
Abstract
Species-specific behavior-modifying chemicals have been used for more than 50 years for monitoring and management of insect pests of agriculture and human health. Elaterid beetle larvae are among insect pests in soil that are increasingly problematic, in part due to the lack of effective management strategies. However, little is known about the insect-produced chemicals that mediate the reproductive behavior of these pests. We used chemical and behavioral studies to identify, synthesize, and field test the sex attractant pheromone of adults of Melanotus communis, commonly called the corn wireworm, the larvae of which are economically important pests of U.S. crops. Our results indicated that a single female-produced chemical, 13-tetradecenyl acetate, was strongly attractive to conspecific male beetles, and did not appear to attract other species. In field evaluations, male M. communis exhibited a dose-dependent response to this compound. In a trial comparing different slow-release dispensers, a small rubber septum impregnated with the chemical was as effective as and easier to use than a plastic bag dispenser. Given that the sex attractant of this insect consists of a single compound that can be readily synthesized, its development for monitoring and management of the corn wireworm may be economically feasible.
Similar content being viewed by others

Introduction
The click beetles (Coleoptera: Elateridae) comprise a large and diverse family of insects, with > 12,000 described species worldwide (P.J. Johnson, unpubl. catalog). The larvae of pest species are known as wireworms, and can be significant agricultural pests. Their importance as pests is increasing due to the phasing out of effective but environmentally detrimental insecticides that had been used in their control over the past several decades. Sex pheromones or sex attractants have been developed for monitoring and management of a number of Eurasian click beetle species1. However, to date, pheromones have been identified from only two North American species, in the genus Cardiophorus Eschscholtz2, from the ~1,000 known species on this continent3,4. Earlier reports of pheromone identifications for the North American species Limonius californicus (Mannerheim)5 and L. canus LeConte6,7 have not been substantiated in subsequent field trials (unpublished data; JMS in California, P. Landolt and D. Horton in Washington state, G. Reddy in Montana).
In the eastern United States and southeastern Canada, the genus Melanotus Eschscholtz includes several economically important species, including M. communis (Gyllenhal), M. depressus (Melsheimer), and M. verberans (LeConte). In particular, M. communis is an important pest of maize, small grains, sugarcane, and vegetable crops in much of the United States east of the Rocky Mountains8,9,10,11,12. Effective insecticidal options for in-season control of wireworms are disappearing, with currently registered insecticides being of limited efficacy13,14. There is some promise with sugarcane genotype resistance15 and some non-lethal methods could help to manage wireworms in sugarcane16,17,18,19. However, even low populations of M. communis are a serious concern to sugarcane growers because it has been reported that just one wireworm per 1.5 m of sugarcane row can cause a reduction in stand and yield by 7.0% and 3.8%, respectively10. There is also a low market tolerance for injury to tuber/root vegetables, such as potatoes and sweetpotatoes, due to the feeding holes caused by wireworms. These feeding holes can also facilitate attack by other invertebrates as well as pathogens. Thus, additional methods of monitoring and managing M. communis would be an asset for multiple cropping systems. One such method that might be explored is the development of practical applications for sex pheromones of this species, as has been done for a congener20,21.
Pheromone glands of female M. fusciceps (Gyllenhal) and M. castanipes (Paykull) from western Russia contained complex mixtures that included esters of 14-carbon saturated and unsaturated alcohols, but the exact structures and their evaluation in field trials have not been reported22. In contrast, research groups in Japan identified and field-tested female-produced sex pheromones for M. okinawensis Ôhira (dodecyl acetate)1,23 and M. sakishimensis Ôhira ((E)-9,11-dodecadienyl butyrate and (E)-9,11-dodecadienyl hexanoate)1,24. (E)-9,11-Dodecadienyl butyrate and (E)-9,11-dodecadienyl hexanoate have also been identified in extracts from M. tamsuyensis Bates, but field trials with the two compounds were not reported25. Practical applications for these pheromones have since been developed for monitoring purposes, and for control via mass trapping and mating disruption20,21,26,27,28,29,30.
The serendipitous discovery that male M. depressus were attracted to (E)-11-tetradecenyl acetate and (E)-11-tetradecenol29,30, the pheromone components of the tufted apple bud moth (TABM), Platynota idaeusalis (Walker)31, suggested that one or both of these compounds might also be pheromone components of Melanotus spp. However, to our knowledge, it was never determined whether M. depressus females actually produced (E)-11-tetradecenyl acetate and/or (E)-11-tetradecenol. This fragmentary data from several Melanotus spp. from North America and Eurasia suggested that the pheromones of species in this genus are likely to be esters of saturated or unsaturated 12- or 14-carbon alcohols.
Thus, the goal of the research reported here was to identify the sex pheromones of one or more North American Melanotus species, with the working hypothesis that the identification of the pheromone for one species would likely provide insight into the pheromones of congeners. As a first model species, we focused our attention on the economically important M. communis. Here, we report the identification, synthesis, and field-testing of 13-tetradecenyl acetate and several related compounds as possible female-produced sex pheromone components for this species.
Methods
Insect collection and rearing
Melanotus communis adults were collected from agricultural fields at the USDA-ARS U.S. Vegetable Laboratory and Clemson University Coastal Research and Extension Center in Charleston, SC, USA, (32°44′44.06″N 80°03′40.35″W, elev. 4 m) intermittently from May to September in 2016 and 2017. Beetles were collected with Malaise traps, blacklight traps, at incandescent lights, and by hand under debris and vegetation in the fields. Malaise traps were used without a killing agent, and were monitored several times daily to collect live beetles. Light traps were monitored at night and again shortly after sunrise. Mating status, age, and other life history factors of the beetles were not known. After collection, adults were held individually in ventilated transparent 45 ml plastic vials (12 dram, No. 55–12, Thornton Plastics Co., Salt Lake City, UT, USA) streaked with honey and containing a piece of paper towel moistened with distilled water. Insects were held (22 °C ± 1, 50% r.h., L:D 16:8) for 1 to 7 d prior to overnight shipment to the University of California, Riverside Entomology Quarantine Facility (USDA-APHIS permits P526P-14-03526 and P526P-17-02384). All Melanotus species were identified by comparison with reference specimens of known identity, supported with genital morphology and taxonomic keys32. Voucher specimens are deposited in the Severin-McDaniel Insect Research Collection at South Dakota State University, the Entomology Research Museum at the University of California, Riverside, and the National Museum of Natural History through the USDA-ARS Systematic Entomology Laboratory, Washington, D.C.
Preparation and analysis of extracts of potential female-produced pheromones
Headspace volatiles were collected from individual live females held in 250 ml glass canning jars, with lids cut from TeflonTM sheet (ca. 1.6 mm thickness; McMaster-Carr, Elmhurst, IL, USA) and fitted with an air inlet and outlet. The beetles were provided with 10% sugar water for nutrition (in a glass vial with a cotton-tipped applicator wick) and a small wire mesh perch. Emitted compounds were trapped on collectors made from a glass tube (0.5 cm ID) with a 1-cm-long bed of activated charcoal (50–200 mesh; Fisher Scientific, Pittsburgh, PA, USA), held in place by glass wool plugs. For the female that was shipped to UC Riverside in June 2016, humidified and charcoal purified air was pushed through the glass jar at a flow rate of 250 ml/min for 4 d. In contrast, for collection of headspace volatiles from two females that were shipped to UC Riverside in June 2017, charcoal filtered air was pulled through the jars at 250 ml/min by applying a slight vacuum to the outlet. Headspace volatiles were collected for 3 d. All collections were conducted in a temperature controlled room (23 °C), equipped with fluorescent lighting and a north-facing window (~5 m × 1 m) that provided ambient lighting (ca. 12:12 L:D). Trapped volatiles were eluted from the collectors with 0.5 ml of dichloromethane (Optima grade, Fisher Scientific, Pittsburg, PA, USA). After collections of headspace volatiles were terminated, volatiles were collected from abdomens of freeze-killed females. Individual abdomens were crushed in a 20 ml glass vial, the vial was sealed with aluminum foil, and the volatiles were sampled using solid phase microextraction (SPME)2. Volatiles were absorbed for 2 h onto a SPME fiber pushed through the aluminum foil sealing the vial (100 μm PDMS; Supelco, Bellefonte, PA, USA), then analyzed as described below.
Extracts of headspace volatiles were analyzed by coupled gas chromatography-mass spectrometry (GC-MS) with an Agilent 7820 A gas chromatograph coupled to an Agilent 5977E mass selective detector (Agilent Technologies, Santa Clara, CA, USA) fitted with an autosampler and an HP-5 column (30 m × 0.25 mm ID, Agilent). Injections were made in splitless mode, with the purge valve opened after 30 sec. The oven was programmed from 40 °C for 1 min, then increased 10 °C/min to 280 °C. Volatiles from crushed abdomens that were adsorbed onto SPME fibers were analyzed with an Agilent 6890 N gas chromatograph interfaced to an Agilent 5975 C mass selective detector. The GC was also fitted with a HP-5 column and the temperature program used was 40 °C for 1 min, then increased 10 °C/min to 280 °C, hold for 20 min. Loaded SPME fibers were desorbed in the injector port (250 °C) in splitless mode for 30 sec prior to starting the run. Compounds were tentatively identified by mass spectral interpretation, matches with database spectra (W8N05ST; Wiley version 8.0 and NIST, version 5.0), and matches with published retention indices33. Identifications were confirmed by matching retention times and mass spectra with those of authentic standards.
Coupled gas chromatography-electroantennogram detection (GC-EAD) was used to test the responses of the antennae of male M. communis beetles to various C14 acetates, specifically tetradecyl acetate, 13-tetradecenyl acetate, (E)-11-tetradecenyl acetate, and (E)-11,13-tetradecadienyl acetate. For all males, the legs were removed and genitalia were glued shut with Super Glue® before they were taken out of the quarantine facility (a requirement of the permit) for use in GC-EAD analyses. Analyses were conducted on an HP 5890 Series II GC (Hewlett-Packard, now Agilent) fitted with a DB-WAX column (30 m × 0.25 mm ID × 0.25 μm film; J&W Scientific, Folsom, CA, USA). The effluent from the column was split using an ‘X’ cross, with half of the sample going to the flame-ionization detector and the other half to the antennal preparation, with the 4th arm of the X-cross providing make-up gas (~0.62 ml/min). The portion directed to the EAD was diluted in a humidified air stream (~550 ml/min) directed over the antennal mounting block2. Because the four acetates have similar retention times, GC-EAD analyses of the standards were done by alternating injections of the four compounds at ~2.5 min intervals using an isothermal oven temperature (220 °C) in split mode (injector temperature 250 °C), so that an antennal preparation was exposed sequentially to each synthetic compound during a run (n = 12 replicates, using one antenna each from four males).
Authentic standards of pheromone candidates
Dodecyl acetate (henceforth 12:OAc) was purchased from TCI America (Portland, OR, USA), and 11-dodecenyl acetate (henceforth 11–12:OAc) and (E)-11-tetradecenyl acetate (henceforth E11-14:OAc) from Bedoukian Research (Danbury, CT, USA). Tufted apple bud moth sex pheromone lures were purchased from Scentry Biologicals Inc. (Billings, MT, USA).
Other compounds were prepared as described below. Unless otherwise specified, solutions were dried over anhydrous Na2SO4, and concentrated by rotary evaporation under partial vacuum. Liquid chromatography purifications were carried out with 230–400 mesh silica gel. Tetrahydrofuran (THF) was purified by distillation from sodium benzophenone ketyl, and all reactions with air- or water-sensitive reagents were carried out in oven-dried glassware under argon.
Synthesis of tetradecyl acetate (henceforth 14:OAc)
Tetradecyl acetate was prepared by dropwise addition of acetyl chloride (2.14 ml, 30 mmol) to an ice-bath cooled solution of tetradecanol (5.35 g, 25 mmol), pyridine (2.42 ml, 30 mmol), and dimethylaminopyridine (~100 mg) in 100 ml CH2Cl2. The mixture was then warmed to room temperature and stirred for 3 h, followed by addition of 1 ml EtOH and stirring for 1 h, to eliminate excess acetyl chloride as ethyl acetate. The mixture was then concentrated, and the residue was partitioned between hexane and water. The hexane layer was washed sequentially with aqueous 1 M HCl, saturated aq. NaHCO3, and brine, then dried and concentrated. The residue was purified by Kugelrohr distillation (oven temp ~105 °C; 0.15 mm Hg), yielding the ester as a clear, colorless liquid (5.60 g, 88%). 1H NMR (400 MHz, CDCl3): δ 4.03 (t, 2H, J = 6.8 Hz), 2.03 (s, 3H), 1.56–1.64 (m, 2H), 1.37–1.20 (m, 22 H), 0.87 (t, 3H, J = 6.4 Hz). 13C NMR (100 MHz, CDCl3): δ 171.20, 64.64, 31.92, 29.65, 29.57, 29.52, 29.35, 29.26, 28.60, 25.91, 22.68, 20.98, 14.09. MS (m/z, %): 241 (M+ -15, trace), 196 (3), 184 (trace), 168 (4), 154 (1), 153 (1), 140 (2), 139 (2), 130 (trace), 125 (7), 116 (4), 111 (19), 97 (39), 83 (52), 69 (51), 61 (41), 55 (53), 43 (100).
Synthesis of 13-tetradecenyl acetate (henceforth 13–14:OAc)
A solution of 11-bromoundecanol (5.02 g, 20 mmol), dihydropyran (2.26 ml, 25 mmol), and ~50 mg p-toluenesulphonic acid (PTSA) in 50 ml ether was stirred overnight at room temperature. The mixture was then diluted with hexane, extracted twice with saturated aq. NaHCO3 and once with brine, then dried and concentrated. The residue was purified by vacuum flash chromatography, eluting with 5% EtOAc in hexane, yielding 6.66 g of the THP-protected bromoalcohol (99%). This compound was taken up in 30 ml dry THF, and ~10 ml of the solution was added to a dry, argon-flushed flask charged with 0.72 g of freshly ground Mg chips. Once the Grignard reaction had started, as evidenced by warming of the flask and a grey color, the remainder of the solution was added to the flask over ~30 min. The mixture was then warmed to 50 °C and stirred for 1.5 h, then cooled to room temperature.
An oven-dried flask flushed with argon was charged with 300 mg CuI and 40 ml dry THF, and cooled to ~−10 °C in an ice-acetone bath. The Grignard solution was then added dropwise over 40 min with a syringe pump, producing a blue-black slurry. Allyl bromide (2.42 g, 20 mmol) in 5 ml THF was then added dropwise by syringe pump over 45 min, and the resulting mixture was stirred an additional 30 min, then quenched with saturated aqueous NH4Cl, and diluted with hexane. The organic layer was washed with saturated aqueous NH4Cl and brine, then dried and concentrated. The residue was taken up in 100 ml MeOH, ~50 mg of PTSA was added, and the mixture was stirred overnight at room temperature. Then, 5 g of solid NaHCO3 were added, and most of the MeOH was removed by rotary evaporation. The residue was partitioned between hexane and water, and the hexane layer was washed with saturated aqueous NaHCO3 and brine, then dried and concentrated. The crude product was purified by vacuum flash chromatography, eluting with 25% EtOAc in hexanes, followed by Kugelrohr distillation (oven temp ~80 °C, 0.05 mm Hg), yielding 3.33 g (15.7 mmol, 79%) of 13-tetradecenol. The alcohol was taken up in CH2Cl2, pyridine (1.58 g, 20 mmol) and dimethylaminopyridine (~100 mg) were added, and the solution was cooled in an ice-bath. Acetyl chloride (1.44 ml, 20 mmol) was added in one aliquot, and the mixture was warmed to room temperature and stirred for 3 h. The reaction was not complete, so a further 1 ml of pyridine and 0.5 ml of acetyl chloride were added, and the mixture was stirred an additional 2 h. EtOH (1.5 ml) was then added, and the mixture was stirred 1 h. The mixture was then concentrated, and the residue was partitioned between hexane and water. The hexane layer was washed sequentially with aqueous 1 M HCl, saturated aqueous NaHCO3, and brine, then dried and concentrated. The crude product was purified by Kugelrohr distillation, first removing a forerun fraction (oven temp < 80 °C at 0.05 mm Hg, ~0.5 g), followed by distillation of the desired product (oven temperature ~90 °C, 0.04 mm Hg), yielding 13-tetradecen-1-yl acetate (2.81 g, 70%), which gave a single peak on GC analysis. 1H NMR (400 MHz, CDCl3): δ 5.80 (m, 1 H), 4.98 (br d, J = 14.1 Hz, 1 H), 4.92 (br d, J = 10.2 Hz, 1 H), 4.04 (t, J = 6.8 Hz, 2 H), 2.07–1.98 (m, 5 H), 1.64–1.56 (m, 2 H), 1.43–1.22 (m, 18 H). 13C NMR (400 MHz, CDCl3): δ 171.19, 139.21, 114.08, 64.64, 33.81, 29.59, 29.55, 29.50, 29.25, 29.14, 28.94, 28.60, 25.91, 20.99. MS (m/z, %): 194 (M+-AcOH, 5), 166 (3), 152 (2), 151 (2), 138 (5), 137 (5), 123 (10), 109 (21), 105 (1), 96 (55), 91 (2), 82 (83), 77 (3), 68 (75), 61 (16), 55 (100), 51 (1), 43 (90).
Synthesis of (E)-11,13-tetradecadien-1-yl acetate (henceforth E11,13–14:OAc)
A solution of THP-protected 11-dodecyn-1-ol (4.2 g, 16 mmol) and ~50 mg triphenylmethane indicator in dry THF was cooled to ~−10 °C in an ice-salt bath and stirred under argon. A solution of BuLi (8 ml, 20 mmol, 2.5 M in hexanes) was added dropwise, resulting in a bright red solution. Dry paraformaldehyde (4 g) was then added in one portion, and the mixture was warmed to room temperature overnight. The mixture was quenched with saturated aqueous NH4Cl and extracted sequentially with hexane and ether. The combined organic layers were washed with water and brine, dried, and concentrated to a colorless oil which was added dropwise to a slurry of LiAlH4 (1.32 g, 35 mmol) in 60 ml dry THF at 0 °C under argon. The mixture was heated to 65 °C for 2 h, then cooled in an ice-bath and quenched by dropwise sequential addition of 1.4 ml water, 1.05 ml 20% aqueous NaOH, and 4.9 ml water. The resulting slurry was stirred overnight, then filtered through a pad of Celite®, rinsing with ether. The filtrate was dried and concentrated, and the residue was purified by vacuum flash chromatography, eluting with 20% EtOAc in hexanes, yielding 2.75 g of the desired alkenol product as one peak by GC analysis. The purified alcohol was taken up in 150 ml CH2Cl2 and the solution was cooled in an ice-bath, then activated MnO2 was added (29 g, 333 mmol), and the mixture was warmed to room temperature and stirred overnight. The mixture was filtered through a pad of Celite®, rinsing well with CH2Cl2. After concentration, the crude aldehyde was used directly in the next step.
A dry flask was charged with methyltriphenylphosphonium bromide (5.54 g, 15.5 mmol) and dry THF (100 ml). The slurry was cooled in an ice-bath, and potassium t-butoxide (1.68 g, 15 mmol) was added in one portion. The mixture was stirred at 0 °C for 15 min, then at room temperature for 3 h, yielding a milky yellow slurry. The slurry was cooled to 0 °C, and a solution of the crude aldehyde (1.9 g, ~6.4 mmol) in 10 ml THF was added by syringe pump over 1 h. The resulting mixture was slowly warmed to room temperature over 3 h, then quenched with saturated aqueous NH4Cl. The mixture was extracted with hexane, and the hexane layer was washed with brine, dried, and concentrated. The residue was taken up in 50 ml hexane to precipitate most of the triphenylphosphine oxide byproduct, and the resulting slurry was filtered. The filtrate was concentrated, the residue was taken up in 25 ml MeOH, and ~50 mg PTSA was added. The solution was stirred for 3 h at room temperature, then 2 g solid NaHCO3 were added, and the mixture was concentrated. The residue was partitioned between hexane and water, and the hexane layer was washed sequentially with water and brine, then dried and concentrated and purified by vacuum flash chromatography, eluting with 25% EtOAc in hexane, yielding the dienol (1.03 g, 77%).
The dienol (0.53 g, 2.5 mmol) was acetylated as described above. The crude product was purified by Kugelrohr distillation (oven temp ~85 °C, 0.05 mm Hg), yielding (E)-11,13-tetradecadien-1-yl acetate as a colorless oil (0.52 g, 82%), 97% pure by GC. 1H NMR (400 MHz, CDCl3): δ 6.30 (ddd, J = 17.0, ~10.3 Hz, ~10.3 Hz, 1 H), 6.04 (br dd, J = 15.2, 10.4 Hz, 1 H), 5.70 (dt, J = 15.2, 6.9 Hz, 1 H), 5.07 (br d, J ~ 17.5 Hz, 1 H), 4.94 (br d, J ~ 10.2 Hz, 1 H), 4.04 (t, J = 6.8 Hz, 2 H), 2.10–2.02 (m, 2 H), 2.04 (s, 3 H), 1.64–1.58 (m, 2 H), 1.42–1.24 (m, 14 H). 13C NMR (400 MHz, CDCl3): δ 171.24, 137.37, 135.59, 130.86, 114.57, 64.66, 32.55, 29.48, 29.46, 29.24, 29.18, 28.60, 25.90, 21.02. MS (m/z, %): 252 (M+, 5), 209 (1), 192 (8), 177 (trace), 163 (4), 149 (7), 121 (23), 110 (16), 107 (16), 95 (39), 81 (71), 79 (71), 73 (3), 67 (100), 61 (9), 54 (53), 43 (92).
Field bioassays of synthetic pheromone candidates
All field bioassays were conducted at the USDA-ARS U.S. Vegetable Laboratory and Clemson University Coastal Research and Extension Center in Charleston, SC. This shared facility encompasses a total of ca. 295 ha, of which ca. 95 ha are crop fields, with the remainder being buildings, wooded areas, and wetlands. In 2016, we tested pheromone candidates for M. communis, including the compounds identified from extracts of the insects, and several related compounds (Table 1). The main focus of this field trial was to determine the activity of the insect-produced 14:OAc and 13–14:OAc on M. communis males. In addition, the 12-carbon acetates were included due to their structural similarities to the pheromone components of M. okinawensis23. E11-14:OH, E11-14:OAc, and E11,13-14:OAc were tested to determine if they were attractive or synergized attraction to 13–14:OAc and 14:OAc, and E11,13–14:OAc was tested due to its structural similarities to pheromone components identified from M. sakishimensis and M. tamsuyensis24,25. The sex pheromone of the tufted apple bud moth (TABM), Platynota idaeusalis, (2:1 ratio of (E)11-14:OAc + (E)11-14:OH) was included because of reports29,30 of attraction of Melanotus spp. to TABM lures. Field trials were conducted for 6 wk (4 August – 13 September 2016) at three fields; one planted to maize (0.52 ha), one to sweetpotato (1.13 ha), and the other to cucumber (0.69 ha). Distance between these three fields ranged from 360 to 570 m. For all treatments (with the exception of the commercial TABM lure), ~10 mg of undiluted blend was pipetted into a 0.2 ml PCR tube (Multiply-Pro; Sarstedt AG & Co., Nümbrecht, Germany) with a pinhole in the lid made with a no. 3 insect pin. Empty, punctured PCR tubes were used as controls. Lures were suspended from the center of each trap. Lures were replaced and treatment locations in each transect were re-randomized biweekly. Linear trap transects were established at each field. In each transect, seven Pherocon® 1 C wing traps (white, ca. 27 × 23 cm; Trécé Inc., Adair, OK, USA) were deployed at the border between the field and an adjacent woodlot. Traps were spaced 12 m apart, and were hung 1 m above the ground from wooden stakes. Click beetles were removed from traps weekly and the adhesive was removed with xylene and an ultrasonic cleaner34.
In 2017 field bioassays were conducted at 11 field sites for 18 wk (1 May – 6 September) with six test treatments and a control (Table 1). These fields were planted to maize, with the exception of two fields, one of which was planted to sweetpotato, and the other was fallow. These 11 fields ranged in size from 0.32 to 0.77 ha, and were from 50 to 1210 m apart. The majority of the treatments were the same as the 2016 field trials, with the exceptions of the 12-carbon acetates and the quaternary mixture of 14-carbon acetates. E11-14:OAc and E11,13-14:OAc were added to a blend of 13–14:OAc and 14:OAc to determine if they synergized attraction to the pheromone candidates. Test blends were diluted with isopropanol ( ≥ 99.5% purity; Sigma-Aldrich, St. Louis, MO, USA) to a concentration of 5.45 mg/ml. One milliliter of each treatment blend was pipetted into 5 × 7.5 cm (~0.05 mm thickness) low-density polyethylene zipper seal bags (#01-816-1 A; Thermo-Fisher Scientific, Waltham, MA, USA), which were hung from the center of the traps. Vernon Pitfall Traps® (ca. 17 × 14 cm) constructed of black polypropylene were deployed in transects in the approximate center of each field, with traps in each transect spaced 12 m apart. These traps combine the attributes of pitfall and pheromone traps, and have been successfully used to test attractants for other click beetle species35. The killing agent in the traps was 12 ml of a 1:1 mixture of water and propylene glycol (Prestone LowTox® Antifreeze/Coolant; Prestone Products Corp., Lake Forest, IL, USA). Click beetles were removed from traps weekly and lures were replaced and treatment locations in each transect were re-randomized biweekly.
Dose-response test of 13-tetradecenyl acetate
Having determined that 13–14:OAc was as good or better as an attractant than any blend of 14-carbon acetates (see results), we conducted field trials to determine the optimal dose of 13–14:OAc for attraction of M. communis. In 2018, a trial was set up for 8 wk (25 June – 20 August) in six fields; three planted to maize, and one field each to cucumber, tomato, and peanut. These six fields ranged in size from 0.38 to 0.73 ha, and were from 50 to 1135 m apart. At each field site a trap transect was established along the field border using five Vernon Pitfall Traps®, each 12 m apart. Test doses were 0.33, 1.0, 3.3, and 10 mg/ml of 13–14:OAc (1 ml loads), with an isopropanol control, dispensed from zipper-seal polyethylene bags as described above. Beetles were collected weekly, and lures were replaced and re-randomized every 2 wk.
Comparison of pheromone dispensers for 13-tetradecenyl acetate
In 2018, a trial was set up for 7 wk (22 June – 10 August) to compare two lure devices at 12 fields different from those in the dose-response study described above. Eight of these fields were planted to maize, two fields to cucumber, and one field each to tomato and peanut. These 12 fields ranged in size from 0.38 to 0.73 ha, and were from 45 to 1779 m apart. At each field site, two Vernon Pitfall Traps® were deployed along the field border, 12 m apart. At each site, one of the traps was baited with a zipper-seal bag (as described above) and the other with a grey rubber septum (9 mm × 19 mm, 6.2 mm ID; West Co., Lionville, PA, USA; item # 1888 grey). The zipper seal bags were loaded with 1.0 ml of a 10 mg/ml solution of 13–14:OAc in isopropanol. Rubber septa were Soxhlet-extracted with dichloromethane for 8 h, then dried in a fume hood before being loaded with 0.2 ml of a hexane solution of 13–14:OAc (50 mg/ml). Loaded septa were allowed to absorb the pheromone solution and dry, then were sealed in glass vials for shipment to South Carolina. Treatment locations were re-randomized and click beetles were collected weekly.
Diversity of Melanotus species at the field sites and cross-attraction
To gain a better understanding of the Melanotus species occurring at our field sites, two unbaited Malaise traps (Townes style; Sante Traps, Lexington, KY, USA) were set up at two of the fields (traps separated by 620 m) at the border between the woodlot and the field. Every year (2016–2018) trapping began in April and was terminated in November. Collection vessels contained ca. 50 ml of a 1:1 mixture of water and propylene glycol as described above. Traps were serviced weekly. Melanotus specimens captured in these traps were identified to species as described above. Voucher specimens are deposited in the same museums noted above for the specimens collected for putative pheromone analysis.
Statistical analysis
Captures (total catch/trap) of M. communis were analyzed using a multi-response permutation procedure (MRPP)36. MRPP is a nonparametric multivariate analysis that does not require distributional assumptions of the dependent variables. Each field was considered as a replicate in the analysis. Data were square-root transformed prior to MRPP analysis using Euclidean distance to conduct the distance matrix (PC-ORD 7.0; MjM Software Design, Gleneden Beach, OR, USA)37. Pairwise comparisons of treatments were Benjamini-Hochberg38 adjusted using a false discovery rate of 0.25. Unadjusted P and A (chance-corrected within-group agreement) values are presented in the results. Capture data from comparison of pheromone dispensers in 2018 were square-root transformed prior to paired t-test.
Results
Identification of pheromone candidates
Coupled GC-MS analyses of extracts of headspace volatiles collected from live female M. communis did not show peaks from any likely pheromone candidates, but only traces of hydrocarbons and contaminants. However, GC-MS analyses of SPME-trapped headspace volatiles from three separate crushed abdomens of these same beetles revealed two major peaks in the total ion chromatograms (Fig. 1A) which were absent in the corresponding chromatograms of extracts of headspace volatiles from the live individuals. The two compounds were tentatively identified from matches of their mass spectra with database spectra as tetradecyl acetate and a monounsaturated analog (Fig. 1B). For the unsaturated 14-carbon acetate, the double bond was tentatively determined to be in the terminal position, by comparison of its Kováts retention index with the retention indices of all possible straight-chain monounsaturated 14-carbon acetates on a DB-5 column33. The identifications of the two female-produced compounds were confirmed by matching their retention times and mass spectra with those of authentic standards.

(A) Representative total ion chromatogram of an SPME extract of volatiles from the crushed abdomen of an adult female Melanotus communis, highlighting the possible pheromone candidates 13-tetradecen-1-yl acetate (peak 1) and tetradecyl acetate (peak 2); (B) EI mass spectrum (70 eV) of the female-produced 13-tetradecen-1-yl acetate.
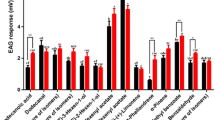
Additionally, in GC-EAD analyses testing the synthesized pheromone components and analogs, antennae of male beetles were challenged sequentially with 14:OAc, 13–14:OAc, E11-14:OAc, and E11,13–14:OAc. Of the four compounds, 13–14:OAc consistently elicited the strongest responses from antennae of males (Fig. 2). There were negligible antennal responses to E11-14:OAc, and smaller and inconsistent responses to 14:OAc and E11, 13–14:OAc (Fig. 2).

Representative GC-EAD analysis of alternating injections of 14:OAc (peak 1), 13–14:OAc (peak 2), E11-14:OAc (peak 3), and E11,13–14:OAc (peak 4), stimulating an antenna from a male M. communis beetle. Top trace: GC chromatogram; bottom, inverted trace: signal from antennal responses. Large peaks in the GC trace are from the solvent used to make sequential injections, and the asterisks indicate the small responses elicited from the antenna by the solvent.
Field bioassays
Comparison of synthetic sex pheromone candidates
In preliminary bioassays in 2016, a total of 26 male M. communis were caught, with lures containing 13–14:OAc being most attractive (Fig. 3). In particular, 13–14:OAc as a single component attracted marginally more beetles (MRPP; P = 0.053, A = 0.081) than the 4:1 binary blend of 13–14:OAc + 14:OAc, but not more (P > 0.05) than the 1:1 blend of these two compounds. Captures in the 1:1 and 4:1 blends of 13–14:OAc + 14:OAc were not significantly different (P > 0.05). The 4:1 blend of 13–14:OAc + 14:OAc did not differ (P > 0.05) from the control. No beetles were caught in the traps baited with 14:OAc, the 12:OAc + 11–12:OAc blend, or TABM pheromone.

Mean total capture ( + SE) per trap of male Melanotus communis beetles in traps baited with 12-carbon and 14-carbon acetates in 2016 (4 August – 13 September). Means with the same letter are not significantly different (P > 0.05).
In 2017, using plastic bag lures and Vernon Pitfall Traps® instead of plastic tube dispensers and sticky traps, 449 male M. communis were attracted to treatments containing the 14-carbon acetates, and trends were similar to those of 2016. The 4:1 and 1:1 blends of 13–14:OAc + 14:OAc and 13–14:OAc as a single component were not significantly different (Fig. 4), whereas all of these were significantly more attractive than the 1:1:1:1 blend of 13–14:OAc + 14:OAc + (E)-11,13–14:OAc + (E)-11-14:OAc (Fig. 4). The TABM pheromone, 14:OAc, and control treatments attracted one, two, and five males, respectively, and did not differ from each other (P > 0.05).

Mean total capture ( + SE) per trap of male Melanotus communis beetles in traps baited with 14-carbon acetates in 2017. Means with the same letter are not significantly different (P > 0.05).
Dose-response tests with 13-tetradecenyl acetate
In the 2018 trial using a semi-log range of doses, a total of 135 male M. communis were captured. The 10 mg dose was significantly more attractive than the lower doses (Fig. 5; MRPP 10 vs 0.33; P = 0.002, A = 0.318) (MRPP 10 vs 1; P = 0.001, A = 0.401) (MRPP 10 vs 3.3; P = 0.018, A = 0.165). Captures with the 0.33, 1.0, and 3.3 mg doses were not significantly different from each other (P > 0.05), but were significantly greater than the control.

Mean total capture ( + SE) per trap of male Melanotus communis beetles in traps baited with 0–10 mg doses over a semi-log range in 2018. Means with the same letter are not significantly different (P > 0.05).
Comparison of pheromone dispensers for 13-tetradecenyl acetate
In this trial comparing equal doses of pheromone released from plastic bag or rubber septum dispensers, a total of 149 male M. communis were captured (septa = 76; bags = 73). Captures in traps baited with the polyethylene bags declined in an exponential manner from 3.33 males per trap in wk 1 (22–29 June) to zero in wk 4 (13–20 July), after which trap catches were negligible (Fig. 6). Captures in traps baited with the rubber septa were relatively stable, i.e., ca. 1.6 males per trap per week for 4 wk (22 June–20 July), after which trap catches declined (Fig. 6). For both dispenser types, trap catches after wk 4 were at or near zero. Catches in traps baited with plastic bags were not significantly different (P > 0.05) than those in traps baited with septa at wk 1 and 2 (22–29 June and 29 June – 6 July), but were significantly less than the septa in wk 3 (6–13 July; T11 = 4.00, P = 0.0021) and wk 4 (13–20 July; T10 = 3.78, P = 0.0036).

Weekly capture (mean ± SE) per trap of male Melanotus communis beetles in traps baited with 10 mg 13–14:OAc released from rubber septa or polyethylene bags in 2018. Dates with an asterisk indicate significant treatment effect (P < 0.05).
Diversity of Melanotus species at the field sites and specificity of pheromone lures
Of the 758 Melanotus individuals captured in traps baited with test chemicals during the pheromone field bioassays, all but two were male M. communis. The exceptions were one male M. lanei Quate (1–8 May 2017) and one female M. piceatus Blatchley (8–15 May 2017), both of which were captured in traps baited with the 1:1 mixture of 13–14:OAc + 14:OAc. An additional 85 Melanotus individuals were captured in Malaise traps, 43 of which were M. communis. The remaining Melanotus, and their numbers, captured in the Malaise traps were: M. piceatus (15), M. verberans (15), M. corticinus (Say) (5), M. morosus Candèze (3), M. similis (3), and M. americanus (Herbst) (1), but no males of any of these species were caught in any of the lure-baited traps, indicating a high species specificity of the M. communis pheromone.
Discussion
Our analytical and bioassay data indicate that 13–14:OAc is the major and likely only component of the female-produced sex pheromone of M. communis. Male M. communis were not attracted to lures that did not have 13–14:OAc as a component. The analog 14:OAc, although present in volatiles collected from crushed abdomens of females, appeared to be inactive, neither increasing or decreasing attraction of males. In addition, 14:OAc elicited minimal responses from antennae of males in GC-EAD bioassays, further suggesting that it is not a crucial component of the pheromone. Thus, 14:OAc may have a role as an inhibitory compound preventing cross-attraction of congeners that might also use 13–14:OAc as a pheromone component, or it may simply be present as a biosynthetic precursor to the pheromone or even an artifact. Our analyses were based on volatiles collected from crushed abdomens rather than the volatiles actually released by calling females, because none of the live females emitted detectable amounts of these or related compounds under laboratory conditions.
In tests of analogs of 13–14:OAc, the TABM pheromone blend was not attractive to M. communis, and E11-14:OAc elicited no responses from antennae of males in GC-EAD assays. In contrast, E11, 13–14:OAc did elicit EAD responses, but if anything, the field bioassay data from 2017 suggested that it might be inhibitory (Fig. 4). Thus, the EAD responses seen may have been a result of the structural similarity between E11, 13–14:OAc and the actual pheromone, 13–14:OAc.
In our initial trials in which test pheromone blends were deployed in plastic PCR tubes with a pinhole in the cap, only low numbers of beetles were caught, suggesting that the release rate of these dispensers might be too low. This result was corroborated in subsequent trials in which either small plastic bags or rubber septum dispensers loaded with similar doses of pheromone attracted substantially more male beetles than the PCR tube dispensers. A dose-response bioassay testing a semi-log range of doses showed that the 10 mg dose, dispensed from plastic bags, was significantly more attractive than any of the lower doses. In a follow-up field trial, the plastic bag dispensers were compared to the rubber septa which have found widespread use as pheromone dispensers for lepidopteran species, with both dispensers loaded with 10 mg of pheromone. Under typical summer climatic conditions, the septa appeared to have longer and more consistent field lifetimes than the polyethylene bags. Specifically, the two dispenser types attracted similar numbers of beetles for the first two weeks after deployment, after which the attractiveness of the plastic bags appeared to decrease more quickly than that of the rubber septa. Five weeks after deployment (i.e., late July), both lure types attracted minimal numbers of beetles, but this may have been due to the natural seasonal decline in M. communis reproductive activity.
During our field trials, conducted over several years with a number of potential pheromone candidates for Melanotus species, we saw no evidence of cross attraction of any other elaterid species to 13–14:OAc, nor did we see any evidence of attraction to any of the analogs or homologs that were tested. These data indicate that 13–14:OAc may be species-specific to M. communis, especially because a number of other Melanotus species were detected at our field sites using other monitoring methods. In fact, it was surprising that no other species were attracted to any of these compounds or blends given that many closely related elaterids have the same or similar pheromone components1,2,39,40,41. In sum, these results suggest that the pheromone components or pheromone blends of Melanotus and other click beetle species may be both quite diverse, but also narrowly tuned, with each species using either relatively unique compounds, or very specific ratios of compounds that may be shared by more than one species.
Our study is the first identification of a sex pheromone for a North American Melanotus species. Of the two other species in the genus for which pheromones have been fully reported, both from Japan, both produce esters of 12-carbon saturated and unsaturated alcohols. Thus, dodecyl acetate (12:OAc) was identified as the single sex pheromone component of M. okinawensis23,42, whereas a blend of (E)-9,11-dodecadienyl butanoate and (E)-9,11-dodecadienyl hexanoate were identified as sex pheromone components of M. sakishimensis24,42. (E)-9,11-Dodecadienyl butanoate and (E)-9,11-dodecadienyl hexanoate have also been identified in extracts of M. tamsuyensis, although bioassay data have not been reported25. Published observations in Europe suggest that M. punctolineatus (Pelerin) may use a tetradecenyl butyrate as a pheromone, whereas M. rufipes (Herbst) may use a tetradecadienyl butyrate43. Pheromone glands of the west-Eurasian M. fusciceps and the Holarctic M. castanipes contained mixtures of > 22 compounds, which included esters of 12-carbon and 14-carbon saturated and unsaturated alcohols, although the exact structures were not identified, nor were any bioassay data reported22. Thus, the compounds that we identified from M. communis in the present study are clearly similar to the pheromones or pheromone candidates found in other Melanotus species from Japan and Europe, especially the 14-carbon compounds of the west-Eurasian beetles. Further studies of the sex pheromones of additional Melanotus species should provide a more complete picture of the chemical diversity of the sex pheromones within this taxonomically large genus.
Insect sex pheromones have been exploited for pest management for more than 50 years, including the pheromones of click beetles. In Europe, pheromone-based methods have been particularly valuable for monitoring pest species44,45,46, as well as threatened non-pest species47. Pheromone-based methods also have been used to monitor several invasive Eurasian click beetle species, i.e., Agriotes spp., that are now established in Canada and the U.S.48. In addition, sex pheromones were exploited in management of pest species, including mass trapping20 and mating disruption21 of M. okinawensis, and may have potential in attract-and-kill strategies using entomopathogens49. Given that the pheromone of M. communis consists of a single component that can be readily synthesized, development of its pheromone for monitoring and control purposes may be both possible and economically feasible.
Ethical approval
The laboratory research was conducted at the University of California, Riverside, and field bioassays were conducted at the USDA-ARS U.S. Vegetable Laboratory and Clemson University Coastal Research and Extension Center in Charleston. All methods met the ethical requirements of the respective university and the United States Department of Agriculture, and followed guidelines of the Committee of Publication Ethics. This article does not involve any studies with human participants or vertebrate animals.
Data availability
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Tóth, M. Pheromones and attractants of click beetles: an overview. J. Pest Sci. 86, 3–17 (2013).
Serrano, J. S., Collignon, R. M., Zou, Y. & Millar, J. G. Identification of sex pheromones and sex pheromone mimics for two North American click beetle species (Coleoptera: Elateridae) in the Genus Cardiophorus Esch. J. Chem. Ecol. 44, 327–338 (2018).
Johnson, P. J. Family 58. Elateridae Leach 1815. In: Arnett Jr., R. H., Thomas, M. C., Skelley, P. E. & Frank, J. H. (eds) American Beetles Volume 2. Polyphaga: Scarabaeoidea through Curculionoidea. CRC Press, Boca Raton, FL, USA, pp 160–173 (2002).
Marske, K. A. & Ivie, M. A. Beetle fauna of the United States and Canada. Coleopt. Bull. 57, 495–503 (2003).
Jacobson, M., Lilly, C. E. & Harding, C. Sex attractant of sugar beet wireworm: identification and biological activity. Science 159, 208–210 (1968).
Onsager, J. A., McDonough, L. M. & George, D. A. A sex pheromone in the Pacific coast wireworm. J. Econ. Entomol. 61, 691–693 (1968).
Butler, L. I., McDonough, L. M., Onsager, J. A. & Landis, B. J. Sex pheromones of the Pacific coast wireworm, Limonius canus. Environ. Entomol. 4, 229–230 (1975).
Riley, T. J. & Keaster, A. J. Wireworms associated with corn: identification of nine species of Melanotus from the north central states. Ann. Entomol. Soc. Am. 72, 408–414 (1979).
Hall, D. G. Insects associated with sugarcane in Florida. Fla. Entomol. 71, 138–150 (1988).
Hall, D. Stand and yield losses in sugarcane caused by the wireworm Melanotus communis (Coleoptera: Elateridae) infesting plant cane in Florida. Fla. Entomol. 73, 298–302 (1990).
Jansson, R. K. & Seal, D. R. Biology and management of wireworms on potato. In: Zehnder, G. W., Powelson, M. L., Jansson, R. K. & Raman, K. V. (eds) Advances in potato pest management. APS Press, St. Paul, MN, USA. pp 31–53 (1994).
Lefko, S. A., Pedigo, L. P., Rice, M. E. & Batchelor, W. D. Wireworm (Coleoptera: Elateridae) incidence and diversity in Iowa conservation reserve environments. Environ. Entomol. 27, 312–317 (1998).
Kuhar, T. P. et al. Current status of insecticidal control of wireworms in potatoes. Pestic. Outlook 14, 265–267 (2003).
Larsen, N. A., Nuessly, G. S. & Cherry, R. H. Efficacy of pyrethroid and neonicotinoid insecticides for Melanotus communis (Gyll.) (Coleoptera: Elateridae) control in Florida sugarcane (Saccharum spp.). J. Appl. Entomol. 140, 517–526 (2016).
Larsen, N. A., Nuessly, G. S., Cherry, R. H. & Glaz, B. Varietal susceptibility to the corn wireworm Melanotus communis (Coleoptera: Elateridae) in sugarcane. J. Pest Sci. 86, 91–98 (2013).
Villani, M. G. & Gould, F. Screening of crude plant extracts as feeding deterrents of the wireworm, Melanotus communis. Entomol. Exp. Applic. 37, 69–75 (1985).
Villani, M. G. & Gould, F. Butterfly milkweed as a feeding deterrent of the wireworm, Melanotus communis. Entomol. Exp. Applic. 37, 95–100 (1985).
Cherry, R. & Nuessly, G. Repellency of the biopesticide, azadirachtin, to wireworms (Coleoptera: Elateridae). Fla. Entomol. 93, 52–55 (2010).
Cherry, R., Taylor, J., Sherrod, D. & Karounos, M. Corn seed treatments for control of the corn wireworm (Coleoptera: Elateridae). J. Entomol. Sci. 52, 161–168 (2017).
Arakaki, N. et al. Control of the sugarcane click beetle Melanotus okinawensis Ôhira (Coleoptera: Elateridae) by mass trapping using synthetic sex pheromone on Ikei Island, Okinawa, Japan. Appl. Entomol. Zool. 43, 37–47 (2008).
Arakaki, N. et al. Mating disruption for control of Melanotus okinawensis (Coleoptera: Elateridae) with synthetic sex pheromone. J. Econ. Entomol. 101, 1568–1574 (2008).
Yatsynin, V. G., Rubanova, E. V. & Okhrimenko, N. V. Identification of female-produced sex pheromones and their geographical differences in pheromone gland extract composition from click beetles (Col., Elateridae). J. Appl. Entomol. 120, 463–466 (1996).
Tamaki, Y., Sugie, H., Nagamine, M. & Kinjyo, M. Female sex pheromone of the sugarcane wireworm Melanotus okinawensis Ôhira (Coleoptera: Elateridae). Jpn. Kokai Tokkyo Koho JP 61–12601 (1986).
Tamaki, Y., Sugie, H., Nagamine, M. & Kinjo, M. 9,11-Dodecadienyl butyrate and 9,11-dodecadienyl hexanoate female sex pheromone of the sugarcane wireworm Melanotus sakishimensis Ôhira (Coleoptera: Elateridae). Jpn. Kokai Tokkyo Koho JP, 2–53753 (1990).
Yen, Y. P. & Chen, P. H. Simple and stereoselective synthesis of attractant of the sugarcane wireworm Melanotus tamsuyensis. Syn. Comm. 28, 4561–4569 (1998).
Nagamine, M. & Kinjo, M. Report on the establishment of the control technique for the sugarcane wireworm Melanotus okinawensis. Okinawa-ken Togyo Sinkou Kyokai, Naha (in Japanese) (1990).
Kishita, M., Arakaki, N., Kawamura, F., Sadoyama, Y. & Yamamura, K. Estimation of population density and dispersal parameters of the adult sugarcane wireworm, Melanotus okinawensis Ôhira (Coleoptera: Elateridae), on Ikei Island, Okinawa, by mark-recapture experiments. Appl. Entomol. Zool. 38, 233–240 (2003).
Arakaki, N. et al. Estimation of abundance and dispersal distance of the sugarcane click beetle Melanotus sakishimensis Ôhira (Coleoptera: Elateridae) on Kurima Island, Okinawa, by mark-recapture experiments. Appl. Entomol. Zool. 43, 409–419 (2008).
Weires, R. W. Evidence of an attractant for the click beetle, Melanotus depressus, in the pheromones of two Platynota spp. (Lepidoptera: Tortricidae). Environ. Entomol. 5, 920–922 (1976).
Brown, E. A. & Keaster, A. J. Field response of Melanotus depressus and M. verberans (Coleoptera: Elateridae) to flight traps and to pheromone of tufted apple budmoth (Platynota idaeusalis Walker) (Lepidoptera: Tortricidae). J. Kansas Entomol. Soc. 56, 47–49 (1983).
Hill, A., Cardé, R., Comeau, A., Bode, W. & Roelofs, W. Sex pheromones of the tufted apple bud moth (Platynota idaeusalis). Environ. Entomol. 3, 249–252 (1974).
Quate, L. W. & Thompson, S. E. Revision of click beetles of genus Melanotus in America north of Mexico (Coleoptera: Elateridae). Proc. US Nat. Mus. Smithsonian Inst. no. 3568. 121, 1–84 (1967).
Marques, F. A., McElfresh, J. S. & Millar, J. G. Kováts retention indexes of monounsaturated C12, C14, and C16 alcohols, acetates and aldehydes commonly found in lepidopteran pheromone blends. J. Braz. Chem. Soc. 11, 592–599 (2000).
Williams, L. III & O’Keeffe, L. E. An ultrasonic procedure for cleaning sticky-trapped arthropods. J. Kansas Entomol. Soc. 63, 461–462 (1990).
van Herk, W. G., Vernon, R. S. & Borden, J. H. A pheromone-baited pitfall trap for monitoring Agriotes spp. click beetles (Coleoptera: Elateridae) and other soil-surface insects. J. Ent. Soc. British Columbia 115, 101–103 (2018).
Peck, J. E. Multivariate Analysis for Ecologists: Step-by-Step. 2nd Edition. MjM Software Design, Gleneden Beach, OR, USA (2016).
McCune, B., Grace, J. B. & Urban, D. L. Analysis of Ecological Communities. MjM Software Design, Gleneden Beach, OR, USA (2002).
Benjamini, Y. & Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. Royal Stat. Soc. B 57, 289–300 (1995).
Tolasch, T., König, C., von Fragstein, M. & Steidle, J. L. M. Identification of the sex pheromone of Idolus picipennis (Bach, 1852) revealed the presence of a cryptic sibling species. J. Chem. Ecol. 39, 1433–1440 (2013).
König, C., Steidle, J. M. L. & Tolasch, T. Decrypting cryptic click beetle species by analysis of sex pheromones. J. Chem. Ecol. 41, 740–745 (2015).
König, C., Szallies, A., Steidle, J. L. M. & Tolasch, T. Sex pheromone of the rare click beetle Betarmon bisbimaculatus. J. Chem. Ecol. 42, 55–59 (2016).
Iwanaga, S. & Kawamura, F. Trapping efficacy of funnel-vane and water pan traps baited with synthetic sex pheromone of the sugarcane wireworms, Melanotus sakishimensis Ôhira and M. okinawensis Ôhira (Coleoptera: Elateridae). Appl. Entmol. Zool. 35, 283–285 (2000).
Tolasch, T., von Fragstein, M. & Steidle, J. L. M. Sex pheromone of Elater ferrugineus L. (Coleoptera: Elateridae). J. Chem. Ecol. 33, 2156–2166 (2007).
Kudryavtsev, I., Siirde, K., Laats, K., Ismailov, V. & Pristavko, V. Determination of distribution of harmful click beetle species (Coleoptera, Elateridae) by synthetic sex-pheromones. J. Chem. Ecol. 19, 1607–1611 (1993).
Blackshaw, R. P. & Vernon, R. S. Spatiotemporal stability of two beetle populations in non-farmed habitats in an agricultural landscape. J. Appl. Ecol. 43, 680–689 (2006).
Vernon, R. S. & Tóth, M. Evaluation of pheromones and a new trap for monitoring Agriotes lineatus and Agriotes obscurus in the Fraser Valley of British Columbia. J. Chem. Ecol. 33, 345–351 (2007).
Svensson, G. P. et al. Chemical ecology and insect conservation: optimising pheromone-based monitoring of the threatened saproxylic click beetle Elater ferrugineus. J. Insect Conserv. 16, 549–555 (2012).
Vernon, B., Lagasa, E. & Philip, H. Geographic and temporal distribution of Agriotes obscurus and A. lineatus (Coleoptera: Elateridae) in British Columbia and Washington as determined by pheromone trap surveys. J. Ent. Soc. British Columbia 98, 257–266 (2001).
Kabaluk, T. J., Lafontaine, J. P. & Borden, J. H. An attract and kill tactic for click beetles based on Metarhizium brunneum and a new formulation of sex pheromone. J. Pest Sci. 88, 707–716 (2015).
Acknowledgements
We are grateful to T. Whitwell and B. Ward (Clemson University, Coastal Research and Extension Center) for facilitating access to some of the field sites, and to W. van Herk and R. Vernon (Agriculture and Agri-Food Canada) for providing the Vernon Pitfall Traps®. We thank E. Anthony, L. Lawrence, and J. Webb for technical assistance. This work was supported by USDA-ARS appropriated funding, by the Robert van den Bosch Scholarship to JMS, and by a UC Riverside CORE Academic Senate research grant to JGM. PJJ was supported through the South Dakota Agricultural Experiment Station. This article reports the results of research only. Mention of a proprietary product does not constitute an endorsement or a recommendation by the USDA for its use. The USDA is an equal opportunity provider and employer. The U.S. Government has the right to retain a non-exclusive, royalty-free license in and to any copyright of this article.
Author information
Authors and Affiliations
Contributions
L.W., J.G.M., and J.M.S. conceived the study and designed the experiments. L.W. collected the beetles, and J.M.S. and J.G.M. conducted the collections of volatiles, pheromone identification, electrophysiology, and pheromone syntheses. L.W. conducted field bioassays and data analysis. P.J.J. contributed to design of field bioassays, and identified the beetles. All authors contributed to the writing and review of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Williams, L., Serrano, J.M., Johnson, P.J. et al. 13-Tetradecenyl acetate, a female-produced sex pheromone component of the economically important click beetle Melanotus communis (Gyllenhal) (Coleoptera: Elateridae). Sci Rep 9, 16197 (2019). https://doi.org/10.1038/s41598-019-52199-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-019-52199-1
- Springer Nature Limited