
Abstract
Mitochondria are organelles that produce cellular energy in the form of ATP through oxidative phosphorylation, and this primary function is conserved among many taxa. Locomotion is a trait that is highly reliant on metabolic function and expected to be greatly affected by disruptions to mitochondrial performance. To this end, we aimed to examine how activity and sleep vary between Drosophila melanogaster strains with different geographic origins, how these patterns are affected by mitochondrial DNA (mtDNA) variation, and how breaking up co-evolved mito-nuclear gene combinations affect the studied activity traits. Our results demonstrate that Drosophila strains from different locations differ in sleep and activity, and that females are generally more active than males. By comparing activity and sleep of mtDNA variants introgressed onto a common nuclear background in cytoplasmic hybrid (cybrid) strains, we were able to quantify the among-line variance attributable to mitochondrial DNA, and we establish that mtDNA variation affects both activity and sleep, in a sex-specific manner. Altogether our study highlights the important role that mitochondrial genome variation plays on organismal physiology and behaviour.
Similar content being viewed by others


Introduction
Mitochondria are key organelles in a range of critical metabolic processes and are the primary energy producers for the eukaryotic cell. In addition to this primary role, they are also involved in a range of other vital processes that control key aspects of cellular growth and regulation such as signalling (Chandel Navdeep 2015), cellular differentiation (Vega-Naredo et al. 2014), cell death (Wang and Youle 2009) and immunity (Buchanan et al. 2018; Salminen and Vale 2020). The mitochondrial machinery responsible for ATP production via oxidative phosphorylation (OXPHOS) is jointly encoded by the mitochondrial (mtDNA) and nuclear genomes. While the mtDNA genome encodes for 13 protein-coding OXPHOS genes, the nuclear genome encodes the majority of the OXPHOS subunits as well as over 1200 other genes required for mitochondrial function (Anderson et al. 1981; Gray et al. 1999a; Gray et al. 1999b; Lang et al. 1999). In addition to vast differences in genome size, there are large differences in copy number between the genomes, with up to hundreds of mtDNA copies inhabiting each diploid cell (Robin and Wong 1988). Consequently, precise and synchronised coordination between the two genomes is required for proper assembly and function of the components of the electron transport chain and mitochondrial functions. Disruptions to this system—by mutations in the mitochondrial or nuclear counterparts—can have consequences on a wide range of life-history phenotypes (Hill et al. 2019), and in severe cases lead to mitochondrial disease (DiMauro and Schon 2003; Salminen et al. 2019; Schon et al. 2012; Smeitink et al. 2001; Wallace 1994). A consequence of this tight intergenomic partnership is that any trait with heavy metabolic underpinnings is reliant on the compatibility between mitochondrial and nuclear genomes.
Mitochondrial dysfunction affects a wide range of metabolic and behavioural traits, given the key role of mitochondria in energetics (Ghaoui and Sue 2018). Two such traits which are vital for everyday function are locomotor activity and sleep. Sleep is integral to regular brain function, influencing processes such as learning and memory (Ganguly-Fitzgerald et al. 2006), and plays a role in cellular processes such as metabolic recovery and oxidative stress (Trivedi et al. 2017). Continued sleep deprivation results in fatality for both invertebrate and vertebrate species (Potdar and Sheeba 2013; Rechtschaffen et al. 1989; Shaw et al. 2002). In humans, sleep deprivation is associated with an increased risk of metabolic and cognitive disorders (Harbison et al. 2013). In Drosophila, sleep deprivation has been shown to be connected to mitochondrial bioenergetics and causes mitochondrial dysfunction (Rodrigues et al. 2018). However, investigation of sleep as a consequence of mitochondrial dysfunction appears to be an understudied aspect of mitochondrial disease (Brunetti et al. 2021). It is also unclear how naturally occurring mtDNA variation may affect sleep.
Locomotion is also heavily reliant on metabolic function, with studies finding a strong positive correlation between activity and resting metabolic rate (Videlier et al. 2019). Moreover, both increased fatigue and exercise intolerance have been linked to several metabolic disorders originating from mitochondrial dysfunction (Filler et al. 2014; Sujkowski et al. 2019). For instance, Drosophila models of neuromuscular degeneration have shown a progressive decrease in fly activity with time (Bar et al. 2018). Additionally, nutritional metabolic interventions in the form of dietary restriction have shown to change activity patterns (Ghimire and Kim 2015), while physiologically costly immune stimulation has also been found to result in reduced locomotor activity in some insect species (Vale and Jardine 2015; Gupta et al. 2017; Vale et al. 2018; Vincent et al. 2021).
Given their tight link with metabolism, mutations affecting mitochondrial function are predicted to affect both sleep and locomotor activity. A previous review of primary mitochondrial diseases described sleep disorders in humans associated with mutations in mitochondrial DNA (Ramezani and Stacpoole 2014), but in general, we know little about how mitochondrial variation may affect activity and sleep patterns (Fogle et al. 2019; Ramezani and Stacpoole 2014). The fruit fly, Drosophila melanogaster, offers a powerful system to address the link between mitochondrial variation and activity and sleep disruption. Drosophila is an established genetic model system, including the study of mito-nuclear effects on various phenotypic traits (Camus and Dowling 2018; Hoekstra et al. 2013; Holmbeck et al. 2015; Zhu et al. 2014). As mtDNA is maternally inherited, introgression enables the generation of flies with specific combinations of nDNA and mtDNA. Generation of cytoplasmic hybrids (cybrids) therefore allows the effects of mtDNA mutations to be disentangled from nuclear genome variation (Rand et al. 2004). A large body of work using Drosophila cybrid lines has established that several measures of life-history phenotypes are modulated by changes in the mitochondrial genome, including aging (Camus et al. 2012; Camus et al. 2020; Rand et al. 2006), fitness (Camus and Dowling 2018; Mossman et al. 2019; Salminen et al. 2017), and metabolic rate (Nagarajan-Radha et al. 2020).
Drosophila is also an established model for the study of sleep and circadian rhythms and displays a state of quiescence that shares critical features of mammalian sleep (Shaw et al. 2000). These similarities include an elevated arousal threshold (Shaw et al. 2000), altered brain electrical activity (Nitz et al. 2002) and a decrease in amount of sleep as flies age (Hendricks et al. 2000). Furthermore, gene expression associated with ‘waking’ in fruit flies has been shown to correlate with ‘waking’ genes in mammals (Shaw et al. 2000). A relevant example is the mtDNA-encoded Cytochrome oxidase C, subunit I, which has been demonstrated to have elevated expression during the initial hours following sleep in both Drosophila and rats (Shaw et al. 2000). This homology between Drosophila and mammalian sleep, combined with the knowledge that many of the genetic and molecular regulators of sleep are conserved between flies and humans (Crocker and Sehgal 2010), has prompted extensive use of the fruit fly as a genetically tractable model organism in the study of sleep.
To specifically address the role of variation in the mitochondrial DNA on activity and sleep, we examined the sleep–wake cycles and activity profiles of a worldwide collection of eight D. melanogaster lines, in addition to a set of derived cybrid lines which contained each of the eight mtDNA variants introgressed onto a single common nuclear background. This experimental setup allowed us to investigate the baseline activity and sleep profiles of each line and to assess the contribution of mtDNA to these phenotypes. Further, because the mitochondrial genome of each line presents unique mtDNA variation at the haplotype level as well as common variants at the haplogroup level (Salminen et al. 2017) we were able to test if variation in sleep and activity patterns were associated with specific haplotypes.
Materials and methods
Fly strains, backcrossing and rearing conditions
We sourced eight wild-type D. melanogaster strains, with distinct geographic origins (Table 1), originally obtained from the Drosophila Stock Center (Bloomington, IN). Based on the mtDNA coding region variation the eight mtDNA variants form two distinct haplogroups with a set of few common replacement variants present in haplogroup I in OXPHOS complexes I (ND1; V190M, ND2; I277L, ND5; M502I) and V (ATP6; S538P and M559V) (Salminen et al. 2017). Haplogroup II contains European mtDNA variants (mtPYR2, mtLS and mtBS1), whereas haplogroup I contains variants from different continents (mtORT, mtKSA2, mtBOG1, mtWT5A and mtM2) (Table 1). Cybrid lines were created earlier by backcrossing females from each strain (carrying unique mtDNA variant) to males from the nuclear-donor strain of the Oregon RT strain (Oregon R strain maintained long-term in Tampere, Finland; ORT) for at least 12 generations (Salminen et al. 2017). This resulted in a total of 16 strains; 8 of which were the Bloomington-derived strains representing coevolved mito-nuclear combinations and 8 were nORT mtDNAx cybrids. All lines were cultured on standard Lewis medium (Lewis 1960), supplemented with yeast, under 12: 12 light: dark cycles at 25 °C and 60% humidity. Flies were propagated by placing 30, 2–4-day old females and males on food vials for 3–4 days, with adults being discarded and egg clutches kept. This rearing regime maintained egg densities low enough to prevent larval overcrowding.
Experimental design
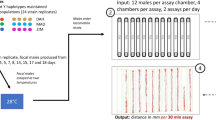
The Drosophila Activity Monitor (DAM2, Trikinetics) was used to collect all experimental data on fly activity and sleep cycles (Chiu et al. 2010). Prior to the experiment, a solution of 8% sucrose: 2% agar was prepared in distilled water and autoclaved for sterilisation and stored at room temperature before re-melting for use. DAM2 tubes (5 mm diameter, 65 mm length) were prepared by adding ~1 cm sucrose-agar medium to one end of the tube and sealing with a rubber cap. This provided a source of food and moisture for the fly during the experiment. Flies from each coevolved and cybrid strain were collected upon eclosion and kept in food vials for 2 days. Flies were anesthetised with CO2, sorted by sex and transferred into a DAM2 tube using a fine paintbrush. After the anesthetised fly had been inserted into the DAM2 tube it was closed with a rubber cap containing a small hole for ventilation (Chiu et al. 2010).
Fifteen individuals per sex and strain were tested across five experimental blocks, with 3–5 replicates per block for each line/sex combination positioned haphazardly in the activity monitors. Each monitor had a null or blank control in which a recorded position contained an empty tube or no tube, respectively. Monitoring of fly activity and sleep lasted for three continuous days. Each DAM2 tube is bisected by an infra-red beam and locomotor activity movement was recorded whenever a fly broke the beam. The number of activity counts (beam breaks) generated by the DAM was used to quantify locomotor activity. Sleep was defined as a 5 min time-bin with no registered activity (Andretic and Shaw 2005). To determine whether individuals/flies are more active simply because they sleep less, we calculated the proportion of time each replicate spends sleeping, as well as the mean activity count during awake periods. The experiments were run under 12:12 light:dark cycles at 25 °C at constant temperature and humidity.
Statistical analysis
Total activity count, mean awake activity and proportion of time sleeping were calculated for each individual fly (see also Vale and Jardine 2015). Four flies (out of 480) died during the experiment and were excluded from analysis. We analysed the co-evolved and cybrid lines separately, using otherwise identical statistical models. Total activity count and mean awake activity were analysed using linear mixed-effects models (LME). Models fitted ‘line’, ‘sex’ and their interaction as categorical fixed effects. Variation in proportion of time asleep was analysed using generalised linear effects models (GLME) assuming binomial distributed error, which also fitted ‘line’, ‘sex’ and their interaction as categorical fixed effects. All models included the random effect of ‘replicate’ nested within ‘block’ to account for variation between individuals among different blocks. We also investigated if breaking up co-evolved mito-nuclear gene complexes affected each of these behavioural outputs. This analysis therefore included the coevolved and cybrid fly lines to analyse the effect of mtDNA variant on its coevolved or cybrid nuclear background. Similar model structure was used as described above for each response variable, where each model fitted ‘type’ (coevolved or cybrid), ‘sex’ and their interactions as categorical fixed effects, and ‘line’ as a random effect nested within type. R version 1.1.4 (Team RC 2019) was used for analysis and plots, using packages ggplot2 (Wickham 2009), dplyr (Wickham et al. 2019), lme4 (Bates et al. 2012), car (Fox and Weisberg 2011) and plotrix (Lemon 2016). All datasets and full R code for all analyses can be found at https://doi.org/10.5281/zenodo.5573904.
Results
Coevolved fly lines show sex-specific natural variation in sleep and activity patterns
We first evaluated the sleep and activity patterns of genetically and geographically diverse fly lines carrying natural, and presumably coevolved, mito-nuclear combinations (Table 1). Activity profiles showed that all lines were crepuscular, exhibiting a peak of activity at the onset of the dark period (Fig. S1). In the majority of strains, females were significantly more active than males (14% more active—female mean total activity: 1988, male: 1750). The extent of this difference was influenced by genotype and in lines BS1 and WT5A, males were more active than females (Table 2 ‘Line × Sex’ effect; Fig. 1A and Fig. S2). Overall, males spent a higher proportion of time asleep (64% of the time) compared to females (55% of the time), and the extent of this difference varied between genetic backgrounds (Table 2 ‘Line × Sex’ effect). Notably, while males slept for a greater proportion of the day, they were slightly more active while awake (5.74 recorded movements per 5-min bout) compared to females (5 recorded movements per 5-min bout), and again the extent of this variation differed between lines (Table 2 ‘Line × Sex’ effect; Fig. 1B, C). This suggests that although females exhibit higher total activity, this is due to females spending more time awake (sleeping less) rather than having higher levels of activity when awake.

The total number of activity events recorded over three days in original (A) and cybrid (D) lines. The total number of activity events recorded when the flies were awake in original (B) and cybrid (E) lines. The proportion of time that flies were asleep, defined as 5 min of inactivity in original (C) and cybrid (F) lines. See Figs S1-S4 for individual actograms and boxplots for each line. See Table 1 for details of each line. See Table 2 for outputs of statistical models and Table S1 for random effect variance components.
Mitochondrial genome variation affects activity and sleep patterns of the cybrid lines
By introgressing the eight mtDNA variants onto a common nuclear background (ORT), we were able to evaluate how much of the variance in locomotor activity was affected by the mtDNA and newly created mito-nuclear combinations (Figs. S3, S4, Table S1). In general, comparing total activity counts showed that as observed with coevolved lines, females of each cybrid line were significantly more active than cybrid males (45% more active - female mean total activity: 3847, male: 2652), and the extent of this variation was mtDNA specific (Table 2 ‘Line × Sex’ effect; Fig. 1E and Fig. S4). Therefore, compared to the coevolved lines, cybrid lines exhibited a substantial 75% increase in the total activity on average (3266 recorded movements compared to 1867 on average in the original lines), and the differences between sexes, while consistent in showing females are more active, were also more pronounced in cybrid lines (45% compared to 14%) (Fig. 1A, D). Males were found to sleep for a significantly larger proportion of time (46% of the time) compared to females (30% of the time) (Fig. 1B, E), although unlike the co-evolved lines, the female cybrids were generally more active when awake (6.3 recorded movements per 5-min bout) when compared to males (5.6 recorded movements per 5-min bout) (Fig. 1C, F). Both of these differences were also mediated by mtDNA variation (Table 2 ‘Line × Sex’ effect). Therefore, when the effects of individual mitochondrial genomes are isolated on a common nuclear background females are both more active while awake and also spend less time sleeping than males.
Haplogroup-specific mtDNA variation can be seen in the activities of cybrid females
Based on the mtDNA coding region variation the eight mtDNA variants form two distinct haplogroups with a set of few common replacement variants present in haplogroup I in OXPHOS complexes I (ND1; V190M, ND2; I277L, ND5; M502I) and V (ATP6; S538P and M559V) (Salminen et al. 2017). Haplogroup I contains the haplotypes mtORT, mtKSA2, mtBOG1, mtWT5A and mtM2, as the haplogroup II contains the haplotypes mtPYR2, mtLS and mtBS1 (Table 1). Haplogroup division did not cause clear differences in the sleep-wake activities when the haplogroup-specific mtDNA variants were present in their coevolved nuclear backgrounds and the sex differences that were observed earlier with the coevolved lines were decreased (Table 3, Fig. 2. upper row). However, when the haplogroup I and II mtDNA variants were placed on a novel common nuclear background in the cybrid lines, we observed that haplogroup II females we more active than haplogroup I females (Table 3, Fig. 2, lower row A). Haplogroup II females were also more active when awake and slept for a smaller proportion of time than haplogroup I females (Table 3, Fig. 2 lower rows B and C).

Locomotor activities and sleep in coevolved and cybrid females and males based on the mtDNA haplogroup division. A Total activity counts over three days. B Activity of the flies when awake. C The proportion of time the flies spent sleeping.
Breaking up co-evolved mito-nuclear combinations affects activity and sleep patterns
As naturally occurring mito-nuclear genome combinations have co-adapted locally over time, we expected that disruption of these combinations could result in maladaptive effects. We also expected that these effects might be more severe, or more variable in males, as postulated under the mother’s curse hypothesis (Carnegie et al. 2021; Dowling and Adrian 2019). Both male and female cybrid flies were significantly more active than flies with co-evolved mito-nuclear combinations (on average, 75% more active), although the extent of this variation differed between sexes and was more prominent in females (Table 2 ‘Line × Sex’ effect; Fig. 3A). Part of the increase in total activity in cybrids was driven by a 57% increase in the awake activity level (2.2–3.8 recorded movements per 5-min bout, on average, in the original and cybrid line, respectively), though this was the case mainly in females (Table 2 ‘Line × Sex’ effect; Fig. 3B). However, the largest driver of the increased total activity in both male and female cybrid flies is that these spent a significantly lower proportion of their time asleep (38% of the time) compared to the co-evolved lines (59% of the time), and the extent of this difference was slightly larger in females (Table 2 ‘Line × Sex’ effect; Fig. 3C).

A total number of activity events recorded over 3 days. B the total number of activity events recorded when flies were not asleep. C The proportion of time that flies were determined to be asleep, defined as 5 min of inactivity, See Table 2 for outputs of statistical models.
Given that our design was not orthogonal and only one nuclear genotype was tested across several mitochondrial genotypes, we cannot conclusively quantify the effect of nuclear variation. However, we are able to calculate the proportion of variance explained in each trait by mtDNA, as ratio of the among-line variance in the cybrids with the among-line variance in original lines (Table 4). This analysis shows that for both total activity and sleep, mtDNA explained only a fraction of variance within lines (total activity 20%; sleep 14%, Table 4). However, we were surprised to see that the among-line variance in awake activity in the original lines was very low (2%) and that when isolated on a common nuclear background, mtDNA explained six times more variance compared to the original lines (Table 4).
Discussion
Mitochondrial dysfunction commonly manifests in tissues with high metabolic demands, such as muscles, with movement disorders being a major feature of mitochondrial diseases (Ghaoui and Sue 2018). However, studying the effect of mtDNA variation is challenging as it is necessary to disentangle the effects of the mitochondrial genome from the effect of the nuclear background. This is feasible with the Drosophila cybrid model, and here we have focused on studying the effect of mtDNA variation on locomotion activity and sleep. We addressed three questions by separately quantifying the contributions of the mitochondrial and nuclear genomes to activity and sleep phenotypes. First, we asked how variation in both the nDNA and mtDNA affects sleep and activity. Second, by isolating mtDNA variants on a common nuclear background we investigated how variation in mtDNA affects sleep and activity. Finally, we assessed how breaking up co-evolved mito-nuclear genetic interactions affected sleep and activity.
Locomotor activity and the proportion of time spent asleep were first measured from eight wild-type Drosophila strains with co-evolved mito-nuclear combination adapted to their local environment. These strains showed variation in their sleep-wake patterns and in general females were more active overall; males exhibited a higher waking activity but slept for a larger proportion of time. It is hypothesised that inseminated females spend more time awake as they have to lay eggs and scavenge for food to maintain high levels of fecundity. These results are supported by previous literature that finds genotype-specific and sexual dimorphism for this trait (Hyde and Jerussi 1983; Long and Rice 2007). We next aimed to test if mtDNA contributed to this variation between lines. The eight studied mtDNA genomes can be subdivided into two haplogroups based on their genetic variation (Salminen et al. 2017). Interestingly, haplogroup II females were shown to have higher total activity levels and spent less time sleeping when compared to haplogroup I females. The same was not observed with males. It is unclear how these patterns might have arisen in nature. However further experimentation, replicated in both lab and natural settings, is required to examine if these patterns are a result of local adaptation, or if they could arise due to independent compensatory mito-nuclear co-evolution, or even just due to drift.
As the eight cybrids also possess mitochondrial haplotype-specific mutations, we examined their effects on locomotion and sleep when introgressed into common nuclear background in the cybrid lines. In general, we were able to see differences in the amounts of activity and sleep, in both females and males, brought upon by mtDNA variation. Cybrid females were more active than males, sleep less and have higher waking activity. It is difficult to say if specific mtDNA mutations are causing the seen variation, as most of these mtDNA genomes contain more than one source of variation, i.e replacement variants in the protein-coding genes, synonymous SNPs, indels in the tRNA and rRNA genes and also length variation in the non-coding A + T region (Salminen et al. 2017). Here we focused on non-synonymous SNPs between our cybrid strains as they are predicted to have larger effect size, however several studies have shown that synonymous SNPs and SNPs in tRNAs also have a significant phenotypic effect (Meiklejohn et al. 2013; Camus et al. 2017). Mitovariant mtM2, the only variant that is originally from Australia, contains most unique non-synonymous mutations when compared to the other mitovariants (Table 1). mtM2 was also one of the lowest activity strains, and females especially spent more time sleeping when compared to the other cybrids. This might be due to altered interactions between mitochondrial and nuclear gene products (Pichaud et al. 2019). M2 flies which have the coevolved nuclear background actually appear to exhibit relatively high activity and low amount of sleep. Disruption of naturally occurring mito-nuclear combinations can result in interruption of precise interactions, leading to mito-nuclear incompatibilities, which have downstream deleterious fitness consequences (Mossman et al. 2019).
mtKSA2 is the only African variant, and it possesses two unique amino acid replacement variants in OXPHOS cIII (CYTB) and cIV (COIII) when compared to other cybrids (Salminen et al. 2017). mtKSA2 cybrids males exhibited the highest proportion of time spent sleeping. Also, the overall activity counts were among the lowest in mtKSA2 females and males, and the mtKSA2 cybrids females also had the lowest awake activity counts. The mtDNA variation studied here is maternally inherited and its effects can be multisystemic, affecting each tissue in Drosophila. However, there are cases where several sporadic missense and nonsense CYTB mutations in muscles have been shown to cause complex III deficiency and exercise intolerance in humans (Andreu et al. 1999). CYTB mutation in mtKSA2 may partially explain the lower activity rates when compared to other strains.
mtKSA2 is also associated with low mtDNA copy number in ORT nuclear background when compared to other cybrid lines (Salminen et al. 2017). Since mtDNA copy number has been shown to be associated with other measures of fitness in Drosophila including fertility and longevity (Camus et al. 2015) and with development time and weight (Salminen et al. 2017) of the same co-evolved and cybrid lines as studied here, it is possible that it might also be associated with variation in activity levels. mtDNA copy number is sexually dimorphic in Drosophila, with females of most strains tending to exhibit a higher copy number than males (Camus et al. 2015; Salminen et al. 2017), and is also affected by the age of the flies, sex specifically (Salminen et al. 2017). Sleep and activity are also shown to be sexually dimorphic traits in Drosophila (Long and Rice 2007; Shaw et al. 2000). In our study, we saw that in both co-evolved and cybrid strains the females were more active overall and slept less, as shown also with previous Drosophila sleep and activity research (Isaac et al. 2010). A future line of enquiry is therefore whether mtDNA copy number levels correlate with the extent of locomotor activity and sleep.
Previous studies have demonstrated that disrupting co-evolved mito-nuclear interactions can lead to decreased OXPHOS function (Sackton et al. 2003; Salminen et al. 2017). Specifically, cytochrome c oxidase activity has been shown to be reduced when an mtDNA haplotype is combined with a more distant nuclear background rather than its coevolved nuclear background in copepod Tigriopus californicus (Willett and Burton 2003). Mito-nuclear incompatibilities can lead to reduction in fitness (Clancy 2008; Clancy et al. 2011; Yee et al. 2013), and maintaining normal levels of activity is beneficial to fitness to allow, for example foraging and mating to take place (Long and Rice 2007). Short sleep duration in Drosophila has previously been shown to be associated with poor memory (Bushey et al. 2007), and reduced longevity (Bushey et al. 2010; Cirelli et al. 2005).
Natural selection is blind to deleterious mitochondrial phenotypes which manifest in males as mtDNA is maternally inherited (Carnegie et al. 2021; Frank 2012; Gemmell et al. 2004). This can indicate that variation which persists in mtDNA is more likely to cause greater phenotypic divergence within males and more likely to cause deleterious effects in males than in females (Frank 2012; Gemmell et al. 2004). While several studies provide evidence in support of this theory, known as the mother’s curse (Camus et al. 2012; Carnegie et al. 2021; Ruiz-Pesini et al. 2000), our data here do not suggest the mother’s curse impacts activity. Although we observed a general increase in activity and decrease in sleep in both males and female cybrids, the magnitude of these effects was larger in females, and not in males, as would be predicted under the mother’s curse hypothesis.
Drosophila sleep and activity research will enable further understanding of the causes of abnormal sleep and activity patterns in humans and the role of mitochondrial variation in these traits. Further studies of mito-nuclear interactions and mtDNA variation are essential in recognising and eventually preventing mito-nuclear mismatches that might occur during mitochondrial replacement therapy.
Data archiving
All datasets and full R code for all analyses can be found at https://doi.org/10.5281/zenodo.5573904 and is fully citable as Vale PF (2021) Data and code for Mitonuclear interactions affect locomotor activity and sleep in Drosophila melanogaster [Data set]. Zenodo. https://doi.org/10.5281/zenodo.5573904.
References
Anderson S, Bankier AT, Barrell BG, de Bruijn MH, Coulson AR, Drouin J et al. (1981) Sequence and organization of the human mitochondrial genome. Nature 290(5806):457–465
Andretic R, Shaw PJ (2005) Essentials of sleep recordings in Drosophila: moving beyond sleep time. Methods Enzymol 393:759–772
Andreu AL, Hanna MG, Reichmann H, Bruno C, Penn AS, Tanji K et al. (1999) Exercise intolerance due to mutations in the cytochrome b gene of mitochondrial DNA. N Engl J Med 341(14):1037–1044
Bar S, Prasad M, Datta R (2018) Neuromuscular degeneration and locomotor deficit in a Drosophila model of mucopolysaccharidosis VII is attenuated by treatment with resveratrol. Dis Models Mech 11(11):dmm036954
Bates D, Maechler M, Bolkler B (2012) lme4: Linear mixed-effects models using S4 classes. R package version 0999999-0. https://cran.r-project.org/web/packages/lme4/index.html
Brunetti V, Della Marca G, Servidei S, Primiano G (2021) Sleep disorders in mitochondrial diseases. Curr Neurol Neurosci Rep. 21(7):30
Buchanan JL, Meiklejohn CD, Montooth KL (2018) Mitochondrial dysfunction and infection generate immunity-fecundity tradeoffs in Drosophila. Integr Comp Biol 58(3):591–603
Bushey D, Huber R, Tononi G, Cirelli C (2007) Drosophila Hyperkinetic mutants have reduced sleep and impaired memory. J Neurosci 27(20):5384–5393
Bushey D, Hughes KA, Tononi G, Cirelli C (2010) Sleep, aging, and lifespan in Drosophila. BMC Neurosci 11:56
Camus MF, Clancy DJ, Dowling DK (2012) Mitochondria, maternal inheritance, and male aging. Curr Biol 22(18):1717–1721
Camus MF, Dowling DK (2018) Mitochondrial genetic effects on reproductive success: signatures of positive intrasexual, but negative intersexual pleiotropy. Proc Biol Sci 285:1879
Camus MF, O’Leary M, Reuter M, Lane N (2020) Impact of mito-nuclear interactions on life-history responses to diet. Philos Trans R Soc Lond B Biol Sci 375(1790):20190416
Camus MF, Wolf JB, Morrow EH, Dowling DK (2015) Single nucleotides in the mtDNA sequence modify mitochondrial molecular function and are associated with sex-specific effects on fertility and aging. Curr Biol 25(20):2717–2722
Camus MF, Wolff JN, Sgrò CM, Dowling DK (2017) Experimental Support That Natural Selection Has Shaped the Latitudinal Distribution of Mitochondrial Haplotypes in Australian Drosophila melanogaster. Mol Biol Evol 34:2600–2612
Carnegie L, Reuter M, Fowler K, Lane N, Camus MF (2021) Mother’s curse is pervasive across a large mito-nuclear Drosophila panel. Evol Lett 5(3):230–239
Chandel NS (2015) Evolution of mitochondria as signaling organelles. Cell Metab 22:204–206
Chiu JC, Low KH, Pike DH, Yildirim E, Edery I (2010) Assaying locomotor activity to study circadian rhythms and sleep parameters in Drosophila. JoVE (Journal of Visualized Experiments), (43):e2157
Cirelli C, Bushey D, Hill S, Huber R, Kreber R, Ganetzky B et al. (2005) Reduced sleep in Drosophila Shaker mutants. Nature 434(7037):1087–1092
Clancy DJ (2008) Variation in mitochondrial genotype has substantial lifespan effects which may be modulated by nuclear background. Aging Cell 7(6):795–804
Clancy DJ, Hime GR, Shirras AD (2011) Cytoplasmic male sterility in Drosophila melanogaster associated with a mitochondrial CYTB variant. Heredity 107(4):374–376
Crocker A, Sehgal A (2010) Genetic analysis of sleep. Genes Dev 24(12):1220–1235
DiMauro S, Schon EA (2003) Mitochondrial respiratory-chain diseases. N Engl J Med 348(26):2656–2668
Dowling DK, Adrian RE (2019) Challenges and prospects for testing the mother’s curse hypothesis. Integr Comp Biol 59(4):875–889
Filler K, Lyon D, Bennett J, McCain N, Elswick R, Lukkahatai N et al. (2014) Association of mitochondrial dysfunction and fatigue: a review of the literature. BBA Clin 1:12–23
Fogle KJ, Mobini CL, Paseos AS, Palladino MJ (2019) Sleep and circadian defects in a Drosophila model of mitochondrial encephalomyopathy. Neurobiol Sleep Circadian Rhythms 6:44–52
Fox J, Weisberg S (2011). An R companion to applied regression, Second Edition
Frank SA (2012) Evolution: mitochondrial burden on male health. Curr Biol 22(18):R797–799
Ganguly-Fitzgerald I, Donlea J, Shaw PJ (2006) Waking experience affects sleep need in Drosophila. Science 313(5794):1775–1781
Gemmell NJ, Metcalf VJ, Allendorf FW (2004) Mother’s curse: the effect of mtDNA on individual fitness and population viability. Trends Ecol Evol 19(5):238–244
Ghaoui R, Sue CM (2018) Movement disorders in mitochondrial disease. J Neurol 265(5):1230–1240
Ghimire S, Kim MS (2015) Enhanced locomotor activity is required to exert dietary restriction-dependent increase of stress resistance in Drosophila. Oxid Med Cell Longev 2015:813801–813801
Gray MW, Burger G, Cedergren R, Golding GB, Lemieux C, Sankoff D et al. (1999a) A genomics approach to mitochondrial evolution. Biol Bull 196(3):400–403
Gray MW, Burger G, Lang BF (1999b) Mitochondrial evolution. Science 283(5407):1476–1481
Gupta V, Stewart CO, Rund SSC, Monteith K, Vale PF (2017) Costs and benefits of sublethal Drosophila C virus infection. J Evol Biol 30:1325–1335
Harbison ST, McCoy LJ, Mackay TF (2013) Genome-wide association study of sleep in Drosophila melanogaster. BMC Genom 14:281
Hendricks JC, Finn SM, Panckeri KA, Chavkin J, Williams JA, Sehgal A et al. (2000) Rest in Drosophila is a sleep-like state. Neuron 25(1):129–138
Hyde JF, Jerussi TP (1983) Sexual dimorphism in rats with respect to locomotor activity and circling behavior. Pharmacol Biochem Behav 18:725–729
Hill GE, Havird JC, Sloan DB, Burton RS, Greening C, Dowling DK (2019) Assessing the fitness consequences of mito-nuclear interactions in natural populations. Biol Rev Camb Philos Soc 94(3):1089–1104
Hoekstra LA, Siddiq MA, Montooth KL (2013) Pleiotropic effects of a mitochondrial–nuclear incompatibility depend upon the accelerating effect of temperature in Drosophila. Genetics 195(3):1129–1139
Holmbeck MA, Donner JR, Villa-Cuesta E, Rand DM (2015) A Drosophila model for mito-nuclear diseases generated by an incompatible interaction between tRNA and tRNA synthetase. Dis Model Mech 8(8):843–854
Isaac RE, Li C, Leedale AE, Shirras AD (2010) Drosophila male sex peptide inhibits siesta sleep and promotes locomotor activity in the post-mated female. Proc Biol Sci 277(1678):65–70
Lang BF, Gray MW, Burger G (1999) Mitochondrial genome evolution and the origin of eukaryotes. Annu Rev Genet 33:351–397
Lemon J (2016) Plotrix: a package in the red light district of R. R-N. 6(4):8–12
Lewis E (1960) A new standard food medium. Drosoph Inform Serv 34(117):1–55
Long TA, Rice WR (2007) Adult locomotory activity mediates intralocus sexual conflict in a laboratory-adapted population of Drosophila melanogaster. Proc Biol Sci 274(1629):3105–3112
Meiklejohn CD, Holmbeck MA, Siddiq MA, Abt DN, Rand DM, Montooth KL (2013) An Incompatibility between a Mitochondrial tRNA and Its Nuclear-Encoded tRNA Synthetase Compromises Development and Fitness in Drosophila. PLoS Genet 9:e1003238
Mossman JA, Ge JY, Navarro F, Rand DM (2019) Mitochondrial DNA fitness depends on nuclear genetic background in Drosophila. G3 9(4):1175–1188
Nagarajan-Radha V, Aitkenhead I, Clancy DJ, Chown SL, Dowling DK (2020) Sex-specific effects of mitochondrial haplotype on metabolic rate in Drosophila melanogaster support predictions of the Mother’s Curse hypothesis. Philos Trans R Soc Lond B Biol Sci 375(1790):20190178
Nitz DA, van Swinderen B, Tononi G, Greenspan RJ (2002) Electrophysiological correlates of rest and activity in Drosophila melanogaster. Curr Biol 12(22):1934–1940
Pichaud N, Berube R, Cote G, Belzile C, Dufresne F, Morrow G et al. (2019) Age-dependent dysfunction of mitochondrial and ROS metabolism induced by mito-nuclear mismatch. Front Genet 10:130
Potdar S, Sheeba V (2013) Lessons from sleeping flies: insights from Drosophila melanogaster on the neuronal circuitry and importance of sleep. J Neurogenet 27(1-2):23–42
Ramezani RJ, Stacpoole PW (2014) Sleep disorders associated with primary mitochondrial diseases. J Clin Sleep Med 10(11):1233–1239
Rand DM, Fry A, Sheldahl L (2006) Nuclear-mitochondrial epistasis and drosophila aging: introgression of Drosophila simulans mtDNA modifies longevity in D. melanogaster nuclear backgrounds. Genetics 172(1):329–341
Rand DM, Haney RA, Fry AJ (2004) Cytonuclear coevolution: the genomics of cooperation. Trends Ecol Evol 19(12):645–653
Rechtschaffen A, Bergmann BM, Everson CA, Kushida CA, Gilliland MA (1989) Sleep deprivation in the rat: X. Integration and discussion of the findings. Sleep 12(1):68–87
Robin ED, Wong R (1988) Mitochondrial DNA molecules and virtual number of mitochondria per cell in mammalian cells. J Cell Physiol 136(3):507–513
Rodrigues NR, Macedo GE, Martins IK, Gomes KK, de Carvalho NR, Posser T et al. (2018) Short-term sleep deprivation with exposure to nocturnal light alters mitochondrial bioenergetics in Drosophila. Free Radic Biol Med 120:395–406
Ruiz-Pesini E, Lapena AC, Diez-Sanchez C, Perez-Martos A, Montoya J, Alvarez E et al. (2000) Human mtDNA haplogroups associated with high or reduced spermatozoa motility. Am J Hum Genet 67(3):682–696
Sackton TB, Haney RA, Rand DM (2003) Cytonuclear coadaptation in Drosophila: disruption of cytochrome c oxidase activity in backcross genotypes. Evolution 57(10):2315–2325
Salminen TS, Cannino G, Oliveira MT, Lillsunde P, Jacobs HT, Kaguni LS (2019) Lethal interaction of nuclear and mitochondrial genotypes in Drosophila melanogaster. G3 9(7):2225–2234
Salminen TS, Oliveira MT, Cannino G, Lillsunde P, Jacobs HT, Kaguni LS (2017) Mitochondrial genotype modulates mtDNA copy number and organismal phenotype in Drosophila. Mitochondrion 34:75–83
Salminen TS, Vale PF (2020) Drosophila as a model system to investigate the effects of mitochondrial variation on innate immunity. Front Immunol 11:521
Schon EA, DiMauro S, Hirano M (2012) Human mitochondrial DNA: roles of inherited and somatic mutations. Nat Rev Genet 13(12):878–890
Shaw PJ, Cirelli C, Greenspan RJ, Tononi G (2000) Correlates of sleep and waking in Drosophila melanogaster. Science 287(5459):1834–1837
Shaw PJ, Tononi G, Greenspan RJ, Robinson DF (2002) Stress response genes protect against lethal effects of sleep deprivation in Drosophila. Nature 417(6886):287–291
Smeitink J, van den Heuvel L, DiMauro S (2001) The genetics and pathology of oxidative phosphorylation. Nat Rev Genet 2(5):342–352
Sujkowski A, Spierer AN, Rajagopalan T, Bazzell B, Safdar M, Imsirovic D et al. (2019) Mito-nuclear interactions modify Drosophila exercise performance. Mitochondrion 47:188–205
Team RC (2019). R: a language and environment for statistical computing. R Foundation for Statistical Computing
Trivedi MS, Holger D, Bui AT, Craddock TJA, Tartar JL (2017) Short-term sleep deprivation leads to decreased systemic redox metabolites and altered epigenetic status. PLoS One 12(7):e0181978
Vale PF (2021) Data and code for Mitonuclear interactions affect locomotor activity and sleep in Drosophila melanogaster [Data set]. Zenodo https://doi.org/10.5281/zenodo.5573904
Vale PF, Jardine MD (2015) Sex-specific behavioural symptoms of viral gut infection and Wolbachia in Drosophila melanogaster. J Insect Physiol 82:28–32
Vale PF, Siva-Jothy JA, Morrill A, Forbes MR (2018). The influence of parasites on insect behavior. In: Insect behavior: from mechanisms to ecological and evolutionary consequences, Oxford University Press
Vega-Naredo I, Loureiro R, Mesquita KA, Barbosa IA, Tavares LC, Branco AF et al. (2014) Mitochondrial metabolism directs stemness and differentiation in P19 embryonal carcinoma stem cells. Cell Death Differ 21(10):1560–1574
Videlier M, Rundle HD, Careau V (2019) Sex-specific among-individual covariation in locomotor activity and resting metabolic rate in Drosophila melanogaster. Am Nat 194(6):E164–E176
Vincent CM, Dionne MS (2021) Disparate regulation of IMD signaling drives sex differences in infection pathology in Drosophila melanogaster Proc Natl Acad Sci 118(32):e2026554118
Wallace DC (1994) Mitochondrial DNA sequence variation in human evolution and disease. Proc Natl Acad Sci USA 91(19):8739–8746
Wang C, Youle RJ (2009) The role of mitochondria in apoptosis*. Annu Rev Genet 43(1):95–118
Wickham H (2009). ggplot2: elegant graphics for data analysis. Use R: 1–212
Wickham H, Francois R, Henry L, Müller K (2019) dplyr: a grammar of data manipulation. R package version 0801:1–212
Willett CS, Burton RS (2003) Environmental influences on epistatic interactions: viabilities of cytochrome c genotypes in interpopulation crosses. Evolution 57(10):2286–2292
Yee WKW, Sutton KL, Dowling DK (2013) In vivo male fertility is affected by naturally occurring mitochondrial haplotypes. Curr Biol 23(2):R55–R56
Zhu C-T, Ingelmo P, Rand DM (2014) G×G×E for lifespan in drosophila: mitochondrial, nuclear, and dietary interactions that modify longevity. PLOS Genet 10(5):e1004354
Acknowledgements
We thank Angela Reid and Lucinda Rowe and the Ashworth Media team for fly medium preparation. Lucy Anderson was supported by the Genetics Honours Programme, School of Biological Sciences, University of Edinburgh. This work was partially supported by a Chancellor’s Fellowship (University of Edinburgh), a Branco Weiss Fellowship (Society in Science—Zürich, https://brancoweissfellowship.org/) and a Leverhulme Trust Research Grant (RPG-2018-369), all awarded to Pedro Vale.
Author information
Authors and Affiliations
Contributions
LA, TSS and PFV conceived the study. LA carried out the experimental work, with help from KM and TSS and supervised by PFV. LA, MFC and PFV analysed the data. PFV generated the final tables and figures and curated the code. LA, MFC, TSS and PFV drafted the manuscript. All authors commented on several drafts and approved the final version.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Associate editor: Gerald Heckel.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Anderson, L., Camus, M.F., Monteith, K.M. et al. Variation in mitochondrial DNA affects locomotor activity and sleep in Drosophila melanogaster. Heredity 129, 225–232 (2022). https://doi.org/10.1038/s41437-022-00554-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/s41437-022-00554-w
- Springer Nature Switzerland AG