
Abstract
Anthropogenic pollutants are known to have adverse effect on ecosystem, biodiversity and human health. Bioremediation is an option that has been widely used to remediate organic contaminants and reduce the risk of these hazardous materials. Microorganisms are readily available to screen and can be rapidly characterized to be applied in many extreme environmental conditions. Actinomycetes have a great potential for the production of bioactive secondary metabolites which have biodegradation activity. This study aimed to screen and characterize Nocardia species with biodegradation potential from diverse Iranian ecosystems. The isolates were screened from 90 collected environmental samples, identified and characterized using conventional and molecular microbiological methods including the PCR amplification and sequencing analysis of 16S rRNA and rpoB genetic markers. Growth rate in presence of pollutants, chromatography, Gibbs and turbidometric methods were used to determine bioremediation ability. A total of 19 Nocardia isolates were recovered from the cultured samples (21.1%) that belonged to 10 various species. The most prevalent Nocardia species was N. farcinica; 4 isolates (21%), followed by N. cyriacigeorgica and N. cashijiensis like; 3 isolates each (15.7%) and N. asteroides and N. kroppenstedtii; 2 isolates each (10.5%). Our results showed that various Nocardia species have great potential for bioremediation purposes, although they have not received much attention of the scholars for such significant usage.
Similar content being viewed by others


Introduction
Emerging contaminants due to pharmaceutical and personal care products, plasticizers, synthetic dyes, flame retardants, and pesticides are frequently detected in the environment we live in. In low- and middle-income countries, certain human activities are at the forefront of environmental pollution. Human made changes like overpopulation, pollution, burning fossil fuels, and deforestation have triggered climate change, soil erosion, poor air quality, and undrinkable water. These negative impacts can affect human behavior and can prompt mass migrations or battles over clean water1,2.
Removal of emerging contaminants by bioremediation that involves biosorption, bio-uptake, and biotransformation by plants and microorganisms offers numerous advantages such as low-cost and high removal efficiency over other cleanup methods. The key players in bioremediation are microorganisms that are existing everywhere and ideally suited to the task of contaminant destruction and the production of extracellular hydrolytic enzymes that allow them to use environmental contaminants to sustain growth and vital processes1,3.
The attractive, generally low-cost bioremediation approach using biodegradation capacity of microorganisms, primarily bacteria, have been considered as an ecological and economical alternative to physicochemical processes to eliminate diffusive contamination of persistent organic pollutants (POPs) in various environments, e.g. soil, sediments and sludge’s. High concentrations of these chemicals act as an environmental stress factor to develop efficient adaptation mechanisms in the adverse environment to metabolize these pollutants3,4,5. Understanding the microbial population and the processes that occur at contaminated sites would be a helpful hint to select and apply the most effective bioremediation method6,7. Therefore, the most important step in bioremediation process is isolation and identification of organisms capable of decontamination of pollutants.
Actinomycetes which includes a phylogenetically coherent group i.e., Corynebacterium, Rhodococcus, Nocardia, Gordonae, and Mycobacterium genera are reported to not only have a high intrinsic resistance to stressful condition but also have potential for degradation of environmental pollutants: members of Rhodococcus are able to degrade hydrocarbons, chlorophenols, polychlorinated biphenyls and sulfonated azo dyes8,9. Mycobacteria have shown ability to degrade polychlorophenols, heavy metals and diverse polycyclic aromatic hydrocarbons (PAH)1,9,10, Nocardia have the ability to decompose polycyclic aromatic hydrocarbons (PAH), polychlorinated biphenyls, chlorophenols, sulfonated azo dyes and alkanes11,12, and Gordoniae can break down alkanes13. Although some genera of this family such as Mycobacterium and Nocardia has a slower growth rate compared to other bioremedial species, yet they are excellent survivors of unfavorable conditions in contaminated environment. Hence, they can successfully compete with fast growing strains such Pseudomonas and related bacteria that are well-known for their ability in degradation of hazardous pollutants like aromatic compounds14.
Iran is a vast country in southwest Asia occupying an area of over 1.64 million square km. This includes an estimation of 8000 species of plants, 535 species of birds, 197 species of mammals and 870 species of fish that indicate biodiversity richness15. This great diversity of ecosystems leads to the diversity of microorganisms with distinct enzymatic capabilities. Due to a number of causes, however, Iran is exposed to rapid environmental and biodiversity degradation processes. There is heavy stress on and pollution of scarce environmental resources. Thus, screening and use of microbial species from this group of bacteria with bioremediation activity is a worthwhile effort that can contribute to the development and enhancement of bioremediation technology. The aim of current study was to screen, identify and characterize bioremediation capability of Nocardia as one of the most diverse species of actinomycetes with high catabolic capacity from Iranian environmental resources.
Materials and methods
Sampling and isolation
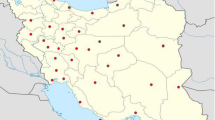
Form July 2015 to August 2017, a cross sectional study was carried out on a total number of 90 environmental samples collected from extreme natural and human-related environments such as sea, salt lake and rivers sediments, drinking and non-drinking water, factories and hospital wastewaters, agricultural soil and deserts, forests, oil wells and mines soil (The location and details of the sampling sites are indicated in Fig. 1 and Table 1. The samples were processed based on standard procedures. In summary, for aquatic samples, they were transported at 4 °C to the laboratory and processed within a maximum period of 24 h. The water samples were decontaminated with cetylpyridinium chloride (CPC) 0.005%, for 15 min, and filtered by vacuum through cellulose nitrate filters (0.45 µm, Sartorius, Gottingen, Germany). The filters were rinsed and macerated in tubes containing 15 ml of distilled water. Almost 100 µL aliquots of dissolved samples were transferred into Sauton’s medium, and incubated as described for the soils samples16.

Geographic distribution of sampling sites from Iranian ecosystems. The Figure source is obtained from Google maps and designed by Adobe Photoshop 2017 v18.1.1.252.
For soil samples, 15–30 g soil was taken from 3–5 cm depth of a sampling point at the polluted site and transferred directly to the laboratory. Five grams of soil was transferred to 50 ml sterile centrifuge tube. Then, 20 ml sterile distilled water was added to the tube and vortexed for 5 min, and centrifuged at 4300 RCF at room temperature for 20 min. The pellet and supernatant were decontaminated in separate tubes by 3% sodium lauryl sulfate and 1% NaOH. Afterwards, 100 µL of the decontaminated sample was used to inoculate into the Sauton’s medium which was supplemented with antifungal and antibacterial antibiotics including kanamycin, nystatin and nalidixic acid (each at 50 µg/ml) and were incubated at 25 °C, 30 °C and 35 °C with 5% CO2 atmosphere for 3 weeks17.
For sediment samples, up to 3 g of samples stirred for 30 min in 100 ml of sterile ringer’s solution (5% v/v). Tenfold serial dilutions of each homogenized suspension was prepared and 200 μl of each of the pretreated 10−2, 10−3, and 10−4 dilutions were inoculated into the Sauton’s medium which was supplemented with antifungal and antibacterial antibiotics including nystatin, kanamycin and nalidixic acid (each at 50 µg/ml). The samples were incubated for 3 weeks at 25 °C, 32 °C and 37 °C with 5% CO2 atmosphere18.
The details of environmental samples tested during this study are given in Table 2.
Conventional identification
The isolates were initially characterized by conventional phenotypic tests including partial acid-fast staining, growth at 25 °C, 37 °C and 42 °C, pigment production and standard biochemical assays, including resistance to hydrolysis of tyrosine, lysozyme, xanthine, and hypoxanthine tests19. The identification was further pursued by molecular testing as follows.
Molecular identification
Chromosomal DNA from Nocardia isolates was extracted using a simple boiling method. In brief, few colonies of bacteria added into 200 ml of TE buffer (Tris EDTA), boiled for 30 min and centrifuged at 10,000 rpm for 10 min. The supernatant was transferred to sterile microtube and centrifuged at 13,000 rpm for 10 min. Precipitated DNA was re-suspended in 50 µl of Milli-Q water and stored at − 20 °C20.
The environmental isolates identified phenotypically as Nocardia were further verified to the genus level using a specific PCR protocol based on a 596-bp region of the 16S rRNA as recommended by Laurent21. For species identification, amplification and direct sequence analysis of 16S rRNA gene was carried out as described by Roth22. Sequencing was performed in Bioneer Company (South Korea). The obtained sequences in the current study were aligned manually with all existing sequences of the closely related microorganism retrieved from GenBank database, compared with the relevant sequences and analyzed using the jPhydit program version 1.023.
Nucleotide sequence accession numbers
The GenBank accession number for the 16S rRNA sequencing of isolated Nocardia in this study are listed below., N.cyriacigeorgica (KX685347), N. coublieae like (KX685350), N. asteroides (KX685345), N. fluminea like (KX685346), N. otitidiscavarum (KX685341), N. kroppenstedii like (KX685348), N. cashijiensis like (KT372140.2), N. sungurluensis like (KT372141.1).
Bioremediation analysis
The sample collection locations have been polluted with crude oil and its derivatives such as PAHs (Polycyclic Aromatic Hydrocarbon), phenol and sodium sulfate. With regard to widespread existence of petroleum resource all around Iran, petrochemical and its derivatives pollutions are the most common environmental pollutants in different region of Iran and have been found in agricultural soil and water, hospital and industrial wastewater and sewages24,25,26. The bioremediation capacity of isolates for these pollutants was evaluated according to Kanali et al.27 as follow:
Media preparation
To evaluate the bacterial growth in presence of PAH, phenol and sodium sulfate, 100 ml aliquots of Mineral Salt Medium (MSM) were prepared in 250 ml flask as the base media for this experiment. The MSM medium containing (7H20. 0.25-MgSO4, 0.5-KH2PO4, 0.5-K2HPO4, 1-NaCl, 0.009-CaCl2.2H2O, 0.5-KNO3, 0.1-Mn Cl2.4H2O, 0.07-ZnCl2, 0.015-CuCl2.2H2O, 0.025-Ni Cl2.6H2O, 0.12- COCl2.6H2O, 0.025-Na2MO4.2H2O (g/l). The MSM medium then mixed with different substrates as follow.
The media supplemented with 1% PAH mix solution (1–1) purchased from AccuStandard. The PAH mix solution content the following ingredients each at a concentration of 0.2 mg/ml dissolved in dichloromethane and methanol: Acenaphthene, Acenaphthylene, Anthracene, Benzo(b)fluoranthene, 1,2-Benzanthracene, Benzo (a)pyrene, Benzo (k)fluoranthene, Benzo(g,h,i)perylene, Chrysene, Dibenz(a,h)anthracene, Fluoranthene, Fluorene, Indeno(1,2,3-cd) pyrene, Phenanthrene, Naphthalene, Pyrene.
Another set of MSM medium were supplemented with 1% phenol (Merck, Germany) dissolved in deionized water, and the third MSM medium set were supplemented 1% sodium sulfate (Merck, Germany) dissolved in deionized water.
An amount of 1 ml of 0.5 McFarland turbidity (1 × 108 CFU ml−1) of the selected isolates was prepared in normal saline, and inoculated into culture media, incubated for 144 h at 30 °C in an orbital shaker (3 RCF). To evaluate the bacterial growth in presence of PAH, phenol or sodium sulfate, samples were collected at 24-h intervals and the optical density of samples measured at a wavelength of 560 nm by spectrophotometer.
Showing sign of growth in the media indicated decomposition and/or consumption of target material by the studied isolates. For final confirmation of degradation, an amount of 5 ml of medium was removed from the flask and examined for the PAH, phenol and sodium sulfate degradation yield according to standard procedures28,29,30 explained as follow.
Determination of PAHs phenol and sodium sulfate degradation
After growth, 5 ml of the MSM medium supplemented with PAH was transferred to a screw cap glass tube and supplemented with 0.6 ml of a mixture of tetrachlorethylene and methanol (1:100) as the extraction solvent, vortexed for 10 s, centrifuged at 3000 rpm for 10 min. The organic phase then collected and transferred to a clean tube for further analysis by HPLC. The amount of PAH was measured by injecting 100 µl of the collected organic phase into the HPLC device (Manager 5000, Knauer, Germany), equipped with C18 ultra-sep ES PAH QC specia 6o × 2 mm ID, with water and acetonitrile as mobile phase at a ratio of 5:95, and 0.3 ml min−1 flow rate. The adsorption at 254 nm was measured and the PAH content of the sample was calculated using the standard curve and function previously calculated using sterile PAH standards28.
Gibbs method was used for determination of phenolic compound29. In brief, 5 ml of MSM medium containing phenol which showed bacterial growth was transferred to a sterile tube and the pH was adjusted at 8.0, subjected to centrifugation at 2700g for 20 min. 150 µl of the collected supernatant was mixed with 30 µl NaHCO3, and 20 µl of Gibbs reagent (2, 6-Dichloroquinone 4-chloroimide) was added to the mixture and shaked for 30 min at 25 °C. The absorbance at 620 nm was recorded and the phenol content determined using the standard curve previously calculated using sterile standards.
From the MSM medium supplemented with sodium sulfate that showed bacterial growth, an amount of 5 ml sample was taken and transferred to a sterile tube. One ml acetic acid (1%) and 1 ml of acetate buffer were added and mixed for 3 min. Afterwards, 1 ml of barium chloride was added and mixed for another 3 min. The turbidity was calculated using spectrophotometry at a wavelength of 420 nm. The amount of consumed sodium sulfate was measured by using the standard curve previously calculated using sterile sodium sulfate standards.
Experiments were conducted in duplicate and average values were computed. The Michaelis-Menten equation was used to compute the in-solution tested materials bio sorption efficiency for each material by each isolate; the results were expressed in percentage terms.
Results
Isolation and characterization of Nocardia strains
The recorded temperature and pH of the soil samples were in the range of 6 °C to 32 °C and 6.2 to 7.8 respectively. For wastewater and sediment samples, these measures were in the range of 5 °C to 28 °C and 6.8 to 8.2, respectively. The corresponding figures for water samples were 4 °C to 29 °C and 6.4 to 8.0, respectively, and the total dissolved solids (TDS) for the water samples ranged between 560 to 1340 mg/L. The details of soil, water, wastewater and sediment samples and the isolates key properties are presented in Table 1.
From 90, wastewater, sediment, soil, and water samples 19 isolates were identified as Nocardia based on phenotypic and biochemical properties, including partial acid-fast staining, pigmentation, resistance to lysozyme and decomposition of xanthine and tyrosine. The presence of a 16S rRNA amplicon of 596 bp in size was a signature band for Nocardia species (Table 2).
The 16S rRNA gene sequencing of the isolates revealed that all isolates had nucleotide signatures of Nocardia at positions 70–98 (A-T), 293–304 (G-T), 307 (C), 328 (T), 614–626 (A-T), 631(G), 661–744 (G-C), 824e876 (T-A), 825–875 (A-T), 843 (C), and 1122–1151 (A-T)31.
Based on phenotypic and molecular data, the Nocardia isolates in this study belonged to 8 validated species and 2 unknown or potentially novel species.
The most prevalent Nocardia species isolated in our study were N. farcinica; 4 isolates (21%), N. cyriacigeorgica and an unknown Nocardia species closely related to N. cashijiensis; 3 isolates (15.7%) each, N. asteroides and N. kroppenstedtii 2 isolates (10.5%) each. Five single isolates belonged to four established species that is N. coublieae, N. carne, N. fluminea and N. otitidiscavarum, as well as one unknown Nocardia species closely related to N. sungurluensis.
The almost complete 16S rRNA gene sequences showed that the isolate AN7 had 99.5% similarities (corresponding to 6 base pair nucleotide differences) with N. coubleae type strain DQ235688T, the isolates AN21, AN20 and AN30 showed 98.92% similarities with N. cashijiensis type strain JCM 11508T (corresponding to 8 base pair nucleotide differences) and the isolate AN8 showed 99.25% similarities (corresponding to 7 base pair nucleotide differences) with N. sungurluensis type strain CR3272T. This degree of nucleotide differences between our isolates and the established aforementioned Nocardia species make them the candidate for further inquiry as the novel species.
The relationship between our isolates and the standard established species of Nocardia was supported by a high bootstrap value in phylogenetic tree based on 16S rRNA gene depicted by MEGA8. (Fig. 2).

16S rRNA sequence based phylogenetic tree for Iranian biodegrading Nocardia isolates and nearest validated species of Nocardia depicted by applying MEGA8 software using the neighbor-joining method with bootstrapping values 1000. The figures at each node represent bootstrapping values. The tree was rooted with G. terrae.
Bioremediation analysis
With regard to the previous studies the isolated strains were classified into two categories based on the bioremediation ability of the isolates:
-
1.
The isolates that showed bioremediation ability and had been reported previously (Table 1). These included AN4, AN26, AN27 and AN73 which were identified as N. farcinica, with capability to degradation of paraffin wax, crude oil and phenol32,33. The Isolates AN18, AN37 and AN47 were identified as N. cyriacigeorgica. This species has been reported for degradation capacity of PAHs and thymol34,35. The isolates AN31and AN32 were identified as N. asteroides. This species has been reported for degradation capacity of the crude oil, rubber, polyethylene, sodium benzoate36,37,38. The isolates AN7 was identified as N. coubleae. This species has been reported for degradation capacity of PAHs and crude oil degradation39. The Isolate AN35 identified as N. carnea. This species has been reported for degradation capacity of the lewisite40. The Isolate AN5 was identified as N. otitidiscaviarum This species has been reported for degradation capacity of PAHs and crude oil41.
-
2.
This group included the strains with unreported bioremediation capacity, that is, the isolates, AN4, AN26, AN27and AN73 which were identified as N. farcinica, the isolate AN22 which was identified as N. fluminea, AN4, AN26, AN27and AN73 which were identified as N. farcinica, the isolates AN3, AN19 which were identified as N. kroppenstedtii, the isolates AN20, AN21 and AN30 which were identified as N. cashijiensis, and the isolate AN8 that was identified as N. sungurluensis, In other words for the first time their bioremediation capacity, These strains were tested for PAHs, phenol and sodium sulfate biodegradation capacity according the standard procedures and the following results were obtained.
Determination of PAHs, phenol and sodium sulfate degradation
In order to determine PAHs degradation capacity of the selected isolates, the growth and growth rate of the isolates were evaluated in MSM medium in presence of PAHs by measuring optical density at 560 nm in the 24 h intervals.
As shown in Table 2 the results indicate that, the isolates AN4, AN26, AN27and AN73 which were identified as N. farcinica, the isolates AN3, AN19 which were identified as N. kroppenstedtii and the isolate AN22 which was identified as N. fluminea, were able to consume PAHs compound as an energy and carbon source. These strains were able to degrade one or several compound of the PAH mix. The results also showed that the isolate AN22 which was identified as N. fluminea has the highest PAHs degradation rates by degrading more than 90% of the PAHs in the culture medium, followed by the isolates AN4, AN26, AN27 and AN73 were identified as N. farcinica and the isolates AN3, AN19 were identified as N. kroppenstedtii with 80% and 70% degrading rate of PAHs respectively (Fig. 3). The other isolates, were not able to degrade PAHs.

Growth curves of Iranian Nocardia isolates over a 24 h. Incubation period at 30 °C in the presence of PAHs. C control sample.
For final confirmation, the degradation yield and the type of PAH compounds consumed (degraded) by the studied isolates were analyzed by HPLC. The result showed that the isolates AN4, AN26, AN27, AN73, AN3, AN19 and AN22 can degrade different type of PAHs and convert the compounds to other ingredients (Fig. 4).

HPLC chromatograms of PAHs mix solution by selected Nocardia isolates, (A) control samples, (B) after 144 h incubation at 30 °C. 1. Naphthalene 2. Acenaphthylene 3. Acenaphthene 4. Fluorene, 5. Phenanthrene, 6. Anthracene, 7. Fluoranthene 8. Pyrene 9. Benzo[a] Anthracene 10. Chrysene 11. Benzo[b]fluoranthene 12. Benzo[k]fluoranthene 13. Benzo[a]pyrene 14. Indeno [1, 2, 3-cd] pyrene 15. Dibenzo [a, h] anthracene.
The phenol biodegradation ability of the isolates was investigated through showing sign of growth in MSM medium supplemented with phenol and by measuring absorbance at 560 nm for samples collected at 24 h intervals.
The results showed that isolates AN4, AN26, AN27 and AN73 which were identified as N. farcinica, the isolates AN3 and AN19 which were identified as N. kroppenstedtii, the isolates AN20, AN21 and AN30 which were identified as N. cashijiensis like and the isolate AN8 that was identified as N. sungurluensis like, had ability to grow in the presence of phenol and degrade this compound (Fig. 5).

Growth curves of Iranian Nocardia isolates over a 24 h. Incubation period at 30 °C in the presence of phenol. C control samples.
The results of phenol consumption by the selected isolates showed that isolates AN3 and AN19 that were identified as N. kroppenstedtii had the highest phenol degradation rates (95% of phenol degradation in media after 144 h), followed by the N. sungurluensis like isolate AN8, N. farcinica isolates AN4, AN26, AN27 and AN73 and N. cashijiensis isolates AN20, AN21 and AN30 with phenol degradation rate of 85%, 80% and 70% respectively. The other tested isolates were not able to grow significantly in present of phenol in culture media.
Sodium sulfate biodegradation ability of the isolates were investigated by checking for the sign of growth in MSM medium supplemented with sodium sulfate and measuring the optical density at 560 nm. Additionally, in case of growth, the growth rate was evaluated by measuring the cell density via absorbance at 560 nm for samples harvested at 24 h intervals.
The results showed that N. cashijiensis like isolates AN20, AN21 and AN30 and N. sungurluensis like isolate AN8 had the ability to degrade sodium sulfate (Fig. 6).

Growth curves of Iranian Nocardia isolates over a 24 h. Incubation period at 30 °C in the presence of sodium sulfate. C control samples.
The amount of sodium sulfate consumption by the selected isolates was measured by turbidimetry. The results indicated that N. cashijiensis isolates AN20, AN21 and AN30 showed highest sodium sulfate biodegradation capability and was able to degrade 100% of the sodium sulfate followed by N. sungurluensis isolate AN8 that was able to degrade 85% of the sodium sulfate in prepared medium.
Discussion
A knowledge of degradation potentials of bacterial strains isolated from polluted sites is pivotal in designing and development of an enduring bioremediation strategy1,2. The most of previous studies has focused on isolation of actinomycetes from normal environments which is easily accessible and actinomycetes from these environment can readily be isolated to determine their biological potential. However this is not the case with special environments and actinomycetes that inhabit special environments42,43. These special environments are unique due to the extreme conditions that is prevent easy access to them and subsequently actinomycetes from these environment are largely unexplored8,9.
Isolation of Nocardia species for implementation in bioremediation processes from environmental sources have been a worldwide theme of research44,45,46. In current study, due to their high catabolic capacity as well as high durability in harsh environments such as ecosystems with industrial, domestic and hospital pollution, we studied the process and optimization methods for both isolation and characterization of Nocardia species from environmental sources especially those that exist in pristine and untouched ecosystems, as well as the bioremediation capability of the isolates.
In the current study we isolated and characterized 19 biodegrading Nocardia strains from 90 samples collected from environmental resources. The isolates belonged to 8 validated species and 2 unknown or potentially novel species. N. farcinica was the most encountered isolates. N. farcinica was the first historically identified Nocardia species and based on the result of current study and previous studies has the ability to degrade the paraffin wax, crude oil and phenol32,33. N. cyriacigeorgica ranked second that included 16% of the isolates, this organism that was first isolated in 2001 from clinical specimens47 has been reported to be able to degrade the PAHs and thymol34,35.
In our study N. asteroides and N. kroppenstedtii ranked the third of the isolates. N. asteroides has been reported to have the capacity to degrade the crude oil, rubber, polyethylene and sodium benzoate36,37,38. N. kroppenstedtii is an opportunistic pathogen that was first isolated and characterized in 2014 from a patient with a pulmonary infection48. However there was no reports on its bioremediation ability. We showed that N. kroppenstedtii has the capability to degrade phenol and PAHs.
In the current study, five species that is, N. coubleae, N. carnea, N. otitidiscavarum, N. fluminea and an unknown potentially novel Nocardia species closely related to N. sungurluensis consisted the single isolates. These species were isolated from special ecosystems of Iran including sulfate sodium salt lake, seashore of Persian Gulf, petrochemical factory and farm land around oil well soil respectively.
Nocardia coubleae first isolated and characterized in 2007 from oil contaminated soil and determined that has capacity to degrade the PAHs and crude oil39. N. carnea first characterized in 1913 from clinical samples, and based on the result of previous studies by Nakamiya, determined that this Nocardia species has the ability to degrade the lewisite (is an organoarsenic compound use as a chemical weapon, acting as a vesicant and lung irritant)40. N. otitidiscavarum first isolated and characterized in 1924 from clinical samples49. This Nocardia species is ubiquitous in environmental resources and has been frequently isolated from the clinical and environmental samples41,50. In subsequent studies it was found that N. otitidiscavarum has the capacity to degrade PAHs and crude oil41. N. fluminea first isolated and characterized in 2000 from soil and water51, however we found no reports on its bioremediation ability. We showed that this species has the capability to degrade phenol and PAHs in prepared media.
In our study four isolates AN20, AN21 and AN30 and AN8 found to have molecular and phenotypic characteristics different from all previously established Nocardia based on 16S rRNA, hsp65, and rpoB nucleotide sequences. These isolates were evaluated for bioremediation activity against PAHs, phenol and sodium sulfate according to standard procedure, and it was determined that the isolates AN21, AN20 and AN30 had a capability to consume and degrade phenol and sodium sulfate compound as a sole carbon and energy source and the isolate AN8 had a capability to degrade sodium sulfate and PAHs. The characterization of these unknown Nocardia species remains to be completed using a thorough phenotypic and molecular analysis including cell wall composition analysis and whole genome sequencing.
Conclusion
Efforts made in the current study have been successful and are supported by evidence from recent reports on the isolation and characterization of novel actinomycetes from poorly researched habitats. Therefore it can be assumed that the screening of untapped regions that possess unexplored actinomycetes will increase the chances of discovering new chemical compounds to be used as a resource for bioremediation significance.
From a biotechnological perspective, the current study showed that the diverse Iranian ecosystems and in particular the nonconventional environments provides vital habitat for actinomycetes and notably Nocardia species with capacity for degradation of organic pollutants. This confirms the idea that despite being abundant in environment, Nocardia species have been simply ignored for such significant usage. Indeed, there is an untapped potential with regard to bioremedial actinomycetes particularly Nocardia that has yet to be discovered and administered in bioremediation of hazardous chemicals.
Normal environments which is easily accessible and actinomycetes isolated from these environment could readily be tapped so that we can analyzed this bacterial species to determine their biological potential. However this is not the case with special environments and actinomycetes that inhabit in these special environments. These special environments are unique due to the extreme conditions that is prevent easy access to them and subsequently actinomycetes from these environment are largely. Thus the group of bacteria from these nascent and unknown habitats are yet to be explored and deserve a special mention.
References
Kumar, A., Bisht, B., Joshi, V. & Dhewa, T. Review on bioremediation of polluted environment: A management tool. Int. J. Environ. Sci. 1, 1079 (2011).
KENSA, V. M. Bioremediation—an overview. I Control Pollution 2011 (2015).
de Lorenzo, V. & Loza-Tavera, H. Microbial bioremediation of chemical pollutants: How bacteria cope with multi-stress environmental scenarios. Bacter. Stress Resps. 2, 481–492 (2010).
Storz, G. & Hengge-Aronis, R. Bacter stress resps. Vol. 791 (Wiley Online Library, 2011).
Zorádová, S., Dudášová, H., Lukáčová, L., Dercová, K. & Čertík, M. The effect of polychlorinated biphenyls (PCBs) on the membrane lipids of Pseudomonas stutzeri. Intern. Biodeter. Biodegrad. 65, 1019–1023 (2011).
Tyagi, M., da Fonseca, M. M. R. & de Carvalho, C. C. Bioaugmentation and biostimulation strategies to improve the effectiveness of bioremediation processes. Biodegradation 22, 231–241 (2011).
Alam, A. & Sharma, V. Environmental biotechnology—a review. Researcher [Internet] 5, 71–93 (2013).
Kuyukina, M. S. & Ivshina, I. B. Biology of Rhodococcus 231–262 (Springer, Berlin, 2010).
Xun, L. Microb Degrad Xenob 1–30 (Springer, Berlin, 2012).
Girma, G. Microbial bioremediation of some heavy metals in soils: an updated review. Ind. J. Sci. Res. 6, 147 (2015).
11Biswas, K. Biological agents of bioremediation: a concise review. (2015).
Thapa, B. & Ghimire, A. A review on bioremediation of petroleum hydrocarbon contaminants in soil. Kathmandu Univ. J. Sci. Eng. Technol. 8, 164–170 (2012).
Liu, P.-W.G. et al. Bioremediation of petroleum hydrocarbon contaminated soil: Effects of strategies and microbial community shift. Intern. Biodeter. Biodegrad. 65, 1119–1127 (2011).
Das, K. & Mukherjee, A. K. Crude petroleum-oil biodegradation efficiency of Bacillus subtilis and Pseudomonas aeruginosa strains isolated from a petroleum-oil contaminated soil from North-East India. Biores. Technol. 98, 1339–1345 (2007).
Farashi, A. M. S. Biodiversity hotspots and conservation gaps in Iran. J. Nat. Conserv. 39, 37–57 (2017).
Thomson, R., Carter, R., Gilpin, C., Coulter, C. & Hargreaves, M. Comparison of methods for processing drinking water samples for the isolation of Mycobacterium avium and Mycobacterium intracellulare. Appl. Environ. Microbiol. 74, 3094–3098 (2008).
Kamala, T., Paramasivan, C., Herbert, D., Venkatesan, P. & Prabhakar, R. Evaluation of procedures for isolation of nontuberculous mycobacteria from soil and water. Appl. Environ. Microbiol. 60, 1021–1024 (1994).
Arifuzzaman, M., Khatun, M. & Rahman, H. Isolation and screening of actinomycetes from Sundarbans soil for antibacterial activity. Afr. J. Biotechnol. 9, 4615–4619 (2010).
Saubolle, M. A. & Sussland, D. Nocardiosis review of clinical and laboratory experience. J. Clin. Microbiol. 41, 4497–4501 (2003).
Rahdar, H. A., Azadi, D. & Shojaei, H. Molecular analysis and species diversity of Nocardia in hospital environment of a developing country, a potential health hazard. J. Med. Microbiol. 2, 2 (2017).
Laurent, F. J., Provost, F. & Boiron, P. Rapid identification of clinically relevant Nocardia species to genus level by 16S rRNA gene PCR. J. Clin. Microbiol. 38, 2805 (2000).
Roth, A., Andrees, S., Kroppenstedt, R. M., Harmsen, D. & Mauch, H. Phylogeny of the genus Nocardia based on reassessed 16S rRNA gene sequences reveals underspeciation and division of strains classified asNocardia asteroides into three established species and two unnamed taxons. J. Clin. Microbiol. 41, 851–856 (2003).
Jeon, Y.-S. et al. jPHYDIT: A JAVA-based integrated environment for molecular phylogeny of ribosomal RNA sequences. Bioinformatics 21, 3171–3173 (2005).
Valipour, M., Mousavi, S. M., Valipour, R. & Rezaei, E. Air, water, and soil pollution study in industrial units using environmental flow diagram. J. Basic Appl. Sci. Res. 2, 12365–12372 (2012).
Bazrafshan, E., Mostafapour, F. K. & Mansourian, H. J. Phenolic compounds: Health effects and its removal from aqueous environments by low cost adsorbents. Health Scope 2, 65–66 (2013).
Jalali, M. Geochemistry characterization of groundwater in an agricultural area of Razan, Hamadan Iran. Environ. Geol. 56, 1479–1488 (2009).
Kanaly, R. A. & Harayama, S. Biodegradation of high-molecular-weight polycyclic aromatic hydrocarbons by bacteria. J. Bacteriol. 182, 2059–2067 (2000).
Manoli, E. & Samara, C. Polycyclic aromatic hydrocarbons in waste waters and sewage sludge: Extraction and clean-up for HPLC analysis with fluorescence detection. Chromatographia 43, 135–142 (1996).
Gibbs, H. Phenol tests III. The indophenol test. J. Biol. Chem. 72, 649–664 (1927).
Chesnin, L. & Yien, C. Turbidimetric determination of available sulfates. Soil Sci. Soc. Am. J. 15, 149–151 (1951).
Stackebrandt, E., Rainey, F. A. & Ward-Rainey, N. L. Proposal for a new hierarchic classification system, Actinobacteria classis nov. Intern. J. Syst. Evol. Microbiol. 47, 479–491 (1997).
Patel, D. D. & Lakshmi, B. Study on the role of Nocardia farcinica in enhancing the flow rate of crude oil. Bioremed. J. 20, 224–232 (2016).
Hristov, A., Christova, N., Nacheva, L., Stoineva, I. & Kabaivanova, L. biodegradation potential of N farcinica BN26 for xenebiotic mineralization. Comp Rendus De L Acad Bulgare Des. Sci. 67, 821–830 (2014).
Hahn, V., Sünwoldt, K., Mikolasch, A. & Schauer, F. Two different primary oxidation mechanisms during biotransformation of thymol by gram-positive bacteria of the genera Nocardia and Mycobacterium. Appl. Microbiol. Biotechnol. 97, 1289–1297 (2013).
Ma, J. et al. Isolation and characterization of oil-degrading microorganisms for bench-scale evaluations of autochthonous bioaugmentation for soil remediation. Water Air Soil Pollut. 226, 272 (2015).
Khan, Z. et al. Nocardia asteroides in the soil of Kuwait. Mycopathologia 137, 159–163 (1997).
Tsuchii, A., Takeda, K. & Tokiwa, Y. Degradation of the rubber in truck tires by a strain of Nocardia. Biodegrad 7, 405–413 (1996).
Bonhomme, S. et al. Environmental biodegradation of polyethylene. Polym Degrad Stab. 81, 441–452 (2003).
Rodríguez-Nava, V. et al. Nocardia coubleae sp nov, isolated from oil-contaminated Kuwaiti soil. Intern. J. Syst. Evol. Microbiol. 57, 1482–1486 (2007).
Nakamiya, K., Nakayama, T., Ito, H., Shibata, Y. & Morita, M. Isolation and properties of a 2-chlorovinylarsonic acid-degrading microorganism. J. Hazard Mater. 165, 388–393 (2009).
Zeinali, M., Vossoughi, M., Ardestani, S. K., Babanezhad, E. & Masoumian, M. Hydrocarbon degradation by thermophilic Nocardia otitidiscaviarum strain TSH1: Physiological aspects. J. Bas Microbiol. 47, 534–539 (2007).
Adams, G. O., Fufeyin, P. T., Okoro, S. E. & Ehinomen, I. Bioremediation, biostimulation and bioaugmention: A review. Intern. J. Environ. Bioremed. Biodegrad. 3, 28–39 (2015).
Allard, A.-S. & Neilson, A. H. Bioremediation of organic waste sites: A critical review of microbiological aspects. Intern. Biodeteriorat. Biodegrad. 39, 253–285 (1997).
Megharaj, M., Ramakrishnan, B., Venkateswarlu, K., Sethunathan, N. & Naidu, R. Bioremediation approaches for organic pollutants: a critical perspective. Environ. Int. 37, 1362–1375 (2011).
Gomes, E. B., Soriano, A. U., Miranda, R. C. M., Sousa, M. F. & Pereira, N. Biodegradation of stored jet fuel by a Nocardia sp. isolated from contaminated soil. Braz. Arch Biol. Technol. 52, 1279–1284 (2009).
Milić, J. S. et al. Bioremediation of soil heavily contaminated with crude oil and its products: Composition of the microbial consortium. J. Serb. Chem. Soc. 74, 455–460 (2009).
Yassin, A., Rainey, F. & Steiner, U. Nocardia cyriacigeorgici sp. nov.. Int. J. Syst. Evol. Microbiol. 51, 1419–1423 (2001).
Jones, A. L. et al. Nocardia kroppenstedtii sp. nov., an actinomycete isolated from a lung transplant patient with a pulmonary infection. Int. J. Syst. Evol. Microbiol. 64, 751–754 (2014).
Clark, N. M., Braun, D. K., Pasternak, A. & Chenoweth, C. E. Primary cutaneous Nocardia otitidiscaviarum infection: case report and review. Clin. Infect. Dis. 20, 1266–1270 (1995).
Aghamirian, M. R. & Ghiasian, S. A. Isolation and characterization of medically important aerobic actinomycetes in soil of Iran (2006–2007). Open Microbiol. J. 3, 2 (2009).
Maldonado, L., Hookey, J. V., Ward, A. C. & Goodfellow, M. The Nocardia salmonicida clade, including descriptions of Nocardia cummidelens sp. nov., Nocardia fluminea sp. nov. and Nocardia soli sp. nov.. Antonie Van Leeuwenhoek 78, 367–377 (2000).
Acknowledgements
The authors are grateful to office of vice-chancellor for Research of Isfahan University of Medical Sciences for the support of the current study (Grant number: 393405).
Funding
This study was funded by Isfahan University of Medical Sciences (Grant number: 393405).
Author information
Authors and Affiliations
Contributions
D.A. Is the first author of this article completed experiment, conducted data analysis and write the manuscript. H.S.H. is the corresponding author and designed this study, conducted data analysis and write the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Azadi, D., Shojaei, H. Biodegradation of polycyclic aromatic hydrocarbons, phenol and sodium sulfate by Nocardia species isolated and characterized from Iranian ecosystems. Sci Rep 10, 21860 (2020). https://doi.org/10.1038/s41598-020-78821-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-78821-1
- Springer Nature Limited
This article is cited by
-
Exploring macrophytes’ microbial populations dynamics to enhance bioremediation in constructed wetlands for industrial pollutants removal in sustainable wastewater treatment
Symbiosis (2024)
-
Purple acid phosphatase 10c modifies the rice rhizobacterial community and its phosphorus cycling potential
Plant and Soil (2024)
-
Bioremediation of petroleum refinery wastewater using Bacillus subtilis IH-1 and assessment of its toxicity
Archives of Microbiology (2024)
-
Poly-cis-isoprene Degradation by Nocardia sp. BSTN01 Isolated from Industrial Waste
Applied Biochemistry and Biotechnology (2022)
-
Biosurfactant, a green and effective solution for bioremediation of petroleum hydrocarbons in the aquatic environment
Discover Water (2022)