
Abstract
Olfaction plays key roles in insect survival and reproduction, such as feeding, courtship, mating, and oviposition. The olfactory-based control strategies have been developed an important means for pest management. Streltzoviella insularis is a destructive insect pest of many street tree species, and characterization of its olfactory proteins could provide targets for the disruption of their odour recognition processes and for urban forestry protection. In this study, we assembled the antennal transcriptome of S. insularis by next-generation sequencing and annotated the main olfactory multi-gene families, including 28 odorant-binding proteins (OBPs), 12 chemosensory proteins (CSPs), 56 odorant receptors (ORs), 11 ionotropic receptors (IRs), two sensory neuron membrane proteins (SNMPs), and 101 odorant-degrading enzymes (ODEs). Sequence and phylogenetic analyses confirmed the characteristics of these proteins. We further detected tissue- and sex-specific expression patterns of OBPs, CSPs and SNMPs by quantitative real time-PCR. Most OBPs were highly and differentially expressed in the antennae of both sexes. SinsCSP10 was expressed more highly in male antennae than in other tissues. Two SNMPs were highly expressed in the antennae, with no significant difference in expression between the sexes. Our results lay a solid foundation for understanding the precise molecular mechanisms underlying S. insularis odour recognition.
Similar content being viewed by others


Introduction
Olfaction plays key roles in insect survival and reproduction, and the antennae are regarded as important olfactory organs in insects; they can sensitively detect chemical signals from the environment and produce behavioural reactions, such as feeding, courtship, mating, and oviposition1,2,3,4,5. However, these processes cannot occur without the involvement of olfactory proteins expressed in the antennae. In general, olfactory proteins are classified into several categories: odorant-binding proteins (OBPs), chemosensory proteins (CSPs), olfactory receptors (ORs), ionotropic receptors (IRs), sensory neuron membrane proteins (SNMPs) and odorant-degrading enzymes (ODEs)6,7,8.
OBPs and CSPs are highly abundant in the sensillar lymph of insect antennae. They bind to hydrophobic odorant molecules and transport them in a pH-dependent manner to ORs during the first step of olfactory recognition8,9,10. OBPs are small soluble proteins with a pattern of six cysteines that form two or three disulfide bridges, which are typically divided into two subfamilies in Lepidoptera species, pheromone-binding proteins (PBPs) and general odorant-binding proteins (GOBPs)11. Besides OBPs, CSPs are smaller and more conserved than OBPs; they are characterized by the presence of four cysteines with two disulfide bridges7. Previous studies have verified that CSPs are expressed both in olfactory tissues and non-olfactory tissues, and seemed to play roles in pheromone transport, moulting, development and leg regeneration12,13,14. In addition, SNMPs are olfactory-specific membrane proteins and are homologous to the human fatty-acid transport protein CD36 receptor family15. Lepidopterans generally have two SNMP subfamilies (SNMP1 and SNMP2). SNMP1 is expressed on the dendrite membrane of pheromone-sensitive neurons in Heliothis virescens, and is a crucial cofactor for the detection of the fatty acid derived pheromone 11-cis-vaccenyl acetate in Drosophila. It might be involved in sex pheromone recognition16,17,18. SNMP2 is also associated with pheromone-sensitive sensilla, but it is only expressed in supporting cells18,19. Moreover, insect olfactory reception involves two receptor types (ORs and IRs). Insect ORs are heteromultimers formed by two proteins, a conventional OR and an obligate olfactory co-receptor (Orco)20. The binding of odorant molecules by the OR/Orco complex triggers the transduction of chemical signals to electrical signals that are transmitted to the brain21,22. IRs are a novel family of chemosensory receptors that are related to ionotropic glutamate receptors (iGluRs), and act as ligand-based ion channels23. Insect IRs include two subfamilies: the “antennal IRs” and the species-specific “divergent IRs”24. Earlier studies have proved that IRs also are involved in odour detection. IRs are narrowly tuned for acids and amines during biological decomposition, and ORs are widely tuned for alcohols and esters23,25. After OR activation, olfactory signals must be degraded rapidly to prevent from prolonged olfactory neuronal excitation, and ODEs inactivate odorant molecules by enzymatic degradation in the sensillar lymph of insect antennae, such as carboxylesterase (CEX), aldehyde oxidases (AOX), alcohol dehydrogenase (AD), cytochrome P450 (CYP) and glutathione S-transferases (GST)26,27,28,29.
Streltzoviella insularis (Staudinger) (Lepidoptera: Cossidae) is an important wood-boring insect pest and occurs in many provinces and cities in China. It mainly attacks various street tree species, such as Fraxinus americana, Ginkgo biloba, Sophora spp. and Ulmus spp. Once a female S. insularis lays eggs in the cracks of the host tree trunk, the larvae of this moth hatch out and then bore into the phloem and xylem, which stresses or kills the infested host trees, seriously threatening urban forestry30,31,32. The difficult detection, high population and long lifecycle of S. insularis pose an immense challenge with respect to pest control. Traditional chemical insecticides do not work well against S. insularis and can lead to pesticide resistance and damage to human health and urban environments30. Based on the key roles of insect olfactory proteins in chemical communication, olfactory-based control strategies have been developed an important means for pest management33,34,35,36,37. For example, Jayanthi et al. described a “computational reverse chemical ecology” approach for the screening of attractants based on the binding ability of OBPs as an alternative to behavioural bioassays in Bactrocera dorsalis38. Applying a reverse chemical ecology approach, Choo et al. discovered CquiOR36 responds to acetaldehyde, a potent oviposition attractant for Culex quinquefasciatus39. However, olfactory proteins in S. insularis have not been identified. Therefore, we assembled the antennal transcriptome of male and female S. insularis and identified a series of putative olfactory proteins (OBPs, CSPs, ORs, IRs, SNMPs and ODEs). We further examined the expression patterns of OBPs, CSPs and SNMPs by quantitative real-time PCR (RT-qPCR) in various tissues of the two sexes. These results lay a solid foundation for understanding the molecular basis of odour recognition in S. insularis and other insects, and they provide a basis for the development of new pest control methods targeting the olfactory system in S. insularis.
Results
Transcriptome sequencing and de novo assembly
We performed transcriptome sequencing of male and female S. insularis antennae to identify olfactory multi-gene families, with three replicates per sex. We obtained approximately 52.6 million and 58.8 million raw reads from the antennal cDNA libraries of male and female S. insularis, respectively (see Supplementary Table S1). After filtering low-quality raw sequences, we generated approximately 49.9 million male and 55.8 million female clean reads, respectively (see Supplementary Table S2). Subsequently, both male and female clean reads were assembled together to produce 38,487 unigenes, with an N50 of 2050 bp, an average length of 1359 bp and a maximum length of 24,019 bp (Fig. 1A). The raw reads have been deposited in the National Center for Biotechnology Information (NCBI) – Sequence Read Archive (SRA) database with the accession number SRP166379.

(A) Size distribution of S. insularis unigenes. (B) Species distribution from a homology search with the S. insularis unigenes against the NCBI Nr protein database. (C) GO classification of S. insularis unigenes. (D) KEGG classification of S. insularis unigenes. (A: processes, B: environmental information processing, C: genetic information processing, D: metabolism, E: organismal systems).
Functional annotation of S. insularis unigenes
Among the 38,487 unigenes, 17,984 (46.73%) matched loci in the NCBI non-redundant protein (Nr) database by a BLASTX homology search with a cut-off E-value of 10−5. The best matches were obtained for Danaus plexippus sequences (27.21%), followed by Bombyx mori (23.85%), Papilio xuthus (2.25%), Acyrthosiphon pisum (1.83%) and Tribolium castaneum sequences (1.64%), as shown in Fig. 1B.
Gene Ontology (GO) annotation was used to classify the unigenes into three functional groups (molecular function, cellular component, and biological process) according to the GO categories. Of 38,487 unigenes of S. insularis, 9023 (23.44%) were annotated. As shown in Fig. 1C, 10,973 unigenes were assigned to the molecular function category, and “binding” and “catalytic activity” were the most highly represented terms with this category. A total of 14,108 unigenes were assigned to GO terms in the cellular component category, and “cell part” and “cell” were the most abundant terms. Furthermore, 23,096 unigenes were assigned to GO terms in the biological process category, and the main terms were “metabolic process” and “cellular process”.
The KEGG Orthology (KO) system was used to classify the unigenes into five branches of Kyoto Encyclopedia of Genes and Genomes (KEGG) metabolic pathways, including processes, environmental information processing, genetic information processing, metabolism, and organismal systems. Most unigenes were assigned to the processes branch, and “global and overview maps” was the most highly represented term (Fig. 1D).
Identification of putative OBPs
We identified 28 putative OBPs in S. insularis, including two GOBPs and three PBPs. Among 28 OBPs, 17 OBPs (SinsOBP1, 3–4, 6, 9–12, 16–19, 21–23, PBP3 and GOBP2) were full-length genes with intact open reading frames (ORFs) of at least 400 bp and a signal peptide (see Supplementary Tables S3 and S4). The BLASTX results indicated that all OBPs of S. insularis shared relatively higher amino acid identities with other Lepidoptera OBPs in the NCBI Nr database (>50%). Three OBPs (SinsOBP9, 11 and 16) belonged to the minus–C OBPs based on the lack of the second and fifth cysteines, and the remaining 25 OBPs were identified as classical OBPs with the motif “C1-X15–39-C2-X3-C3-X21–44-C4-X7–12-C5-X8-C6” (where X represents any amino acid). A neighbor–joining phylogenetic tree was constructed using OBPs of Lepidoptera species (Fig. 2). Most OBPs of S. insularis had high homology with those of Eogystia hippophaecolus. SinsGOBP1–2 were clustered with the GOBP family, whereas SinsPBP1–3 formed part of a PBP family clade. Lepidopteran GOBPs and PBPs were highly conserved base on their different functions. Based on reads per kilobase of exon model per million mapped reads (FPKM) values, 13 OBPs (SinsOBP4–6, 9, 14–15, 18, 23, PBP1–3 and GOBP1–2) were highly abundant in male and female antennae of S. insularis (FPKM value > 1,000).

Neighbor-joining phylogenetic tree of putative OBPs from S. insularis, E. hippophaecolus, B. mori, Carposina sasakii, Epiphyas postvittana, Spodoptera litura, Grapholita molesta, Helicoverpa armigera, Ostrinia furnacalis, Spodoptera exigua, Ectropis obliqua and Cydia pomonella. The protein sequences of OBPs used to build phylogenetic trees are listed in Supplementary Table S10. The stability of the nodes was assessed by a bootstrap analysis with 1,000 replications, and only bootstrap values of ≥0.5 are shown at the corresponding nodes. The scale bar represents 0.05 substitutions per site. S. insularis sequences are shown in red.
Identification of putative CSPs
In the antennal transcriptome of S. insularis, we identified 12 putative CSPs with lengths ranging from 564 bp to 2794 bp, including ten CSPs (SinssCSP2–10 and 12) with intact ORFs and signal peptides (see Supplementary Tables S3 and S5). All putative CSPs had four cysteine residues and fit the motif “C1-X6–8-C2-X16–21-C3-X2-C4” (X represents any amino acid). The BLASTX results showed that 12 CSPs had relatively higher amino acid identities with Lepidoptera CSPs in the Nr database (>50%). Based on FPKM values, four CSPs (SinsCSP2, 6, 8 and 10) had relatively high expression levels in the antennal transcriptome of S. insularis (FPKM value > 1,000). According to the neighbor-joining phylogenetic tree of CSPs (Fig. 3), we observed that CSPs are distributed in various clades throughout the cladogram. SinsCSP1, SinsCSP3–7 and SinsCSP9–11 clustered together with EhipCSPs with high bootstrap support.

Neighbor-joining phylogenetic tree of putative CSPs from S. insularis, E. hippophaecolus, B. mori, S. litura, H. armigera, S. exigua, C. pomonella and Agrotis ipsilon. The protein sequences of CSPs used to build phylogenetic trees are listed in Supplementary Table S10. The stability of the nodes was assessed by a bootstrap analysis with 1,000 replications, and only bootstrap values of ≥0.5 are shown at the corresponding nodes. The scale bar represents 0.05 substitutions per site. S. insularis sequences are shown in red.
Identification of putative ORs
We identified 56 putative ORs in the male and female transcriptome, 52 of which were likely full-length OR genes, encoding proteins of more than 300 amino acids with intact ORFs (see Supplementary Tables S3 and S6). The sequence identities of the best BLASTX matches in the Nr database ranged from 45% to 99%. In a FPKM analysis, SinsOrco displayed the highest expression levels in male and female antennae, with FPKM values of 472.06 and 437.42, respectively. However, the other 55 ORs showed the relatively low expression levels, with FPKM values of 0 to 260.09. The expression levels of ORs were relatively lower than those of OBPs and CSPs in S. insularis antennae. Of 56 ORs, 22 ORs sequences were shorter than 300 bp or had no common sites for computing distances; accordingly, we only used the 34 OR sequences of S. insularis to construct a phylogenetic tree. In the neighbor-joining tree of ORs (Fig. 4), two ORs (SinsOR10 and 20) were clustered into the pheromone receptor (PR) clade, and SinsOrco sequence showed high homology to the conserved insect odorant co-receptor clustered in the odorant co-receptor clade (Orco). The remaining ORs were divided to different Lepidoptera ORs ortholog clades.

Neighbor-joining phylogenetic tree of putative ORs from S. insularis, E. hippophaecolus, B. mori, C. sasakii, O. furnacalis, Chilo suppressalis and Plodia interpunctella. The protein sequences of ORs used to build phylogenetic trees are listed in Supplementary Table S10. The stability of the nodes was assessed by bootstrap analysis with 1,000 replications, and only bootstrap values of ≥0.5 are shown at the corresponding nodes. The scale bar represents 2.0 substitutions per site. S. insularis sequences are shown in red.
Identification of putative IRs
We identified 11 putative IRs in the antennal transcriptome of S. insularis, of which five sequences (SinIR93a1, 75q2, 75p2, 75a1 and 21a) were predicted to be full-length sequences with intact ORFs and signal peptides (see Supplementary Tables S3 and S7). The FPKM results showed that SinsIRs had relatively low expression levels (FPKM values ranged from 0.39 to 109.58). Among the 11 IRs, SinsIR93a2 and SinsIR75q1 sequences were shorter than 400 bp. We ued the remaining nine IRs sequences to construct a phylogenetic tree (Fig. 5). The IRs identified in the antennal transcriptome of S. insularis were assigned to the different clades of the conserved IRs with reliable bootstrap support, including IR21a, IR41a, IR68a, IR93a, IR76b and IR75. IR75 included four SinsIRs (SinsIR75p2, SinsIR75q2, SinsIR75q1 and SinsIR75a2).

Neighbor-joining phylogenetic tree of putative IRs from S. insularis, E. hippophaecolus, B. mori C. sasakii, O. furnacalis, C. suppressalis, P. interpunctella and Conogethes punctiferalis. The protein sequences of IRs used to build phylogenetic trees are listed in Supplementary Table S10. The stability of the nodes was assessed by bootstrap analysis with 1,000 replications, and only bootstrap values of ≥0.5 are shown at the corresponding nodes. The scale bar represents 1.0 substitutions per site. S. insularis sequences are shown in red.
Identification of putative SNMPs
Two putative SNMPs with intact ORFs (1473 and 1566 bp, respectively) were identified in the combined male and female antennal transcriptome of S. insularis (see Supplementary Tables S3 and S8) and shared relatively high identities (over 95%) with SNMPs of E. hippophaecolus. The FPKM analysis showed that the expression levels of SinsSNMPs in male antennae were higher than those in female antennae. Based on a phylogenetic tree (Fig. 6), SinsSNMP1 and SinsSNMP2 clustered within the SNMP1 subfamily and SNMP2 subfamily, respectively.

Neighbor-joining phylogenetic tree of putative SNMPs from S. insularis, E. hippophaecolus, S. litura, C. suppressalis, Sesamia inferens and Ostrinia nubilalis. The protein sequences of SNMPs used to build phylogenetic trees are listed in Supplementary Table S10. The stability of the nodes was assessed by a bootstrap analysis with 1,000 replications, and only bootstrap values of ≥0.5 are shown at the corresponding nodes. The scale bar represents 0.05 substitutions per site. S. insularis sequences are shown in red.
Identification of putative ODEs
We also identified 101 putative ODEs in the antennal transcriptome of S. insularis. ODEs were divided into five families, including 19 CEXs, ten AOXs, eight ADs, 47 CYPs and 17 GSTs (see Supplementary Tables S3 and S9). Among the 101 putative ODE genes, 12 had intact ORFs and signal peptides (SinsCEX3, 5, 7, 9–11, 16, 19, SinsAD4, SinsCYP18 and SinsGST8, 13). The BLASTX results indicated that 101 identified ODEs shared relatively higher amino acid identities ranging from 50% to 98% with Lepidoptera ODEs in the NCBI Nr protein database. An FPKM analysis showed that only three ODEs (SinsCEX3, SinsCYP20 and 36) were highly abundant in the antennal transcriptome of S. insularis (FPKM value > 1,000).
Tissue-specific and sex-specific expression of putative OBPs, CSPs and SNMPs
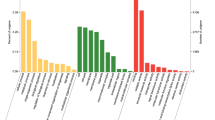
To better understand the functions of 28 putative OBPs, 12 CSPs and two SNMPs in the different tissues of male and female S. insularis, we used RT-qPCR to evaluate expression patterns. We found that 20 OBPs were specifically and highly expressed in both female and male antennae (Fig. 7), of which 14 OBPs (SinsOBP2, 4, 6–7, 9–10, 12, 14–15, 22–23, PBP1–2 and GOBP2) were expressed at significantly higher levels in males than in females. Two OBPs (SinsOBP13 and 20) were expressed at significantly higher levels in female antennae than in male antennae. The expression of four OBPs (SinsOBP5, 18, PBP3 and GOBP1) were not significantly different between male and female antennae. In addition, three OBPs (SinsOBP8, 16 and 19) were more highly expressed in the male genitalia than in other tissues. SinsOBP17 was highly expressed in the male legs. Finally, the remaining four OBPs (SinsOBP1, 3, 11 and 21) were expressed in various tissues of both sexes. In CSPs, six CSPs (SinsCSP2–4, 6, 8, 10) were highly expressed in the antennae, and SinsCSP10 expression in male antennae was highly significantly different from expression in female antennae. The other six CSPs (SinsCSP1, 5, 7, 9, 11–12) were expressed in all tissues. In addition, the expression of SinsSNMP1 and SinsSNMP2 were highly expressed in the antennae of S. insularis, with no significant difference in expression level was found between the sexes.

Expression profiles of the putative OBPs, CSPs and SNMPs in various S. insularis tissues. A: antennae; L: legs; G: genitalia (copulatory organ for male moths; ovipositor for female moths). β-actin was used as a reference gene for the normalization of target gene expression. Blue and red represents males and females, respectively. The standard errors are represented by the error bars, and different lowercase letters (a–d) above the bars denote significant differences (p < 0.05).
Discussion
The gene sets reported in this study substantially increase the pool of genes encoding olfactory protein genes in Cossidae. Prior to our study, members of the major olfactory gene families in Cossidae were identified only from the antennal transcriptome of E. hippophaecolus (excluding ODEs)40. What’s more, the previous studies in S. insularis had focused on biology and ecology. Recently, next-generation sequencing is widely used for the identification of olfactory proteins in insects owing to their important functional roles40,41,42,43,44,45. Our results will provide potential targets for the disruption of the olfactory system in S. insularis for pest control purposes.
We identified 38,487 unigenes with a mean length of1359 bp from the male and female S. insularis antennal transcriptome, indicating the high quality and great depth of sequencing at the transcriptome level. BLASTX homology search in the NCBI Nr protein database found that S. insularis unigenes shared the relatively high homology with sequences from other Lepidoptera species, further supporting the accuracy of our transcriptome data. Additionally, we identified large numbers of transcripts encoding putative olfactory proteins, including 28 OBPs, 12 CSPs, 56 ORs, 11 IRs, two SNMPs and 101 ODEs. These olfactory protein counts in S. insularis were similar to these in E. hippophaecolus (29 OBPs, 18 CSPs, 63 ORs, 12 IRs, and two SNMPs)40, Carposina sasakii (29 OBPs, 13 CSPs, 52 ORs, eight IRs, and one SNMPs)45, Plutella xyllostella (24 OBPs, 15 CSPs, 54 ORs, 16 IRs, and two SNMPs)46 and other Lepidoptera insects. By contrast, previous studies have detected fewer ODEs; Zhang et al. (2017) identified 18 putative CXEs and four AOXs genes from Cnaphalocrocis medinalis antennal transcriptome27, Liu et al. (2015) reported 19 putative CXEs and 16 GSTs in Chilo suppressalis antennae47, and Leitch et al. (2015) identified 97 candidate CYPs and 39 CEXs in the antennal transcriptome of Calliphora stygia28.
Hydrophobic odours are thought to interact with OBPs and CSPs prior to the ligand–receptor interaction. An OBP has a mass of approximately 15–17 kDa48. We detected fewer OBPs in the S. insularis antennal transcriptome than previously reported in Plodia interpunctella49, Mythimna separate50, and Bombyx mori51, but more than in Sesamia inferens52. In the phylogenetic tree of OBPs, we observed the differentiation of Lepidopteran OBPs into several branches over a long evolutionary timescale, in accordance with previous results40,41,42,43,44,45,46. GOBPs and PBPs subfamilies formed separate clusters, suggesting that they diverged from a common ancestral gene due to speciation and reproductive isolation49. Furthermore, we observed distinct tissue-biased of OBPs in insects, strongly indicative of biological functions53. In general, an antenna-enriched expression profile is correlated with a role in olfactory perception, whereas genes that are highly expressed in gustatory organs, such as the proboscis, leg and ovipositor, could be involved in taste detection54,55,56,57,58. Remarkably, 20 OBPs (SinsOBP2, 4–7, 9–10, 12–15, 18, 20, 22–23, SinsPBP1–3 and SinsGOBP1–2) were highly expressed in the antennae and may have vital roles in the detection of odorants, such as sex pheromones and host plant volatiles. These results were consistent with the expression patterns observed in other insects belonging to Lepidoptera, such as E. hippophaecolus40, Helicoverpa assulta59, and Agrotis ipsilon60. SinsOBP17 was highly expressed in the legs and may therefore be involved in the recognition of these contact substances on host plant surfaces61. In addition, we observed three OBPs (SinsOBP8, 16 and 19) that were highly expressed in male genitalia; these might contribute to the controlled release of semiochemicals into the environment62. In comparison, the molecular weights of CSPs (10–15 kDa) were lower than those of OBPs7. The number of CSPs reported in our antennal transcriptome was in accordance with the number detected for A. ipsilon60, but less than those in the P. interpunctella49 and M. separate50. SinsCSPs were distributed among various clades in the CSP phylogeny, indicating that these genes may have various functions in chemical communication12,13,14. In our RT-qPCR analysis, CSPs were expressed in both olfactory organ and non-olfactory organ, suggesting that they have broad functions in S. insularis, including roles in non-olfactory functions. We identified two SNMPs (SinsSNMP1 and SinsSNMP2) assigned to the SNMP1 subfamily and SNMP2 subfamily in the phylogenetic tree; the high divergence suggests that these SNMPs have different function in vivo. SinsSNMP1 and SinsSNMP2 were specifically expressed in antennae. Combined with previous findings, SNMPs may be indispensable for the identification of sex pheromones16,17,18,19,63.
Moth olfactory receptors belong to two families: ORs and IRs. ORs are seven transmembrane domain proteins with an inverted topology compared with that of vertebrates ORs and play a crucial role in chemosensory reception20,64,65. We identified 56 ORs were identified in the S. insularis antennal transcriptome, similar to the numbers identified in other moths (63 in E. hippophaecolus and 58 in Cydia pomonella)40,66. In a phylogenetic tree of ORs, SinsOR10 and SinsOR20, and other Lepidopteran PRs, were assigned to the same clade in the tree, particularly those encoding PR orthologues in E. hippophaecolus. SinsOR10 and SinsOR20 could be the PRs in S. insularis. Additionally, SinsOrco was clustered into the Orco clade, with Orco orthologues also in E. hippophaecolus. Accordingly, SinsOrco is most likely the Orco. In addition, IRs are a conserved family of synaptic ligand-gated ion channels, but the specificity of ligand recognition by IRs is still unclear23,24,67. We identified 11 IRs in S. insularis, similar to the numbers in other Lepidoptera species, such as E. hippophaecolus40, Athetis dissimilis68 and P. interpunctella49. A phylogenetic analysis suggested that SinsIR93a1, SinsIR76b, SinsIR41a, SinsIR21a, and SinsIR68a are the IR93a, IR76b, IR41a, IR21a, IR68a, respectively, and SinsIR75p2, SinsIR75q2, SinsIR75q1 and SinsIR75a2 are the IR75 genes of S. insularis. IR8a and IR25a are were predicted to be the co-receptors present in the IR group and are conserved in many insects23,25; however, we did not find them in the S. insularis antennal transcriptome, which may be explained by a lower expression level in S. insularis, this indicated a further study was needed to identify.
In insects, the rapid degradation of odorant molecules in antennae is important for the sensitivity of olfactory receptor neurons. Odorant degradation in antennae is mediated by various enzymes. We identified 19 CEXs, ten AOXs, eight ADs, 47 CYPs and 17 GSTs in S. insularis. Of these, many antennae-specific CXEs have been identified in various insects, uch as Spodoptera exigua, S. littoralis, S. litura and D. melanogaster, and are involved in olfactory signal (host plant volatiles and sex pheromones) termination69,70,71. AOXs are also found in insect antennae and are thought to degrade aldehyde odorant compounds in B. mori and Mamestra brassicae72,73. Recently, ADs and CYPs were cloned from insect antennae and may play roles in xenobiotic degradation, detoxification and biotransformation of endobiotic compounds74,75. GSTs are also related to odorant degradation. For example, BmGSTD4 is specifically expressed in male B. mori antennae, suggesting it is involved in sex pheromone degradation76. It is possible that many transcripts of putative enzymes are ODEs of S. insularis, but further biochemical analyses of their specific physiological roles are necessary.
Conclusions
S. insularis is regarded as a destructive wood-boring pest affecting various street trees in China. However, the olfactory system of this moth has not been deciphered so far. In this study, we first assembled the antennal transcriptome of male and female S. insularis and annotated a set of olfactory genes, including 28 OBPs, 12 CSPs, 56 ORs, 11 IRs, two SNMPs, and 101 ODEs. Then, Sequence and phylogenetic analyses confirmed the characteristics of these proteins. Additionally, using RT-qPCR, we observed both tissue- and sex-specific expression of OBPs, CSPs and SNMPs, indicating that these loci play crucial roles in courtship, mating and oviposition behaviour. Further studies are needed to determine the function of putative PBPs and PRs of S. insularis, to reveal their specificity and binding to sex pheromones. These analyses could provide novel targets for the disruption of chemical communication in S. insularis for pest control purposes.
Methods
Ethics statement
S. insularis is not on the “List of Endangered and Protected Animals in China”. The Beijing Municipal Bureau of Landscape and Forestry issued a permit for the field collection.
Insect and tissue collection
S. insularis individuals were collected from Fraxinus americana on Beijing Forestry University North Road, Haidian District, Beijing, China (40°0′N, 116°20′E), in May 2017. Damaged trunks were cut off and taken to the laboratory. Larvae inside the trunks were fed on the phloem and xylem of the host under natural environmental conditions. After their eclosion, the moths were sorted by sex according to the genitalia. 150 antennae, 60 legs, and 60 genitalia (copulatory organ for male moths; ovipositor for female moths) were excised from the two sexes, immediately placed in RNAlater (Ambion, Austin, TX, USA), and then stored at −80 °C.
RNA extraction
Total RNA was extracted from 25 antennae of each sex using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) following the manufacturer’s instructions, with three replicates per sex. RNA purity was evaluated using the NanoDrop 2000 (Thermo, Waltham, MA, USA), RNA concentration was measured using the QubitRNA Assay Kit with a Qubit2.0 Fluorometer (Life Technologies, Carlsbad, CA, USA), RNA integrity was detected using the Agilent Bioanalyzer 2100 system (Agilent Technologies, Santa Clara, CA, USA), and RNA degradation and contamination were monitored on a 1% agarose gel to ensure the quality of the RNA samples for subsequent transcriptome sequencing.
cDNA library construction and Illumina sequencing
cDNA library construction and Illumina sequencing were performed at Shanghai Majorbio Bio-pharm Technology Co., Ltd. (Shanghai, China). According to the TruSeq RNA Sample Preparation Guide V2 (Illumina), mRNA was purified from total RNA using oligo (dT) magnetic beads and then fragmented by the addition of fragmentation buffer. Random hexamer primers were used to synthesize the first-strand cDNA, followed by the synthesis of the second-strand cDNA using dNTPs, RNase H and DNA polymerase I. Remaining overhangs were passivated via polymerase activity. After the end-repairing, Poly-A tailing and the ligation of adapters, cDNA fragments of 150–200 bp were purified using the AMPure XP system (Beckman Coulter, Beverly, MA, USA). Then, 3 μl USER Enzyme (NEB, Ipswich, MA, USA) was used with size-selected, adaptor-ligated cDNA at 37 °C for 15 min, followed by 5 min at 95 °C prior to PCR amplification. PCR products were purified using the AMPure XP system and library quality was assessed on the Agilent Bioanalyzer 2100 system. The cDNA libraries of S. insularis were sequenced on the Illumina HiseqTM 4,000 platform, and paired-end reads were generated.
Sequence assembly and functional annotation
To obtain the clean reads, the raw reads were processed to remove low-quality reads and adapter sequences. Then, GC Content, Q20 and Q30 were used to assess the sequencing quality. Clean reads were assembled de novo with Trinity77 after combined the male and female S. insularis clean reads. The largest alternative splicing variants in the Trinity results were called unigenes. BLASTX searches were used to align unigenes and compare them with the NCBI Nr database, using a cut-off E-value of 10−5. Then, Nr BLASTX results were subjected to GO annotation using Blast2GO78. Pathway annotations for unigenes were determined using KO79. ORF of each unigenes were then predicted using ORF finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). FPKM values calculated by RSEM (RNA-Seq by Expectation-Maximization) with default parameters represented gene expression levels in the male and female S. insularis antennae80.
Identification of olfactory genes and phylogenetic analyses
Using TBLASTN, the sequences of OBP, CSP, OR, IR, SNMP and ODE from insecta species as queries to identify putative unigenes encoding putative OBPs, CSPs, ORs, IRs, SNMPs and ODEs in S. insularis. All putative OBPs, CSPs, ORs, IRs, SNMPs and ODEs were manually checked by the BLASTX program in NCBI online. The N-terminal signal peptides of putative OBPs, CSPs, ORs, IRs, SNMPs, and ODEs were found using SignalP4.0 (http://www.cbs.dtu.dk/services/SignalP/). All putative OBP, CSP, OR, IR, SNMP and ODE amino acid sequences of S. insularis and other insect species were aligned using ClustalW implemented in MEGA (version 5.0). Phylogenetic trees were generated by the neighbor-joining method as implemented in MEGA (version 5.0), with the p-distance model and pairwise deletion of gaps. Bootstrap support for tree branches was assessed by re-sampling amino acid positions 1000 times81. Phylogenetic trees were color-coded and arranged using FigTree (Version 1.4.2). The amino acid sequences of olfactory proteins used to build phylogenetic trees are listed in Supplementary Table S10.
Expression patterns of putative OBPs by RT-qPCR
Expression patterns of putative OBPs in various tissues (antennae, legs and genitals) of the two sexes were analysed by RT-qPCR using the Bio-Rad CFX96 PCR System (Hercules, CA, USA). Total RNA was extracted from 25 antennae, ten legs and ten genitals from each sex following the method described above, and was transcribed into cDNA using the PrimeScriptRT Reagent Kit with gDNA Eraser (No. RR047A; TaKaRa, Shiga, Japan). Gene-specific primers were designed using Primer 3 Plus (http://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi) (see Supplementary Table S11). β-actin from S. insularis was used as a reference gene. The RT-qPCR mixtures were composed of 12.5 µl of TB Green™ Premix Ex Taq™ II (Tli RNaseH Plus) (No. RR820A; TaKaRa), 1 µl of forward primer (10 µM), 1 µl of reverse primer (10 µM), 2 µl of cDNA and 8.5 µl of sterilized H2O. RT-qPCR cycling parameters were as follows: 95 °C for 30 s, followed by 40 cycles of 95 °C for 5 s and 60 °C for 30 s, and 65 °C–95 °C in increments of 0.5 °C for 5 s to generate the melting curves. To check reproducibility, each reaction for each tissue was performed in three biological replicates and three technical replicates. Negative controls without the template were included in each experiment. The relative expression levels were calculated according to the comparative 2−ΔΔCT method (the amplification efficiency for 42 genes was close to 100%)82, and the female leg sample was used as the calibrator, β-actin was used for calculating and normalizing the target gene expression and correcting for sample to sample variation. Data (means ± SE) from different samples were subjected to one-way nested analysis of variance, followed by Tukey’s honestly significance difference tests implemented in SPSS Statistics 22.0 (IBM, Chicago, IL, USA).
References
Asahina, K., Pavlenkovich, V. & Vosshall, L. B. The survival advantage of olfaction in a competitive environment. Curr. Biol. 18, 11531–155 (2008).
Field, L. M., Pickett, J. A. & Wadhams, L. J. Molecular studies in insect olfaction. Insect Mol. Biol. 9, 545–551 (2000).
Leal, W. S. Odorant reception in insects: roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 58, 373–391 (2013).
Smadja, C. & Butlin, R. K. On the scent of speciation: the chemosensory system and its role in premating isolation. Heredity 102, 77–97 (2009).
Zhang, J., Walker, W. B. & Wang, G. Pheromone Reception in Moths: From Molecules to Behaviors. Prog. Mol. Biol. Transl. Sci. 130, 109–28 (2015).
Pelosi, P., Iovinella, I., Felicioli, A. & Dani, F. R. Soluble proteins of chemical communication: an overview across arthropods. Front. Physiol. 5, 320 (2014).
Pelosi, P., Zhou, J. J., Ban, L. P. & Calvello, M. Soluble proteins in insect chemical communication. Cell Mol. Life Sci. 63, 1658–1676 (2006).
Sánchez-Gracia, A., Vieira, F. G. & Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 103, 208–216 (2009).
Fan, J., Francis, F., Liu, Y., Chen, J. L. & Cheng, D. F. An overview of odorant-binding protein functions in insect peripheral olfactory reception. GMR 10, 3056–3069 (2011).
Lautenschlager, C., Leal, W. S. & Clardy, J. Coil-to-helix transition and ligand release of Bombyx mori pheromone-binding protein. Biochem. Biophys. Res. Commun. 335, 1044–1050 (2005).
Pelosi, P. & Maida, R. Odorant-binding proteins in insects. Comp. Biochem. Physiol. 111, 503–514 (1995).
Wanner, K. W., Isman, M. B., Feng, Q., Plettner, E. & Theilmann, D. A. Developmental expression patterns of four chemosensory protein genes from the Eastern spruce budworm, Chroistoneura fumiferana. Insect Mol. Biol. 14, 289–300 (2005).
Li, Z. Q. et al. Gene Identification of pheromone gland genes involved in type II sex pheromone biosynthesis and transportation in female tea pest Ectropis grisescens. G3 8, 899–908 (2018).
Kitabayashi, A. N., Arai, T., Kubo, T. & Natori, S. Molecular cloning of cDNA for p10, a novel protein that increases in the regenerating legs of Periplaneta americana (American cockroach). Insect Biochem. Mol. Biol. 28, 785–790 (1998).
Vogt, R. G. et al. The insect SNMP gene family. Insect Biochem. Mol. Biol. 39, 448–456 (2009).
Benton, R., Vannice, K. S. & Vosshall, L. B. An essential role for a CD36-related receptor in pheromone detection in Drosophila. Nature 450, 289–293 (2007).
Jin, X., Ha, T. S. & Smith, D. P. SNMP is a signaling component required for pheromone sensitivity in. Drosophila. Proc. Natl. Acad. Sci. USA 105, 10996–11001 (2008).
Forstner, M. et al. Differential expression of SNMP-1 and SNMP-2 proteins in pheromone-sensitive hairs of moths. Chem. Senses 33, 291–299 (2008).
Rogers, M. E., Krieger, J. & Vogt, R. G. Antennal SNMPs (sensory neuron membrane proteins) of Lepidoptera define a unique family of invertebrate CD36-like proteins. J. Neurobiol. 49, 47–61 (2001).
Clyne, P. J. et al. A novel family of divergent seven-transmembrane proteins: candidate odorant receptors in Drosophila. Neuron 22, 327–338 (1999).
Hallem, E. A. & Carlson, J. R. Coding of odors by a receptor repertoire. Cell 125, 143–160 (2006).
Vosshall, L. B. & Hansson, B. S. A unified nomenclature system for the insect olfactory coreceptor. Chem. Senses 36, 497–498 (2011).
Benton, R., Vannice, K. S., Gomez-Diaz, C. & Vosshall, L. B. Variant ionotropic glutamate receptors as chemosensory receptors in Drosophila. Cell 136, 149–162 (2009).
Croset, V. et al. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. Plos genetics 6, e1001064 (2010).
Abuin, L. et al. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 69, 44–60 (2011).
Liu, S., Gong, Z. J., Rao, X. J., Li, M. Y. & Li, S. G. Identification of Putative Carboxylesterase and Glutathione S-transferase Genes from the Antennae of the Chilo suppressalis (Lepidoptera: Pyralidae). J. Insect Sci. 15, 103 (2015).
Zhang, Y. X., Wang, W. L., Li, M. Y., Li, S. G. & Liu, S. Identification of putative carboxylesterase and aldehyde oxidase genes from the antennae of the rice leaffolder, Cnaphalocrocis medinalis, (Lepidoptera: Pyralidae). J. Asia-Pacific Entomol. 20, 907–913 (2017).
Leitch, O., Papanicolaou, A., Lennard, C., Kirkbride, K. P. & Anderson, A. Chemosensory genes identified in the antennal transcriptome of the blowfly Calliphora stygia. BMC Genomics 16, 255 (2015).
Yang, B., Ozaki, K., Ishikawa, Y. & Matsuo, T. Identification of candidate odorant receptors in Asian corn borer Ostrinia furnacalis. Plos One 10, e0121261 (2015).
Gao, R. T. & Qin, X. X. Preliminary study on Holcocerus insularis. For. Pest Dis. 1, 3–5 (1983).
Liu, H. X. et al. Sensilla on the antennae and ovipositor of the carpenterworm, Streltzoviella insularis (Staudinger, 1892) (Lepidoptera, Cossidae). Oriental Insects 52, 420–433 (2018).
Xu, L. L., Pei, J. H., Wang, T., Ren, L. L. & Zong, S. X. The larval sensilla on the antennae and mouthparts of five species of Cossidae (Lepidoptera). Can. J. Zool. 95, 611–622 (2017).
Zhao, Y. Y., Liu, F., Yang, G. & You, M. S. PsOr1, a potential target for RNA interference-based pest management. Insect Mol. Biol. 20, 97–104 (2011).
Lu, Y. L., Yao, Y., Li, S., Zhang, Q. & Liu, Q. J. Olfactory biosensor based on odorant-binding proteins of Bactrocera dorsalis with electrochemical impedance sensing for pest management. Sensor Rev. 37, 397–403 (2017).
Chu, W. T. et al. Constant pH molecular dynamics (CpHMD) and molecular docking studies of CquiOBP1 pH-induced ligand releasing mechanism. J. Mol. Model 19, 1301–1309 (2013).
Leal, W. S. et al. Reverse and conventional chemical ecology approaches for the development of oviposition attractants for Culex mosquitoes. Plos One 3, e3045 (2008).
Leal, W. S. Pheromone reception. Topics in Curr. Chem. 240, 1–36 (2005).
Jayanthi, P. D. K. et al. Computational reverse chemical ecology: Virtual screening and predicting behaviorally active semiochemicals for Bactrocera dorsalis. BMC Genomics 15, 209 (2014).
Choo, Y. M. et al. Reverse chemical ecology approach for the identification of an oviposition attractant for Culex quinquefasciatus. P. Natl. Acad. Sci. USA 115, 714–719 (2018).
Hu, P. et al. Antennal transcriptome analysis and expression profiles of odorant binding proteins in Eogystia hippophaecolus (Lepidoptera: Cossidae). BMC Genomics 17, 651 (2016).
Hu, P., Wang, J. Z., Cui, M. M., Tao, J. & Luo, Y. Q. Antennal transcriptome analysis of the Asian longhorned beetle Anoplophora glabripennis. Sci. Rep. 6, 26652 (2016).
Wang, J. Z., Hu, P., Gao, P., Tao, J. & Luo, Y. Q. Antennal transcriptome analysis and expression profiles of olfactory genes in Anoplophora chinensis. Sci. Rep. 7, 15470 (2017).
Bin, S. Y., Qu, M. Q., Pu, X. H., Wu, Z. Z. & Lin, J. T. Antennal transcriptome and expression analyses of olfactory genes in the sweetpotato weevil Cylas formicarius. Sci. Rep. 7, 11073 (2017).
Feng, B., Guo, Q. S., Zheng, K. D., Qin, Y. X. & Du, Y. J. Antennal transcriptome analysis of the piercing moth Oraesia emarginata (Lepidoptera: Noctuidae). Plos One 12, e0179433 (2017).
Tian, Z. Q. et al. Antennal transcriptome analysis of the chemosensory gene families in Carposina sasakii, (Lepidoptera: Carposinidae). BMC Genomics 19, 544 (2018).
Yang, S. Y., Cao, D. P., Wang, G. R. & Liu, Y. Identification of genes involved in chemoreception in Plutella xyllostella by antennal transcriptome analysis. Sci. Rep. 7, 11941 (2017).
Liu, S., Gong, Z. J., Rao, X. J., Li, M. Y. & Li, S. G. Identification of Putative Carboxylesterase and Glutathione S-transferase Genes from the Antennae of the Chilo suppressalis (Lepidoptera: Pyralidae). J. Insect Sci. 15, 103 (2015).
Briand, L., Nespoulous, C., Huet, J. C. & Pernollet, J. C. Disulfide pairing and secondary structure of ASP1, an olfactory-binding protein from honeybee (Apis mellifera L). J. Peptide Res. 58, 540–545 (2002).
Jia, X. J., Zhang, X. F., Liu, H. M., Wang, R. Y. & Zhang, T. Identification of chemosensory genes from the antennal transcriptome of Indian meal moth Plodia interpunctella. Plos One 13, e0189889 (2018).
Chang, X. Q. et al. De novo analysis of the oriental armyworm Mythimna separata antennal transcriptome and expression patterns of odorant-binding proteins. CPB 22, 120–130 (2017).
Gong, D. P., Zhang, H. J., Zhao, P., Xia, Q. Y. & Xiang, Z. H. The odorant binding protein gene family from the genome of silkworm, Bombyx mori. BMC Genomics 10, 332 (2009).
Zhang, Y. N. et al. Differential expression patterns in chemosensory and non-Chemosensory tissues of putative chemosensory genes identified by transcriptome analysis of insect pest the purple stem borer Sesamia inferens (Walker). Plos One 8, e69715 (2013).
Hull, J. J., Perera, O. P. & Snodgrass, G. L. Cloning and expression profiling of odorant-binding proteins in the tarnished plant bug, Lygus lineolaris. Insect Mol. Biol. 23, 78–97 (2014).
Pelosi, P., Iovinella, I., Felicioli, A. & Dani, F. R. Soluble proteins of chemical communication: an overview across arthropods. Front. Physiol. 5, 320 (2014).
Brito, N. F., Moreira, M. F. & Melo, A. C. A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 95, 51–65 (2016).
Swarup, S., Morozova, T. V., Sridhar, S., Nokes, M. & Anholt, R. R. Modulation of feeding behavior by odorant-binding proteins in Drosophila melanogaster. Chem. Senses 39, 125–132 (2014).
Jeong, Y. T. et al. An odorant-binding protein required for suppression of sweet taste by bitter chemicals. Neuron 79, 725–737 (2013).
Gu, S. H. et al. Functional characterization and immunolocalization of odorant binding protein 1 in the lucerne plant bug, Adelphocoris lineolatus (Goeze). Arch. Insect Biochem. Physiol. 77, 81–99 (2011).
Zhang, J. et al. Antennal transcriptome analysis and comparison of chemosensory gene families in two closely related Noctuidae moths, Helicoverpa armigera and H. assulta. Plos One 10, e0117054 (2015).
Gu, S. H. et al. Molecular characterization and differential expression of olfactory genes in the antennae of the black cutworm moth Agrotis ipsilon. Plos One 9, e103420 (2014).
Gu, S. H. et al. Identification and tissue distribution of odorant binding protein genes in the lucerne plant bug Adelphocoris lineolatus (Goeze). Insect Biochem. Mol. Biol. 41, 254–263 (2011).
Sun, Y. L., Huang, L. Q., Pelosi, P. & Wang, C. Z. Expression in antennae and reproductive organs suggests a dual role of an odorant-binding protein in two sibling Helicoverpa species. Plos One 7, e30040 (2012).
Liu, C. C., Zhang, J., Liu, Y., Wang, G. R. & Dong, S. L. Expression of SNMP1 and SNMP2 genes in antennal sensilla of Spodoptera exigua (HÜBNER). Arch. Insect Biochem. Physiol. 85, 114–126 (2014).
Vosshall, L., Amrein, H., Morozov, P., Rzhetsky, A. & Axel, R. A spatial map of olfactory receptor expression in the Drosophila antenna. Cell 96, 725–736 (1999).
Benton, R., Sachse, S., Michnick, S. W. & Vosshall, L. B. Atypical membrane topology and heteromeric function of Drosophila odorant receptors in vivo. Plos Biol. 4, 240–257 (2006).
Walker, W. B., Gonzalez, F., Garczynski, S. F. & Witzgall, P. The chemosensory receptors of codling moth Cydia pomonella-expression in larvae and adults. Sci. Rep. 6, 23518 (2016).
Rytz, R., Croset, V. & Benton, R. Ionotropic receptors (IRs): chemosensory ionotropic glutamate receptors in Drosophila and beyond. Insect Biochem. Mol. Biol. 43, 888–897 (2013).
Dong, J. F., Song, Y. Q., Li, W. L., Shi, J. & Wang, Z. Y. Identification of putative chemosensory receptor genes from the Athetis dissimilis antennal transcriptome. Plos One 11, e0147768 (2016).
Durand, N. et al. A diversity of putative carboxylesterases are expressed in the antennae of the noctuid moth Spodoptera littoralis. Insect Mol. Biol. 19, 87–97 (2010).
He, P., Li, Z. Q., Liu, C. C., Liu, S. J. & Dong, S. L. Two esterases from the genus Spodoptera degrade sex pheromones and plant volatiles. Genome 57, 201–208 (2014).
Younus, F. et al. Identification of candidate odorant degrading gene/enzyme systems in the antennal transcriptome of Drosophila melanogaster. Insect Biochem. Mol. Biol. 53, 30–43 (2014).
Pelletier, J. et al. Identification of candidate aldehyde oxidases from the silkworm Bombyx mori potentially involved in antennal pheromone degradation. Gene 404, 31–40 (2007).
Merlin, C. et al. A new aldehyde oxidase selectively expressed in chemosensory organs of insects. Biochem. Biophys. Res. Commun. 332, 4–10 (2005).
Zhang, Y. N., Xia, Y. H., Zhu, J. Y., Li, S. Y. & Dong, S. L. Putative pathway of sex pheromone biosynthesis and degradation by expression patterns of genes identified from female pheromone gland and adult antenna of Sesamia inferens (Walker). J. Chem. Ecol. 40, 439–451 (2014).
Keeling, C. I. et al. CYP345E2, an antenna-specific cytochrome P450 from the mountain pine beetle, Dendroctonus ponderosae Hopkins, catalyses the oxidation of pine host monoterpene volatiles. Insect Biochem. Mol. Biol. 43, 1142–1151 (2013).
Tan, X. et al. Antenna-specific glutathione S-transferase in male silkmoth Bombyx mori. Int. J. Mol. Sci. 15, 7429–7443 (2014).
Grabherr, M. G. et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 29, 644 (2011).
Conesa, A. et al. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 21, 3674–3676 (2005).
Moriya, Y., Itoh, M., Okuda, S., Yoshizawa, A. C. & Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 35, 182–185 (2007).
Trapnell, C. et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nature Biotechnol. 28, 511–515 (2010).
Saitou, N. & Nei, M. The neighbor-joining method-a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425 (1987).
Livak, K. J. & Schmittgen, T. D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 25, 402–408 (2001).
Acknowledgements
This research was funded by Beijing’s Science and Technology Planning Project (Z171100001417005).
Author information
Authors and Affiliations
Contributions
Y.Y. performed the experiments, analysed the data and drafted the manuscript. W.L. collected samples. Y.Y., J.T. and S.Z. designed the study. J.T. and S.Z. reviewed the manuscript. All authors approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yang, Y., Li, W., Tao, J. et al. Antennal transcriptome analyses and olfactory protein identification in an important wood-boring moth pest, Streltzoviella insularis (Lepidoptera: Cossidae). Sci Rep 9, 17951 (2019). https://doi.org/10.1038/s41598-019-54455-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-54455-w
- Springer Nature Limited
This article is cited by
-
Use of semiochemicals for surveillance and control of hematophagous insects
Chemoecology (2020)