
Abstract
During Arabidopsis seed coat development, copious amounts of mucilage polysaccharides are produced in the epidermal cells. When hydrated on imbibition, these polysaccharides expand and are released to encapsulate the seed as a two-layered hydrogel. Polysaccharides are synthesized from UDP-sugars by glycosyltransferases (GTs) and several GTs, with differing activities, have been identified that contribute to mucilage polysaccharide synthesis. How these GTs orchestrate production of the complex polysaccharides found in mucilage remains to be determined. In this study, we generated a range of multiple GT mutants using either CRISPR/Cas9 targeted mutation or genetic crosses of existing T-DNA insertion mutants. Four traits for mucilage amounts or macromolecular properties were examined for four replicate seed lots from 31 different GT mutant combinations. This data provides a valuable resource for future genetic, biochemical, structural, and functional studies of the roles and properties of polysaccharides present in Arabidopsis mucilage and the relative contributions of different GTs to mucilage production.
Similar content being viewed by others
Background & Summary
Mucilage is formed of polysaccharides which are produced in copious amounts in seed coat epidermal cells during seed development. The polysaccharides are hydrated and expand upon seed imbibition, leading to their release from the epidermal cells and the encapsulation of the seeds as a hydrogel1,2. Arabidopsis mucilage is structured in two layers: an outer diffuse layer which can be removed easily by shaking with water and an inner adherent layer which remains attached to the seed coat3. Both layers are primarily composed of rhamnogalacturonan-I (RGI), a pectic domain. The inner layer also contains small amounts of another pectic domain, homogalacturonan (HG), together with cellulose and some hemicelluloses1,3,4. As Arabidopsis mucilage shares common polysaccharides with cell walls in a more accessible format, it can be used as a simple proxy to investigate polysaccharide synthesis and physico-chemical properties.
Polysaccharides are synthesized from UDP- and GDP-sugars by a range of glycosyltransferases (GTs) having different activities, some of which have been implicated in the production of mucilage pectin based on mutant phenotypes. RGI: RHAMNOSYLTRANSFERASE1 (RRT1) was the first rhamnosyltransferase demonstrated to transfer Rha from UDP-Rha onto RGI primers5. While RRT1 is highly expressed in the seed coat at the developmental stage when mucilage polysaccharides are being produced, rrt1 mutant seeds have only slightly reduced mucilage RGI amounts. Expression analysis showed that two other GTs, MUCILAGE-RELATED70 (MUCI70) and GALACTURONOSYLTRANSFERASE-LIKE5 (GATL5), are also highly expressed in the seed coat6,7. In addition to dramatically reduced mucilage polysaccharide amounts, mucilage from muci70 mutant seeds is composed of shorter RGI polymer chains8. A recent study demonstrated the in vitro galacturonosyltransferase activity of MUCI70 towards RGI, which was subsequently named RHAMNOGALACTURONAN GALACTURONOSYLTRANSFERASE1 (RGGAT1)9. In contrast, seed mucilage RGI polymers in gatl5 are larger than those of wild type (WT), even though the amount of mucilage polysaccharides is still reduced7. While GALACTURONOSYLTRANSFERASE11 (GAUT11) was shown to catalyse the addition of UDP-galacturonic acid (GalA) to HG primers, gaut11 mutants surprisingly exhibited reduced amounts of RGI in mucilage.
Other GT mutants show an altered distribution of mucilage polysaccharides between the outer and the inner layers of mucilage, with no effect on global amounts. This is the case for mutants of MUCILAGE MODIFIED 5 (MUM5)/MUCI21 and IRREGULAR XYLEM 14 (IRX14), which have more outer mucilage layer polysaccharides and a corresponding reduction in the inner mucilage layer10,11. MUM5 and IRX14 have been proposed to be xylosyltransferases that synthesize xylan, a hemicellulose, which plays a pivotal role in the formation of the inner layer by anchoring RGI to cellulose attached to the seed surface11. The six GT mutants cited above (rrt1, muci70, gatl5, gaut11, irx14, mum5) have each been shown individually to impact mucilage polysaccharide characteristics. Nevertheless, it is currently difficult to appraise how these GTs collectively contribute to RGI synthesis and the nature of the RGI-xylan relationship, due to the lack of integrated studies.
Here, we have generated double and triple mutant combinations of the six GTs indicated above by a CRISPR/Cas9 targeted gene-editing approach to create novel, knock-out mutant alleles and by genetic crosses between existing T-DNA insertion mutant alleles. Mucilage from seeds of these mutants was then analysed to create a novel data set cataloguing the amounts and macromolecular properties of the different genotypes. These data will be a rich resource for determining how different GT activities impact RGI production and modulate polymer macromolecular properties, as well as for the choice of genotypes in studies examining the role of mucilage using seeds with different amounts and properties.
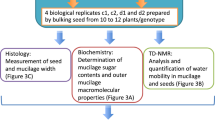
In total, data for 31 different GT mutants and their corresponding WT (Col-0) were obtained; 11 single, 17 double and 3 triple mutants (Table 1). These were used to generate data records which correspond to mucilage amounts quantified for the outer and inner layers using a medium-throughput method, and values for macromolecular characteristics of polymers present in the outer mucilage layer. The experimental procedures and different steps in data production are summarised in Fig. 1.

Schematic representation of the experimental workflow used to generate data records for glycosyltransferase mutant mucilage traits. Mutants were generated by either a CRISPR/cas9 based-approach or crosses between previously characterised T-DNA mutants. Seeds had been produced at two different times corresponding to cultivation series e or f, and up to four replicates were used for biochemical analyses. Analyses of mucilage GalA contents and macromolecular characteristics for the 32 genotypes are available in two data records. cfg: centrifugation; RGase: rhamnogalacturonan hydrolase.
Methods
Plant materials and growth conditions
The Arabidopsis thaliana T-DNA insertion mutants rrt1-1, gaut11-4, muci70-1, gatl5-1, irx14-2 and mum5-3 were previously described5,6,7,11 (Table 2); all mutants are in the Col-0 accession background. The genotypes of homozygous lines, and homozygous double mutants derived from crosses between two T-DNA lines, were confirmed by PCR using the primers listed in Table 2 and Supplemental Table 1. Other mutant alleles and mutant combinations were generated by CRISPR/Cas9 targeted gene editing as described below. Surface-sterilized seeds were sown onto Arabidopsis Gamborg B5 growth medium (Duchefa; https://www.duchefa-biochemie.com/ pH 6 supplemented with 30mM Sucrose, 0.08% (w/v) bromocresol purple and 0.7% (w/v) agar. Following stratification for three days (4 °C in the dark), seeds were germinated in a growth chamber for 14 days (16 h photoperiod, 50 µmol m−2 s−1 light intensity, 60% relative humidity, 18 °C). Seedlings were transplanted to soil (Tref Substrates; https://jiffygroup.com) and transferred to the glasshouse (18 °C–28 °C, minimum photoperiod of 13 h provided by supplementary lighting) and watered for 3 weeks with tap water, then with Plan-Prod nutritive solution (Fertil; https://www.fertil.fr/). The plants were grown at two different times, series ‘e’ was grown from February 2022 to April 2022 and series ‘f’ from March 2022 to May 2022. Seeds for each genotype were harvested as bulks from eight to twelve plants from a given series (e or f) so that each series represents a biological replicate. Subsequent analyses used up to four replicates (e.g. e_1, e_2, f_1 and f_2, where numbers indicate the technical replicates within a series) (Fig. 2).

Nomenclature for seed samples analyzed to generate dataset. This is based on, and compatible with, that used in Poulain et al.20.
Generation of GT mutants using a CRISPR/Cas9 targeted gene editing strategy
Two different guide RNAs were designed for each of the six target GTs (twelve guide RNAs in total), using the CRISPOR Web site (http://crispor.tefor.net), and selecting from proposed guide sequences based on their predicted efficiency and off-target probability. The specificity of the guides for their corresponding genes was checked by a BLAST search (https://blast.ncbi.nlm.nih.gov/Blast.cgi) against the Arabidopsis genome. Guide sequences were also selected for their position close to the 5′ end of each gene targeted, in order to increase the likelihood that induced mutations would produce truncated, inactive proteins. The target sequences of the twelve guide RNAs are listed in Table 3. Forward and reverse primers for each guide RNA were synthesized with additional sequences (Eurofins Genomics, https://eurofinsgenomics.eu/), that are complementary to the cohesive termini generated by BsaI digestion of either the pUPD:pU6-26 vector containing the Arabidopsis U6 promoter or the pUPD:psgRNA vector containing a scaffold guideRNA. Both plasmids enabling the guide to be cloned directionally into the domestication vector pDGB3_α1 or pDGB3_α2, to generate a transcription unit (TU) through a GoldenBraid (GB) cyclic digestion/ligation level 0 reaction as detailed by Vazquez-Vilar et al.12. These 12 guide RNA TUs were then stacked into a single vector through a 5 level pairwise cycle that alternated omega and alpha pDGB3 vectors between levels. The transcription units proRPS5A:hCas9:ter, composed of the Ribosomal Protein 5A promoter (pRPS5A) with the hCAS9 CDS (GB0575) and the RbcsE9 terminator, and proCMV:DsRED:terNos (gifts of Lionel Gissot), with the final 12 guide RNA TUs-proRPS5A:hCas9:ter-proCMV:DsRED:terNos in pDGB3_α2. The correct assembly was confirmed by PCR and sequencing at each step in E. coli (DH10B) or A. tumefaciens (C58C1) using the primers listed in Supplemental Table S1. Arabidopsis (Col-0 accession) was transformed with the final binary vector by the agrobacterium-mediated floral dip method13. Transgenic seeds were identified by DsRED fluorescence and DNA extracts from resulting plants subsequently genotyped for edited genes by sequencing PCR products amplified with the GT-CF and GT-CR primers (e.g. RRT1-CF and RRT1-CR) listed in Table 2 and Supplemental Table S1. To fix the mutation, Cas9-containing constructs were removed from segregating lines by selecting for seeds that did not exhibit DsRED fluorescence in the progeny of selected edited lines14. A further round of genome sequence analysis was carried out to confirm the edited mutations indicated in Fig. 3 and Table 2.

Position of mutations in the glycosyltransferase mutants used in this study and the sequence of guide RNAs used for gene-editing. Graphic representation of six glycosyltransferase genes (RRT1, GATL5, GAUT11, MUCI70, IRX14 and MUM5) with black boxes representing exons and red symbols and text the site and type of mutation in the corresponding mutants. The positions of the gene-specific primers used for genotyping gene-edited mutants are indicated with black arrows. For the precise position of mutations see Table 2. Guide RNAs, gRNA; bp, base pair.
Mucilage Extraction and biochemical analyses
A sequential extraction of the outer and the inner mucilage layers was performed essentially as previously described8,15, but adapted to a much smaller quantity of seeds. Twenty mg of intact seeds were mixed head-over-tail in 1 mL of distilled water for 3 h at room temperature. The suspension was centrifuged (17,500 g, 10 min) and exactly 760 µl of supernatant was recovered and filtered through a disposable glass microfiber filter (13 mm diameter, 0.45 µm pore size; A.I.T. France FS13PVDF45) for analysis as an outer mucilage extract. Seeds were resuspended by adding exactly 800 µl of 50 mM sodium acetate buffer pH 4.5. Then, 10.5 μL of rhamnogalacturonan hydrolase (EC 3.2.1.171, glycoside hydrolase family 28) provided by Novozymes, at 0.1 mg/mL in 50 mM sodium acetate buffer pH 4.5, were added. The seed suspension was then incubated for 16 h at 40 °C. Samples were centrifuged (17,500 g, 10 min), and the supernatants were collected for analysis as inner mucilage extracts.
The GalA content of the outer and inner mucilage extracts was determined colorimetrically by the automated m-hydroxybiphenyl method16. This quantification method is based on the ability of sugars to be converted into furfuric derivatives in the presence of hot sulfuric acid. Furfuric derivatives can then condense with various phenolic compounds to produce a coloured complex that can be quantified using a spectrometer. Acidic sugars can be quantified specifically using meta-hydroxy biphenyl (mphenyl-phenol or 3 phenyl-phenol; 530 nm)17. GalA solutions at 20, 40, 60, 80, and 100 µg/mL were used to generate a standard curve. To take residual outer mucilage extract present in inner mucilage extract into account, GalA amounts for the inner mucilage layer were calculated by subtraction of the GalA contents corresponding to this residual outer mucilage, as detailed in the following equation:
where GR is the GalA contents remaining in the inner mucilage extraction, GO presents the GalA contents in outer mucilage extracts, GI presents the actual inner mucilage contents in the inner mucilage extraction, and GIE presents the GalA contents in inner mucilage extracts. The 240 µL corresponds to the volume of water added to the seed (1000 µL) after deduction of the volume recovered as outer mucilage extract (760 µL). The 1050.5 µL corresponds to 240 µL plus the volumes of buffer (800 µL) and enzyme (10.5 µL) added for adherent mucilage hydrolysis. The GalA contents for outer and inner mucilage layers were then expressed with regard to seed mass.
The outer mucilage extracts were also analyzed for their weight-average molar mass (Mw) and intrinsic viscosity (IV) on a high-performance size exclusion chromatography (HPSEC) system (OMNISEC RESOLVE-REVEAL - Malvern Panalytical). The filtered (0.45 µm PVDF filter) outer mucilage extracts were boiled for 5 min, then samples were injected (50 µL) onto the HPSEC system maintained at 30 °C comprising a Shodex OH SB-G precolumn and a Shodex OH-Pack SB-805 HQ column eluted with 50 mM sodium nitrate at a flow rate of 0.7 mL min−1. Measurements were performed using a differential refractometer (OMNISEC REVEAL), a multi angle laser light scattering detector (λ = 660 nm, 44°, 60°, 76°, 90°, 108°, 124°, 140°, VISCOTEK SEC-MALS 9) and a differential pressure viscometer (OMNISEC REVEAL). Detectors were calibrated with a pullulan standard having narrow molecular mass distribution (weight-average molar mass = 40.611 Da, number-average molar mass = 38.931 Da, IV = 23.6 mL/g at 30 °C in 0.1 M sodium nitrate, refractive index increment [dn/dc] = 0.147 mL/g). Data analyses were carried out using OmniSec version 11.32 software (Malvern Panalytical) and a dn/dc value of 0.147 mL/g was used for mucilage extracts.
Data Records
The data record contains values for GalA content (mg/g seeds) in both the outer and inner layers of mucilage, as well as the weight-average molar mass (Mw) (g/mol) and intrinsic viscosity (IV) (dL/g) of the outer mucilage layer. The data were obtained from four replicates as described above, excepting three genotypes for Mw and IV and one genotype for GalA (genotypes coded 4004, 4012, 4030 and 4005, respectively, see Table 1), which had three replicates each. In addition, the four replicates analysed for the genotype coded 4010 were all from culture series f (f_1, f_2, f_3, and f_4), and for the genotype coded 4028 exclusively from culture series e (e_1, e_2, e_3, and e_4). All other genotypes were evaluated using two replicates from each culture series (e_1, e_2, f_1, and f_2) (Fig. 2). Note that certain values for Mw and IV were below the detection limit (B.D.L.) (genotypes coded 4007, 4008, 4018, 4019, and 4022).
The extent of variability in inner and outer mucilage GalA amounts observed between different glycosyltransferase mutants, compared to wild-type, is shown in Fig. 4 for one replicate from each of the 31 different mutant genotypes. This large diversity demonstrates the potential of these genotypes and the dataset for exploitation in future studies. The dataset is available at Data INRAE18. The sample nomenclature for input is outlined in Fig. 2, genotype codes in Table 1 and the description of variables in Table 4. An overview of the dataset is shown in Table 5 with the following seven columns:
-
1.
sample_code: the sample code (see Fig. 2)
-
2.
genotype_code: the genotype code (see Table 1)
-
3.
cultivation_series: e or f
-
4.
technical_replicate: the technical replicate (1, 2, 3, 4)
-
5.
sample_type: the sample type analyzed (0 or 1) (see Fig. 2)
-
6.
variable: the code of the variable (see Table 4 for the description)
-
7.
value: the measured value

Glycosyltransferase mutants exhibit a range of GalA amounts in (a) outer and (b) inner mucilage layers. Values represent one replicate from each of the 32 different genotypes. Red bar indicates the value for wild type (WT).
Technical Validation
The technical quality of the dataset was validated through the use of four replicates from two different culture series, series e grown from February 2022 to April 2022 and series f grown from March 2022 to May 2022, except for four genotype/variable combinations, as indicated above. The reproducibility of results was examined for biochemical analyses based on the variation between four replicates. The variation is presented as standard errors expressed as a % of the average value of the four replicates of WT, and the highest variation observed was less than 10% (Table 6). Furthermore, values obtained for WT mucilage were similar to those previously published from independent studies, validating the reproducibility of measurements (Fig. 5)3,11,15,19,20,21,22,23. This technical validation also confirms that the calculation used to compensate for the absence of a wash step in the medium-throughput method used here produced robust inner mucilage GalA values.

Technical validation of values obtained by comparison of values for wild-type extracts in this study to previously obtained values. GalA amounts (mg/g seed) for (a) outer mucilage and (b) inner mucilage extracts, or (c) intrinsic viscosity (mL/g) of outer mucilage extracts. Blue bars indicate the values for two technical replicates obtained for samples analyzed in this study (culture e or f, respectively). Grey bars indicate published values corresponding to the following references: 1, Macquet et al.3; 2, Saez et al.19; 3, Ralet et al.11; 4, Griffith et al.22; 5, Poulain et al.20; 6, Saez et al.23; 7, Cambert et al.21.
For quantification of GalA concentrations, a standard curve was established both before and after a series of samples, using a serial dilution of GalA at 20, 40, 60, 80, and 100 μg/mL, to confirm technical rigour. HP-SEC columns were calibrated for IV using both a calibrant and a standard sample passed at the beginning, middle and end of a series of samples to check that no drift occurred over time.
Code availability
The different available software and the versions used to acquire and process data presented in the dataset are summarized in Table 7.
References
North, H. M., Berger, A., Saez-Aguayo, S. & Ralet, M.-C. Understanding polysaccharide production and properties using seed coat mutants: future perspectives for the exploitation of natural variants. Ann. Bot. 114, 1251–1263 (2014).
Francoz, E., Ranocha, P., Burlat, V. & Dunand, C. Arabidopsis seed mucilage secretory cells: regulation and dynamics. Trends Plant Sci. 20, 515–524 (2015).
Macquet, A., Ralet, M.-C., Kronenberger, J., Marion-Poll, A. & North, H. M. In situ, chemical and macromolecular study of the composition of Arabidopsis thaliana seed coat mucilage. Plant Cell Physiol. 48, 984–999 (2007).
Yu, L. et al. CELLULOSE SYNTHASE-LIKE A2, a glucomannan synthase, is involved in maintaining adherent mucilage structure in Arabidopsis seed. Plant Physiol. 164, 1842–1856 (2014).
Takenaka, Y. et al. Pectin RG-I rhamnosyltransferases represent a novel plant-specific glycosyltransferase family. Nature Plants 4, 669–676 (2018).
Voiniciuc, C. et al. Identification of Key Enzymes for Pectin Synthesis in Seed Mucilage. Plant Physiol. 178, 1045–1064 (2018).
Kong, Y. et al. GALACTURONOSYLTRANSFERASE-LIKE5 Is Involved in the Production of Arabidopsis Seed Coat Mucilage. Plant Physiol. 163, 1203–1217 (2013).
Fabrissin, I. et al. Natural Variation Reveals a Key Role for Rhamnogalacturonan I in Seed Outer Mucilage and Underlying Genes. Plant Physiol. 181, 1498–1518 (2019).
Amos, R. A. et al. Polymerization of the backbone of the pectic polysaccharide rhamnogalacturonan I. Nature Plants 8, 1289–1303 (2022).
Voiniciuc, C., Günl, M., Schmidt, M. H.-W. & Usadel, B. Highly Branched Xylan Made by IRREGULAR XYLEM14 and MUCILAGE-RELATED21 Links Mucilage to Arabidopsis Seeds. Plant Physiol. 169, 2481–2495 (2015).
Ralet, M.-C. et al. Xylans Provide the Structural Driving Force for Mucilage Adhesion to the Arabidopsis Seed Coat. Plant Physiol. 171, 165–178 (2016).
Vazquez-Vilar, M. et al. A modular toolbox for gRNA-Cas9 genome engineering in plants based on the GoldenBraid standard. Plant Methods 12, 10 (2016).
Clough, S. J. & Bent, A. F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16, 735–743 (1998).
Aliaga-Franco, N. et al. Identification of Transgene-Free CRISPR-Edited Plants of Rice, Tomato, and Arabidopsis by Monitoring DsRED Fluorescence in Dry Seeds. Front. Plant Sci. 10, 1150 (2019).
Sullivan, S. et al. CESA5 is required for the synthesis of cellulose with a role in structuring the adherent mucilage of Arabidopsis seeds. Plant Physiol. 156, 1725–1739 (2011).
Thibault, J. F. Automatisation du dosage des substances pectiques par la méthode au métahydroxydiphényle. Lebensml Wiss Technol 12, 247–251 (1979).
Blumenkrantz, N. & Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 54, 484–489 (1973).
Aoi et al. Biochemical data documenting variations in mucilage polysaccharide amount and macromolecular properties in a range of glycosyltransferase mutants. Portail Data INRAE https://doi.org/10.57745/DBSZ5X (2023).
Saez-Aguayo, S. et al. PECTIN METHYLESTERASE INHIBITOR6 promotes Arabidopsis mucilage release by limiting methylesterification of homogalacturonan in seed coat epidermal cells. Plant Cell 25, 308–323 (2013).
Poulain, D., Botran, L., North, H. M. & Ralet, M.-C. Composition and physicochemical properties of outer mucilage from seeds of Arabidopsis natural accessions. AoB Plants 11, lz031 (2019).
Cambert, M. et al. Datasets of seed mucilage traits for Arabidopsis thaliana natural accessions with atypical outer mucilage. Scientific Data 8, 1–9 (2021).
Griffiths, J. S., Crepeau, M.-J., Ralet, M.-C., Seifert, G. J. & North, H. M. Dissecting Seed Mucilage Adherence Mediated by FEI2 and SOS5. Front. Plant Sci. 7, 1073 (2016).
Saez-Aguayo, S. et al. Transport of UDP-rhamnose by URGT2, URGT4, and URGT6 modulates rhamnogalacturonan-I length. Plant Physiol. 185, 914–933 (2020).
Acknowledgements
We thank Fabien Nogue, Zsolt Kelemen and Anne Frey for technical advice on the CRISPR/Cas9 gene-editing technique used to generate mutants. We also thank Lionel Gissot for providing the pGBD3 TUs containing proRPS5A:hCas9:ter and proCMV:DsRED:terNos used in GoldenBraid cloning. Y.A was supported by PhD fellowship funding from INRAE (BAP and TRANSFORM divisions) and the Pays de Loire region. The IJPB benefits from the support of Saclay Plant Sciences-SPS (ANR-17-EUR-0007, EUR SPS-GSR).
Author information
Authors and Affiliations
Contributions
Y.A., M.-C.R. and H.M.N. conceived and designed the research. Y.A. and A.B. genotyped and produced seeds. Y.A. and L.S. performed biochemical analyses. Y.A., L.S., M.-C.R. and H.M.N. analyzed the data. Y.A., M.-C.R. and H.M.N. wrote the manuscript. All authors read and approved the manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Aoi, Y., Benamar, A., Saulnier, L. et al. Biochemical data documenting variations in mucilage polysaccharides in a range of glycosyltransferase mutants. Sci Data 10, 702 (2023). https://doi.org/10.1038/s41597-023-02604-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41597-023-02604-2
- Springer Nature Limited