
Abstract
Mandarin fish Siniperca chuatsi, a valuable piscivorous fish, have been stocked into many lakes in China since the 1990s. This study did the first attempt to evaluate the ecological effects of hatchery-reared mandarin fish stocking in the Yangtze River basin lakes. Our study demonstrated a significant change in fish community composition after mandarin fish stocking, but no fish extinction was observed. No significant difference was observed in the total density of 13 forage fish before and after mandarin fish stocking, but the total biomass showed a significant decline after mandarin fish stocking. Significant differences in length-frequency distributions were observed for Carassius auratus, Pseudorasbora parva and Toxabramis swinhonis captured before and after stocking mandarin fish. No significant change in habitat distribution was detected before and after mandarin fish stocking. A marked decline in total nitrogen and a slight decline in total phosphorus were observed while a slight increasing trend for Secchi depth was found after stocking. Our findings suggested that mandarin fish stocking can increase predation pressure on forage fish and subsequently optimize the food web structure. Also, mandarin fish stocking has the potential to improve water quality and may be a feasible strategy to alleviate eutrophication of shallow Yangtze lakes.
Similar content being viewed by others


Introduction
Stock enhancement is a fisheries management approach involving the release of cultured organisms to increase abundance and yield of natural fish or invertebrate stocks1. In general, successful stock enhancement can improve socioeconomic outcomes by (1) creating new economic opportunities for fisheries-related livelihoods, (2) increasing skills of local communities, and (3) providing high-quality food fish and protein2,3. However, stocking or introduction of predators, especially piscivorous fish, may have strong impacts on freshwater ecosystems through predation, competition, mixing of exotic genes, habitat modification, and the introduction of pathogens4,5,6,7. When predation is intense, predators create both direct and indirect effects that cascade down the food web affecting both food web structure and biodiversity8,9,10,11,12. Direct predation on fish can change the species composition and size structure of the fish community at lower trophic levels and reduce population densities13,14. Indirect effects include habitat changes associated with predator avoidance behavior and increases of emigration rates of forage fish13,15. Therefore, understanding the changes on prey fish abundance and cascading effects down to lower trophic levels is essential for evaluating the ecological effects of stocking piscivorous fish in freshwater ecosystems.
Piscivorous fish stocking can be used as a lake restoration tool to improve water quality by creating a trophic cascade16,17. Direct predation on zooplanktivorous fish can initiate a trophic cascade that reduces predation pressure on zooplankton, especially on large cladocerans, leading to increased grazing on planktonic algae18,19,20. Predators may also reduce benthivorous fish, leading to a decrease in suspended material in the water column, thereby increasing Secchi depth21. Based on this evidence, stocking piscivorous fish in eutrophic lakes is commonly referred to as biomanipulation and is a possible strategy to alleviate eutrophication19.
Mandarin fish, or Chinese perch, Siniperca chuatsi (Basilewsky), is one of the most important commercial freshwater fish inhabiting Asian waters from the south in the Zhujiang River system to the north in the Amur River system22. The fish is a piscivore with specialized feeding habits, feeding only on live fish or shrimp throughout its life23,24,25. In the past several decades, mandarin fish populations in the Yangtze lakes have declined due to overfishing and habitat loss25. Thus, hatchery-reared mandarin fish have been stocked since the 1990s to increase populations25, to pursue commercial returns26, and to ease conflicts between fishery development and water quality conservation, based on the principle of trophic cascades16. By far, the area of lakes and reservoirs stocked mandarin fish exceeds 133000 ha in the middle and lower basins of the Yangtze River. The economic benefits of mandarin fish stocking in several lakes is well documented26, and studies on stocked mandarin fish have focused on survival, growth, recapture rate and foraging success27,28,29. In contrast, relatively little is known about the impacts of stocking mandarin fish on the ecology of Yangtze Lakes.
Stocked mandarin fish always incorporate both fish and shrimp into their diets25 and thus can alter the composition, densities, size distributions, and niche characteristics of wild fish populations directly or indirectly through predation or competition6,15,19,30. There is some evidence that stocked mandarin fish do not compete with wild Siniperca fishes during the critical periods of the early stocking stages25, but we know of no research that has explicitly focused on stocked mandarin fish impacts on wild fish community at lower trophic levels in lakes. To address this, we used a before-after study design to evaluate impacts of mandarin fish stocking on the wild forage fish community. Specifically, we compared the composition, abundance, size structure and spatial patterns of habitat use of wild forage fish before and after mandarin fish stocking in a shallow Yangtze lake, Central China. We predicted that mandarin fish stocking would significantly change the community structure of wild fish. Also, water quality was compared before and after mandarin fish stocking to evaluate whether piscivorous fish stocking can improve water quality through trophic cascade effects in shallow lakes along the Yangtze River.
Results
Mandarin fish density and biomass
After six years continuous hatchery-reared mandarin fish stocking, their density and biomass showed an apparent increase trend. The density and biomass of mandarin fish from 2012 to 2014 were significantly higher than those of 2009 before stocking (all P < 0.001, Table 1).
Fish composition and abundance
A total of 19 fish species were collected by multiple mesh-sized gill nets from 2007 to 2014 in Biandantang Lake, of which 13 species were found in stomach contents of mandarin fish25. In 2007–2008, prior to the mandarin fish stocking, Toxabramis swinhonis, Sharpbelly Hemiculter leucisculus, Culter dabryi, crucian carp Carassius auratus and topmouth gudgeon Pseudorasbora parva were the dominant species in forage fish community, with an average numerical percentages of 24%, 21%, 14%, 10% and 10%, respectively. While in 2010–2014, after the mandarin fish stocking, T. swinhonis dominated the net samples with an average numerical percentage of 77%, followed by H. leucisculus (6%) and C. dabryi (6%). The percentage of C. auratus and P. parva declined gradually, and only accounted for 1.1% and 0.25% of the total number in 2014, respectively (Table 2). Although T. swinhonis, H. leucisculus, C. dabryi, C. auratus and P. parva were the most commonly captured forage fish before and after mandarin fish stocking in Biandantang Lake, the community composition of forage fish differed significantly between periods (χ2 = 63.65, df = 12, P < 0.001). Comparing the food habits of 13 forage fishes31,32, we found all 13 fish species feed more or less on zooplankton. Among them, the larvae of C. auratus and C. dabryi, and P. parva mainly feed on zooplankton (Table 2).
Forage fish density and biomass
The total density of 13 forage fish ranged from 0.143 to 0.171 individuals∙m−2∙h−1 during our study period, and no significant difference was observed before and after mandarin fish stocking (Mann-Whitney test, P = 0.081) (Fig. 1a). However, there was a significant difference in the total biomass of 13 forage fish before (mean: 2.59 g∙m−2∙h−1) and after (mean ± SD: 1.57 ± 0.31 g∙m−2∙h−1) mandarin fish stocking (Mann-Whitney test, P < 0.001). The total biomass ranged from 1.275 to 2.637 g∙m−2∙h−1 and show an apparent declining trend after mandarin fish stocking (Fig. 1b). For several dominant forage fish, a significant decreased trend was observed in the density and biomass of C. auratus, H. leucisculus, C. dabryi and Rhodeus spp. following mandarin fish stocking (Mann-Whitney test, P < 0.001). In contrast, the density and biomass of T. swinhonis showed an apparent increase after mandarin fish stocking, respectively (Mann-Whitney test, P < 0.001) (Fig. 1).

Changes in forage fish density (a) and biomass (b) before and after mandarin fish stocking in Biandantang Lake.
Size structure of forage fish
Significant differences in the length-frequency distributions were found for C. auratus (K-S test, Dmax = 0.365, P < 0.001), P. parva (K-S test, Dmax = 0.442, P < 0.001), and T. swinhonis (K-S test, Dmax = 0.320, P < 0.001) captured before and after stocking mandarin fish in Biandantang Lake, but no significant difference was observed for H. leucisculus (K-S test, Dmax = 0.034, P = 0.064) and C. dabryi (K-S test, Dmax = 0.020, P = 0.103) (Fig. 2). Much larger C. auratus and P. parva and much smaller T. swinhonis were found in Biandantang Lake after mandarin fish stocking compared to before stocking. Indeed, mean total lengths of C. auratus and P. parva after mandarin fish stocking were significantly larger than those before stocking (t-test, C. auratus: t234 = −5.264, P < 0.001; P. parva: t256 = −5.403, P < 0.001), while mean total lengths of T. swinhonis were significantly small after mandarin fish stocking (t-test, t2076 = 18.426, P < 0.001) (Fig. 2).

Total length frequency histograms for C. auratus (a) P. parva (b) H. leucisculus (c) C. dabryi (d) T. swinhonis (e) captured before (2007–2008, dark bars) and after (2010–2014, open bars) stocking mandarin fish in Biandantang Lake.
Habitat distribution
Both before and after mandarin fish stocking, forage fish were predominantly captured in nearshore habitat in similar proportions (percentage of total catch: before: \(\bar{\chi }\) = 60.8 ± 1.3%; after: \(\bar{\chi }\) = 54.3 ± 4.8%, Fig. 3). The percentages of forage fish were significantly higher nearshore than offshore during both periods (F = 35.424, P < 0.001), but those differences declined after stocking as indicated by significant interactions between treatment and habitat (F = 6.584, P = 0.028). For C. auratus and C. dabryi, the percentage in nearshore habitat was significantly higher than those in offshore habitat prior to stocking (C. auratus: F = 39.718, P < 0.001; C. dabryi: F = 32.559, P < 0.001), but no significant difference was observed after stocking (C. auratus: F = 0.078, P = 0.124; C. dabryi: F = 0.066, P = 0.298, Fig. 3). For T. swinhonis and H. leucisculus, the percentage in nearshore habitat was lower than in offshore habitat before stocking, but became higher nearshore than offshore after stocking. For P. parva and Rhodeus spp., the percentage nearshore was significantly higher than offshore both before and after mandarin fish stocking (before: P. parva, F = 45.114, P < 0.001; Rhodeus, F = 40.359, P < 0.001; after: P. parva, F = 23.581, P < 0.001; Rhodeus, F = 25.441, P < 0.001), but the ratio of the percentage in nearshore to offshore habitat before stocking was lower than after stocking (Fig. 3).

Mean (SD) percentages of total catch of forage fish at two different habitats (nearshore and offshore) before (2007–2008) and after (2010–2014) mandarin fish stocking in Biandantang Lake.
Water quality
The concentration of total nitrogen (TN) was significantly reduced after mandarin fish stocking (Mann-Whitney test, P < 0.001, Fig. 4a). Total phosphorus (TP) concentration showed more variability after stocking, but no apparent trend between mean values 0.056 mg L−1 during 2006–2008, to 0.048 mg L−1 during 2010–2014 (Mann-Whitney test, P = 0.349, Fig. 4b). Secchi depth was similar before and after mandarin fish stocking despite two years of improved visibility immediately following the start of stocking (2010 to 2011) (before: \(\bar{\chi }\) = 55.2 ± 4.8 cm; after: \(\bar{\chi }\) = 58.9 ± 8.6 cm. Mann-Whitney test, P = 0.370, Fig. 4c). Chl a concentration in 2010 increased immediately following stocking, but the means were not different before and after stocking (Mann-Whitney test, P = 0.410, Fig. 4d).

Changes in concentrations of (a) total nitrogen (TN), (b) total phosphorous (TP), (c) chlorophyll-a (Chl. a) and (d) Secchi depth before (open bars) and after (dark bars) mandarin fish stocking in Biandantang Lake.
Discussion
Apparent effects of top predators on lower trophic levels in freshwater ecosystems are common, and stocking or introductions of top predators into new ecosystems can reduce prey fish abundance and cause cascading effects through lower trophic levels19,20,33,34. Our study in Biandantang Lake demonstrated a significant change in forage fish community composition from one with similar proportions of T. swinhonis, H. leucisculus, C. dabryi, C. auratus and P. parva prior to mandarin fish stocking to a community dominated by T. swinhonis after stocking. The observation of reduced species diversity following stocking with a top piscivore is consistent with previous studies for piscivorous fish in North American and European countries10,11,20,34, but this is the first evidence that mandarin fish stocking exert strong effects on the community structure of wild forage fish in the shallow Yangtze lakes. The observed changes in forage fish community in our study was mainly embodied with the apparent difference in percentage of abundance of different forage fish, as no reduction of species richness of forage fish was observed after mandarin fish stocking. This was probably due to the wide food spectrum and selective predation of mandarin fish24,25.
Our before- and after-stocking comparative study provided no evidence that mandarin fish stocking reduced the total densities of wild forage fish populations, nor did the richness of wild forage fish decline, despite the well documented negative impacts that stocked piscivores have on many wild species10,35. Some factors could contribute to why piscivores stocking did not lead to reductions in total densities of forage fish: (i) feeding behaviour of predators, (ii) prey fish species and availability in environment, (iii) morphology and behavioural responses of forage fish, and (iv) habitat heterogeneity15,35. Unlike oligotrophic alpine lakes, mesotrophic or eutrophic shallow lakes of the Yangtze River basin including Biandantang Lake support an abundance of small-sized fish, which can serve as ample alternative prey for mandarin fish, who as a generalist predator can feed on a wide spectrum of available prey24. Over the initial five years after stocking mandarin fish, populations of these other forage fishes were reduced, but not extirpated. Therefore, the apparent increase of T. swinhonis population compensated for the reduction of other forage fish commonly consumed by mandarin fish, which accounted for the lack of a significant reduction in the total densities of forage fish after mandarin fish stocking.
In contrast to total densities, the total biomass of forage fish showed a significant reduction following stocking of mandarin fish. This reduction was apparently a result of both the direct effects of predation and indirect effects such as habitat shifts in an attempt to avoid predators, and a decrease in the average size of forage fish considered most vulnerable to predation11. The lack of trends in the total densities and biomass could be attributed to variations in community composition of forage fish. The reduction in abundance of relatively large-bodied forage fish species including C. auratus, H. leucisculus and C. dabryi directly caused the reduction of the total biomass of forage fish after mandarin fish stocking in Biandantang Lake.
For several dominant forage fish, their density in Biandantang Lake changed during the years of mandarin fish stocking in accordance with the increased predation pressure from stocked mandarin fish. The density of C. auratus, H. leucisculus, C. dabryi and Rhodeus declined by more than 50% after mandarin fish stocking. Rhodeus and C. auratus were affected more than the other two species, which can be explained by the difference of habitat choice. Rhodeus and C. auratus associate with the nearshore zones more than H. leucisculus, and C. dabryi36. Mandarin fish stocked in the nearshore zone occupied similar deeper depths and had easier access to Rhodeus spp. and C. auratus as a prey. In contrast, H. leucisculus, and C. dabryi occupied the upper layers of the water column further offshore, likely reducing their encounter rate with predators31. For T. swinhonis, one of the dominant forage fish in Biandantang Lake before stocking, its density increased dramatically following stocking of mandarin fish. T. swinhonis have a robust spiny dorsal fin, and thus greater anti-predation ability than the soft fin species31,37.
Apart from the stocked mandarin fish, other factors may cause the change in forage fish community composition after hatchery-reared mandarin fish stocking. Generally, temperature is often considered to be an important factor determining reproductive success and survival for fish19,38. Actually, no unusual weather conditions were occurred during the present study. In addition, the decline in biomass of forage fish might be caused by a fish-kill resulting from low oxygen concentrations, rather than by predation19. However, there is no direct evidence that such a fish kill observed. The fact that crucian carp, a species exceptionally tolerant to low oxygen concentrations, also declined, further indicates that the fish-kill hypothesis is unlikely.
Significant differences in the length-frequency distributions were observed for C. auratus, P. parva, and T. swinhonis captured before and after stocking mandarin fish. This effect was caused by size-selective predation of mandarin fish24. Size-selective predation commonly leads to increased mortality at early life stages of prey24,39 and can consequently alter the size structure of forage fish communities37,40. In this study, the mean total lengths of C. auratus and P. parva after mandarin fish stocking were significantly larger than those prior to stocking, suggesting that their populations are often dominated by larger size classes after mandarin fish stocking. The similar results were seen following stocking of other piscivorous fish, such as trout15,41. These results also indicated that more of the smaller C. auratus (120–159 mm) and P. parva (60–79 mm) were consumed with the increased density of mandarin fish. For forage fish P. parva, mandarin fish consistently preferred to consume individuals of 60–79 mm TL, despite consuming much larger topmouth gudgeon than expected from size distributions in Xiaosihai Lake near to Biandantang Lake24. This indicated that increased mandarin fish may cause increased mortality on 60–79 mm TL individuals of P. parva in the environment. For the forage fish C. auratus, disproportionate consumption of small individuals may result from gape limitations of mandarin fish, because crucian carp is a deep bodied fish and have the largest body depth of the five dominant forage fish over similar lengths. In addition, the mechanisms other than morphological limitations, such as greater abilities of larger individuals to escape predators, via spatial segregation or greater evasive capabilities, should also be taken into consideration. In contrast to C. auratus and P. parva, the mean total lengths of T. swinhonis were significantly smaller after stocking than before. The reduction in modal size and increased abundance was likely due to: (1) The spiny dorsal fin on T. Swinhonis, reduced the risk of occupying the same spatial niche as mandarin fish, as suggested by their low electivity as prey for mandarin fish24,25. (2) The food competition between T. Swinhonis and other zooplanktivores was relaxed substantially because of the reduced density of zooplanktivorous fish.
The nearshore-offshore habitat shifts by varied among species of forage fish after mandarin fish stocking, although no significant difference in the composite percentages of forage fish in nearshore versus offshore zones was observed. Generally, forage fish commonly occurred in or around nearshore zones with complex habitat as refuge and support more food resource36,42,43. We attribute the observed offshore shift by most species to increased predation risk in nearshore zones following stocking of mandarin fish which predominantly utilize nearshore habitats31. Typical nearshore forage fishes like C. auratus, P. parva and Rhodeus spp. shifted to offshore habitats after mandarin fish were stocked. However, T. swinhonis and H. Leucisculus, which were relatively more abundant in offshore habitat than in nearshore habitat before mandarin fish stocking, became the predominant forage fishes in both habitats after stocking. The inconsistent response to increased predation risk can possibly be ascribed to the different anti-predation ability among forage fish species. Although abundant food resources were available in nearshore zones, C. auratus, P. parva and Rhodeus spp. shifted offshore to escape predation. In contrast, with increased food competition offshore, T. Swinhonis and H. Leucisculus further exploited the nearshore zones to feed where the spiny dorsal fin of T. Swinhonis and faster swimming speeds of H. Leucisculus afforded greater anti-predation ability to mandarin fish than the other forage fishes31.
A marked decline in total nitrogen concentration and slight increase in Secchi depth were observed after mandarin fish stocking, suggesting that stocking mandarin fish has the potential to improve water quality of shallow Yangtze lakes. Our findings support Potthoff et al. opinion that stocking piscivorous fish can improve ecological characteristics of shallow lakes20. Generally, there was a positive relationships between chlorophyll-a concentration and concentrations of total nitrogen and total phosphorous44,45,46, however, the chlorophyll-a concentration in the study showed a slight but significant increase after mandarin fish stocking. This may have contributed to the increased density of T. swinhonis that feeds predominantly on insect larvae, copepods31. Although P. parva, juvenile C. auratus and C. dabryi also mainly feed on zooplankton31 and their reduced densities can reduce predation pressure on zooplankton, the significant increase in T. swinhonis may compensate for this reduced predation pressure and even intensify the pressure, and consequently cause the increase of phytoplankton through trophic cascade effects. Despite chlorophyll-a concentration showed a slight increase after mandarin stocking, Secchi depth also raised slightly. This is inconsistent with widespread agreement that Secchi depth generally negatively correlated with chlorophyll-a47,48. The inconsistence may be explained by the change of suspended solids. Previous study demonstrated that suspended solid is also an important factor influencing Secchi depth21. And the important effects of benthivorous fish on sediment resuspension has been frequently observed in other studies21,49. In this study, mandarin fish has specialized feeding habits of only feeding on live fish or shrimp throughout their life, and crucian carp, topmouth gudgeon, sharpbelly were the dominant food in their stomach in many lakes in the middle reach of the Yangtze River24,25. Although there is no direct evidence that suspended solids decreased in our study, the fact that reduced density of benthivorous C. auratus induced by increase predation pressure of mandarin fish after continuous stocking was able to diminish sediment resuspension in Biandantang Lake, seems possibly causing a resultant beneficial effect on Secchi depth.
Another experiment conducted in a highly eutrophic shallow Yangtze lake showed that the ecological status could be rapidly reversed from eutrophic to oligotrophic using an integrity measure combining a ban on fertilizer and a reduction of planktivorous fish stocking along with the introduction of both mandarin fish and Chinese mitten crab50. However, there is direct documentation that stocking piscivorous fish alone is useful for water quality protection in the shallow Yangtze lake, which means it could be a feasible bio-manipulation measure to control eutrophication for these lakes located in the middle and lower Yangtze River. Nowadays, water and food security have been and remain two of the most important priorities for the growing population of China. However, hundreds of lakes on the floodplain of the Yangtze River, like Lake Biandantang, are eutrophic and abundant with low value small-sized fish due to anthropogenic activities26,51. Although catchment-level restoration measures, especially nutrient abatement, were considered to have priority for shallow lake protection52, another study suggested that catchment-level restoration alone has limited potential to mitigate effects of eutrophication in shallow lakes53. So, managers as well as policy makers might consider using bio-manipulation based piscivorous fish stocking, in conjunction with catchment-level restoration measures, to stimulate improvement of ecological characteristics and development of sustainable fisheries in shallow Yangtze lakes.
In conclusions, this study is the first attempt to evaluating the ecological effects of stocking mandarin fish in shallow Yangtze lakes. This study supports the hypothesis that piscivorous fish may be a significant structuring force in shallow eutrophic lakes. Our results suggested that mandarin fish stocking have significant impacts on community composition, total biomass, size distribution and habitat use of forage fish by selective predation. Our data also indicated that stocking mandarin fish has the potential to improve water quality of shallow Yangtze lakes. As no significant difference in total density and no fish extinction was observed after mandarin fish stocking, it is suggested that mandarin fish stocking could be a feasible bio-manipulation measure, in conjunction with catchment-level restoration measures, to alleviate eutrophication for managers in shallow Yangtze lakes. However, the stocking balance needs to be carefully weighed as overstocking could have strong negative effects on fish community by replacing wild top consumers and changing ecosystem configuration through strong cascading effects on lower trophic levels. So, studies on optimizing stocking density and size of the stocked individuals and its long-term improvements should be taken into consideration in future.
Materials and Methods
Study site
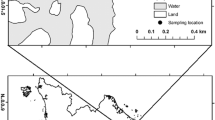
Biandantang Lake (30°15′N, 114°43′E) is located on the south bank of the middle reach of the Yangtze River, Hubei Province, Central China. The lake, with an area of 3.333 km2 and depths ranging from 1.2 to 3.2 m (mean 2.0 m), has been separated from the larger Baoan Lake by a dyke (Fig. 5). Fifteen years ago, Biandantang Lake was heavily covered with submerged macrophytes, with Vallisneria spiralis L., Myriophyllum spicatum L. and Nelunbo nucefera G. the dominant species54. For six years prior to this study, the major fishery had been supported by stocking Chinese mitten crab Eroichier sinensis. Subsequently submerged macrophytes, one of the most important food resources for E. sinensis, gradually decreased due to overstocking of E. sinensis in the past several years. During our study periods, the lake was only sparsely vegetated with M. spicatum and Trapa bispinosa Roxb in a small area of the littoral zone, and was slightly eutrophic. The fish community in the lake comprised 47 fish species belonging to 14 families, of which bighead carp Aristichthys nobilis (Richardson), common carp Cyprinus carpio (Linnaeus), silver carp Hypophthalmichthys molitrix (Valenciennes), crucian carp Carassius auratus (Linnaeus), Culter dabryi and mandarin fish were the most important commercial fish.

Map showing the geographic location and broad outline of Biandantang Lake. The figure was generated by the software Adobe Photoshop, version CS 6 (Adobe Inc., San Jose, California).
Mandarin fish stocking
Each spring since 2009, Biandantang Lake has been stocked with juvenile hatchery-reared mandarin fish at densities of 14–120 individuals∙ha−1 (Table 3). The mandarin fish were hatched and raised in a nearby national fish hatchery. At an age of about 1–2 months, they were transported to the lake and released as much as possible over the littoral zone from a boat. In late June 2009, 4,800 juveniles (mean total length = 69.3 mm), marked with coded wire tags (tags with 0.25 mm diameter and 1 mm length, Northwest Marine Technology (NMT), Shaw Island, Washington) were stocked in order to evaluate the diet composition between hatchery-reared and wild mandarin fish23. From 2010 to 2014, juveniles were stocked in mid-June at a size of 25–35 mm total length.
Fish sampling
The composition and abundance of fish community was determined in October each year in 2007–2008 and during 2010–2014 using multi-mesh Nordic 12 gillnets (Lindeman, Replot, Finland) (stretches mesh sizes 5, 6.25, 8, 10, 12.5, 15.5, 19.5, 24, 29, 35, 43 and 55 mm, that were 30 m long and 1.5 m high). Six sampling sites were selected (three in nearshore zones and three in offshore zones), and two nets were set at each site. The 12 gill nets were set overnight (12–15 h) in the same locations every year. Fishing was conducted over three nights in each year. Catches from each net were processed separately. Captured fish were identified, counted by species, and measured for total length (TL) to the nearest mm, and body weight (BW) to the nearest 0.1 g.
Mandarin fish were quantitatively sampled by electrofishing in June from 2009 to 2014 in order to evaluate the changes of mandarin fish abundance. Electrofishing was conducted in the night (from 20:00 h to 23:00 h) along nearshore zone near to gillnet sampling sites. A total of nine sampling points (each point cover area: 2500 m2) were selected and fishing was conducted at three consecutive nights in each year. Catches from each point were processed separately and counted, and then measured for total length (TL) to the nearest mm, and body weight (BW) to the nearest 0.1 g.
All fish stocking and capturing procedures complied with the animal welfare laws of the Government of China and the ethical rules of the Institutional Animal Care and Use Committee of the Institute of Hydrobiology (Approval ID: Keshuizhuan 08529).
Density and biomass estimation
The fish species that were potential prey of mandarin fish were selected for density and biomass estimation. Catch per unit effort (CUPE) in any 1 year was based on the mean CPUE of the two nets in each of the six zones. Mandarin fish density and biomass were estimated by point-sampling (nine points) electrofishing each year. The CPUE in any 1 year was based on the mean CPUE of nine sampling points. The relative fish density and biomass were determined as number per unit effort (NPUE) and biomass (g) per unit effort (BPUE), respectively.
Size structure of forage fish
For dominant forage fish, two trap-nets were used to assess prey availability and size structure at the same time as gill nets sampling. Each trap-net set had two cod-ends, with 2 m depth and 6 mm nylon mesh. Six trap-net sets were completed in October each year. All forage fish were measured for total length (TL) to the nearest mm, and body weight (BW) to the nearest 0.1 g. We evaluated the effects of mandarin fish stocking on the size structure of dominant forage fish populations by comparing size-frequency distributions and average lengths of forage fish populations before and after mandarin fish stocking.
Water quality
Water was sampled between 2006 and 2014 at around 10:00–12:00 hours in five different zones throughout the lake after gill net sampling in each year. Water Secchi depth was monitored in situ by a Secchi disk. Total nitrogen (TN) and total phosphorus (TP) were analyzed following the ‘Standard methods for the examination of the water and wastewater’55. Chlorophyll-a concentration (Chl. a) was determined using a fluorimeter with methanol extraction of the filtrate56.
Data analysis
The differences in the fish community composition from the Biandantang Lake with respect to year were assessed by a chi-square test (χ2)24. Significant difference in biomass or density before and after mandarin fish stocking was identified using the Mann-Whitney test. The same method was adopted for Secchi depth and concentrations of total phosphorus, total nitrogen and chlorophyll-a. The Kolmogoro-Smirnov (K-S) test was used to examine differences in the length-frequency distribution of five dominant forage fish before and after mandarin fish stocking. A student’s t-test was used to test for the differences in the total length of five dominant forage fish before and after mandarin fish stocking. To test for differences between treatments in distribution of forage fish across the two habitat (nearshore and offshore), we used two-factor ANOVAS with treatment (pre-stocking and post-stocking) and habitat (nearshore and offshore) as the factors, and percentage (arcsine square-root transformed) of fish in each habitat as the response variable. Statistical analysis was carried out with SPSS ver. 16.0 for Windows (SPSS Inc., Chicago, Illinois). Results of statistical tests were considered significant when P < 0.05. Assumptions of normality and homoscedasticity for parametric tests were confirmed in SPSS.
References
Lorenzen, K. Population dynamics and potential of fisheries stock enhancement: practical theory for assessment and policy analysis. Philos. Trans. R. Soc. 360, 171–189 (2005).
Pinkerton, E. Economic and management benefits from the coordination of capture and culture fisheries: the case of Prince William Sound pink salmon. North Am. J. Fish Manage. 14, 262–277 (1994).
Lorenzen, K. & Garaway, C. J. How predictable is the outcome of stocking? FAO Fisheries Technical Paper 374, 133–152 (1998).
Kerfoot, W. C. & Sih, A. Predation: direct and indirect impacts on aquatic communities. Bioscience 38, 39–47 (1987).
Vejřík, L. et al. European catfsh (Silurus glanis) as a freshwater apex predator drives ecosystem via its diet adaptability. Sci. Rep. 7, 15970 (2017).
Weber, E. D. & Fausch, K. D. Interactions between hatchery and wild salmonids in streams: differences in biology and evidence for competition. Can. J. Fish. Aquat. Sci. 60, 1018–1036 (2003).
Allendorf, F. W. et al. Cutthroat trout hybridization and the US Endangered Species Act: one species, two policies. Conserv. Biol. 19, 1326–1328 (2010).
Hasegawa, K., Ohta, T. & Takahashi, S. Are hatchery chum salmon fry a native invader? Direct and indirect effects of stocking salmon fry on stream organisms. Hydrobiologia 806, 111–121 (2018).
Carey, M. P., Sanderson, B. L., Barnas, K. A. & Olden, J. D. Native invaders - challenges for science, management, policy, and society. Front. Ecol. Environ. 10, 373–381 (2012).
Berg, S., Jeppesen, E. & Søndergaard, M. Pike (Esox lucius L.) stocking as a biomanipulation tool. 1. Effects on the fish population in Lake Lyng, Denmark. Hydrobiologia 342/343, 311–318 (1997).
Findlay, C. S., Bert, D. G. & Zheng, L. G. Effect of introduced piscivores on native minnow communities in Adirondack lakes. Can. J. Fish. Aquat. Sci. 57, 570–580 (2000).
Davis, M. A. Biotic globalization: does competition from introduced species threaten biodiversity? Bioscience 53, 481–489 (2003).
He, X. & Kitchell, J. F. Direct and indirect effects of predation on a fish community: a whole lake experiment. Trans. Am. Fish. Soc. 119, 825–835 (1990).
Bronmark, C., Paszkowski, C. A., Tonn, W. M. & Hargeby, A. Predation as a determinant of size structure in populations of crucian carp (Carassius carassius) and tench (Tinca tinca). Ecol. Freshw. Fish. 4, 82–92 (1995).
Nasmith, L. E., Tonn, W. M., Paszkowski, C. A. & Scrimgeour, G. J. Effects of stocked trout on native fish communities in boreal foothills lakes. Ecol. Freshw. Fish 19, 279–289 (2010).
Liere, L. V. & Gulati, R. D. Restoration and recovery of shallow eutrophic lake ecosystems in the Netherlands: epilogue. Hydrobiologia 233, 283–287 (1992).
Skov, C. Pike stocking for lake restoration. In Skov, C. & Nilsson, P. A., editors, Biology and Ecology of Pike. pp. 269–288. CRC Press (2018).
Lauridsen, T. L., Jeppesen, E. & Søndergaard, M. Colonization and succession of submerged macrophytes in shallow Lake Væng during the first five years following fish manipulation. Hydrobiologia 275/276, 233–242 (1994).
Skov, C., Perrow, M. R., Berg, S. & Skovgaard, H. Changes in the fish community and water quality during seven years of stocking piscivorous fish in a shallow lake. Freshw. Biol. 47, 2388–2400 (2002).
Potthoff, A. J. et al. Cascading food-web effects of piscivore introductions in shallow lakes. J. Appl. Ecol. 45, 1170–1179 (2008).
Breukelaar, A. W., Lammens, E. H. R. R., Breteler, J. G. P. K. & Tatrai, I. Effects of benthivorous bream (Abramis brama) and carp (Cyprinus carpio) on sediment resuspension and concentrations of nutrients and chlorophyll a. Freshw. Biol. 32, 113–121 (1994).
Liang, X., Oku, H., Ogata, H. Y., Liu, J. & He, X. Weaning Chinese perch Siniperca chuatsi (Basilewsky) onto artificial diets based upon its specific sensory modality in feeding. Aquac. Res. 32, 76–82 (2001).
Liang, X., Liu, J. & Huang, B. The role of sense organs in the feeding behaviour of Chinese perch. J. Fish Biol. 52, 1058–1067 (1998).
Li, W., Zhang, T., Ye, S., Liu, J. & Li, Z. Feeding habits and predator-prey size relationships of mandarin fish Siniperca chuatsi (Basilewsky) in a shallow lake, central China. J. Appl. Ichthyol. 29, 56–63 (2013).
Li, W. et al. Does hatchery-reared Siniperca chuatsi (Actinopterygii, Perciformes) compete significantly with two wild Siniperca populations for diets in a shallow lake? Hydrobiologia 741, 125–138 (2014).
Cui, Y. & Li, Z. Fishery Resources and Conservation of Environment in Lakes of the Yangtze River Basin. Science Press (2005).
Zhang, B. Effects of stocking enhancement and genetic diversity of mandarin fish (Siniperca chuatsi) in shallow lakes along the middle and lower reaches of Yangtze River. Doctoral dissertation (2007).
Li, W. Ecological studies on the stocking fishery of mandarin fish Siniperca chuatsi (Basilewsky) in shallow lakes along the middle reach of the Yangtze River. Doctoral dissertation (2011).
Li, W. et al. Effects of turbidity and light intensity on foraging success of juvenile mandarin fish Siniperca chuatsi (Basilewsky). Environ. Biol. Fishes 96, 995–1002 (2013).
Townsend, C. R. Invasion biology and ecological impacts of brown trout Salmo trutta in New Zealand. Biol. Conserv. 78, 13–22 (1996).
Fish Research Laboratory of Institute of Hydrobiology (FRLIH). Fish of the Yangtze River. Science Press (1976).
Zhang, T. Life-history strategies, trophic patterns and community structure in the fishes of Lake Biandantang. Doctoral dissertation (2005).
Carpenter, S. R. & Kitchell, J. F. The Trophic Cascade in Lakes. Cambridge University press (1993).
Findlay, D. L. et al. Dynamics of a boreal lake ecosystem during a long-term manipulation of top predators. Ecosystems 8, 603–618 (2005).
Guillerault, N. et al. Does the non-native European catfish Silurus glanis threaten French river fish populations? Freshw. Biol. 60, 922–928 (2015).
Li, W., Zhang, T. & Li, Z. Spatial distribution and abundance of small fishes in Xiaosihai Lake, a shallow lake along the Changjiang (Yangtze) River, China. Chin. J. Oceanol. Limnol. 28, 470–477 (2010).
Tonn, W. M. & Paszkowski, C. A. Size-limited predation, winterkill, and the organization of Umbra-Perca fish assemblages. Can. J. Fish. Aquat. Sci. 43, 194–202 (1986).
Mills, C. A. Reproduction and life history. In Winfield, I. J. & Nelson, J. S., editors, Cyprinid Fishes: Systematics, Biology and Exploitation. pp. 483–508. Chapman & Hall, London (1991).
Werner, E. E. & Gilliam, J. F. The ontogenetic niche and species interactions in size-structured populations. Annu. Rev. Ecol. Evol. Syst. 15, 393–425 (1984).
Heibo, E. & Magnhagen, C. Variation in age and size at maturity in perch (Perca fluviatilis L.), compared across lakes with different predation risk. Ecol. Freshw. Fish 14, 344–351 (2005).
McIntosh, A. R., Crowl, T. A. & Townsend, C. R. Size-related impacts of introduced brown trout on the distribution of native common river galaxias. Z. J. Mar. Freshw. Res. 28, 135–144 (1994).
Hall, D. J. & Werner, E. E. Seasonal distribution and abundances of fishes in the littoral zone of a Michigan lake. Trans. Am. Fish. Soc. 106, 545–555 (1977).
Mittlebach, G. G. Foraging efficiency and body size: a study of optimal diet and habitat use by bullgills. Ecology 62, 1370–1386 (1981).
Dillon, P. J. & Rigler, F. H. The phosphorous-chlorophyll relationship in lakes. Limnol. Oceanogr. 19, 767–773 (1973).
Guildford, S. J. & Hecky, R. E. Total nitrogen, total phosphorus, and nutrient limitation in lakes and oceans: Is there a common relationship? Limnol. Oceanogr. 45, 1213–1223 (2000).
Hakanson, L. & Eklund, J. M. Relationships between chlorophyll, salinity, phosphorus, and nitrogen in lakes and marine areas. J. Coast. Res. 26, 412–423 (2010).
Brezonik, P. L. Effect of organic color and turbidity of Secchi disc transparency. J. Fish. Res. Board. Can. 35, 1410–1416 (1978).
Jr, D. E. C. & Bachmann, R. W. Prediction of total phosphorous concentrations, chlorophyll a, and secchi depth in natural and artificial lakes. Can. J. Fish. Aquat. Sci. 38, 414–423 (1981).
Zambrano, L., Scheffer, M. & Martinez-Ramos, M. Catastrophic response of lakes to benthivorous fish introduction. Oikos 94, 344–350 (2001).
Lin, M. et al. Maintaining economic value of ecosystem services whilst reducing environmental cost: a way to achieve freshwater restoration in China. Plos One 10, e0120298 (2015).
Jin, G. Studies status of lake eutrophication in China. Mod. Agric. Technol. 16, 334–336 (2008).
Moss, B., Madgwick, J. & Phillips, G. A Guide to the Restoration of Nutrient-Enriched Shallow Lakes. Broads Authority and Environment Agency, Norwich, UK (1996).
Havens, K. E. et al. Nutrient dynamics and the eutrophication of shallow lakes Kasumigaura (Japan), Donhu (PR China), and Okeechobee (USA). Environ. Pollut. 111, 263–272 (2001).
Xie, S. et al. The Spatial Pattern of the Small Fish Community in the Biandantang Lake – A Small Shallow Lake Along the Middle Reach of the Yangtze River, China. Environ. Biol. Fishes 57, 179–190 (2000).
APHA—American Public Health Association. Standard methods for the examination for water and wastewater, 19th ed. Washington, D.C (1995).
Holm-Hansen, O. & Rieman, B. Chlorophyll-a determination: improvement in methodology. Oikos 30, 438–447 (1978).
Acknowledgements
This study was financially supported by the R and D Project of Ministry of Science and Technology of China (Grant No. 2015BAD13B02), the Projects of the National Natural Science Foundation of China (Grant No. 31201994 and 30830025), and the Special Fund for Agro-scientific Research in the Public Interest (Grant No. 201303056), and China Agriculture Research System (CARS-45-23). Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Author information
Authors and Affiliations
Contributions
W.L., T.L.Z., J.S.L. and Z.J.L. conceived and designed the study. W.L., M.L.L. and C.B.G. performed the study. W.L., B.J.H. and D.A.B. conducted the statistical analysis. W.L. wrote the first draft. All authors contributed substantial revisions and comments during manuscript preparation.
Corresponding author
Ethics declarations
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Li, W., Hicks, B.J., Lin, M. et al. Impacts of hatchery-reared mandarin fish Siniperca chuatsi stocking on wild fish community and water quality in a shallow Yangtze lake. Sci Rep 8, 11481 (2018). https://doi.org/10.1038/s41598-018-29758-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-018-29758-z
- Springer Nature Limited
This article is cited by
-
Characterizing the development of Sea ranching in China
Reviews in Fish Biology and Fisheries (2022)