth, although results in maximum biomass but ultimately becomes inadequate for the expression of recombinant proteins, consequently costs at prolonged induction time. Reinforcing the lactose supplementation with glycerol or glucose may relieves the protracted induction time. However, lactose and IPTG have their own merits and demerits as inducers12,13. Recombinant production as well as activity of enzymes diminutions when fluctuating from the optimum temperature and pH. Bacteria have an ability to survive in an array of environmental conditions including extreme temperatures, acidic or alkaline pH levels14.
Aerobic fermentation processes demand provision of oxygen at the rate to sufficiently satisfy the growth need of organism. Aeration is the most important and critical factor for the efficiency of fermentation process and is achieved by supplying the bacterial culture with adequate efflux of oxygen to the fermentation broth15. Proper aeration leads to saturation of dissolved oxygen in bacterial cell culture, resulting in an enhanced bacterial growth and production of cellulase up to many folds in comparison to non-agitated or static cultures16. Selection of E. coli preferred codon and introduction of single codon silent mutation in the primer sequence aids to achieve lower structural stability leading towards higher expression17. On the basis of above background, current study intended to achieve overproduction and overexpression of recombinant cellulase by employing certain molecular engineering techniques and optimization of fermentation parameters by response surface methodology. Optimization of growth media, supplementing with enriched nutrients, and fermentation crucial parameters will help to develop cost-effective media to achieve higher productivity with reduced induction time for the industrial benefits.
Results
Fermentation conditions optimization using response surface methodology (RSM)
Significant fermentation parameters including fermentation temperature, pH and agitation rate were optimized by varying the levels of these factors using central composite design for achieving higher cell growth with maximum CMCase activity. Table 2 depicted the obtained values of the responses i.e. cell growth and CMCase activity. Statistical testing of the model using analysis of variance (ANOVA) is presented in the Table 3A and B for cell growth and CMCase activity, respectively. The F-values of the response surface quadratic model were evaluated to be 56.56 and 30.19 for cell growth and CMCase activity, respectively, which implies that the model is significant. Fit statistical values well augmented the ANOVA. The determination coefficient value (R2) was estimated to be 0.9826 and 0.9679, for growth and activity, respectively, that in accordance with the insignificant lack of fit (F-value 0.25) depicted that the predicted quadratic model was able to explain 99.23% of the results. The predicted determination coefficient (R2) of 0.8287 and 0.9002 was in rational agreement with the adjusted R2 of 0.9653 and 0.9359, for growth and activity respectively, i.e. acquiring the difference of less than 0.2, indicated that the predicted model was unable to explain merely approximately 1% of the variations. These estimations confirmed that the predicted quadratic models were significant (p < 0.0001) adequately suitable for predicting both the responses i.e. cell growth and CMCase activity as well as the relationship between the response and the independent variables. Moreover, the quadratic factors (A2, B2 and C2) showed significance at p < 0.0001.
To investigate the relationship between dependent and independent variables and for determination of their optimum levels, 2D contour plots as well as 3D response surface plot were generated by plotting one of the responses against any two independent variables whereas the third variable was maintained at its middle (0) levels. Figure 1(A) representing the interaction effect between fermentation temperature and pH on CMCase activity at constant agitation rate (200 rpm) revealed that higher activity was achieved at temperature higher than 35 °C but lowered than 45 °C corresponding to the neutral pH. At pH towards basic environment with the lower temperature caused negative effect on the activity. Figure 1(B) showing the probable interactive effect of pH and agitation rate at constant temperature (37 °C) on the activity depicted that increasing agitation rate above 200 rpm increasing or decreasing pH. This fluctuations of pH does not show any significant increase in the activity rather acidic and basic fermentation environment leads to decline in the activity irrespective of agitation rate. Figure 1(C) displaying the effect on temperature and agitation rate showed insignificant relation on the activity as lowered and extremely high temperatures caused decline in the activity irrespective of increase in the agitation rate.

2D contour plots and 3D response surface plot for recombinant cellulase production depicting interactive effects between (A) temperature and pH, (B) pH and agitation rate and (C) temperature and agitation rate. Design expert version 11 software used from web site: www.statease.com.
PCR amplification and cloning in expression vector
Various combinations of silent mutations at initial codons were examined to minimize the translation-hindering stable secondary structures at 5′ of mRNA that has been reported to negatively affect the protein expression. ΔG values were estimated from RBS (ribosomal binding site) of mRNA to + 10 codon. Out of all possible combinations examined (Table 1), silent mutation at second codon (GTA > GTT) with ΔG − 2.80 was found to have promising influence on the translation as compared to the native sequence having ΔG − 4.0. Purified plasmid containing recombinant cellulase gene (celA-BC) was amplified through polymerase chain reaction and PCR product analysed by 1% agarose gel electrophoresis, showed successful amplification with respect to the expected size i.e. 1.6 kb. The amplified product was purified followed by the double restriction and ligation of the restricted gene and pET22b(+). The analysis of the clones verified the successful cloning into the expression vector, pET22b(+) and formation of recombinant vector of 7.1 kb.
Validation of RSM through shake flask fermentation
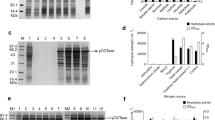
To augment the bacterial growth and production of recombinant cellulase, numerous fermentation parameters including culture medium, temperature, pH and agitation rate were optimized during shake flask fermentation. Recombinant E. coli was best fermented in modified M9NG media when auto-induced with 10 mM lactose overnight that produced biomass concentration of 6.9 gl−1 with 30% expression of recombinant cellulase. Bacterial growth obtained from IPTG induction in modified M9NG medium was 2.5 gl−1 with 30% expression and was plateaued after 4 h of induction. Therefore, it indicated that both the inducers in modified M9NG media are proficient for aggrandized production of recombinant cellulase. However, bacterial growth obtained in LB media was much lowered (3.5 gl−1) that resulted in least expression i.e. 18%. Expression analysis with 12% SDS-PAGE confirmed the expression of all the induced samples at the band corresponding to their respective molecular weight (61 kDa) against un-induced sample as a negative control. Confirmation of expressed protein as cellulase was also done by zymography by using CMC as substrate. Zymogram also presented recombinant cellulase expressed at its corresponding molecular weight i.e. 61 kDa (Fig. 2). Moreover, activity of recombinant cellulase against CMC was estimated to be 1185 U/mg which was in reasonable agreement with the predicted activity by the model.

SDS-PAGE expression analysis for recombinant cellulase. M: Protein marker (BioLabs-P7706L), Lane U, U′: Un-induced samples, Lane I, I′: Samples with IPTG induction, Lane L, L′: Lactose induction, Lane Z, Z′: Zymogram of recombinant cellulase against CMC; in modified M9NG and LB media respectively.
Apposite culture media for enhanced recombinant cellulase production
Recombinant E. coli fermented in modified M9NG media yielded the higher biomass concentration of 6.9 gl−1 with 30% expression of recombinant cellulase (Fig. 3A). However, the enzyme was not pronouncedly expressed in LB media over the 16 h of fermentation. Lowered production and expression in LB media specified that solely complex media components viz. tryptone and yeast extract are not sufficient for the higher production and expression of recombinant cellulase in short time period. Modified M9NG media supplemented with 3 mM glucose and 24 mM glycerol along with resting salts and nutrients resulted in better production and expression in possible least induction time.

(A) Time course of recombinant cellulase fermentation by E. coli BL21 (DE3) in LB (□) and modified M9NG media (♦) over 16 h of shake flask fermentation. Effect of (B) temperature, (C) pH and (D) agitation rate on recombinant cellulase fermentation by E. coli BL21 (DE3) determined by incubating at different fermentation temperature, pH and agitation rates respectively. Wet cell weight (♦), recombinant cellulase expression (□).
Optimum temperature for enhanced recombinant cellulase production
A trend of optimal growth at mesophilic temperature was observed for recombinant cellulase during the present study. Greater expression (30%) corresponding to higher biomass (6.9 gl−1) was achieved when incubated at 37 °C (Fig. 3B). At low temperature, the cellulase expression was minimal due to less microbial growth and the cells underwent death phase at the high temperature.
Optimum pH for enhanced recombinant cellulase production
Highest biomass concentration (6.8 gl−1) was yielded when fermentation run was carried out in pH 7, that superlatively expressed recombinant cellulase with 29% expression. Beyond this limit in either way, the recombinant cellulase gradually lost the expression however, acidic pH adversely affected the fermentation resulted in null expression (Fig. 3C).
Optimum agitation rate for enhanced recombinant cellulase production in E. coli
Limiting oxygen supply caused by inadequate stirring is the major bottleneck in high cell density fermentation run. Higher yield (6.9 gl−1) was obtained from the culture of E. coli BL21 (DE3) when fermented with moderate agitation rate i.e. 200 rpm sufficient for the surface aeration, dissolved oxygen concentration and oxygen supply to the site of oxidative phosphorylation in the cell. Increasing agitation speed did not exhibited pronounced effect on the production (Fig. 3D).
Discussion
Escherichia coli is a widely used advantageous host cell for the heterologous gene18,19. Several factors have been reported to control the bacterial growth conditions and yield of recombinant proteins. These factors include the concentration, pH, temperature of medium, and composition of various salts/additives, plasmid number, IPTG concentration. E. coli has an ability to show growth in culture even at lag phase of growth20,21. We have previously reported the cloning, purification and characterization of endoglucanase from Clostridium thermocellum22. The present study describes our experiments for the scaling-up of fermentation conditions to improve the yield and quality of endoglucanase by exploiting various influencing factors.
Response surface methodology (RSM), is a bunch of multivariant statistical, and mathematical tools for optimization of the culture conditions and bacterial response23. In the first step, the effect of agitation (rpm), temperature and pH of culture was determined. Maximum bacterial biomass, and CMCase activity was found at pH 7, 37 °C and 200 rpm (Table 2). The quadratic model for cell growth optimized in the present study has shown significant improvement in the bacterial biomass and enzyme activity (Table 3A, B). Large F-values with corresponding small p values (i.e. < 0.0001) for both the responses highly signifies the respected coefficients as generally depicted24. The effect of independent variables has been described by 2D contour plots as well as 3D response surface plot. In the literature, these plots have been applied to provide a clear idea about the impact of variables25,26. It indicates that a temperature above 35 °C but lowered than 45 °C has positive effect on enzyme activity at neutral pH (Fig. 1A–C). Similar findings have been reported in the literature22. Different silent mutations were examined at initial codons to minimize the reported translation-hindering stable secondary structures at 5′ of mRNA27. The binding affinities (ΔG values) of native and mutated codons were determined by Mfold web server28,29. The mutation GTA > GTT has shown a promoting impact on mRNA binding with the ribosomes30. The best yield of active enzyme was obtained in modified M9NG media under 10 mM lactose. It has given 6.9 gl−1 biomass and about 30% expression level. Similar expression level was obtained by 0.5 mM IPTG, however the final biomass was restricted to 2.5 gl−1. On the other hand LB media could generate 3.5 gl−1 biomass with only 18% expression. The expressed enzyme gave a band at 61 kDa on SDS-PAGE (Fig. 2). Lactose based induction has been already reported for its merits in recombinant protein synthesis31. It can also promote the soluble fraction of proteins. Recently, the strains with high lactose intake abilities have been introduced in for the optimization of experiments with better yield32,33. The enzyme activity with CMC was found 1185 U/mL which is similar to the model predicted activity. In our studies, the enriched M9NG media with 3 mM Glucose and 24 mM glycerol gave highest yield in the shortest time (Fig. 3A). Glucose and glycerol are in limelight for researchers desiring to achieve higher volumetric yield with reduced induction time for a wide range of bacterial recombinant proteins34. The optimum enzyme yield was obtained at mesophilic growth temperatures (Fig. 3B). According to the literature, for the production of various recombinant enzymes, E. coli BL21 (DE3) promotes higher yield and expression when fermented at around 37 °C of incubation temperature35,36. Moreover, higher temperatures corresponded to the lowered growth with least expression predominantly due the reduced glucose uptake rate and lowered saturated oxygen concentration37. Owing to the extreme stress and metabolic burden imposed onto the bacterial cells by the induction and growth at higher temperatures, fermentation resulted in decreased production and expression38. Moreover, deficient oxygen solubility at higher temperature may responsible for the alteration of TCA-cycle’s gene expression ultimately resulted in heavily accumulation of organic acid viz. acetic acid consequently occasioned in lowest yield and expression39,40. Optimum bacterial biomass and endoglucanase activity was found at neutral pH. Most probably, owing to the neutrophilic nature of E. coli, neutral pH was found to be quintessential environment for the excessive bacterial growth with top-notch expression i.e. likewise reckoned by previous researches41,42. Fluctuation of proton electrochemical gradient or imbalanced extra-intracellular pH that foist huge stress on the cells and affects growth and production can be considered as main factor43. Acidic pH corresponded to the least cell growth with negligible expression primarily due the cytoplasmic pH acidification below the threshold level and consequent hindrance to the cellular function44. Therefore, lowered pH of the medium attributed to the lowered growth rate of E. coli subsequently resulted in lowered growth and production of recombinant cellulase. The agitation rate of 200 rpm gave maximum yield, there was negative effect of further increase in agitation. On the other hand, static fermentation accountable for oxygen downshift resulted in lowered biomass and production. Owing to the inequity in rapid glucose uptake and restricted oxygen solubility in the cell culture due to the high viscosity of the culture medium43, a representative conflict between raised oxygen demand by the facultative cells at exponential phase and low level of dissolved oxygen may ascribed to the metabolism overflow and subsequent high glycolytic flux that adversely affected the bacterial growth and enzymatic production. Agitation greater than moderate rate is not recommended due to the possible sheering effects on the cells45. The present study signifies the predicted quadratic model using central composite design for achieving overproduction with maximum activity of recombinant cellulase. The study enlightened the importance of numerous fermentation parameters and their influence on the bacterial growth and activity. The interaction effect of temperature and pH on the CMCase activity was found to be highly significant. The recombinant cellulase was superlatively fermented in modified M9NG media when incubated at 37 °C, pH 7.0 with the agitation rate of 200 rpm.
Conclusions
The RSM model developed in the present study along with the selected media and additives provides better yield and activity of endoglucanase. Optimization of these fermentation parameters will help to enhance the volumetric production of recombinant cellulase in cost-effective media in least time during batch fermentation.
Methods
Predicted model for fermentation conditions optimization using response surface methodology (RSM)
Response surface methodology was designed to achieve highest culture density of E. coli BL21 (DE3) with maximum recombinant cellulase activity using a set of fermentation conditions. Various factors including fermentation temperature (°C), pH and agitation rate (rpm) were investigated systemically using central composite design (CCD) (Design Expert software, v11, Stat-Ease Inc, 11.1.2.0), known to be suited well to fit a quadratic surface for process optimization. Each numeric factor (temperature, pH and agitation rate) was set to five levels i.e. + α and − α (axial points), + 1 and − 1 (factorial points) and 0 (center point). The whole design comprised of 20 experimental runs including six replicates at the center of the design to estimate the pure error. The experiments were run in triplicate at each design points conducted in randomized order. Table 2 enlisted all the chosen ranges and levels of all the three factors whereby 0 (center point value) represented the frequently reported factor level in the literature. Moreover, low and high values of the factors were determined in accordance with the center point that remained as a middle value of the low and high ranges. Responses of all sets of experimental runs with respect to optimized design and conditions were estimated in terms of cell growth and CMCase activity using the Design Expert software. Analysis of variance (ANOVA) was examined for the statistical testing of the predicted model. F-value and p-value were analyzed to check the significance of the quadratic model.
Validation of the predicted quadratic model
For verification of the predicted model, successful recombinant cellulase production was carried out in E. coli BL21 (DE3) and overproduction with overexpression was achieved in modified M9NG media by employing the optimized conditions.
Bacterial strain and vector
Anaerobic cellulolytic, Clostridium thermocellum ATCC 27405 was used for endoglucanase gene having catalytic domain of celA (Genebank Ac. No. K03088) at C-terminus of a family 3a carbohydrate binding module from the scaffolding protein, cipA (Genbank Ac. No. L08665). Escherichia coli BL21 (DE3)—CodonPlus RIPL strain [E. coli B F– ompT hsdS (rB– mB–) dcm + Tetr gal λ (DE3) endA Hte (argU ileY proL leuW Camr)] from Novagen (Madison, Wis.) and pET22b (+) as an expression vector were used for the expression of recombinant cellulases.
PCR amplification and product analysis
PCR amplification was carried out by using the primers as described by Sajjad et al., 2012. However, silent mutation was introduced at the second codon (GTA → GTT) of the native sequence of the primer used for the cloning, previously12. The native primer having nucleotide sequence 5′ cacatatggtatcaggcaatttgaagg 3′ (ΔG − 4.0 kcal/mol) was replaced by a primer with above mentioned mutation i.e. 5′ cacatatggtttcaggcaatttgaagg 3′ (ΔG − 2.8 kcal/mol). PCR amplification conducted over 30 cycles underwent denaturation at 94 °C for 45 s, annealing at 58 °C for 45 s and extension at 72 °C for 90 s. Final extension was executed for 20 min at 72 °C. PCR product was checked on 1% agarose gel and DNA purification or gene cleaning was done with the help of MONARCH DNA Gel Extraction Kit (NEB #T1020) by following manufacturer’s instructions.
Cloning in expression vector pET22b (+)
Purified plasmid containing cellulase gene and pET22b (+) were restricted with SacI and NdeI restriction enzymes. Restriction of the plasmid was checked with the help of 1% agarose gel electrophoresis and desired DNA fragments were purified using MONARCH DNA Gel Extraction Kit Protocol (NEB #T1020) using the manufacturer’s guide. Purified restricted cellulase was then ligated into linearized pET22b (+). The ligation mixture was incubated on ice for 1 h and then at 20 °C for overnight. Competent cells of E. coli BL21 (DE3) were then transformed with this ligation mixture containing desired heterologous plasmid by conventional heat shock method. For the screening of transformants, the transformed cells were spread over the LB-agar plates (1.5% agar containing 50 μg ml−1 ampicillin) and was incubated at 37℃ overnight. A single colony was isolated and inoculated in 10 ml of LB medium and 20 ml modified M9NG media, both containing 100 µg ml−1 ampicillin. Both the flasks were incubated at 37 °C, 200 rpm on shaking incubator to grow overnight.
Recombinant cellulase production
Recombinant cellulase production was examined in LB and modified M9NG media in shake flask fermentation. Components of modified M9NG media included 1% tryptone, 0.5% yeast extract and 0.5% NaCl, along with resting salts and components i.e. 25 mM K2HPO4, 25 mM NaH2PO4, 15 mM NH4Cl, 8 mM MgSO4.7H2O, 1 mM Trace metals; each (CuSO4, MnCl2, CaCl2, MoO, FeCl3), 24 mM Glycerol, 3 mM Glucose and 10 mM Lactose. Two sets of 20 ml each of LB and modified M9NG media were prepared and inoculated with 20 µl of their respective overnight cultures, both supplemented with 100 μg ml−1 ampicillin and incubated at 37℃, 200 rpm on incubator shaker till OD600nm of 0.6–0.7 was attained. Expression of recombinant cellulase was induced, separately with 10 mM lactose and 0.5 mM IPTG and again incubated at 37℃, 200 rpm on incubator shaker for further 2–6 h. Samples were drawn after every 2 h of fermentation. In a parallel experiment, cells expressing recombinant cellulase were also auto-induced with 10 mM lactose in 20 ml of LB and modified M9NG media containing with 100 μg ml−1 ampicillin. The cultures were grown overnight (12–15 h of induction) at 37℃, 200 rpm. Samples were drawn after 4 h and 15 h (minimum and adequately maximum hours for auto-induction, respectively). After fermentation, the induced cells were centrifuged at 6500 × g for 10 min to be harvested. Supernatant was decanted off and air-dried cell pellets were resuspended in 20 mM Tris buffer; pH 7.0 to a final OD600nm of 10 and samples were prepared for conducting expression analysis and CMCase activity assay.
Optimization of fermentation conditions
Different fermentation parameters were optimized during the shake flask fermentation so as to operate fermentation at large scale with these optimized conditions in order to evade unsuccessful batches, complexities and time wastage. For determination of suitable and appropriate nourished growth media, recombinant cellulase containing culture was fermented in LB and modified M9NG to enhance the bacterial growth and production of enzyme. The culture was fermented at 28 °C, 32 °C, 37 °C, 40 °C and 42 °C to determine the optimum temperature. The pH was optimized over a broad range of pH ranging from 5.0 to 9.0. Optimum agitated rate for the optimum bacterial growth was also determined by fermenting the culture statically and agitated at 150, 200 and 250 rpm. Moreover, culture was auto-induced as well as induced on the basis of time/OD600nm over the period of time ranging from 4 to 16 h.
Expression analysis and CMCase activity assay for recombinant cellulase
Expression of recombinant cellulase in terms of total cell protein was analyzed on 12% SDS-polyacrylamide gel, stained with coomassie brilliant blue R-250 (Sigma Aldrich, #27816), by the standard electrophoresis protocol. Confirmation of the expression of recombinant cellulase was done by the zymographic analysis on CMC-agar plate. Zymography and CMCase activity assay was done according to the modified protocol as described by Sajjad et al., 2012.
Data availability
All the relevant data is available with the corresponding author.
Abbreviations
- RSM:
-
Response surface methodology
- CCD:
-
Central composite design
- RPM:
-
Revolution per minute
- LB:
-
Luria-Bertani
- RBS:
-
Ribosome binding site
- SDS-PAGE:
-
Sodium dodecyl sulphate
- CMC:
-
Carboxy methyl sephadex
- IPTG:
-
Isopropyl β-d-1-thiogalactopyranoside
References
Gupta, V. K. & Kumar, V. Microbial Enzymes in Bioconversions of Biomass 27–75 (Springer, 2016).
Jayasekara, S. & Ratnayake, R. Microbial cellulases: an overview and applications. Cellulose, 22 (2019).
Yang, H. et al. Molecular engineering of industrial enzymes: recent advances and future prospects. Appl. Microbiol. Biotechnol. 98, 23–29 (2014).
Sajjad, M. et al. Influence of positioning of carbohydrate binding module on the activity of endoglucanase CelA of Clostridium thermocellum. J. Biotechnol. 161, 206–212 (2012).
Maki, M., Leung, K. T. & Qin, W. The prospects of cellulase-producing bacteria for the bioconversion of lignocellulosic biomass. Int. J. Biol. Sci. 5, 500 (2009).
Nieß, A., Siemann-Herzberg, M. & Takors, R. Protein production in Escherichia coli is guided by the trade-off between intracellular substrate availability and energy cost. Microb. Cell Factor 18, 8 (2019).
Pope, B. & Kent, H. M. High efficiency 5 min transformation of Escherichia coli. Nucl. Acids Res. 24, 536–537 (1996).
Kent, R. & Dixon, N. Contemporary tools for regulating gene expression in bacteria. Trends Biotechnol. 38, 316–333 (2019).
Tuan, N. A. & Huong, N. T. Optimization of the fermentation medium to receive the highest biomass yield by Bacillus subtilis natto and the initial test of nattokinase yield. IOSR J. Eng. 4, 35–40 (2014).
Terol, G. L., Gallego-Jara, J., Martínez, R. A., Díaz, M. C. & de Diego Puente, T. Engineering protein production by rationally choosing a carbon and nitrogen source using E. coli BL21 (DE3) acetate metabolism knockout strains. Microb. Cell Factor. 18, 1–9 (2019).
Ismael Morales-Camacho, J., Domínguez-Domínguez, J. & Paredes-Lopez, O. Overexpression of a modified amaranth protein in Escherichia coli with minimal media and lactose as inducer. Recent Pat. Biotechnol. 7, 61–70 (2013).
Li, Z., Zhang, X. & Tan, T. Lactose-induced production of human soluble B lymphocyte stimulator (hsBLyS) in E. coli with different culture strategies. Biotechnol. Lett. 28, 477–483 (2006).
Zafar, A. et al. Cloning, purification and characterization of a highly thermostable amylase gene of Thermotoga petrophila into Escherichia coli. Appl. Biochem. Biotechnol. 178, 831–848 (2016).
Acharya, S. & Chaudhary, A. Bioprospecting thermophiles for cellulase production: a review. Brazil J. Microbiol. 43, 844–856 (2012).
Stanbury, P. F., Whitaker, A. & Hall, S. J. Principles of Fermentation Technology 243–272 (Elsevier, 2013).
Shahriarinour, M., Wahab, M. N., Ariff, A. B., Mohamad, R. & Mustafa, S. Kinetics of cellulase production by Aspergillus terreus at various levels of dissolved oxygen tension in a stirred tank bioreactor. Bio Resource 6, 4909–4921 (2011).
Rahmen, N. et al. A particular silent codon exchange in a recombinant gene greatly influences host cell metabolic activity. Microb. Cell Fact. 14, 156 (2015).
Mergulhão, F. J. M., Summers, D. K. & Monteiro, G. A. Recombinant protein secretion in Escherichia coli. Biotechnol. Adv. 23, 177–202 (2005).
Chen, R. Bacterial expression systems for recombinant protein production: E. coli and beyond. Biotechnol. Adv. 30, 1102–1107 (2012).
Zheng, S. et al. Optimization of medium components for plasmid production by recombinant E. coli DH5α pUK21CMVβ1.2. Biotechnol. Bioprocess. Eng. 12, 213 (2007).
Rosano, G. L. & Ceccarelli, E. A. Recombinant protein expression in Escherichia coli: advances and challenges. Front. Microbiol. 5, 172 (2014).
Kusuma, S. A. F. et al. Optimization of culture conditions for Mpt64 synthetic gene expression in Escherichia coli BL21 (DE3) using surface response methodology. Heliyon 5, e02741 (2019).
Zare, H., Sadeghi, H. M. M. & Akbari, V. Optimization of fermentation conditions for reteplase expression by Escherichia coli using response surface methodology. Avicenna J. Med. Biotechnol. 11, 162 (2019).
Deka, D. et al. Enhanced cellulase production from Bacillus subtilis by optimizing physical parameters for bioethanol production. ISRN Biotechnol. 2013, 1–11 (2013).
You, Z., Zhang, S., Liu, X. & Wang, Y. Enhancement of prodigiosin synthetase (PigC) production from recombinant Escherichia coli through optimization of induction strategy and media. Prep. Biochem. Biotechnol. 48, 226–233 (2018).
Lakshmi, E. S., Rao, M. N. & Sudhamani, M. Response surface methodology-artificial neural network based optimization and strain improvement of cellulase production by Streptomyces sp. Biosci. J. 36 (2020).
Khan, M. A., Sadaf, S. & Akhtar, M. W. Role of silent gene mutations in the expression of caprine growth hormone in Escherchia coli in the expression of caprine growth hormone in Escherichia coli. Biotechnol. Prog. 23, 1049–1052 (2007).
Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucl. Acids Res. 31, 3406–3415 (2003).
Mathews, D. H. RNA secondary structure analysis using RNA structure. Curr. Protoc. Bioinform. 46, 1–25 (2014).
Behzadipour, Y. & Hemmati, S. Considerations on the rational design of covalently conjugated cell-penetrating peptides (CPPs) for intracellular delivery of proteins: a guide to CPP selection using glucarpidase as the model cargo molecule. Molecules 24, 4318 (2019).
Dalla Venezia, N., Vincent, A., Marcel, V., Catez, F. & Diaz, J. J. Emerging role of eukaryote ribosomes in translational control. Int. J. Mol. Sci. 20, 1226 (2019).
Hausjell, J., Kutscha, R., Gesson, J. D., Reinisch, D. & Spadiut, O. The effects of lactose induction on a plasmid-free E. coli T7 expression system. Bioengineering (Basel) 7, 8 (2020).
Hausjell, J., Weissensteiner, J., Molitor, C., Halbwirth, H. & Spadiut, O. E. coli HMS174 (DE3) is a sustainable alternative to BL21 (DE3). Microb. Cell Factor. 17(1), 169 (2018).
Kopp, J. et al. Impact of glycerol as carbon source onto specific sugar and inducer uptake rates and inclusion body productivity in E. coli BL21 (DE3). Bioengineering (Basel) 5, 1 (2018).
Kishore, V., Nishita, K. P. & Manonmani, H. K. Cloning, expression and characterization of l-asparaginase from Pseudomonas fluorescens for large scale production in E. coli BL21. 3 Biotech 5, 975–981 (2015).
Duan, X., Hu, S., Qi, X., Gu, Z. & Wu, J. Optimal extracellular production of recombinant Bacillus circulans β-galactosidase in Escherichia coli BL21 (DE3). Process Biochem. 53, 17–24 (2017).
Soini, J. et al. Transient increase of ATP as a response to temperature up-shift inEscherichia coli. Microb. Cell Factor. 4, 9–23 (2005).
Harcum, S. W. & Fu’ad, T. H. Global transcriptome response of recombinant Escherichia coli to heat-shock and dual heat-shock recombinant protein induction. J. Ind. Microbiol. Biotechnol. 33, 801–814 (2006).
Shimizu, K. Regulation systems of bacteria such as Escherichia coli in response to nutrient limitation and environmental stresses. Metabolites 4, 1–35 (2013).
Leone, S., Sannino, F., Tutino, M. L., Parrilli, E. & Picone, D. Acetate: friend or foe? Efficient production of a sweet protein in Escherichia coli BL21 using acetate as a carbon source. Microb. Cell Factor. 14, 106 (2015).
Seneesrisakul, K., Guralp, S. A., Gulari, E. & Chavadej, S. Escherichia coli expressing endoglucanase gene from Thai higher termite bacteria for enzymatic and microbial hydrolysis of cellulosic materials. Electron. J. Biotechnol. 27, 70–79 (2017).
Amraini, S. Z. et al. Production and characterization of cellulase from E. coli EgRK2 recombinant based oil palm empty fruit bunch. Biotechnol. Bioprocess. Eng. 22, 287–295 (2017).
Farliahati, M. R., Mohamed, M. S., Rosfarizan, M., Tri Puspaningsih, N. N. & Ariff, A. B. Kinetics of xylanase fermentation by recombinant Escherichia coli DH5α in shake flask culture. J. Am. Biochem. Biotechnol. 5, 110–118 (2009).
Wilks, J. C. & Slonczewski, J. L. pH of the cytoplasm and periplasm of Escherichia coli: rapid measurement by green fluorescent protein fluorimetry. J. Bacteriol. 189, 5601–5607 (2003).
Fenice, M., Barghini, P., Selbmann, L. & Federici, F. Combined effects of agitation and aeration on the chitinolytic enzymes production by the Antarctic fungus Lecanicillium muscarium CCFEE 5003. Microb. Cell Factor. 11, 12–22 (2002).
Acknowledgements
This article was funded by the Deanship of Scientific Research (DSR) at King Abdulaziz University Jeddah. The authors, therefore, acknowledge and thank DSR for providing technical and financial support.
Funding
The research was funded by Deanship of Scientific Research at King Abdulaziz University Jeddah KSA.
Author information
Authors and Affiliations
Contributions
I.S. and M.S.N. contributed equally in this research. M.S.N., I.S. and M.S. conceived the research idea and made the study plan. I.S., M.S. and A.A. contributed in the lab experiments. I.S., M.A.A., M.S.N., J.A.K. and I.K. analysed data and prepared manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Shahzadi, I., Al-Ghamdi, M.A., Nadeem, M.S. et al. Scale-up fermentation of Escherichia coli for the production of recombinant endoglucanase from Clostridium thermocellum. Sci Rep 11, 7145 (2021). https://doi.org/10.1038/s41598-021-86000-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-021-86000-z
- Springer Nature Limited