
Abstract
We used geographically weighted regression to investigate the relationship between biodiversity and the spatial stability of forest productivity (SSFP) in a subtropical mountain forest. We examined the effect of elevation on this relationship and on its spatial non-stationarity. We found that higher woody plant diversity reduced SSPF. Higher woody plant diversity strengthened the asynchrony of species responses to spatial heterogeneity of forest habitats, which contributed to SSFP, but reduced two factors that enhanced SSFP: species dominance and the spatial stability of the dominant species. The percentage of variation in SSFP explained by diversity measures was highest for the Shannon-Wiener index, lowest for functional dispersion, and intermediate for species richness. The correlations of woody plant diversity with SSFP became stronger with elevation and varied among plots, indicating that the spatial non-stationarity existed in the biodiversity-SSFP relationship. These correlations became weaker in most cases after controlling for elevation. Our results suggest that in the subtropical mountain forest higher woody plant diversity has a spatially destabilising effect on forest productivity, particularly at higher elevations.
Similar content being viewed by others
Explore related subjects
Find the latest articles, discoveries, and news in related topics.Introduction
The accelerating loss of biodiversity and the degradation of ecosystem functions represent increasing threats to humans. Understanding the biodiversity-ecosystem functioning relationship is thus crucial for maintaining the delivery of ecosystem services on which humans depend1,2,3,4. As one of the most fundamental ecosystem functions5, stability refers to the resistance to change or disturbance to ecosystem structure and function across space and time6,7,8,9. Ecosystems with high stability can maintain a dynamic equilibrium of structure and consequently can guarantee the delivery of functions or services when faced with disturbances, especially when faced with environmental deterioration caused by anthropogenic activities10, 11. As an important aspect of ecosystem stability, temporal stability of ecosystem productivity (TSEP) is defined as the ratio of the mean value to the standard deviation of productivity across time and is strongly influenced by biodiversity12, 13. Mounting evidence has shown that greater biodiversity enhances the TSEP14, 15. Analogous to TSEP, spatial stability of ecosystem productivity is another important aspect of ecosystem stability, which reflects the response of ecosystem productivity to spatially environmental heterogeneity7, 10. However, whether biodiversity also enhances the spatial stability of ecosystem productivity remains unclear, particularly in forest ecosystems7, 10. Forests are major terrestrial ecosystems, and the stability of forest productivity plays an essential role in maintaining global biodiversity, as well as in stabilising the productivity of the global terrestrial ecosystems11. If biodiversity is positively correlated with the spatial stability of forest productivity (SSFP), then humans’ effort to maintain the terrestrial biodiversity7 will simultaneously contribute to SSFP.
Some ecologists believe that the effects of biodiversity on SSFP should be similar to its effects on TSEP7, 10. To explain the stabilising effect of biodiversity on TSEP, ecologists have proposed a variety of theories, including the overyielding effect, the portfolio effect, and species asynchrony15,16,17. Species asynchrony is regarded as one of the most important driving forces of TSEP. Plant species show different preferences for environmental conditions because of niche partitioning18. When a species-rich community experiences a disturbance, species that maximize fitness to the changed habitat will benefit, while others that are not adaptable to the new environment will be inhibited. Therefore, the overall productivity of a species-rich community will respond more stably to fluctuating environmental conditions than a species-poor community12, 15, 17, 19. Biodiversity may also affect TSEP through species dominance and the stability of the dominant species. A higher relative abundance of the most abundant species decreases the standard deviation relative to the mean value of productivity. Thus, species dominance is positively correlated with community stability5. A dominant species contributes most to community productivity. The stability of the dominant species plays an essential role in community stability20. Previous studies have demonstrated that higher biodiversity may lead to lower population stability16 and lower stability of the dominant species may destabilise community productivity20. However, we know little of how species asynchrony, species dominance, and the stability of the dominant species affect the relationship between biodiversity and SSFP7, 10.
Although most studies dealing with the effect of biodiversity on the stability of ecosystem productivity have used species richness as a diversity measure, species richness alone fails to account for the special roles of common and rare species in the community, the evenness of species distribution, species coexistence, and species-habitat associations13, 21, 22. In contrast, the Shannon-Wiener index involves both abundance and richness as predictors to quantify community heterogeneity23, 24. Similarly, the functional dispersion index considers not only richness but also niche complementarity of coexisting species and the functional traits of individual plant species25, 26. These diversity measures will presumably differ in their ability to explain the variations in the stability of ecosystems.
Both plant diversity and the stability in productivity are affected by temperature, moisture, and soil nutrients27, 28, which are in turn strongly influenced by elevation29. Soil nutrient content and availability change with temperature and precipitation30, 31. As elevation increases, temperature linearly decreases, whereas precipitation exhibits various patterns, such as linear, unimodal, or bimodal trends32, 33. Therefore, the biodiversity-stability relationship may also change with elevation-driven changes in environmental factors. Understanding the complex associations among plant diversity, SSFP, and the influence of elevation is crucial to the management and conservation of forests, especially mountain forests, and should also provide insight into the relationship between biodiversity and ecosystem functioning.
Here, we used geographically weighted regression (GWR) to investigate the relationship between biodiversity and SSFP, based on the data collected from 129 mountain forest plots established in the eastern part of Guangzhou. We used species richness, the Shannon-Wiener index, and functional dispersion as diversity measures, total tree basal area as a proxy variable for forest productivity, and the reciprocal of the coefficient of variation in the total tree basal areas among subplots within a plot as a proxy variable for SSFP. We also used Berger-Parker index to measure species dominance, and the reciprocal of the coefficient of variation in the tree basal areas of the dominant species among subplots within a plot as a proxy variable for spatial stability of the dominant species. We attempted to answer the following questions: (1) How are woody plant diversity, species asynchrony, the stability of the dominant species, and species dominance related to SSFP? (2) How do different diversity measures differ in explaining variations in SSFP? and (3) Does elevation affect the biodiversity-SSFP relationship? If so, how?
Materials and Methods
Study area
This study was conducted in the eastern part of Guangzhou (112°57′ - 114°3′ E, 22°26′ - 23°56′ N), which is located in the central south of Guangdong Province, China. Guangzhou has a subtropical monsoon climate regime and is consequently hot and wet in summer and cold and dry in winter, and is occasionally affected by typhoons and thunderstorms. The mean annual temperature and precipitation are 21.5 °C and 1667.5 mm, respectively. The wet season is from April to September. Most of the area has a south subtropical lateritic red soil whose parent rock is granite and sand shales34. The forests we investigated were evergreen broadleaved mountain forests with little disturbance. No signs of tree cutting and domestic animal grazing were observed in our monitoring plots.
Data collection
To investigate the relationship between biodiversity and spatial stability of productivity in natural forest ecosystems, we established plots at 43 sites in the subtropical forests in eastern part of Guangzhou. At each site, we established three rectangular plots that were separated by at least 100 m. Each plot had an area of 1200 m2 (30 m × 40 m or 20 m × 60 m), and the total area of the 129 plots (43 sites × 3 plots/site) was 15.48 ha. Woody plant species richness in these plots ranged from 4 to 49. Each plot was further divided into 12 subplots of 10 m × 10 m.
We recorded the latitude, longitude, slope aspect, slope steepness, and elevation of each plot. Plot elevation ranged from 25 to 819 m a.s.l. The plots were grouped into four elevation classes for further analysis: 1 = 25–200 m; 2 = 201–400 m; 3 = 401–600 m; and 4 = 601–819 m. A tree census was conducted in each plot. We measured all woody plants with a diameter at breast height (DBH) ≥ 3 cm and labeled them with unique numbers. We recorded the species name, DBH (measured to the nearest 0.1 cm), and tree height (measured to the nearest 0.1 m) for all stems measured. All stems were identified to species during the survey (i.e., on site) except for those with uncertain identity; in the latter case, voucher specimens were collected, labeled, and subsequently identified by the South China Agricultural University Herbarium (CANT). Plant nomenclature follows Ye & Peng35. Field work for data collection was conducted and completed in 2014.
Statistical analysis
We calculated species richness (S), the Shannon-Wiener index (H′), and functional dispersion (FDis) for plant diversity at the plot level using the following formulas:
where p i is the relative individual density of species i; a i is the individual density of species i, and z i is the distance of species i to the weighted centroid c, which is determined by the individual density and functional trait value of each species36. Functional dispersion is based on maximum tree height, which is calculated as the 99th percentile of the tree height values measured for each species.
Functional dispersion can be calculated based on any type and number of functional traits and any dissimilarity or distance measure, and is independent of the effect of species richness36,37,38. We used maximum tree height to calculate functional dispersion because height is associated with competition for light among trees and directly correlates with woody productivity39. In a few cases (18 in 25948 cases), we used multiple imputations to solve the problem of missing data in tree height values.
We used basal area as the proxy variable for forest productivity40, 41. SSFP was calculated as the ratio of the mean value to the standard deviation of productivity among subplots within each plot7, 10:
Similarly, stability of the dominant species, the species with maximum total basal area at the plot level, was calculated as the ratio of the mean value to the standard deviation of productivity of the dominant species among subplots within each plot. We used the same formula that is used to calculate asynchronous responses of species across time15, 28, 42 in order to calculate asynchronic responses of species across space, but the meanings of the parameters changed:
where σ i is the standard deviation of the productivity of species i among subplots within a plot, σ 2 is the variance of the productivity among subplots within a plot, and S is the species richness within a plot. Because some species within a plot occurred in only one or two subplots and because the calculation of standard deviation requires at least 3 replicates, we assumed that all species within a certain plot have individuals in each subplot of that plot. In cases where a species within a plot had no individual in any subplot of that plot, the productivity of that species in that subplot was recorded as zero. Asynchrony ranged from 0 to 1, where 0 indicated a perfectly synchronic response and 1 indicated a perfectly asynchronic response among different species to spatial heterogeneity in habitat. We calculated the Berger-Parker index to measure species dominance:
where D B-P is the Berger-Parker index, N max is the number of individuals of the most abundant species, and N is the total number of individuals within each plot. Before conducting further analyses, we tested the normality of variables, and those violating the normality assumption were log10-transformed.
Although mounting evidence has demonstrated positive correlations between biodiversity and stability, the correlations and significance has differed among studies12, 14, 15. In one study, the biodiversity-stability relationships were even inconsistent among different sites19. These findings indicate spatial non-stationarity in correlations between biodiversity and temporal stability. Because the plots in our study were located in different areas of the eastern part of Guangzhou, we used geographically weighted regression (GWR) modeling to assess the relationship between woody plant diversity and SSFP because GWR models can efficiently deal with spatial non-stationarity43, 44. Model performance was assessed using the corrected Akaike Information Criterion (AICc), residual sum of squares (RSS), and the coefficient of determination (R2). Lower AICc and RSS values and higher R2 values indicate better model performance44, 45.
The effect of elevation was assessed by using the Kruskal-Wallis test to examine the significance of variation, along an elevational gradient, in the correlations of woody diversity with SSFP and of species asynchrony, species dominance, and stability of the dominant species with SSFP or woody plant diversity. After controlling for elevation, we used GWR models to evaluate the relationship between woody plant diversity and SSFP; elevation was controlled for by using the residuals of SSFP on elevation45. Correlations of species asynchrony, species dominance, and stability of the dominant species with plant diversity or SSFP after controlling for elevation were examined using the same methods.
R software version 3.2.546 was used for all calculations and statistical analyses. Multiple imputations were conducted using the mice package47. The Shannon-Wiener index and functional dispersion were calculated using vegan 48 and FD 36 packages, respectively. Normality tests and Kruskal-Wallis tests were performed using the stats package46. GWR analyses were conducted with the spgwr package49, using a Gaussian spatial weighting function with an adaptive spatial kernel.
Results
Plant diversity-SSFP relationship
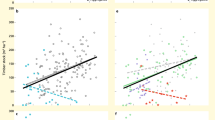
Although the three measures of woody plant diversity explained significant proportions of the variances of SSFP (overall R2 > 0.20, Table 1), the measures differed in explanatory power. The Shannon-Wiener index explained the most variance (overall R2 = 0.280), while functional dispersion explained the least (overall R2 = 0.219). According to the AICc and RSS values, the GWR model for SSFP against Shannon-Wiener index was fitted better than those against species richness or functional dispersion (Table 1). Spatial non-stationarity existed in the correlations between SSFP and plant diversity. SSFP was negatively correlated with plant diversity in most plots and was positively correlated with plant diversity in only a few plots. Local R2 of GWR models for SSFP against plant diversity varied strongly among plots (Fig. 1a–c). Correlations between SSFP and species richness or the Shannon-Wiener index were highly consistent in both magnitude and direction among plots (Fig. 1a,b). Correlations between SSFP and species diversity (species richness or Shannon-Wiener index) showed larger ranges of variations among plots than the correlations between SSFP and functional diversity (functional dispersion) (Fig. 1a–c).

Local R2 for geographically weighted regressions of SSFP against various predictive variables. The predictive variables: (a) species richness, (b) the Shannon-Wiener index, (c) functional dispersion, (d) species asynchrony, (e) species dominance, and (f) stability of the dominant species. Filled circles represent negative correlations, and triangles represent positive correlations.
Explanatory power of predictors for the plant diversity-SSFP relationship
GWR model for SSFP against stability of the dominant species was fitted better than those against species asynchrony or species dominance. As assessed by AICc and RSS values, GWR models for species asynchrony, species dominance, or stability of the dominant species against Shannon-Wiener index were fitted better than those against species richness or functional dispersion (Table 1). Overall, more of the variance in SSFP was explained by species asynchrony, species dominance, or stability of the dominant species than by plant diversity measures. Among species asynchrony, species dominance, and stability of the dominant species, stability of the dominant species explained the most variance (overall R2 = 0.408), and species dominance explained the least variance (overall R2 = 0.260) in SSFP. Plant diversity had a strong explanatory power for variances of species asynchrony, species dominance, and stability of the dominant species. Similarly, the Shannon-Wiener index explained the most variance and functional dispersion explained the least variance in species asynchrony, species dominance, and stability of the dominant species (Table 1). Spatial non-stationarity also existed in correlations of species asynchrony, species dominance, or stability of the dominant species with SSFP and with plant diversity measures. These correlations varied strongly among plots (Figs 1 and 2). SSFP was positively correlated with species asynchrony and species dominance in most plots and was negatively correlated with these variables in only a few plots (Fig. 1d,e). SSFP was positively correlated with stability of the dominant species in all plots (Fig. 1f). Species asynchrony was positively correlated with plant diversity in most plots and was negatively correlated with plant diversity in only a few plots (Fig. 2a–c). Both species dominance and stability of the dominant species were negatively correlated with plant diversity in most plots and were positively correlated with plant diversity in only a few plots (Fig. 2d–i). Species dominance was negatively correlated with species diversity in all plots. Stability of the dominant species was also negatively correlated with the Shannon-Wiener index in all plots (Fig. 2d,e,h).

Local R2 for geographically weighted regressions of species asynchrony, species dominance, and stability of the dominant species, respectively, against diversity measures. 1) species asynchrony against (a) species richness, (b) Shannon-Wiener index, and (c) functional dispersion; 2) species dominance against (d) species richness, (e) the Shannon-Wiener index, and (f) functional dispersion; 3) stability of the dominant species against (g) species richness, (h) the Shannon-Wiener index, and functional dispersion. Filled circles represent negative correlations, and triangles represent positive correlations.
Influence of elevation
Changes in elevation influenced the relationships of SSFP to plant diversity, species asynchrony, species dominance, and the stability of the dominant species. As elevation increased, the local R2 tended to increase significantly for regressions of SSFP against species diversity (P < 0.0001, Fig. 3a,b), species asynchrony (P = 0.0004, Fig. 3d), species dominance (P = 0.0044, Fig. 3e), and stability of the dominant species (P < 0.0001, Fig. 3f), but fluctuate significantly for the regression of SSFP against functional dispersion (P = 0.0072, Fig. 3c). The local R2 for regressions of species asynchrony, species dominance, and stability of the dominant species, respectively, against plant diversity varied, either significantly increased, significantly decreased, significantly fluctuated, or remained relatively constant with increasing elevation (see Supplementary Fig. S1).

Effects of elevation on the local R 2 for geographically weighted regression of SSFP against various predictive variables. The predictive variables: (a) species richness (KW-H(3, 129) = 39.789, P < 0.0001), (b) the Shannon-Wiener index (KW-H(3, 129) = 42.592, P < 0.0001), (c) functional dispersion (KW-H(3, 129) = 12.040, P = 0.0072), (d) species asynchrony (KW-H(3, 129) = 18.299, P = 0.0004), (e) species dominance (KW-H(3, 129) = 13.091, P = 0.0044), and (f) stability of the dominant species (KW-H(3, 129) = 29.666, P < 0.0001). Boxes represent the 25th and 75th percentile values, the horizontal line in the box is the median, the whiskers show the non-outlier range, and the solid points represent outliers. Differences along the elevation gradient were tested using Kruskal-Wallis H test. Elevation gradient: 1 = 0–200 m; 2 = 201–400 m; 3 = 401–600 m; 4 = 601–819 m.
After elevation was controlled for, the overall R2 decreased for all models, but the independent variables still explained more than 10% of the variances of the dependent variables (Table 1). Controlling for elevation did not change the direction of the correlations in most plots for any model, but decreased the magnitude of the variation in correlations, except for the correlations of SSFP with species asynchrony, for correlations of species dominance with Shannon-Wiener index or functional dispersion, or for the correlations of stability of the dominant species with functional dispersion; for these exceptions, controlling for elevation increased the magnitude of variations (Figs 1 and 2; Supplementary Figs S2 and S3).
Discussion
Species with different preferences for certain habitats respond differently to environmental changes across time or space. Some species are enhanced while others are inhibited or unaffected by changes in the environment, reflecting different adaptations to the habitat changes. In addition, the speed at which organisms respond to environmental change differs among species. Communities with higher diversity contain more species with contrasting preferences for environmental changes, and this diversity therefore tends to increase community stability15, 17. This view is to some extent supported by the results of our study. We found that the SSFP was positively associated with species asynchrony across space, which in turn was positively associated with woody plant diversity. However, SSFP was actually reduced by plant diversity in most plots. This can be explained by our finding that both species dominance and the stability of the dominant species were positively correlated with SSFP but negatively correlated with plant diversity. The effects of species dominance and spatial stability of the dominant species overrode that of species asynchrony. These results suggest that plant diversity has more destabilising than stabilising effects on forest productivity across space and that species dominance and the stability of the dominant species, rather than species asynchrony, determine the relationship of plant diversity to SSFP. Higher species dominance increased temporal stability by lowering the standard deviation relative to the mean value of forest productivity across time, indicating that species dominance was positively correlated with temporal stability5. Species dominance was also positively correlated with SSFP (Fig. 1e). As plant diversity increased, species dominance decreased, consequently leading to lower SSFP. A dominant species occupies larger ecological niches, is more adaptable to environmental changes50, and is more stable than other species20, 51. Furthermore, the most dominant species is the major contributor of total community productivity and thus contributes most to the stability of forest productivity. Changes in the stability of the dominant species will affect temporal stability of ecosystem productivity20. Higher plant diversity leads to lower population stability, particularly the stability of both the dominant and rare species, because of competition16, and decreases in the stability of the dominant species destabilise community productivity across time20. In our study, higher plant diversity also reduced SSFP by decreasing the spatial stability of the dominant species. We therefore regarded the decrease in species dominance and in the stability of the dominant species with the increase in plant diversity as key factors explaining the destabilising effect of diversity on SSFP. We also found that the Shannon-Wiener index explained more of the variance in the diversity-SSFP relationship than the other diversity measures used in this study. This might be because the Shannon-Wiener index combines the richness, abundance, and evenness of species, and thereby considers the special roles of the dominant, common, and rare species in the community24, and because the dominant species greatly affects community stability5, 52.
The responses of spatial stability of productivity to biodiversity can differ among ecosystems. In contrast to the results from our forest study, results of the Jena grassland study suggested that biodiversity is positively correlated with spatial stability of productivity and that only functional trait diversity can explain the variations in spatial stability10. Previous studies have demonstrated that woody plants are more sensitive to environmental change than grasses, which might explain the discrepancy in the relationship between biodiversity and spatial stability in forest versus grassland ecosystems53, 54.
We found that the correlation of woody plant diversity with SSFP and correlations of species asynchrony, species dominance, and stability of the dominant species with plant diversity and with SSFP were affected by elevation. As temperature linearly decreases and precipitation distribution patterns change with increasing elevation32, 33, soil moisture and nutrients also change30, 31. Changes in these environmental factors can drive variations in woody plant diversity, productivity, species asynchrony, species dominance, and the stability of the dominant species in a forest ecosystem, thus leading to variations in SSFP20, 27, 28, 42, 55. As elevation increased in the current study, the effects of species diversity on SSFP significantly increased and quickly saturated at higher elevations. These findings suggest that higher woody plant diversity, particularly species diversity, reduced the stability of forest productivity across space in higher-elevation regions and that the destabilising effect of plant diversity on forest productivity across space might be stronger at higher elevations. That species asynchrony, species dominance, and the stability of the dominant species had stronger associations with SSFP as elevation increased demonstrated that their stabilising effects on SSFP became stronger as elevation increased. The different rates at which these correlations (coefficients of determination) increased with elevation demonstrated that the relative importance of species asynchrony to species dominance and the stability of the dominant species fluctuated as elevation increased. Correlations between plant diversity and SSFP, and correlations of species asynchrony, species dominance, and the stability of the dominant species with SSFP and plant diversity decreased when elevation was controlled for, demonstrating that elevation had a strong effect on the SSFP, and that the effect of elevation might be even stronger than that of plant diversity.
Spatial non-stationarity was evident in the diversity-SSFP relationship and decreased in most cases when elevation was controlled for, indicating that elevation affects but does not determine the spatial non-stationarity in the correlations of diversity with SSFP. Environmental heterogeneity and the interactions between species and environment are regarded as two important drivers of spatial non-stationarity in the diversity-stability relationship15. The growth, development, and distribution of plants are associated with habitat heterogeneity54, 56, 57, which is controlled by elevation-driven variations in temperature, moisture, and soil nutrients30,31,32,33. Thus, both plant diversity and ecosystem stability, and consequently their correlations, are affected by elevation-driven changes in a habitat32, 42, 55. However, habitat variations are driven by many biotic and abiotic factors, including slope aspect, slope steepness, anthropogenic disturbance, and biological invasion58,59,60,61. Further studies are therefore needed to determine how diversity affects SSFP and how the relationship between diversity and SSFP is influenced by spatial non-stationarity at multiple spatial scales.
In conclusion, our results demonstrated that plant diversity increased species asynchrony, which in turn promoted SSFP; however, due to its inverse association with species dominance and the stability of the dominant species, which are the major drivers of SSFP, higher plant diversity reduced SSFP. The destabilising effect of plant diversity on SSFP became stronger with elevation, indicating that higher plant diversity further reduced the SSFP at higher elevations. The proportion of variation in SSFP explained by diversity measures was highest for the Shannon-Wiener index, lowest for functional dispersion, and intermediate for species richness. These results will help expand our understanding of the biodiversity-ecosystem functioning relationship and will have potential implications for biodiversity conservation and forest management.
Data Availability
The datasets generated during the current study are available from the corresponding author on reasonable request.
References
Cardinale, B. Impacts of biodiversity loss. Science. 336, 552–553 (2012).
Hooper, D. U. et al. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature. 486, 105–108 (2012).
Naeem, S., Duffy, J. E. & Zavaleta, E. The functions of biological diversity in an age of extinction. Science. 336, 1401–1406 (2012).
Reich, P. B. et al. Impacts of biodiversity loss escalate through time as redundancy fades. Science. 336, 589–592 (2012).
Sasaki, T. & Lauenroth, W. K. Dominant species, rather than diversity, regulates temporal stability of plant communities. Oecologia. 166, 761–768 (2011).
Loreau, M. et al. A new look at the relationship between diversity and stability. In: Loreau M, Naeem S, Inchausti P, eds. Biodiversity and ecosystem functioning 79–91 (Oxford University Press, Oxford, Oxfordshire, 2002).
Eisenhauer, N. et al. Impact of above- and below-ground invertebrates on temporal and spatial stability of grassland of different diversity. J. Ecol. 99, 572–582 (2011).
MacDonald, R. L., Chen, H. Y. H., Bartels, S. F., Palik, B. J. & Prepas, E. E. Compositional stability of boreal understorey vegetation after overstorey harvesting across a riparian ecotone. J. Veg Sci. 26, 733–741 (2015).
Saavedra, S., Rohr, R. P., Fortuna, M. A., Selva, N. & Bascompte, J. Seasonal species interactions minimize the impact of species turnover on the likelihood of community persistence. Ecology. 97, 865–873 (2016).
Weigelt, A., Schumacher, J., Roscher, C. & Schmid, B. Does biodiversity increase spatial stability in plant community biomass? Ecol Lett. 11, 338–347 (2008).
Allesina, S. & Tang, S. Stability criteria for complex ecosystems. Nature. 483, 205–208 (2012).
Morin, X., Fahse, L., de Mazancourt, C., Scherer-Lorenzen, M. & Bugmann, H. Temporal stability in forest productivity increases with tree diversity due to asynchrony in species dynamics. Ecol Lett. 17, 1526–1535 (2014).
Hautier, Y. et al. Anthropogenic environmental changes affect ecosystem stability via biodiversity. Science. 348, 336–340 (2015).
Cardinale, B. J. et al. Biodiversity simultaneously enhances the production and stability of community biomass, but the effects are independent. Ecology. 94, 1697–1707 (2013).
Jucker, T., Bouriaud, O., Avacaritei, D. & Coomes, D. A. Stabilizing effects of diversity on aboveground wood production in forest ecosystems: Linking patterns and processes. Ecol Lett. 17, 1560–1569 (2014).
Lehman, C. L. & Tilman, D. Biodiversity, stability, and productivity in competitive communities. Am Nat. 156, 534–552 (2000).
Loreau, M. & de Mazancourt, C. Biodiversity and ecosystem stability: A synthesis of underlying mechanisms. Ecol Lett. 161, 106–115 (2013).
Loreau, M. & de Mazancourt, C. Species synchrony and its drivers: Neutral and nonneutral community dynamics in fluctuating environments. Am Nat. 172, E48–E66 (2008).
Hector, A. et al. General stabilizing effects of plant diversity on grassland productivity through population asynchrony and overyielding. Ecology. 91, 2213–2220 (2010).
Grman, E., Lau, J. A. Jr., Schoolmaster, D. R. & Gross, K. L. Mechanisms contributing to stability in ecosystem function depend on the environmental context. Ecol Lett. 13, 1400–1410 (2010).
Gross, K. et al. Species richness and the temporal stability of biomass production: A new analysis of recent biodiversity experiments. Am Nat. 183, 1–12 (2014).
Isbell, F. et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature. 526, 574–577 (2015).
Jabot, F. & Chave, J. Inferring the parameters of the neutral theory of biodiversity using phylogenetic information and implications for tropical forests. Ecol Lett. 12, 239–248 (2009).
Bernhardt-Roemermann, M. et al. Drivers of temporal changes in temperate forest plant diversity vary across spatial scales. Global Change Biol. 21, 3726–3737 (2015).
Ruiz-Benito, P. et al. Diversity increases carbon storage and tree productivity in Spanish forests. Global Ecol Biogeogr. 23, 311–322 (2014).
Zuo, X. et al. Plant functional diversity mediates the effects of vegetation and soil properties on community-level plant nitrogen use in the restoration of semiarid sandy grassland. Ecol Indic. 64, 272–280 (2016).
Zhou, G. et al. Substantial reorganization of China’s tropical and subtropical forests: Based on the permanent plots. Global Change Biol. 20, 240–250 (2014).
Zhang, Y. et al. Nitrogen enrichment weakens ecosystem stability through decreased species asynchrony and population stability in a temperate grassland. Global Change Biol. 22, 1445–1455 (2016).
Looby, C. I., Maltz, M. R. & Treseder, K. K. Belowground responses to elevation in a changing cloud forest. Ecol Evol. 6, 1996–2009 (2016).
Nottingham, A. T. et al. Climate warming and soil carbon in tropical forests: Insights from an elevation gradient in the Peruvian Andes. Bioscience. 65, 906–921 (2015).
Duran, J. et al. Climate change decreases nitrogen pools and mineralization rates in northern hardwood forests. Ecosphere. 7, e12513 (2016).
McCain, C. M. & Grytnes, J. Elevational gradients in species richness. In: Jonsson, R. ed. Encyclopedia of Life Sciences (eLS) 1–10 (John Wiley & Sons, Ltd, Chichester, 2010).
Acharya, K. P., Vetaas, O. R. & Birks, H. J. B. Orchid species richness along Himalayan elevational gradients. J. Biogeogr. 38, 1821–1833 (2011).
Wang, J., Huang, L., Ren, H., Sun, Z. & Guo, Q. Regenerative potential and functional composition of soil seed banks in remnant evergreen broad-leaved forests under urbanization in south China. Community Ecol. 16, 86–94 (2015).
Ye, H. & Peng, S. Plant diversity inventory of Guangdong. (World Publishing Corporation, Guangzhou, 2006).
Laliberte, E. & Legendre, P. A distance-based framework for measuring functional diversity from multiple traits. Ecology. 91, 299–305 (2010).
Anderson, M. J. Distance-based tests for homogeneity of multivariate dispersions. Biometrics. 62, 245–253 (2006).
Laliberte, E., Legendre, P. & Shipley, B. FD: Measuring functional diversity from multiple traits, and other tools for functional ecology. R package version 1.0–12. (2014).
Kunstler, G. et al. Plant functional traits have globally consistent effects on competition. Nature. 529, 204–207 (2016).
DeClerck, F. A. J., Barbour, M. G. & Sawyer, J. O. Species richness and stand stability in conifer forests of the Sierra Nevada. Ecology. 87, 2787–2799 (2006).
Huong, N., Herbohn, J., Firn, J. & Lamb, D. Biodiversity-productivity relationships in small-scale mixed-species plantations using native species in Leyte province, Philippines. Forest Ecol Manag. 274, 81–90 (2012).
Xu, Z. et al. Environmental changes drive the temporal stability of semi-arid natural grasslands through altering species asynchrony. J. Ecol. 103, 1308–1316 (2015).
Wang, Q., Ni, J. & Tenhunen, J. Application of a geographically-weighted regression analysis to estimate net primary production of Chinese forest ecosystems. Global Ecol Biogeogr. 14, 379–393 (2005).
Xu, X., Wang, Z., Rahbek, C., Lessard, J. & Fang, J. Evolutionary history influences the effects of water-energy dynamics on oak diversity in Asia. J. Biogeogr. 40, 2146–2155 (2013).
Roll, U., Geffen, E. & Yom-Tov, Y. Linking vertebrate species richness to tree canopy height on a global scale. Global Ecol Biogeogr. 24, 814–825 (2015).
R Core Team. R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/ (2016).
Buuren, S. V. & Groothuis-Oudshoorn, K. Mice: Multivariate Imputation by Chained Equations in R. J. Stat Softw. 45, 1–67 (2011).
Oksanen, J. et al. Vegan: Community Ecology Package. R Package Version 2.3–5. https://CRAN.R-project.org/package=vegan (2016).
Bivand, R. & Yu, D. Spgwr: Geographically Weighted Regression. R package version 0.6–28. https://CRAN.R-project.org/package=spgwr (2015).
Tanentzap, A. J. et al. When do plant radiations influence community assembly? The importance of historical contingency in the race for niche space. New Phytol. 207, 468–479 (2015).
Roscher, C. et al. Identifying population- and community-level mechanisms of diversity-stability relationships in experimental grasslands. J. Ecol. 99, 1460–1469 (2011).
Hillebrand, H., Bennett, D. M. & Cadotte, M. W. Consequences of dominance: A review of evenness effects on local and regional ecosystem processes. Ecology. 89, 1510–1520 (2008).
Hu, L., Li, M. & Li, Z. Geographical and environmental gradients of lianas and vines in China. Global Ecol Biogeogr. 19, 554–561 (2010).
Fang, J. et al. Forest community survey and the structural characteristics of forests in China. Ecography. 35, 1059–1071 (2012).
Hautier, Y. et al. Eutrophication weakens stabilizing effects of diversity in natural grasslands. Nature. 508, 521–525 (2014).
Lippok, D. et al. Topography and edge effects are more important than elevation as drivers of vegetation patterns in a neotropical montane forest. J. Veg Sci. 25, 724–733 (2014).
Stein, A., Gerstner, K. & Kreft, H. Environmental heterogeneity as a universal driver of species richness across taxa, biomes and spatial scales. Ecol Lett. 17, 866–880 (2014).
Vila, M. et al. Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems. Ecol Lett. 14, 702–708 (2011).
Shooner, S., Chisholm, C. & Davies, T. J. The phylogenetics of succession can guide restoration: An example from abandoned mine sites in the subarctic. J. Appl Ecol. 52, 1509–1517 (2015).
Martinez-Ramos, M., Ortiz-Rodriguez, I. A., Pinero, D., Dirzo, R. & Sarukhan, J. Anthropogenic disturbances jeopardize biodiversity conservation within tropical rainforest reserves. P. Natl Acad Sci USA. 113, 5323–5328 (2016).
Alexander, C., Deak, B. & Heilmeier, H. Micro-topography driven vegetation patterns in open mosaic landscapes. Ecol Indic. 60, 906–920 (2016).
Acknowledgements
We thank Dr. Zhenkui Li and Mr. Wenbin Li for plant identification. We acknowledge the help from Jiayu Cui, Keke Wang, Ying Liu, and Qiming Tang for their participation in field work. This research is funded in part by the Science and Technology Plan Programs of Guangdong Province (Grant No. 2013B020305008) and a grant from the Administration of Forestry and Gardening of Guangzhou Municipality (Grant No. GZFC-KT1-ZT2).
Author information
Authors and Affiliations
Contributions
Y.L.Z. and Z.S. conceived the study and wrote the first draft. Y.L.Z., Y.S., M.X., Y.Z., and Y.W. participated in field surveys and collected the data. Y.L.Z. performed all analyses and prepared the figures. Z.S. revised the draft and completed the final manuscript. All the authors contributed substantially to the discussion and review of the manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors declare that they have no competing interests.
Additional information
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Zhong, Y., Sun, Y., Xu, M. et al. Spatially destabilising effect of woody plant diversity on forest productivity in a subtropical mountain forest. Sci Rep 7, 9551 (2017). https://doi.org/10.1038/s41598-017-09922-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-017-09922-7
- Springer Nature Limited