
Abstract
Creativity pervades many areas of everyday life and is considered highly relevant in several human living domains. Previous literature suggests that the posterior parietal cortex (PPC) is related to creativity. However, none of previous studies have compared the effect of transcranial random noise stimulation (tRNS) over bilateral PPC on both verbal and visual divergent thinking (DT) and Remote Associates Test (RAT) in the same experimental design. Forty healthy participants were randomly assigned to tRNS (100–500 Hz) over bilateral PPC or sham group, for 15 min and current was set at 1.5 mA. Participants’ creativity skills were assessed before and after brain stimulation with the Unusual Uses and the Picture Completion subtests from the Torrance Test of Creative Thinking and the RAT. ANCOVA (baseline scores as covariate) results indicated that tRNS group had significantly higher scores at post-test in RAT and visual originality compared to sham group. Unusual Uses, on the other hand, was not significant. Improvement in RAT suggests the involvement of PPC during via insight solution which may reflect internally directed attention that helps the recombination of remotely associated information. The improvement in visual originality dimension from DT may be due to a higher internally directed attention while reducing externally oriented attention.
Similar content being viewed by others


Introduction
Creativity is considered one of the most important and prevalent human accomplishments1 but incredibly difficult to measure and define. However, in the scientific literature there is a relative agreement that creative solutions can be defined as both potentially novel/original and useful/effective2,3,4. According to several authors, creative thinking includes two main components1,5,6, including convergent thinking (CT) and divergent thinking (DT). CT involves finding a single solution to a closed-ended problem by using deductive reasoning7. According to many authors8,9,10,11,12,13,14,15,16,17,18,19,20 the most used instrument for CT has been the Remote Associates Test (RAT)21. Although RAT seems to measure mainly CT, there is also evidence that it may be reflecting DT, at least to some extent22.
DT, on the other hand, is usually defined as the ability to form remote associations between unconnected ideas from distant categories and produce multiple alternative and original responses to an open-ended problem6,23. In this context, originality is argued to contain both novelty and authenticity24 and the research during idea generation activates an exploratory DT process25.
Given the importance of creativity, the interest from a neuroscientific perspective has increased during the last years26,27. In this context, several studies have used neuroimaging and transcranial stimulation techniques in order to investigate the neural underpinnings of creative thinking, including both CT and DT. According to a meta-analysis of transcranial stimulation28, most of these studies have targeted dorsolateral prefrontal cortex (DLPFC) and frontotemporal areas8,29,30,31,32,33,34. Transcranial direct current stimulation (tDCS)7,31,34 and transcranial random noise stimulation (tRNS)35 over the DLPFC have consistently shown an improvement in RAT.
The literature regarding DT, on the other hand, has shown more inconsistent results after frontotemporal stimulation. Anodal tDCS over the DLPFC30 and prefrontal cortex32 has been related to higher verbal DT. In contrast, various previous studies have shown a significant general verbal DT improvement (but not originality) after cathodal (but not anodal) tDCS8,36 and transcranial magnetic stimulation29 applied to the left inferior frontal gyrus (IFG). Similarly, 10 Hz Transcranial Alternating Current Stimulation (tACS) over the DLPFC37,38 has been linked to an improvement in verbal DT (more precisely with ideational verbal fluency but not in originality)38 and a composite of visual DT (but not originality)37. Finally, Chi and Snyder39,40 showed that the inhibition of the anterior temporal lobe produced an enhancement in insight problem-solving tasks (matchstick arithmetic problems and the nine dot problem).
Even though there is not a completely consistent pattern of results, all these previous studies suggest that both DLPFC and fronto-temporal areas of the brain are more related to both verbal CT and DT (especially to verbal fluency dimension from DT), but not so much to originality.
On the other hand, the role of parietal cortex has also been related to creativity by several neuroimaging25,41,42,43,44,45,46,47,48,49,50 and non-invasive brain stimulation studies1,7,51,52, based mainly on its role as part of the default-mode network50. One of the most replicated findings in the literature is the strong synchronization of EEG alpha activity over PPC during performance on mainly verbal DT tasks25,46,48,53,54.Although less studied, EEG alpha power over PPC has also been related to visual DT55. Since the parietal alpha synchronization is suggested to reflect a higher internally directed attention while reducing externally oriented attention56, according to previous authors57 it may indicate that “divergent thinkers are better able to continually exclude interfering external stimuli for the sake of generating creative responses”. Additional evidence for the role of PPC on DT comes from studies using functional magnetic resonance imaging (fMRI) also suggests that PPC is related to DT41,58.
A closer look at these studies on DT suggests that PPC involvement in creativity may be more related to the originality dimension of DT rather than fluency45,46,59,60. On the contrary, as previously mentioned, prefrontal areas seem to be more related to fluency dimension of DT8,32,37,38.
Similar to the relationship between DT and PPC, RAT or Compound Remote Associates scores have also been related to alpha power in PPC studies when participants generated solutions with insight61,62. According to some authors25, a possible explanation for the finding of alpha power increase in PPC during insightful solutions may be interpreted as a suppression of sensory information, thus favoring the combination of remotely associated semantic information25. Studies that included fMRI63,64 suggest that along with frontal brain regions (including DLPFC, ventrolateral PFC, medial PFC and IFG), PPC was involved in Compound Remote Associates Task performance. There are only two studies that used tDCS over the PPC1,7 and their the results are in line with the evidence from neuroimaging studies. In the study carried out by Zmigrod et al.7 they found that anodal stimulation of both left and right PPC produced an increase of insight type solutions to RAT. However, the same study did not show a significant improvement in verbal DT. They did not include any visual DT task, so we cannot conclude if the lack of improvement found in verbal DT would be also true for visual DT. Similarly, Pick and Lavidor1 found an improvement in RAT after anodal bilateral stimulation of PPC. In another study using tDCS, Ghavanati et al.51 found that anodal stimulation over the right PPC produced more unique and novel figures than anodal stimulation over the DLPFC and sham groups in a figural fluency task. These unique and novel figures may be considered an originality measure, although it is not directly comparable to other originality tasks from visual DT (such as Torrance Test of Creative Thinking65) used in the neuroimaging studies previously mentioned.
However, as far as the authors are aware, none of the previous studies targeting the PPC has examined its role on visual DT tasks.
On the other hand, most previous studies that investigated creative thinking through transcranial stimulation have used tDCS. TRNS is a more recent form of transcranial electrical stimulation of random noise to modulate cortical plasticity, especially when using high frequencies66 even though the underlying mechanisms are not yet completely understood. There are two main suggested mechanisms of tRNS effects, including the increase of neuronal excitability via stochastic resonance (whereby weak neural signal detection in the central nervous system is enhanced when ‘noise’ is added67) and the repetitive opening of NaC channels68. Although more research is needed, previous studies have suggested some advantages that tRNS may show compared to tDCS and tACS. For example, the effect of tRNS may be larger than tDCS in visual perceptual learning tasks66 and have a greater long-term effect69,70. Moreover, tRNS has been proposed to be more tolerable than tDCS regarding possible adverse effects, such as irritation and burning71. Finally, a meta-analysis72 indicated that the effect of tRNS on language and mathematics was stronger than that of tDCS.
Therefore, based on previous studies that used tDCS over PPC1,7, our first hypothesis was that participants receiving tRNS over bilateral PPC would improve their CT scores (RAT) compared to sham.
Considering the impact of tDCS over PPC on figural creativity51 but not in verbal DT7, our second hypothesis was that tRNS would improve more in visual DT than in verbal DT. Finally, given that the literature suggests that prefrontal brain regions are more related to the fluency dimension of DT8,32,37,38 whereas PPC is more related to the originality dimension45,46,59,60, we expect to find a higher positive effect of tRNS over PPC on the originality dimension of DT compared to fluency.
Results
Baseline characteristics of tRNS and sham groups
Baseline characteristics of the groups are shown in Table 1. There were no significant differences in age, years of education, gender, number of hours slept, number of stimulants (tea, coffee or similar), and handedness (Edinburgh Handedness Inventory). We asked if the number of stimulant drinks ingested and number of hours slept was more than usual, less than usual or as usual at the time of assessment. There were no significant differences between groups in any of the proportion of these responses regarding stimulant drinks (X2 = 0.01, p = 0.924) and sleeping condition (X2 = 0.58, p = 0.748).
Effect of brain stimulation on convergent thinking, verbal divergent thinking and visual divergent thinking
Table 2 shows baseline and post-treatment scores for all creative sub-domains. In Table 3 we show marginal means (with baseline scores as covariates) of RAT, visual DT and verbal DT scores after brain stimulation. ANCOVA results indicated that there were significant differences between tRNS and sham groups in RAT, suggesting a higher number of correct responses after stimulation in the tRNS group compared to sham. Results regarding visual DT scores and verbal DT, on the other hand, were not statistically significant.
A closer inspection of the visual DT subdomains revealed that only visual originality was significant (F = 6.91, p = 0.012, \({\mathrm{n}}_{\mathrm{p}}^{2}\) = 0.157). In this subdomain, scores of the tRNS group increased from 1.40 (SD = 1.18) to 2.60 (SD = 1.42), whereas the sham group changed from 2.10 (SD = 1.07) to 1.70 (SD = 1.21).
Regarding verbal DT subdomains (data not shown), none of them were significant (F values from 0.00 to 2.37, p values from 0.966 to 0.132).
Adverse effects and blinding
None of the participants reported having experienced any significant adverse effects. There were not significant differences between real stimulation groups vs sham in the number of adverse effects assessed (F = 1.17, p = 0.285). However, the percentage of participants experiencing concentration problems was significantly higher in the experimental group compared to sham (See Table 4).
A Chi-square test was performed to test blinding of the stimulation based on participants´ report. In the stimulation group, 10.0% guessed that they had received stimulation, 25.0% placebo and 65.0% did not know. in the sham group, 10.0% guessed that they had received stimulation, 55.0% placebo, 35.0% and did not know. There were no significant differences in stimulation guess between sham and real tRNS conditions (χ2 = 4.05, p = 0.132).
Discussion
The present study investigated the effects of increasing cortical excitability with tRNS over left and right PPC on RAT, visual DT and verbal DT tasks. We found that tRNS produced a significant and large increase in RAT scores when compared to sham. This result is consistent with previous studies that used tDCS over PPC1,7 and with neuroimaging studies, including EEG61,62 and fMRI63,64. Zmigrod et al.7 found that anodal stimulation over right and anodal over left PPC produced a higher number of insight solutions to a Compound Remote Associates Task, a similar task to RAT used in the present study. Similarly, Pick and Lavidor reported a significant increase in RAT after anodal bilateral stimulation over PPC1. In the present study, participants only had 165 s for the whole task, which resulted in 5.5 s per item. According to various authors57,73, CRA problems may be solved in at least two ways; (1) via insight (also called the “Aha moment”) that appears suddenly and with little ability to report on the processing that enabled the solution and (2) via a deliberate, trial-and-error analytical approach. Given the time constriction, participants did not have much time to think about possible responses, so we believe that the main way to obtain the solution was insight type (AHA moment) rather than an analytical approach. This hypothesis goes in line with Kounios et al.´s findings74, who found less alpha over the posterior cortex related to non-insightful, analytical problem-solving. Authors74,75 suggest that this kind of problem-solving is characterized by directing attention outward in a bottom-up way whereas insight type solution is associated with focusing attention more inwardly. However, we cannot assure it was mainly via insight since we did not ask them to report how they had obtained the correct answer directly. We did not include another condition giving more time to find the solution for RAT. Therefore, we cannot know if the effect of tRNS over PPC would have disappeared or attenuated if they had been given the chance to use other different cognitive mechanisms to solve the problem. In this context, future transcranial stimulation studies could investigate if PPC vs prefrontal brain regions improve RAT scores through different cognitive mechanisms.
Our results regarding visual DT indicate that participants receiving tRNS over PPC significantly increased the visual originality dimension of DT (but not fluency of flexibility) showing a large effect size. As far as the authors are aware, there is no previous study that examined visual DT after stimulating bilateral PPC either with tRNS or tDCS, so direct comparisons are not possible. However, this result goes in line with the previously mentioned results on neuroimaging studies55,76 and tDCS over PPC on figural fluency51,52. In the tDCS studies, the authors compared anodal stimulation over the right PPC with anodal over the left DLPFC and sham. In their first study51, results indicated that participants produced significantly more unique designs under anodal right PPC tDCS compared to anodal left DLPFC and sham. Similarly, in a more recent study52, the same authors found very similar results regarding unique and novel figures in a figural fluency task, whereas left anodal DLPFC stimulation generated more verbal fluency scores.
Taking together both RAT and visual originality results, we could speculate that the alpha power effect could reflect more internally oriented attention53, which may reflect selective inhibition of interfering external input48,77,78. This internally-directed attention could therefore facilitate the combination of remotely associated semantic information25 and prevent functional fixedness during the creative ideation phase of creativity79.However, there are other possible explanations for the role of PPC, such as its role in efficient effective memory search and retrieval that could have a positive impact on these tasks53, working memory for object location80 or spatial representation and updating81.
Regarding verbal DT, we did not find any significant improvement after tRNS compared to sham. These results are also consistent with the lack of significant changes after tDCS over PPC reported by the only study that used tDCS over PPC7. In this study, the authors assessed fluency, flexibility and elaboration but not originality. The studies that reported significant changes after transcranial stimulation in verbal DT, on the other hand, have focused on more frontal regions8,30,32. The lack of significant results after stimulating PPC observed in this study and the previous one7, altogether with significant improvement after targeting more frontal areas7,8,30,32 suggest that PPC might not be as strongly related to verbal DT as frontal areas. The results were non-significant even for verbal originality, a dimension more closely related to PPC as shown in different neuroimaging studies45,46,59,60.
Despite these interesting results, the present study has several limitations. First, we did not compare tDCS directly with tRNS, so we cannot conclude if tRNS is superior or not. Second, we did not include additional groups targeting right and left PPC separately. Previous results7 found significant results on CT after both anodal left and anodal right PPC. However, we cannot rule out if there would have been differential effects on visual originality. Third, we did not include any measure of attention, memory or working memory, so we cannot rule out the possible mediating role of visual attention in the visual originality results obtained in the present study. Finally, although the highest electric-field strength was estimated to be under the PPC, adjacent areas may have also been receiving electrical stimulation so we cannot completely rule out the influence of adjacent areas on these results.
Future studies could directly compare if the stimulation of prefrontal brain regions increases fluency scores compared to a differential effect of PPC stimulation on originality scores. Regarding stimulation techniques, there is still much to research on the long-term effect of tRNS. Since most studies have used single session transcranial brain stimulation so far, future studies could also investigate if repetitive brain stimulation generates more pronounced and lasting effects. Additionally, future studies could also investigate if tRNS increases alpha activity or if the effects of 10 Hz tACS effects on alpha activity over PPC are related to creative enhancement.
Material and methods
Participants
A priori power analysis was conducted to determine the sample size using the G Power*3 software82. According to a previous study7, a sample size of 40 participants, 20 in each group, was enough to attain a large effect size (f = 0.54) to detect differences in a Compound Remote Associates task with 90% power and a 5% level of significance. Inclusion criteria included age of 18 years or above and both genders. Exclusion criteria included: (1) previous history of brain surgery; (2) being pregnant; (3) suffering from frequent or severe headaches or migraines; (4) previous history or presence of neurological disorder or injury (epileptic or convulsive seizure, brain stroke, severe brain injury; and (5) presence of any type of metallic implant in the brain.
Participants did not receive any financial compensation for participating in the study. Ethical approval was obtained from the Deusto University Ethics Committee (Ref: ETK-31/17-18).
All volunteers provided written informed consent to participate in the study and all experiments were performed in accordance with the tenets of the Declaration of Helsinki and they were free to withdraw at any time. Once the study was finished, participants allocated to the sham group were offered the opportunity to receive real brain stimulation.
Measures
The Edinburgh Handedness Inventory
We assessed handedness with the Edinburgh Handedness Inventory83. In this inventory participants are asked to indicate their preference of hand use for 10 everyday activities. Scores ranged from 100 (perfectly right-handed) to − 100 (perfectly left-handed).
The Remote Associates Test (RAT)
The Spanish version of the original RAT21 was administered in order to assess CT. This activity involves identifying a solution word that is associated with three cue words. The solution can be related to the three cue words either semantically or by forming a compound word. Different forms of the test were employed for the baseline evaluation and after intervention. Each form included 30 items. Participants were given 2 min and 45 s. The number of correct responses was recorded and scores ranged from 0 to 30. Internal consistency was high (Cronbach´s alpha = 0.81).
The Torrance Test of Creative Thinking (TTCT)
The Picture Completion and the Unusual Uses (UU) subtests from the TTCT65,84 were administered in order to assess visual DT and verbal DT, respectively. Two minutes were given to perform each task. Different forms of the test (Form A and B) were administered for the baseline and after the stimulation. The Picture Completion task requires the participants to complete ten unfinished figures by adding additional elements. For this study three dimensions were measured: fluency, originality, and flexibility. These dimensions were scored using the Torrance Test of Creative Thinking scoring manual84. Fluency was measured by the total number of figures completed, awarding 1 point to each figure completed. Originality was based on the statistical infrequency of each response. Responses were classified as original (1 point) or unoriginal (0 points) according to a list that had been developed for each item by Torrance84 on the basis of normative data. Flexibility was defined as the number of different ideational categories produced in the pictures. For the correction of this dimension, each figure was classified according to the corresponding category, using the list of categories from the Spanish adaptation of the Torrance Test of Creative Thinking85. One point was given for each different category used. Fluency, originality and flexibility were converted into z scores based on the pooled group and a visual DT composite was obtained, showing a satisfactory internal consistency (Cronbach´s alpha = 0.84).
In the UU task, participants were asked to write as many unusual uses as possible for an item. In Form A of the test, the stimulus was Cardboard Boxes, while in Form B Tin Cans was used. Three dimensions were measured: Fluency, Originality, and Flexibility. Fluency was based upon the number of different unusual uses generated, assigning 1 point to each unusual use. Originality was based on the statistical unusualness of each response. This dimension was scored using the list from the manual65,84, giving 1 point for original or uncommon responses, and 0 points for unoriginal responses. Flexibility score was obtained from the number of different categories represented in the responses, using the list of categories from the manual84, awarding 1 point to each different category. Fluency, originality, and flexibility were used to obtain a verbal DT composite (Cronbach´s alpha = 0.85).
Questionnaire of adverse effects
After each session, subjects filled out a questionnaire to assess any perceived side effects, which consisted of 11 items (including headache, throat sore, scalp pain, skin tingling, skin itching, skin burning sensation, redness of the skin, numbness, concentration problems, mood change and phosphenes).
Transcranial stimulation protocol
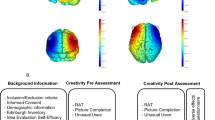
In the tRNS and sham groups, the electrodes were placed over the left and right PPC (P3 and P4, respectively). TRNS group received 1.5 mA (tRNS 100–500 Hz) via two saline-soaked (5 ml per sponge), 16 cm2 (8 × 8 cm2) circular sponges. They were attached under designated electrode positions (P3, P4) using a wireless neoprene cap that followed the International 10–20 system. Figure 1 (part A) shows the simulated electric field of this tRNS group (based on Stim Weaver) using the finite element model86. TRNS was applied with a light, battery-operated device (Neuroelectrics Inc., Barcelona) attached to the back of the neoprene cap, delivering electrical current for 15 min with additional ramp-up and ramp-down phases of 30 s. In the sham condition, current was applied using a 30 s ramp up followed 15 min after by a 30 s ramp-down of activity. The impedance of the electrodes was checked before and during tRNS application to guarantee that it was below 10 kΩ.

Study design and transcranial random noise stimulation (tRNS) montage.
Procedure
The study had a double-blind, sham-controlled, parallel-group design. The participants were randomly allocated to either the tRNS or the sham group (see Fig. 1, Part A). Assignment was conducted based on a computer-generated randomization (randomizer.org). All raters (JP, AS, and AA) were blind to treatment condition.
Figure 1 (part B) shows the study design and procedure. After signing the consent form, participants reported sociodemographic information along with hours of sleep, tobacco consumption, and stimulant drinks ingested before the session. Afterwards, the Edinburgh Handedness Inventory was administered. Baseline creativity assessment was carried out before starting stimulation (tRNS or sham). They have 2 min and 45 s to complete the RAT, 2 min for UU and 2 min Figure Completion.
Five minutes after the beginning of stimulation, they started performing the parallel versions of RAT, UU and Figure completion tests with the same time limitations. The order of RAT, UU, and Figure Completion versions were counterbalanced. Afterwards, participants completed the adverse effects questionnaire. Finally, in order to examine the blinding efficacy, participants were asked to answer the following question: “Please, tell us if you think you were receiving real stimulation, no stimulation (placebo) or you do not know”.
Statistical analyses
Categorical data were analyzed with the χ2 test. Baseline characteristics were compared using ANOVA. Analyses of covariance (ANCOVA) were performed independently for CT, visual DT and verbal DT with the post test scores as dependent variable and the baseline scores as covariate. Effect size (\({\mathrm{n}}_{\mathrm{p}}^{2}\)) was reported and according to Cohen87, an effect size of 0.01 is considered small, 0.06 medium, and 0.14 large. IBM SPSS Statistics Version 25 was used for all statistical analyses. The significance level was set at 0.05. All tests were two-tailed.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Pick, H. & Lavidor, M. Modulation of automatic and creative features of the Remote Associates Test by angular gyrus stimulation. Neuropsychologia 129, 348–356 (2019).
Sternberg, R. J. & Lubart, T. I. The concept of creativity: prospects and paradigms. In Handbook of Creativity (ed. Sternberg, R. J.) 3–15 (Cambridge University Press, Cambridge, 1999). https://doi.org/10.1017/CBO9780511807916.003.
Runco, M. A. Divergent thinking. In Encyclopedia of Creativity, Vol. 49 (eds Runco, M. A. & Pritzker, S. R.) 577–582 (Elsevier Academic Press, Amsterdam, 1999).
Sternberg, R. J. & Lubart, T. I. Investing in creativity. Am. Psychol. 51, 677–688 (1996).
Cropley, A. In praise of convergent thinking. Creat. Res. J. 18, 391–404 (2006).
Guilford, J. P. Creativity. Am. Psychol. https://doi.org/10.1037/h0063487 (1950).
Zmigrod, S., Colzato, L. S. & Hommel, B. Stimulating creativity: modulation of convergent and divergent thinking by transcranial direct current stimulation (tDCS). Creat. Res. J. 27, 353–360 (2015).
Mayseless, N. & Shamay-Tsoory, S. G. Enhancing verbal creativity: modulating creativity by altering the balance between right and left inferior frontal gyrus with tDCS. Neuroscience 291, 167–176 (2015).
Hommel, B., Colzato, L. S., Fischer, R. & Christoffels, I. K. Bilingualism and creativity: benefits in convergent thinking come with losses in divergent thinking. Front. Psychol. 2, 273 (2011).
Hoşgören Alici, Y., Devrimci Özgüven, H., Kale, E., Yenihayat, I. & Baskak, B. Prefrontal activity measured by functional near infrared spectroscopy during divergent and convergent thinking in bipolar disorder. Noropsikiyatri Ars 56, 86–91 (2019).
Zhu, W., Shang, S., Jiang, W., Pei, M. & Su, Y. Convergent thinking moderates the relationship between divergent thinking and scientific creativity. Creat. Res. J. 31, 320–328 (2019).
Ma, K. & Hommel, B. Metacontrol and body ownership: divergent thinking increases the virtual hand illusion. Psychol. Res. 84, 120–127 (2020).
Colzato, L. S., Szapora, A., Pannekoek, J. N. & Hommel, B. The impact of physical exercise on convergent and divergent thinking. Front. Hum. Neurosci. 7, 824 (2013).
Colzato, L. S., van den Wildenberg, W. P. M. & Hommel, B. Increasing self-other integration through divergent thinking. Psychon. Bull. Rev. 20, 1011–1016 (2013).
Dewhurst, S. A., Thorley, C., Hammond, E. R. & Ormerod, T. C. Convergent, but not divergent, thinking predicts susceptibility to associative memory illusions. Personal. Individ. Differ. 51, 73–76 (2011).
Chermahini, S. A. & Hommel, B. Creative mood swings: divergent and convergent thinking affect mood in opposite ways. Psychol. Res. 76, 634–640 (2012).
Chermahini, S. A. & Hommel, B. The (b)link between creativity and dopamine: spontaneous eye blink rates predict and dissociate divergent and convergent thinking. Cognition 115, 458–465 (2010).
Yamada, Y. & Nagai, M. Positive mood enhances divergent but not convergent thinking. Jpn. Psychol. Res. 57, 281–287 (2015).
Sellaro, R., Hommel, B., de Kwaadsteniet, E. W., van de Groep, S. & Colzato, L. S. Increasing interpersonal trust through divergent thinking. Front. Psychol. 5, 561 (2014).
Colzato, L. S., Szapora, A., Lippelt, D. & Hommel, B. Prior meditation practice modulates performance and strategy use in convergent- and divergent-thinking problems. Mindfulness (N. Y.) 7, 152–159 (2017).
Mednick, S. The associative basis of the creative process. Psychol. Rev. 69, 220–232 (1962).
Cortes, R. A., Weinberger, A. B., Daker, R. J. & Green, A. E. Re-examining prominent measures of divergent and convergent creativity. Curr. Opin. Behav. Sci. 27, 90–93 (2019).
Barbot, B. & Lubart, T. Creative thinking in music: its nature and assessment through musical exploratory behaviors. Psychol. Aesthet. Creat. Arts 6, 231–242 (2012).
Corazza, G. E. Potential originality and effectiveness: the dynamic definition of creativity. Creat. Res. J. 28, 258–267 (2016).
Agnoli, S., Zanon, M., Mastria, S., Avenanti, A. & Corazza, G. E. Predicting response originality through brain activity: an analysis of changes in EEG alpha power during the generation of alternative ideas. Neuroimage 107, 116385 (2020).
Beaty, R. E., Benedek, M., Silvia, P. J. & Schacter, D. L. Creative cognition and brain network dynamics. Trends Cogn. Sci. 20, 87–95 (2016).
Beaty, R. E., Seli, P. & Schacter, D. L. Network neuroscience of creative cognition: mapping cognitive mechanisms and individual differences in the creative brain. Curr. Opin. Behav. Sci. 27, 22–30 (2019).
Lucchiari, C., Sala, P. M. & Vanutelli, M. E. Promoting creativity through transcranial direct current stimulation (tDCS). A critical review. Front. Behav. Neurosci. 12, 167 (2018).
Kleinmintz, O. M. et al. Participation of the left inferior frontal gyrus in human originality. Brain Struct. Funct. 223, 329–341 (2018).
Colombo, B., Bartesaghi, N., Simonelli, L. & Antonietti, A. The combined effects of neurostimulation and priming on creative thinking. A preliminary tDCS study on dorsolateral prefrontal cortex. Front. Hum. Neurosci. 9, 1–12 (2015).
Cerruti, C. & Schlaug, G. Anodal transcranial direct current stimulation of the prefrontal cortex enhances complex verbal associative thought. J. Cogn. Neurosci. 21, 1980–1987 (2009).
Chrysikou, E. G. et al. Noninvasive transcranial direct current stimulation over the left prefrontal cortex facilitates cognitive flexibility in tool use. Cogn. Neurosci. 4, 81–89 (2013).
Salvi, C., Beeman, M., Bikson, M., McKinley, R. & Grafman, J. TDCS to the right anterior temporal lobe facilitates insight problem-solving. Sci. Rep. 10, 946 (2020).
Metuki, N., Sela, T. & Lavidor, M. Enhancing cognitive control components of insight problems solving by anodal tDCS of the left dorsolateral prefrontal cortex. Brain Stimul. 5, 110–115 (2012).
Peña, J., Sampedro, A., Ibarretxe-Bilbao, N., Zubiaurre-elorza, L. & Ojeda, N. Improvement in creativity after transcranial random noise stimulation (tRNs) over the left dorsolateral prefrontal cortex. Sci. Rep. 9, 7116 (2019).
Ivancovsky, T., Kurman, J., Morio, H. & Shamay-Tsoory, S. Transcranial direct current stimulation (tDCS) targeting the left inferior frontal gyrus: effects on creativity across cultures. Soc. Neurosci. https://doi.org/10.1080/17470919.2018.1464505 (2018).
Lustenberger, C., Boyle, M. R., Foulser, A. A., Mellin, J. M. & Fröhlich, F. Functional role of frontal alpha oscillations in creativity. Cortex 67, 74–82 (2015).
Grabner, R. H., Krenn, J., Fink, A., Arendasy, M. & Benedek, M. Effects of alpha and gamma transcranial alternating current stimulation (tACS) on verbal creativity and intelligence test performance. Neuropsychologia 118, 91–98 (2018).
Chi, R. P. & Snyder, A. W. Facilitate insight by non-invasive brain stimulation. PLoS ONE 6, e16655 (2011).
Chi, R. P. & Snyder, A. W. Brain stimulation enables the solution of an inherently difficult problem. Neurosci. Lett. 515, 121–124 (2012).
Fink, A. et al. Enhancing creativity by means of cognitive stimulation: evidence from an fMRI study. Neuroimage 52, 1687–1695 (2010).
Jung, R. E. et al. Neuroanatomy of creativity. Hum. Brain Mapp. 31, 398–409 (2010).
Jung-Beeman, M. et al. Neural activity when people solve verbal problems with insight. PLoS Biol. 2, E97 (2004).
Sandkühler, S. & Bhattacharya, J. Deconstructing insight: EEG correlates of insightful problem solving. PLoS ONE 3, e1459 (2008).
Fink, A., Graif, B. & Neubauer, A. C. Brain correlates underlying creative thinking: EEG alpha activity in professional vs. novice dancers. Neuroimage 46, 854–862 (2009).
Agnoli, S., Zanon, M., Mastria, S., Avenanti, A. & Corazza, G. E. Enhancing creative cognition with a rapid right-parietal neurofeedback procedure. Neuropsychologia 118, 99–106 (2018).
Benedek, M., Schickel, R. J., Jauk, E., Fink, A. & Neubauer, A. C. Alpha power increases in right parietal cortex reflects focused internal attention. Neuropsychologia 56, 393–400 (2014).
Benedek, M., Bergner, S., Könen, T., Fink, A. & Neubauer, A. C. EEG alpha synchronization is related to top-down processing in convergent and divergent thinking. Neuropsychologia 49, 3505–3511 (2011).
Di Bernardi Luft, C., Zioga, I., Thompson, N. M., Banissy, M. J. & Bhattacharya, J. Right temporal alpha oscillations as a neural mechanism for inhibiting obvious associations. Proc. Natl. Acad. Sci. USA 115, E12144–E12152 (2018).
Beaty, R. E., Benedek, M., Barry Kaufman, S. & Silvia, P. J. Default and executive network coupling supports creative idea production. Sci. Rep. 5, 1–14 (2015).
Ghanavati, E., Nejati, V. & Salehinejad, M. A. Transcranial direct current stimulation over the posterior parietal cortex (PPC) enhances figural fluency: implications for creative cognition. J. Cogn. Enhanc. 2, 88–96 (2018).
Ghanavati, E., Salehinejad, M. A., Nejati, V. & Nitsche, M. A. Differential role of prefrontal, temporal and parietal cortices in verbal and figural fluency: implications for the supramodal contribution of executive functions. Sci. Rep. 9, 3700 (2019).
Fink, A. & Benedek, M. EEG alpha power and creative ideation. Neurosci. Biobehav. Rev. 44, 111–123 (2014).
Benedek, M. et al. Brain mechanisms associated with internally directed attention and self-generated thought. Sci. Rep. 6, 22959 (2016).
Rominger, C. et al. The creative brain in the figural domain: Distinct patterns of EEG alpha power during idea generation and idea elaboration. Neuropsychologia 18, 13–19 (2018).
Benedek, M. The neuroscience of creative idea generation. In Exploring Transdisciplinarity in Art and Sciences (eds Kapoula, Z. et al.) 31–48 (Springer, Berlin, 2018). https://doi.org/10.1007/978-3-319-76054-4_2.
Stevens, C. E. & Zabelina, D. L. Creativity comes in waves: an EEG-focused exploration of the creative brain. Curr. Opin. Behav. Sci. 27, 154–162 (2019).
Wu, X. et al. A meta-analysis of neuroimaging studies on divergent thinking using activation likelihood estimation. Hum. Brain Mapp. 36, 2703–2718 (2015).
Fink, A. & Neubauer, A. C. EEG alpha oscillations during the performance of verbal creativity tasks: differential effects of sex and verbal intelligence. Int. J. Psychophysiol. 62, 46–53 (2006).
Grabner, R. H., Fink, A. & Neubauer, A. C. Brain correlates of self-rated originality of ideas: evidence from event-related power and phase-locking changes in the EEG. Behav. Neurosci. 121, 224–230 (2007).
Gebuis, T., Gevers, W. & Cohen Kadosh, R. Topographic representation of high-level cognition: numerosity or sensory processing?. Trends Cogn. Sci. 18, 1–3 (2014).
Rothmaler, K., Nigbur, R. & Ivanova, G. New insights into insight: Neurophysiological correlates of the difference between the intrinsic “aha” and the extrinsic “oh yes” moment. Neuropsychologia 95, 204–214 (2017).
Kizilirmak, J. M. et al. Learning of novel semantic relationships via sudden comprehension is associated with a hippocampus-independent network. Conscious Cogn. 69, 113–132 (2019).
Becker, M., Sommer, T. & Kühn, S. Inferior frontal gyrus involvement during search and solution in verbal creative problem solving: a parametric fMRI study. Neuroimage 206, 116294 (2020).
Torrance, E. P. The Torrance Tests of Creative Thinking-Norms-Technical Manual Research Edition-Verbal Tests, Forms A and B-Figural Tests, Forms A and B (Personnel Press, Princeton, 1966).
Terney, D., Chaieb, L., Moliadze, V., Antal, A. & Paulus, W. Increasing human brain excitability by transcranial high-frequency random noise stimulation. J. Neurosci. 28, 14147–14155 (2008).
van der Groen, O. & Wenderoth, N. Transcranial random noise stimulation of visual cortex: stochastic resonance enhances central mechanisms of perception. J. Neurosci. 36, 5289–5298 (2016).
Schoen, I. & Fromherz, P. Extracellular stimulation of mammalian neurons through repetitive activation of Na + channels by weak capacitive currents on a silicon chip. J. Neurophysiol. 100, 346–357 (2008).
Snowball, A. et al. Long-term enhancement of brain function and cognition using cognitive training and brain stimulation. Curr. Biol. 23, 987–992 (2013).
Brevet-Aeby, C., Mondino, M., Poulet, E. & Brunelin, J. Three repeated sessions of transcranial random noise stimulation (tRNS) leads to long-term effects on reaction time in the Go/No Go task. Neurophysiol. Clin. 49, 27–32 (2019).
Fertonani, A., Pirulli, C. & Miniussi, C. Random noise stimulation improves neuroplasticity in perceptual learning. J. Neurosci. 31, 15416–15423 (2011).
Simonsmeier, B. A., Grabner, R. H., Hein, J., Krenz, U. & Schneider, M. Electrical brain stimulation (tES) improves learning more than performance: a meta-analysis. Neurosci. Biobehav. Rev. 84, 171–181 (2018).
Kounios, J. & Beeman, M. The cognitive neuroscience of insight. Annu. Rev. Psychol. 65, 71–93 (2014).
Kounios, J. et al. The origins of insight in resting-state brain activity. Neuropsychologia 46, 281–291 (2008).
Kounios, J. & Beeman, M. The Aha! Moment: the cognitive neuroscience of insight. Curr. Dir. Psychol. Sci. 18, 210–216 (2009).
Aziz-Zadeh, L., Liew, S. L. & Dandekar, F. Exploring the neural correlates of visual creativity. Soc. Cogn. Affect. Neurosci. 8, 475–480 (2013).
Klimesch, W., Sauseng, P. & Hanslmayr, S. EEG alpha oscillations: the inhibition-timing hypothesis. Brain Res. Rev. 53, 63–88 (2007).
Rihs, T. A., Michel, C. M. & Thut, G. Mechanisms of selective inhibition in visual spatial attention are indexed by α-band EEG synchronization. Eur. J. Neurosci. 25, 603–610 (2007).
Camarda, A. et al. Neural basis of functional fixedness during creative idea generation: an EEG study. Neuropsychologia 118, 4–12 (2018).
Rottschy, C. et al. Differentiated parietal connectivity of frontal regions for ‘what’ and ‘where’ memory. Brain Struct. Funct. 218, 1551–1567 (2013).
Bonino, D. et al. Spatial imagery relies on a sensory independent, though sensory sensitive, functional organization within the parietal cortex: a fMRI study of angle discrimination in sighted and congenitally blind individuals. Neuropsychologia 68, 59–70 (2015).
Faul, F., Erdfelder, E., Lang, A.-G. & Buchner, A. G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 39, 175–191 (2007).
Oldfield, R. C. The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9, 97–113 (1971).
Torrance, E. P. Torrance Tests of Creative Thinking (Scholastic Testing Service. Inc., Bensenville, 2016).
Jiménez, J. E., Artiles, C., Rodríguez, C., & García, E. Adaptación y baremación del test de pensamiento creativo de Torrance: expresión figurada (Consejería de Educación, Cultura y Deportes del Gobierno de Canarias, Dirección General de Ordenación e Innovación Educativa, 2007).
Miranda, P. C., Mekonnen, A., Salvador, R. & Ruffini, G. The electric field in the cortex during transcranial current stimulation. Neuroimage 15, 48–58 (2013).
Cohen, J. Statistical Power Analysis for the Behavioral Sciences (Lawrence Erlbaum, Mahwah, 1988).
Acknowledgements
We would like to thank all the participants that took part in the experiment.
Funding
This work was supported by a 2018 Leonardo Grant for Researchers and Cultural Creators, BBVA Foundation (The Foundation accepts no responsibility for the opinions, statements and contents included in the project and/or the results thereof, which are entirely the responsibility of the authors), the Department of Education and Science of the Basque Government (Team A) [IT946‐16] and AS was supported by a Predoctoral Fellowship from the Fundación Tatiana Pérez de Guzmán el Bueno.
Author information
Authors and Affiliations
Contributions
J.P., N.O., A.A. and N.I.B. contributed to the study design and conceptualization. Statistical analyses and interpretation were performed by J.P., A.S. and L.Z., A.S, A.A. and J.P. contributed to data collection and management. All authors contributed to the writing and revision of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peña, J., Sampedro, A., Ibarretxe-Bilbao, N. et al. The effect of transcranial random noise stimulation (tRNS) over bilateral posterior parietal cortex on divergent and convergent thinking. Sci Rep 10, 15559 (2020). https://doi.org/10.1038/s41598-020-72532-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-020-72532-3
- Springer Nature Limited
This article is cited by
-
Neuromodulation to Enhance Creative Cognition: a Review of New and Emerging Approaches
Journal of Cognitive Enhancement (2023)
-
The impact of creativity on functional outcome in schizophrenia: a mediational model
npj Schizophrenia (2021)