
Abstract
Skin is a complex ecosystem colonized by millions of microorganisms, including bacteria, fungi, and viruses. Skin microbiota is believed to exert critical functions in maintaining host skin health. Profiling the structure of skin microbial community is the first step to overview the ecosystem. However, the community composition is highly individualized and extremely complex. To explore the fundamental factors driving the complexity of the ecosystem, namely the selection pressures, we review the present studies on skin microbiome from the perspectives of ecology. This review summarizes the following: (1) the composition of substances/nutrients in the cutaneous ecological environment that are derived from the host and the environment, highlighting their proposed function on skin microbiota; (2) the features of dominant skin commensals to occupy ecological niches, through self-adaptation and microbe–microbe interactions; (3) how skin microbes, by their structures or bioactive molecules, reshape host skin phenotypes, including skin immunity, maintenance of skin physiology such as pH and hydration, ultraviolet (UV) protection, odor production, and wound healing. This review aims to re-examine the host–microbe interactions from the ecological perspectives and hopefully to give new inspiration to this field.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The skin is considered a barrier organ against the entry of foreign physical, chemical, and biological insults, thereby maintaining the internal homeostasis of the human body. In the past decades, Human Microbiome Project (HMP) has expanded our perception of the skin as not only a piece of placid “soil” but a vast “ecosystem” that harbors a myriad of microbial inhabitants (Human Microbiome Project Consortium 2012). It has been believed that the colonization of diverse microbes resulted from millions of years of mutual adaptation and functional integration (Lousada et al. 2021), and thus the human body forms a complex, synergistic entity, termed a holobiont or meta-organism (Bosch and McFall-Ngai 2011; Rosenberg et al. 2007). The environmental and nutrient conditions define the unique microhabitats for skin microbes (Flowers and Grice 2020), and in turn, these microbes can influence their survival environment (host skin) by stabilizing, mutually beneficial host–microbe interactions (Postler and Ghosh 2017). In various disease conditions, the host–microbe interactions became imbalanced, termed “dysbiosis”, presenting various shifts in microbiome from “healthy” to “diseased” states (Thomas and Jobin 2020).
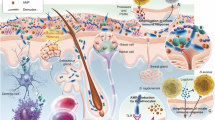
Profiling the structure of skin microbial community is the first step to overview the ecosystem and to address host–microbe interactions. However, this system was proven to be highly individualized and extremely complex. Many factors were identified influencing the composition of the system, including race, gender, age, lifestyle (e.g., occupation, hygiene, skin product and medication usage, and diet) and environment (e.g., climate, geographical location, pollution, UV, and other radiation) (Wei et al. 2022; Grice and Segre 2011; Harris-Tryon and Grice 2022). Nevertheless, from the perspectives of classical ecology, most of these factors may only indirectly influence, but not drive the establishment and maintenance of the system. The primary selection pressures that form the driving forces for the ecosystem, include resource availability (presence of nutrients), environmental conditions (temperature, geographical access) and biological factors (predators and pathogens) (Williams 1996). In this review, we will sum-up related studies centered on these essential selection pressures, including the presence of different types of nutrients and favored micro-environment for dominant skin commensals, the occupation of the ecological niches through self-adaptation or microbe–microbe interactions, and eventually we will discuss how skin microbes, by their structures or bioactive molecules, reshape host skin phenotypes (Fig. 1).

Skin microbiome, metabolome and skin phenome, from the perspective of skin as an ecosystem. From left to right: (Blue box) Diverse substances, derived from the host (stratum corneum, skin appendages, and plasma), environment (xenobiotics) and microbial metabolism, cover the skin surface, forming the micro-environment for skin microbiota; (Green box) occupation of ecological niches by self-adaptation and microbe–microbe interactions, promoting commensals or inhibiting pathogens; (Yellow box) the skin microbes, by their own structures or bioactive molecules, reshape the host skin phenotypes
Micro-environment of the Human Skin Ecosystem
The host skin offers nutrients and shelters for microbial survival, competition, and cooperation (Roth and James 1988). Nutrient substances may directly affect microbial colonization, growth and metabolism either through nourishing (Brüggemann et al. 2004) or persecuting (Ferrer et al. 2017); on the other hand, these substances may also finetune the local microenvironment, such as pH or moisture state, and thus exert indirect impact on microbial survival. The microbial energy substances are mainly from the host skin and the outside environment. The host skin-derived nutrients consist of lipids embedded in the “brick and mortar” structure (Chen 2018), piles of dead enucleated corneocytes in the stratum corneum (SC) (Abhishek and Palamadai Krishnan 2016), and the secretions from skin appendages [hair follicles (HFs) and glands]. The environment-derived nutrients include personal skincare products, medication, and other environmental xenobiotics. Here, we summarized the metabolites detected on the skin by various metabolome studies (Table1).
It is known that individuals, even the same individual at different life stages, vary markedly in regards to the delicate structure or secretion function of the skin and appendages, which produce metabolites consistently and thus play an essential role in shaping diverse microenvironments with distinct pH, salt, moisture, sebum content, and extent of anaerobiosis (Grice and Segre 2011; Capone et al. 2011; Grice et al. 2009). Factors that influence systemic metabolisms, such as diet and gut microbiota, and hormone levels, can also significantly impact the skin’s local microhabitats (Prescott et al. 2017). Furthermore, one’s exposome, such as environmental pollution, UV levels, occupation environment, drug or skincare habits, is highly individualized (Khmaladze et al. 2020). These together form highly complex physical and chemical landscapes on the skin surface, likely to be the real biological explanation that underlies the substantial inter-individual variability in the skin microbiota. Indeed, our previous study showed two robust “cutotypes” of microbial networks on Chinese facial skin, C-cutotype and M-cutotype, possessed distinct patterns of skin properties (Li et al. 2021). The dominant two species, C. acnes and Moraxella osloensis, exhibited vastly varied nutrient-demand: whereas C. acnes was high nutrient demanding, M. osloensis was a non-fastidious bacterium that was able to grow in a mineral medium supplemented with a single organic carbon source (Juni 1974; Juni and Bøvre 2015). This species was shown to be incapable of utilizing any carbohydrates or possessing any saccharolytic activity, but strictly depend on other carbon sources such as acetic or lactic acid (Baumann et al. 1968; Juni 1974; Juni and Bøvre 2015; Moss et al. 1988).
Occupation of Ecological Niches by Self-adaptation and Microbe–Microbe Interactions
The skin surface formed diverse microhabitats, and many studies favored to divide them into four types (sebaceous, moist, dry, and foot) according to the physical properties of anatomical locations (Oh et al. 2014). Although such water/oil-based classification was not delicate enough, some prominent features for the growth and colonization of the microbiota were well identified. Other metabolites and physical properties were also identified in modulating microbial communities. Furthermore, microbe–microbe interactions are essential for shaping the skin ecosystem. In general, microbes deploy strategies to adapt to the living environment and compete for ecological niches via the following: (1) Self-adaptation to the specific environment conditions: skin microbiota changes their characteristic like metabolism pathways to adapt to the skin microenvironment. For example, Staphylococcus synthesized tensioactive agent to withstand the low pH and high salt content of sweat (Hentati et al. 2021; Scharschmidt and Fischbach 2013); (2) Competition for ecological niches through microbe–microbe interactions, for example, coagulase-negative Staphylococcus (CoNS) species can either directly kill or limit the virulence of Staphylococcus aureus through the secretion of different regulators (Flowers and Grice 2020). Here we will sum-up the findings of this part (Table 2).
Compared to the skin surface, HFs provide a more moisture and acidic environment with ultraviolet light protection, facilitating the colonization of multiple bacteria, fungi, and viruses. The most abundant bacteria in the HFs were P. acnes spp. (Lousada et al. 2021). M. restricta and M. globosa are the dominant fungi (Lousada et al. 2021). Meanwhile, the HF virome comprises dependoviruses, Propionibacterium phage P100D and 101A, papillomaviruses and adeno-associated viruses (Hall et al. 2018). In addition, the mite (Demodex folliculorum) groups are often found in the distal infundibulum, usually with their dorsal body oriented against the hair shaft (Elston and Elston 2014).
From Microbes to Host Skin: How Microbes Reshape the Skin Phenome
Skin microbiota leverage “nutrients” from the host skin and environment and produce a series of bioactive molecules with vital functions (Chen et al. 2018). For example, skin microbiota can convert host proteins into amino acids by their protease (Holland et al. 1979; Byrd et al. 2018), ferment carbohydrates into lactic acids (Ong et al. 2020) or decompose sebum lipids such as triglycerides into free fatty acids (FFAs) (Traisaeng et al. 2019; Belkaid and Segre 2014). In addition, skin microbiota produces AMPs, phenol-soluble modulins (PSMs), and antibiotics (Belkaid and Segre 2014; Gallo and Hooper 2012). These metabolism products may further act on the host or other microbes, exert biological effects and reshape the skin phenome.
The most well-studied functions of skin commensals include the following: (1) pathogen colonization resistance by ecological niche blocking for the invasion of opportunistic or pathogenic microbiota, (2) immune education during early phases, and (3) regulation of immunity and inflammation. Given many comprehensive reviews already on these functions, we will take a particular focus on other functions that were usually missed, including the maintenance of skin physiology, such as pH and SC hydration, UV protection, odor production, and wound healing, which were also important functions in skin homeostasis.
Regulation of Immunity and Inflammation
The microbiota is a rich source of short-chain fatty acids (SCFAs) (Traisaeng et al. 2019). For example, C. acnes fermented carbohydrates into propionic acid (Traisaeng et al. 2019); S. epidermidis was able to ferment glycerol to butyric acid and acetic acid in vitro (Traisaeng et al. 2019; Keshari et al. 2019). SCFAs can regulate several immune cell functions, including the production of cytokines (TNF-α, IL-2, IL-6, and IL-10) (Traisaeng et al. 2019), activate resident skin regulatory T (Treg) cells, mitigate inflammatory skin reactions and thus contribute to the preservation of skin homeostasis in mice and human (Schwarz et al. 2017). Butyric acid significantly attenuated lipopolysaccharide (LPS)-induced nuclear factor-κB (NF-κB) activation and nitric oxide production in murine macrophage cell line (Chakravortty et al. 2000), reduced interferon-gamma (IFNγ)-induced proinflammatory IL-6 and TNF-α production in a macrophage cell line (Park et al. 2007) and mediated short-chain fatty acid receptor 2 (FFAR2) to modulate the production of proinflammatory cytokines induced by ultraviolet B (UVB) in mice (Keshari et al. 2019). Furthermore, the ability of immune cells to migrate to the foci of infection can be regulated by SCFAs (Vinolo et al. 2011). Given the potential anti-inflammatory of SCFAs, they are applied on psoriatic skin in vitro. This study found that decreased expression of G-protein-coupled receptors (GPR) GPR43 and GPR109a in psoriatic skin can be restored and expression of inflammatory factors can be inhibited by topical application of sodium butyrate (Krejner et al. 2018). However, SCFAs are not always anti-inflammatory. C. acnes-derived SCFAs inhibit histone deacetylase (HDAC) activity in skin keratinocytes (KCs) and stimulate inflammation through Toll-like receptor (TLR) signaling (Sanford et al. 2016). SCFAs from C. acnes conferred a robust proinflammatory effect in human sebocytes (Sanford et al. 2019). Expression of a major component of the Corynebacterium accolens cell wall, mycolic acid, promotes inflammation in an IL-23-dependent manner under a high-fat diet condition in mice (Ridaura et al. 2018).
The essential amino acid tryptophan (Trp) can be metabolized by human skin microbiota into 5-hydroxytryptophan (5-HTP), indole-3-aldehyde (IAId) and other metabolites (Yu et al. 2019). IAId was able to suppress thymic stromal lymphopoietin (TSLP) and thereby inhibited calcipotriol (MC903)-induced AD-like dermatitis in mice (Yu et al. 2019). IAId can also activate aryl hydrocarbon receptor (AhR), producing indoleamine 2,3-dioxygenase (IDO) and IL-10 in Langerhans cells (LCs), and thus negatively regulate skin inflammation (Liu et al. 2020).
S. epidermidis and other Gram-positive bacteria release adhesion molecules upon bacteriolysis, such as lipoteichoic acid (LTA) (Ginsburg 2002). LTA from Staphylococcal species suppressed inflammation during tissue injury through a Toll-like receptor 2 (TLR2)-dependent mechanism to prevent excessive damage (Lai et al. 2009). Staphylococcal LTA may also have applications in the treatment of inflammatory disease. For example, in an acne model of C. acnes-induced skin inflammation, staphylococcal LTA application abrogated inflammatory effects via induction of a microRNA, miR-143, destabilizes the TLR2 mRNA and decreases protein production (Xia et al. 2016).
In addition, many commensal species contain virulence strains. One major virulence factor of the microorganism is a secretory lipase that acts on triglycerides to release FFAs (Holland et al. 2010). C. acnes exist both in health and patients, but C. acnes from acne patients harbored unique genomic elements encoding virulence factors, including camp5, gehA, sialidases, neuraminidases, endoglicoceraminidases, lipases, proteases and hemolysins that were rarely present in C. acnes genomes from healthy controls (Brüggemann 2005; Burkhart et al. 1999). Several commensals are opportunistic pathogens that encode virulence factors such as toxins, exoenzymes, and adhesins (Brown et al. 2012). Skin microbiota may directly or indirectly mediate inflammatory responses by releasing various virulence factors under unhealthy conditions. Malassezia spp. can be the causative agents in disease. Many Malassezia spp. secrete extracellular vesicles that signal KCs to secrete proinflammatory cytokines (Vallhov et al. 2020; Watanabe et al. 2001; Zhang et al. 2019). Malassezia spp. metabolize sebum to different fatty acids such as phosphatidylcholine (PC) and phosphatidylserine (PS), which then act as irritants, causing flaking and irritation under dandruff, a frequent scalp issue and seborrheic dermatitis conditions (Celis Ramírez et al. 2020; DeAngelis et al. 2005; Han et al. 2019; Johansson et al. 2018).
Pathogen Colonization Resistance
Commensals compete for niches through microbe–microbe interactions, as mentioned above (Table 2). Direct induction of AMPs or cytokine expression in KCs is one of the main strategies used by skin commensals, such as Propionibacterium and S. epidermidis, in defending against pathogen invasion and shaping the skin microbiota community (Midorikawa et al. 2003; Wanke et al. 2011). In addition, commensals function as endogenous cofactors of the skin immune system to promote skin local immune response. Skin harbor considerable commensal-specific T-cell, e.g., Staphylococcus epidermidis-specific IL-17A+ CD8+ T cells (Naik et al. 2015). The activation of these cells can promote AMP production by keratinocytes, thereby promoting heterologous protection against pathogens infections (Braff et al. 2005). Staphylococcus epidermidis can also induce KC to express IL-1α, thus promoting skin αβ T cells to produce IL-17A and IFNγ in mice (Naik et al. 2012). IL-17A induces chemokines that recruit neutrophils and AMP production, thus protecting the host from pathogen infection. In adults, cutaneous mucosal-associated invariant T cells (MAIT cells) are a dominant population of IL-17A-producing lymphocytes (Constantinides et al. 2019). MAIT cells are absent in germ-free (GF) mice, and their development are controlled by microbial metabolites such as vitamin B2 (Treiner et al. 2003; Koay et al. 2016; Legoux et al. 2019). MAIT cells can respond to skin commensals or commensal-derived metabolites in an IL-1-, IL-18-, and antigen-dependent manner (Constantinides et al. 2019), thus enhancing inhibition of pathogen invasion.
Immune Education
The commensals play an essential role in regulating the development, proliferation, maturation and activation of immune cells of innate immunity. A previous study found that GF mice contain mast cells (MCs) that are largely undifferentiated and express abnormally low amounts of stem cell factor (SCF). Commensal bacteria induce KC-produced SCF, promote skin MCs mature. The migration of MCs in the skin is fully dependent on high levels of SCF, as produced by KCs (Wang et al. 2017b). In addition, γδT cells, which play an essential role in recognizing lipids, one of the microbial metabolites (Belkaid and Tamoutounour 2016), significantly reduced IL-17 secretion capacity in GF mice (Naik et al. 2012). Varying from the immune responses to invasive pathogens, adaptive immune responses respond to commensals under noninflammatory conditions, which help build immune homeostasis (Naik et al. 2015).
The skin contains one of the highest frequencies of FOXP3+ Treg cells within the body in mice (Suffia et al. 2006). In the skin of both mice and humans, Tregs reside in the dermis, and a large fraction of these cells can be found in close proximity to HFs, which serve as a natural habitat for skin-resident microorganisms (Ali et al. 2017; Sanchez Rodriguez et al. 2014). Tregs are essential in establishing and regulating immune tolerance to commensal microbes during a defined period of neonatal life in mice (Scharschmidt et al. 2015). S. epidermidis colonization on the skin surface two weeks after birth induces Treg cells’ tolerance to S. epidermidis in adult mice (Scharschmidt et al. 2015). Furthermore, it promotes the accumulation and migration of Treg cells into the skin (Scharschmidt et al. 2017). Further study found that Treg cell migration in Neonatal Skin is influenced by hair follicle development and microbes colonized in the hair follicle. In turn, colonization of microbes in HFs during the early stage is resisted and regulated by Treg cells (Scharschmidt et al. 2017). These results suggest a dynamic balance between microbe and host immune system.
Maintain pH and SC Hydration
Skin microbiota metabolizes dead corneocytes, sweat and sebum components, and other wastes (Pistone et al. 2021) and converts them into amino acids, such as glutamate and aspartate, proteins and various FFAs (Pistone et al. 2021; Timm et al. 2020). They also secrete lactic acid (Ong et al. 2020), a series of SCFAs (Christensen and Brüggemann 2014) and other organic acids (Garrote et al. 2000; Wang et al. 2017a; Bengoa et al. 2019). These acidic metabolites can regulate skin surface pH and SC hydration level (Watabe et al. 2013; McGrath 2008; Caspers et al. 2001; Cui et al. 2016; Pappas 2009).
The skin surface pH is slightly acidic, ranging from 4.5 to 5.5 in human (Braun-Falco and Korting 1986). The pH of the SC is crucial for many vital epidermal functions, including permeability barrier homeostasis, desquamation of corneocytes, initiation of inflammation, processing of secreted lamellar body (LB) polar lipids and antimicrobial defense (Lee and Lee 2014). In addition, variation in pH also affects the SC thickness and pigmentation (Sandby-Møller et al. 2003). These results indicate that many skin traits may intertwine, such as pH, trans-epidermal water loss (TEWL), skin thickness, SC hydration and pigmentation, and thereby may be modulated by skin microbiota and their metabolites.
Our previous study also revealed that cutotypes of microbial networks on Chinese facial skin possess distinct skin traits: C-cutotype skin is more hydrated and more oily, and the levels of skin surface sebum and its microbial metabolite porphyrin are increased; In contrast, M-cutotype skin is dryer and often occurs in the elder (Li et al. 2021). A study on the skin microbiome of Koreans found that Lawsonella had a negative correlation with skin moisture and brown spots; Staphylococcus and Corynebacterium both had negative correlations with the number of UV spots and positive correlations with TEWL; Staphylococcus aureus had a negative correlation with skin moisture parameters (Kim et al. 2021a). Moreover, two studies found a linkage between the skin microbiome and skin metabolites (Howard et al. 2022; Roux et al. 2022). A recent study demonstrated that S. epidermidis can significantly increase skin ceramide levels and thereby prevent water loss of damaged skin dependent on its sphingomyelinase in mice (Zheng et al. 2022).
Skin aging is a dynamic process with a series of changes in the skin phenome (Farage et al. 2008; Pochi et al. 1979; Cotterill et al. 1972; Howard et al. 2022) and skin metabolism, e.g., altered levels of natural moisturizing factors (NMFs), AMPs, vitamins and coenzyme Q10, and many other metabolites (Howard et al. 2022; MacLaughlin and Holick 1985; Kuehne et al. 2017). These changes may underlie the alterations in the microbiome. For example, age-related decrease in sebocyte area is positively correlated with Cutibacterium and negatively correlated with Streptococcus, Acinetobacter, Corynebacterium and Methylobacterium‒Methylorubrum abundance (Howard et al. 2022). Furthermore, anti-aging skincare products were reported able to persist on the skin for weeks and provide long-term contributions to the chemical environment (Bouslimani et al. 2019), thus shaping the specific skin microbial communities (Bouslimani et al. 2015). For example, lipid components of moisturizers could provide nutrients and promote the growth of lipophilic bacteria such as Staphylococcus and Propionibacterium (Bouslimani et al. 2015; Unno et al. 2017; Holland et al. 2010). More details regarding cosmetics can be found in Table 1.
UV Protection
Some skin commensals can protect skin from UV damage by secreting different metabolites (Souak et al. 2021). For example, S. epidermidis can produce 6-HAP to suppress UV-induced tumor in mice (Nakatsuji et al. 2018). Skin microflora produces cis-urocanic acid from l-histidine, affects UV-induced immune suppression and suppresses melanoma growth (Hug et al. 1999; Laihia et al. 2010). Some Streptomyces-derived compounds, such as amides exhibited UV-absorbing, antioxidant, and anti-inflammatory properties (Sánchez-Suárez et al. 2020). Propionic acid produced by Cutibacterium acnes fermentation ameliorates UVB-induced melanin synthesis (Kao et al. 2021). Cyanobacteria develop a diversity of defense mechanisms, including the biosynthesis of UV-absorbing/screening compounds, such as mycosporine-like amino acids (MAAs), and enzymes, including superoxide dismutases (SOD), which counteract oxidative stress (Souak et al. 2021).
Ultraviolet radiation (UV-R) is well known to inhibit the cellular growth of Malassezia furfur (Wikler et al. 1990). On the other hand, Malassezia furfur can produce pityriacitrin, a UV-filtering compound believed to be protective (Machowinski et al. 2006). It is hypothesized that this fungus developed the UV-filter compound to reduce UV damage and compete for survival over other commensals (Machowinski et al. 2006). However, they did not find any adverse effect of pityriacitrin on commensals such as S. aureus, S. epidermidis, or Candida albicans (Machowinski et al. 2006).
Odor Production
The metabolic activities of some skin microbes produce special odors. For example, human body odor is believed to result from bacterial growth and decomposition of secretions from specialized glands in the axillary region (Lam et al. 2018; Decréau et al. 2003; Natsch et al. 2003). Microbes are present in specific scent glands or tissue in mammals and modulate specific odors (Ezenwa et al. 2012). Skin microbes metabolize host sweat and produce volatile metabolites, enhancing the attractiveness of human sweat for the malaria mosquito (Brouwer 1960; Takken and Kline 1989). A recent study specified acetophenone, a volatile from the skin microbiota, promoted mosquito attractiveness in flavivirus-infected hosts (Zhang et al. 2022).
Skin commensal Moraxella osloensis (Li et al. 2021), a species highly tolerant to desiccation and UV irradiation, existed in various living environments, particularly in the laundry. This species has the potential to generate 4-methyl-3-hexenoic acid (4M3H), which is often described as a “wet-and-dirty-dustcloth-like malodor” or an “acidic or sweaty odor” (Kubota et al. 2012). In addition to bacteria, fungi are important sources of many volatile organic compounds (VOCs), including alcohols, aldehydes, esters, FAs, and terpenes (Belinato et al. 2019). In malignant fungating wounds (MFWs), metabolites such as dimethyl trisulfide (DMTS), four fatty acid volatiles (acetic acid, isobutyric acid, butyric acid, and isovaleric acid) and putrescine are linked with components of malignant fungating wound odor (Vardhan et al. 2019).
Wound Healing
Wound healing is a complex but highly regulated process critical for skin barrier function (Han and Ceilley 2017). The presence and abundance of microbes in skin wounds depend on wound type (chronic/acute wound) (Johnson et al. 2018) and shifts over time (Loesche et al. 2017). Studies demonstrated that skin microbiota was also involved in wound healing in multifaceted ways. S. epidermidis promotes rapid KC progression via upregulation of TLR and downstream modulation of TNF-α in skin CD8+ T cells (Linehan et al. 2018; Naik et al. 2015). A study with a wound-induced hair follicle neogenesis (WIHN) mouse model revealed that skin microbiota promoted skin regeneration via IL-1β and KC-dependent IL-1R-MyD88 signaling (Wang et al. 2021). Metabolites from microbiota promote wound healing, e.g., lipoteichoic acid from S. epidermidis can decrease inflammation via TLR2 signaling (Lai et al. 2009). On the other hand, some potential pathogens do not promote cutaneous wound healing. For example, S. aureus (Kirker et al. 2009; den Reijer et al. 2016), Acinetobacter. baumanni and A. junii (de Breij et al. 2012) form biofilms on the SC and have a detrimental effect on human dermal fibroblast migration and ultimately result in cellular apoptosis (Kirker et al. 2012). Microbial stability was believed to be essential for skin health; however, temporal stability in the chronic wound is associated with poor healing as instability in the microbiome reflects effective control of wound bacteria, which prevents any community structure from stabilizing (Loesche et al. 2017).
Conclusion
The present review centers on the current knowledge on skin microbiome from a perspective of skin as an ecosystem and tries to explore the fundamental driving force for the establishment and the balance of the highly personalized microbial feature. We believe that microenvironments that define the physical (e.g., pH, oxygen) and chemical (carbon sources and metabolites) conditions drive the microbiome composition. In turn, these microbes may reshape this environment via microbe–microbe or microbe–host interactions. Skin surface metabolome may be a critical approach to address causative correlations between the skin microbiome and skin phenome; therefore, future skin microbiome research should leverage those multi-omics to reveal these strong correlations and then validate them with the principle of Koch’s postulates. Furthermore, considering the higher complexity of the system due to the host genome and exposome, the longitudinal time-series study should be taken more into consideration for the control of these variables and for addressing the direction of those networks. Based on solid causative correlations, we can develop accurate interventions targeting specific skin microbe(s) and eventually reshape the skin conditions.
Of note, recent studies revealed that microbiota at strain level varies in the local microenvironment (Conwill et al. 2022), suggesting studies on higher resolution should be emphasized, which means deeper sequencing until strain level and more refined sampling sites up to single pore level. However, the greatest challenge for these designs is biomass, including metabolites and metagenomic biomass. This strongly relies on the technology development and iterative update of detection instruments to improve the sensitivity.
The significance of the human skin microbiome is increasingly appreciated. The approach from metagenomic sequencing (profiling) was gradually shifted to isolation/culturomics and function validation (mechanisms). However, some significant issues still exist, such as the lack of ideal ex-vivo skin models (e.g., reconstructed human epidermis (RHEs) and skin explants) that can reliably simulate the complexity of the host–microbe interactions (Harris-Tryon and Grice 2022; Larson et al. 2021). Some recent studies performed the function experiments with three-dimensional (3D) human skin equivalent. For example, a study using 3D skin tissue cultures revealed that a model microbiome or a mixed community of skin microbiome representatives led to pronounced changes in epidermal thickness, epidermal cell proliferation, and filaggrin production (Loomis et al. 2021). Another study investigated the interaction between the skin microbiota and environmental pollutant benzo[a]pyrene (B[a]P), with a microbially competent 3D skin model and demonstrated that commensal metabolism of xenobiotics can influence host toxicity (Lemoine et al. 2021). However, the limitations of these ex-vivo skin models are apparent, i.e., the lack of the histological/physiological/immunological complexity of RHEs, the paucity of inter-donor variability of skin explants, as well as short lifespan and the relatively high costs (Larson et al. 2021). Nevertheless, this is a matter of time to address these issues and push forward the skin microbiota targeted new intervention based on solid experimental evidence.
Data Availability
Not applicable.
Code Availability
Not applicable.
References
Abhishek S, Palamadai Krishnan S (2016) Epidermal differentiation complex: a review on its epigenetic regulation and potential drug targets. Cell J 18:1–6. https://doi.org/10.22074/cellj.2016.3980
Agrawal K, Waller JD, Pedersen TL et al (2018) Effects of stimulation technique, anatomical region, and time on human sweat lipid mediator profiles. Prostaglandins Other Lipid Mediat 134:84–92. https://doi.org/10.1016/j.prostaglandins.2017.09.007
Al-Fatimi M, Wurster M, Lindequist U (2016) Chemical composition, antimicrobial and antioxidant activities of the volatile oil of Ganoderma pfeifferi Bres. Medicines. https://doi.org/10.3390/medicines3020010
Ali N, Zirak B, Rodriguez RS et al (2017) Regulatory T cells in skin facilitate epithelial stem cell differentiation. Cell 169:1119–29.e11. https://doi.org/10.1016/j.cell.2017.05.002
Allhorn M, Arve S, Brüggemann H et al (2016) A novel enzyme with antioxidant capacity produced by the ubiquitous skin colonizer Propionibacterium acnes. Sci Rep 6:36412. https://doi.org/10.1038/srep36412
Angiolella L, Rojas F, Mussin J et al (2020) Biofilm formation, adherence, and hydrophobicity of M. sympodialis, M. globosa, and M. slooffiae from clinical isolates and normal skin virulence factors of M. sympodialis, M. globosa and M. slooffiae. Med Mycol 58:1162–1168. https://doi.org/10.1093/mmy/myaa017
Ansari MN, Nicolaides N, Fu HC (1970) Fatty acid composition of the living layer and stratum corneum lipids of human sole skin epidermis. Lipids 5:838–845. https://doi.org/10.1007/bf02531977
Arezki NR, Williams AC, Cobb AJ et al (2017) Design, synthesis and characterization of linear unnatural amino acids for skin moisturization. Int J Cosmet Sci 39:72–82. https://doi.org/10.1111/ics.12351
Autiero M, Abrescia P, Guardiola J (1991) Interaction of seminal plasma proteins with cell surface antigens: presence of a CD4-binding glycoprotein in human seminal plasma. Exp Cell Res 197:268–271. https://doi.org/10.1016/0014-4827(91)90432-t
Badiu DL, Luque R, Dumitrescu E et al (2010) Amino acids from Mytilus galloprovincialis (L.) and Rapana venosa molluscs accelerate skin wounds healing via enhancement of dermal and epidermal neoformation. Protein J 29:81–92. https://doi.org/10.1007/s10930-009-9225-9
Baechle D, Flad T, Cansier A et al (2006) Cathepsin D is present in human eccrine sweat and involved in the postsecretory processing of the antimicrobial peptide DCD-1L. J Biol Chem 281:5406–5415. https://doi.org/10.1074/jbc.M504670200
Bajo-Grañeras R, Sanchez D, Gutierrez G et al (2011) Apolipoprotein D alters the early transcriptional response to oxidative stress in the adult cerebellum. J Neurochem 117:949–960. https://doi.org/10.1111/j.1471-4159.2011.07266.x
Baker LB (2019) Physiology of sweat gland function: the roles of sweating and sweat composition in human health. Temperature 6:211–259. https://doi.org/10.1080/23328940.2019.1632145
Barresi C, Stremnitzer C, Mlitz V et al (2011) Increased sensitivity of histidinemic mice to UVB radiation suggests a crucial role of endogenous urocanic acid in photoprotection. J Investig Dermatol 131:188–194. https://doi.org/10.1038/jid.2010.231
Bastos MC, Ceotto H, Coelho ML et al (2009) Staphylococcal antimicrobial peptides: relevant properties and potential biotechnological applications. Curr Pharm Biotechnol 10:38–61. https://doi.org/10.2174/138920109787048580
Baumann P, Doudoroff M, Stanier RY (1968) Study of the Moraxella group. I. Genus Moraxella and the Neisseria catarrhalis group. J Bacteriol 95:58–73. https://doi.org/10.1128/jb.95.1.58-73.1968
Belinato JR, Silva E, De Souza DS et al (2019) Rapid discrimination of fungal strains isolated from human skin based on microbial volatile organic profiles. J Chromatogr B Anal Technol Biomed Life Sci 1110–1111:9–14. https://doi.org/10.1016/j.jchromb.2019.02.007
Belkaid Y, Segre JA (2014) Dialogue between skin microbiota and immunity. Science 346:954–959. https://doi.org/10.1126/science.1260144
Belkaid Y, Tamoutounour S (2016) The influence of skin microorganisms on cutaneous immunity. Nat Rev Immunol 16:353–366. https://doi.org/10.1038/nri.2016.48
Bengoa AA, Iraporda C, Garrote GL et al (2019) Kefir micro-organisms: their role in grain assembly and health properties of fermented milk. J Appl Microbiol 126:686–700. https://doi.org/10.1111/jam.14107
Bhattacharyya AK, Connor WE, Spector AA (1972) Excretion of sterols from the skin of normal and hypercholesterolemic humans. implications for sterol balance studies. J Clin Investig 51:2060–2070. https://doi.org/10.1172/jci107012
Bocheva G, Slominski RM, Slominski AT (2021) The impact of vitamin D on skin aging. Int J Mol Sci. https://doi.org/10.3390/ijms22169097
Bomar L, Brugger SD, Yost BH et al (2016) Corynebacterium accolens releases antipneumococcal free fatty acids from human nostril and skin surface triacylglycerols. Mbio 7:e01725-15. https://doi.org/10.1128/mBio.01725-15
Bosch TC, Mcfall-Ngai MJ (2011) Metaorganisms as the new frontier. Zoology 114:185–190. https://doi.org/10.1016/j.zool.2011.04.001
Bouslimani A, Porto C, Rath CM et al (2015) Molecular cartography of the human skin surface in 3D. Proc Natl Acad Sci USA 112:E2120–E2129. https://doi.org/10.1073/pnas.1424409112
Bouslimani A, Da Silva R, Kosciolek T et al (2019) The impact of skin care products on skin chemistry and microbiome dynamics. BMC Biol 17:47. https://doi.org/10.1186/s12915-019-0660-6
Boxberger M, Cenizo V, Cassir N et al (2021) Challenges in exploring and manipulating the human skin microbiome. Microbiome 9:125. https://doi.org/10.1186/s40168-021-01062-5
Braff MH, Zaiou M, Fierer J et al (2005) Keratinocyte production of cathelicidin provides direct activity against bacterial skin pathogens. Infect Immun 73:6771–6781. https://doi.org/10.1128/iai.73.10.6771-6781.2005
Braun-Falco O, Korting HC (1986) Normal pH value of human skin. Hautarzt 37:126–129
Brouwer R (1960) The attraction of carbon dioxide excreted by the skin of the arm for malaria mosquitoes. Trop Geogr Med 12:62–66
Brown SK, Shalita AR (1998) Acne vulgaris. Lancet 351:1871–1876. https://doi.org/10.1016/s0140-6736(98)01046-0
Brown SP, Cornforth DM, Mideo N (2012) Evolution of virulence in opportunistic pathogens: generalism, plasticity, and control. Trends Microbiol 20:336–342. https://doi.org/10.1016/j.tim.2012.04.005
Brüggemann H (2005) Insights in the pathogenic potential of Propionibacterium acnes from its complete genome. Semin Cutan Med Surg 24:67–72. https://doi.org/10.1016/j.sder.2005.03.001
Brüggemann H, Henne A, Hoster F et al (2004) The complete genome sequence of Propionibacterium acnes, a commensal of human skin. Science 305:671–673. https://doi.org/10.1126/science.1100330
Büchau AS, Gallo RL (2007) Innate immunity and antimicrobial defense systems in psoriasis. Clin Dermatol 25:616–624. https://doi.org/10.1016/j.clindermatol.2007.08.016
Burke RC, Lee TH, Buettner-Janusch V (1966) Free amino acids and water soluble peptides in stratum corneum and skin surface film in human beings. Yale J Biol Med 38:355–373
Burkhart CG, Burkhart CN, Lehmann PF (1999) Acne: a review of immunologic and microbiologic factors. Postgrad Med J 75:328–331. https://doi.org/10.1136/pgmj.75.884.328
Burkovski A (2018) The role of corynomycolic acids in Corynebacterium-host interaction. Antonie Van Leeuwenhoek 111:717–725. https://doi.org/10.1007/s10482-018-1036-6
Byrd AL, Belkaid Y, Segre JA (2018) The human skin microbiome. Nat Rev Microbiol 16:143–155. https://doi.org/10.1038/nrmicro.2017.157
Capone KA, Dowd SE, Stamatas GN et al (2011) Diversity of the human skin microbiome early in life. J Investig Dermatol 131:2026–2032. https://doi.org/10.1038/jid.2011.168
Caspers PJ, Lucassen GW, Carter EA et al (2001) In vivo confocal Raman microspectroscopy of the skin: noninvasive determination of molecular concentration profiles. J Investig Dermatol 116:434–442. https://doi.org/10.1046/j.1523-1747.2001.01258.x
Celis Ramírez AM, Amézquita A, Cardona Jaramillo JEC et al (2020) Analysis of malassezia lipidome disclosed differences among the species and reveals presence of unusual yeast lipids. Front Cell Infect Microbiol 10:338. https://doi.org/10.3389/fcimb.2020.00338
Chakravortty D, Koide N, Kato Y et al (2000) The inhibitory action of butyrate on lipopolysaccharide-induced nitric oxide production in RAW 264.7 murine macrophage cells. J Endotoxin Res 6:243–247. https://doi.org/10.1177/09680519000060030501
Chamberlain NR, Brueggemann SA (1997) Characterisation and expression of fatty acid modifying enzyme produced by Staphylococcus epidermidis. J Med Microbiol 46:693–697. https://doi.org/10.1099/00222615-46-8-693
Chen X (2018) Current and future technological advances in transdermal gene delivery. Adv Drug Deliv Rev 127:85–105. https://doi.org/10.1016/j.addr.2017.12.014
Chen YE, Fischbach MA, Belkaid Y (2018) Skin microbiota–host interactions. Nature 553:427–436. https://doi.org/10.1038/nature25177
Chen D, He J, Li J et al (2021) Microbiome and metabolome analyses reveal novel interplay between the skin microbiota and plasma metabolites in psoriasis. Front Microbiol 12:643449. https://doi.org/10.3389/fmicb.2021.643449
Choi H, Kim DJ, Nam S et al (2018) Substance P restores normal skin architecture and reduces epidermal infiltration of sensory nerve fiber in TNCB-induced atopic dermatitis-like lesions in NC/Nga mice. J Dermatol Sci 89:248–257. https://doi.org/10.1016/j.jdermsci.2017.11.013
Christensen GJM, Brüggemann H (2014) Bacterial skin commensals and their role as host guardians. Benef Microbes 5:201–215. https://doi.org/10.3920/bm2012.0062
Claesen J, Spagnolo JB, Ramos SF et al (2020) A Cutibacterium acnes antibiotic modulates human skin microbiota composition in hair follicles. Sci Transl Med. https://doi.org/10.1126/scitranslmed.aay5445
Clayton RW, Göbel K, Niessen CM et al (2019) Homeostasis of the sebaceous gland and mechanisms of acne pathogenesis. Br J Dermatol 181:677–690. https://doi.org/10.1111/bjd.17981
Cogen AL, Yamasaki K, Sanchez KM et al (2010) Selective antimicrobial action is provided by phenol-soluble modulins derived from Staphylococcus epidermidis, a normal resident of the skin. J Investig Dermatol 130:192–200. https://doi.org/10.1038/jid.2009.243
Cohn JR, Emmett EA (1978) The excretion of trace metals in human sweat. Ann Clin Lab Sci 8:270–275
Consolazio CF, Matoush LO, Nelson RA et al (1962) The dermal excretion of minerals and its possible relation to mineral balance and requirements (Sodium, potassium, iron, magnesium and phosphorus). Rep US Army Med Res Nutr Lab Denver
Consolazio CF, Matoush LO, Nelson RA et al (1966) Comparisons of nitrogen, calcium and iodine excretion in arm and total body sweat. Am J Clin Nutr 18:443–448. https://doi.org/10.1093/ajcn/18.6.443
Constante M, Fragoso G, Lupien-Meilleur J et al (2017) Iron supplements modulate colon microbiota composition and potentiate the protective effects of probiotics in dextran sodium sulfate-induced colitis. Inflamm Bowel Dis 23:753–766. https://doi.org/10.1097/mib.0000000000001089
Constantinides MG, Link VM, Tamoutounour S et al (2019) MAIT cells are imprinted by the microbiota in early life and promote tissue repair. Science. https://doi.org/10.1126/science.aax6624
Conwill A, Kuan AC, Damerla R et al (2022) Anatomy promotes neutral coexistence of strains in the human skin microbiome. Cell Host Microbe 30:171–82.e7. https://doi.org/10.1016/j.chom.2021.12.007
Cornbleet T, Klein R, Pace E (1936) Vitamin C content of sweat. Arch Derm Syphilol 34:253–254
Cotterill JA, Cunliffe WJ, Williamson B et al (1972) Age and sex variation in skin surface lipid composition and sebum excretion rate. Br J Dermatol 87:333–340. https://doi.org/10.1111/j.1365-2133.1972.tb07419.x
Craig SS, Craig SA, Ganio MS et al (2010) The betaine content of sweat from adolescent females. J Int Soc Sports Nutr 7:3. https://doi.org/10.1186/1550-2783-7-3
Csősz É, Emri G, Kalló G et al (2015) Highly abundant defense proteins in human sweat as revealed by targeted proteomics and label-free quantification mass spectrometry. J Eur Acad Dermatol Venereol 29:2024–2031. https://doi.org/10.1111/jdv.13221
Cui L, Jia Y, Cheng ZW et al (2016) Advancements in the maintenance of skin barrier/skin lipid composition and the involvement of metabolic enzymes. J Cosmet Dermatol 15:549–558. https://doi.org/10.1111/jocd.12245
Dai X, Okazaki H, Hanakawa Y et al (2013) Eccrine sweat contains IL-1alpha, IL-1beta and IL-31 and activates epidermal keratinocytes as a danger signal. PLoS ONE 8:e67666. https://doi.org/10.1371/journal.pone.0067666
Dam B (1978) Vitamins and sport. Br J Sports Med 12:74–79
De Breij A, Haisma EM, Rietveld M et al (2012) Three-dimensional human skin equivalent as a tool to study Acinetobacter baumannii colonization. Antimicrob Agents Chemother 56:2459–2464. https://doi.org/10.1128/aac.05975-11
Deangelis YM, Gemmer CM, Kaczvinsky JR et al (2005) Three etiologic facets of dandruff and seborrheic dermatitis: Malassezia fungi, sebaceous lipids, and individual sensitivity. J Investig Dermatol Symp Proc 10:295–297. https://doi.org/10.1111/j.1087-0024.2005.10119.x
Decréau RA, Marson CM, Smith KE et al (2003) Production of malodorous steroids from androsta-5,16-dienes and androsta-4,16-dienes by Corynebacteria and other human axillary bacteria. J Steroid Biochem Mol Biol 87:327–336. https://doi.org/10.1016/j.jsbmb.2003.09.005
Den Reijer PM, Haisma EM, Lemmens-Den Toom NA et al (2016) Detection of alpha-toxin and other virulence factors in biofilms of Staphylococcus aureus on polystyrene and a human epidermal model. PLoS ONE 11:e0145722. https://doi.org/10.1371/journal.pone.0145722
Diep BA, Gill SR, Chang RF et al (2006) Complete genome sequence of USA300, an epidemic clone of community-acquired meticillin-resistant Staphylococcus aureus. Lancet 367:731–739. https://doi.org/10.1016/s0140-6736(06)68231-7
Drake DR, Brogden KA, Dawson DV et al (2008) Thematic review series: skin lipids. Antimicrobial lipids at the skin surface. J Lipid Res 49:4–11. https://doi.org/10.1194/jlr.R700016-JLR200
Du B, Bian Z, Xu B (2014) Skin health promotion effects of natural beta-glucan derived from cereals and microorganisms: a review. Phytother Res 28:159–166. https://doi.org/10.1002/ptr.4963
Elpa DP, Chiu HY, Wu SP et al (2021) Skin metabolomics. Trends Endocrinol Metab 32:66–75. https://doi.org/10.1016/j.tem.2020.11.009
Elston CA, Elston DM (2014) Demodex mites. Clin Dermatol 32:739–743. https://doi.org/10.1016/j.clindermatol.2014.02.012
Ezenwa VO, Gerardo NM, Inouye DW et al (2012) Microbiology. Anim Behav Microbiome Sci 338:198–199. https://doi.org/10.1126/science.1227412
Farage MA, Miller KW, Elsner P et al (2008) Functional and physiological characteristics of the aging skin. Aging Clin Exp Res 20:195–200. https://doi.org/10.1007/bf03324769
Feingold KR (2009) The outer frontier: the importance of lipid metabolism in the skin. J Lipid Res 50(Suppl):S417–S422. https://doi.org/10.1194/jlr.R800039-JLR200
Ferrer M, Méndez-García C, Rojo D et al (2017) Antibiotic use and microbiome function. Biochem Pharmacol 134:114–126. https://doi.org/10.1016/j.bcp.2016.09.007
Findley K, Oh J, Yang J et al (2013) Topographic diversity of fungal and bacterial communities in human skin. Nature 498:367–370. https://doi.org/10.1038/nature12171
Fischer CL, Drake DR, Dawson DV et al (2012) Antibacterial activity of sphingoid bases and fatty acids against gram-positive and gram-negative bacteria. Antimicrob Agents Chemother 56:1157–1161. https://doi.org/10.1128/aac.05151-11
Flowers L, Grice EA (2020) The skin microbiota: balancing risk and reward. Cell Host Microbe 28:190–200. https://doi.org/10.1016/j.chom.2020.06.017
Fluhr JW, Darlenski R, Surber C (2008) Glycerol and the skin: holistic approach to its origin and functions. Br J Dermatol 159:23–34. https://doi.org/10.1111/j.1365-2133.2008.08643.x
Gallo RL, Hooper LV (2012) Epithelial antimicrobial defence of the skin and intestine. Nat Rev Immunol 12:503–516. https://doi.org/10.1038/nri3228
Gannesen AV, Borrel V, Lefeuvre L et al (2019) Effect of two cosmetic compounds on the growth, biofilm formation activity, and surface properties of acneic strains of Cutibacterium acnes and Staphylococcus aureus. Microbiologyopen 8:e00659. https://doi.org/10.1002/mbo3.659
Gariboldi S, Palazzo M, Zanobbio L et al (2008) Low molecular weight hyaluronic acid increases the self-defense of skin epithelium by induction of beta-defensin 2 via TLR2 and TLR4. J Immunol 181:2103–2110. https://doi.org/10.4049/jimmunol.181.3.2103
Garrote GL, Abraham AG, De Antoni GL (2000) Inhibitory power of kefir: the role of organic acids. J Food Prot 63:364–369. https://doi.org/10.4315/0362-028x-63.3.364
Gehring W (2004) Nicotinic acid/niacinamide and the skin. J Cosmet Dermatol 3:88–93. https://doi.org/10.1111/j.1473-2130.2004.00115.x
Ginsburg I (2002) Role of lipoteichoic acid in infection and inflammation. Lancet Infect Dis 2:171–179. https://doi.org/10.1016/s1473-3099(02)00226-8
Gläser R, Harder J, Lange H et al (2005) Antimicrobial psoriasin (S100A7) protects human skin from Escherichia coli infection. Nat Immunol 6:57–64. https://doi.org/10.1038/ni1142
Greene RS, Downing DT, Pochi PE et al (1970) Anatomical variation in the amount and composition of human skin surface lipid. J Investig Dermatol 54:240–247. https://doi.org/10.1111/1523-1747.ep12280318
Gribbon EM, Cunliffe WJ, Holland KT (1993) Interaction of Propionibacterium acnes with skin lipids in vitro. J Gen Microbiol 139:1745–1751. https://doi.org/10.1099/00221287-139-8-1745
Grice EA, Dawson TLJ (2017) Host–microbe interactions: Malassezia and human skin. Curr Opin Microbiol 40:81–87. https://doi.org/10.1016/j.mib.2017.10.024
Grice EA, Segre JA (2011) The skin microbiome. Nat Rev Microbiol 9:244–253. https://doi.org/10.1038/nrmicro2537
Grice EA, Kong HH, Conlan S et al (2009) Topographical and temporal diversity of the human skin microbiome. Science 324:1190–1192. https://doi.org/10.1126/science.1171700
Gum ET, Swanson RA, Alano C et al (2004) Human serum albumin and its N-terminal tetrapeptide (DAHK) block oxidant-induced neuronal death. Stroke 35:590–595. https://doi.org/10.1161/01.Str.0000110790.05859.Da
Hall JB, Cong Z, Imamura-Kawasawa Y et al (2018) Isolation and identification of the follicular microbiome: implications for acne research. J Investig Dermatol 138:2033–2040. https://doi.org/10.1016/j.jid.2018.02.038
Han G, Ceilley R (2017) Chronic wound healing: a review of current management and treatments. Adv Ther 34:599–610. https://doi.org/10.1007/s12325-017-0478-y
Han Y, Zhang YJ, Wang HX et al (2019) Malassezia furfur promoting growth of Staphylococcus epidermidis by increasing pH when cultured in a lipid-free environment. Chin Med J 132:873–876. https://doi.org/10.1097/cm9.0000000000000152
Hardy BL, Dickey SW, Plaut RD et al (2019) Corynebacterium pseudodiphtheriticum exploits Staphylococcus aureus virulence components in a novel polymicrobial defense strategy. Mbio. https://doi.org/10.1128/mBio.02491-18
Harris-Tryon TA, Grice EA (2022) Microbiota and maintenance of skin barrier function. Science 376:940–945. https://doi.org/10.1126/science.abo0693
Harshman SW, Pitsch RL, Smith ZK et al (2018) The proteomic and metabolomic characterization of exercise-induced sweat for human performance monitoring: a pilot investigation. PLoS ONE 13:e0203133. https://doi.org/10.1371/journal.pone.0203133
Hassan MI, Waheed A, Yadav S et al (2009) Prolactin inducible protein in cancer, fertility and immunoregulation: structure, function and its clinical implications. Cell Mol Life Sci 66:447–459. https://doi.org/10.1007/s00018-008-8463-x
Hentati D, Cheffi M, Hadrich F et al (2021) Investigation of halotolerant marine Staphylococcus sp. CO100, as a promising hydrocarbon-degrading and biosurfactant-producing bacterium, under saline conditions. J Environ Manag 277:111480. https://doi.org/10.1016/j.jenvman.2020.111480
Hoiby N, Pers C, Johansen HK et al (2000) Excretion of beta-lactam antibiotics in sweat—a neglected mechanism for development of antibiotic resistance? Antimicrob Agents Chemother 44:2855–2857. https://doi.org/10.1128/aac.44.10.2855-2857.2000
Holland KT, Greenman J, Cunliffe WJ (1979) Growth of cutaneous propionibacteria on synthetic medium; growth yields and exoenzyme production. J Appl Bacteriol 47:383–394. https://doi.org/10.1111/j.1365-2672.1979.tb01198.x
Holland KT, Aldana O, Bojar RA et al (1998) Propionibacterium acnes and acne. Dermatology 196:67–68. https://doi.org/10.1159/000017870
Holland C, Mak TN, Zimny-Arndt U et al (2010) Proteomic identification of secreted proteins of Propionibacterium acnes. BMC Microbiol 10:230. https://doi.org/10.1186/1471-2180-10-230
Houpt ER (2005) Microbial inhabitants of humans: their ecology and role in health and disease by Michael Wilson Cambridge, U.K.: Cambridge University Press, 2005. 476 pp., illustrated. $65.00 (cloth). Clin Infect Dis 41:768–868. https://doi.org/10.1086/432586
Howard B, Bascom CC, Hu P et al (2022) Aging-associated changes in the adult human skin microbiome and the host factors that affect skin microbiome composition. J Investig Dermatol 142:1934–46.e21. https://doi.org/10.1016/j.jid.2021.11.029
Hug DH, Dunkerson DD, Hunter JK (1999) The degradation of l-histidine and trans- and cis-urocanic acid by bacteria from skin and the role of bacterial cis-urocanic acid isomerase. J Photochem Photobiol B 50:66–73. https://doi.org/10.1016/s1011-1344(99)00072-x
Human Microbiome Project Consortium (2012) Structure, function and diversity of the healthy human microbiome. Nature 486:207–214. https://doi.org/10.1038/nature11234
Ianiri G, Heitman J, Scheynius A (2018) The skin commensal yeast Malassezia globosa thwarts bacterial biofilms to benefit the host. J Investig Dermatol 138:1026–1029. https://doi.org/10.1016/j.jid.2018.01.008
Ianiri G, Coelho MA, Ruchti F et al (2020) HGT in the human and skin commensal Malassezia: a bacterially derived flavohemoglobin is required for NO resistance and host interaction. Proc Natl Acad Sci USA 117:15884–15894. https://doi.org/10.1073/pnas.2003473117
Imayama S, Shimozono Y, Hoashi M et al (1994) Reduced secretion of IgA to skin surface of patients with atopic dermatitis. J Allergy Clin Immunol 94:195–200. https://doi.org/10.1016/0091-6749(94)90040-x
Iwase T, Uehara Y, Shinji H et al (2010) Staphylococcus epidermidis Esp inhibits Staphylococcus aureus biofilm formation and nasal colonization. Nature 465:346–349. https://doi.org/10.1038/nature09074
Jacob S, Mcclintock MK (2000) Psychological state and mood effects of steroidal chemosignals in women and men. Horm Behav 37:57–78. https://doi.org/10.1006/hbeh.1999.1559
Janek D, Zipperer A, Kulik A et al (2016) High frequency and diversity of antimicrobial activities produced by nasal staphylococcus strains against bacterial competitors. PLoS Pathog 12:e1005812. https://doi.org/10.1371/journal.ppat.1005812
Jeong S, Ledee DR, Gordon GM et al (2012) Interaction of clusterin and matrix metalloproteinase-9 and its implication for epithelial homeostasis and inflammation. Am J Pathol 180:2028–2039. https://doi.org/10.1016/j.ajpath.2012.01.025
Johansson HJ, Vallhov H, Holm T et al (2018) Extracellular nanovesicles released from the commensal yeast Malassezia sympodialis are enriched in allergens and interact with cells in human skin. Sci Rep 8:9182. https://doi.org/10.1038/s41598-018-27451-9
Johnson TR, Gómez BI, Mcintyre MK et al (2018) The cutaneous microbiome and wounds: new molecular targets to promote wound healing. Int J Mol Sci. https://doi.org/10.3390/ijms19092699
Jokura Y, Ishikawa S, Tokuda H et al (1995) Molecular analysis of elastic properties of the stratum corneum by solid-state 13C-nuclear magnetic resonance spectroscopy. J Investig Dermatol 104:806–812. https://doi.org/10.1111/1523-1747.ep12607005
Juni E (1974) Simple genetic transformation assay for rapid diagnosis of Moraxella osloensis. Appl Microbiol 27:16–24. https://doi.org/10.1128/am.27.1.16-24.1974
Juni E, Bøvre K (2015) Bergey's manual of systematics of archaea and bacteria
Kaneko T, Shiota R, Shibuya S et al (2010) Human external ear canal as the specific reservoir of Malassezia slooffiae. Med Mycol 48:824–827. https://doi.org/10.3109/13693780903514880
Kang BS, Seo JG, Lee GS et al (2009) Antimicrobial activity of enterocins from Enterococcus faecalis SL-5 against Propionibacterium acnes, the causative agent in acne vulgaris, and its therapeutic effect. J Microbiol 47:101–109. https://doi.org/10.1007/s12275-008-0179-y
Kao MS, Huang S, Chang WL et al (2017) Microbiome precision editing: using PEG as a selective fermentation initiator against methicillin-resistant Staphylococcus aureus. Biotechnol J. https://doi.org/10.1002/biot.201600399
Kao HJ, Wang YH, Keshari S et al (2021) Propionic acid produced by Cutibacterium acnes fermentation ameliorates ultraviolet B-induced melanin synthesis. Sci Rep 11:11980. https://doi.org/10.1038/s41598-021-91386-x
Keshari S, Balasubramaniam A, Myagmardoloonjin B et al (2019) Butyric acid from probiotic Staphylococcus epidermidis in the skin microbiome down-regulates the ultraviolet-induced pro-inflammatory IL-6 cytokine via short-chain fatty acid receptor. Int J Mol Sci. https://doi.org/10.3390/ijms20184477
Khmaladze I, Leonardi M, Fabre S et al (2020) The skin interactome: a holistic, “genome-microbiome-exposome” approach to understand and modulate skin health and aging. Clin Cosmet Investig Dermatol 13:1021–1040. https://doi.org/10.2147/ccid.S239367
Kim SK, Karadeniz F (2012) Biological importance and applications of squalene and squalane. Adv Food Nutr Res 65:223–233. https://doi.org/10.1016/b978-0-12-416003-3.00014-7
Kim H, Lim YJ, Park JH et al (2012) Dietary silk protein, sericin, improves epidermal hydration with increased levels of filaggrins and free amino acids in NC/Nga mice. Br J Nutr 108:1726–1735. https://doi.org/10.1017/s0007114511007306
Kim JH, Son SM, Park H et al (2021a) Taxonomic profiling of skin microbiome and correlation with clinical skin parameters in healthy Koreans. Sci Rep 11:16269. https://doi.org/10.1038/s41598-021-95734-9
Kim SH, Yum HW, Kim SH et al (2021b) Topically applied taurine chloramine protects against UVB-induced oxidative stress and inflammation in mouse skin. Antioxidants. https://doi.org/10.3390/antiox10060867
Kirker KR, Secor PR, James GA et al (2009) Loss of viability and induction of apoptosis in human keratinocytes exposed to Staphylococcus aureus biofilms in vitro. Wound Repair Regen 17:690–699. https://doi.org/10.1111/j.1524-475X.2009.00523.x
Kirker KR, James GA, Fleckman P et al (2012) Differential effects of planktonic and biofilm MRSA on human fibroblasts. Wound Repair Regen 20:253–261. https://doi.org/10.1111/j.1524-475X.2012.00769.x
Kluytmans J, Van Belkum A, Verbrugh H (1997) Nasal carriage of Staphylococcus aureus: epidemiology, underlying mechanisms, and associated risks. Clin Microbiol Rev 10:505–520. https://doi.org/10.1128/cmr.10.3.505
Koay HF, Gherardin NA, Enders A et al (2016) A three-stage intrathymic development pathway for the mucosal-associated invariant T cell lineage. Nat Immunol 17:1300–1311. https://doi.org/10.1038/ni.3565
Krejner A, Bruhs A, Mrowietz U et al (2018) Decreased expression of G-protein-coupled receptors GPR43 and GPR109a in psoriatic skin can be restored by topical application of sodium butyrate. Arch Dermatol Res 310:751–758. https://doi.org/10.1007/s00403-018-1865-1
Krogsgård Nielsen C, Kjems J, Mygind T et al (2016) Effects of Tween 80 on growth and biofilm formation in laboratory media. Front Microbiol. https://doi.org/10.3389/fmicb.2016.01878
Kubota H, Mitani A, Niwano Y et al (2012) Moraxella species are primarily responsible for generating malodor in laundry. Appl Environ Microbiol 78:3317–3324. https://doi.org/10.1128/aem.07816-11
Kuehne A, Hildebrand J, Soehle J et al (2017) An integrative metabolomics and transcriptomics study to identify metabolic alterations in aged skin of humans in vivo. BMC Genomics 18:169. https://doi.org/10.1186/s12864-017-3547-3
Lai Y, Di Nardo A, Nakatsuji T et al (2009) Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat Med 15:1377–1382. https://doi.org/10.1038/nm.2062
Laihia JK, Kallio JP, Taimen P et al (2010) Protodynamic intracellular acidification by cis-urocanic acid promotes apoptosis of melanoma cells in vitro and in vivo. J Investig Dermatol 130:2431–2439. https://doi.org/10.1038/jid.2010.151
Lam TH, Verzotto D, Brahma P et al (2018) Understanding the microbial basis of body odor in pre-pubescent children and teenagers. Microbiome 6:213. https://doi.org/10.1186/s40168-018-0588-z
Larson PJ, Chong D, Fleming E et al (2021) Challenges in developing a human model system for skin microbiome research. J Investig Dermatol 141:228-231.e4. https://doi.org/10.1016/j.jid.2020.05.096
Lee HJ, Lee SH (2014) Epidermal permeability barrier defects and barrier repair therapy in atopic dermatitis. Allergy Asthma Immunol Res 6:276–287. https://doi.org/10.4168/aair.2014.6.4.276
Lee B, Bowden GH, Myal Y (2002) Identification of mouse submaxillary gland protein in mouse saliva and its binding to mouse oral bacteria. Arch Oral Biol 47:327–332. https://doi.org/10.1016/s0003-9969(01)00113-3
Lee DY, Huang CM, Nakatsuji T et al (2009) Histone H4 is a major component of the antimicrobial action of human sebocytes. J Investig Dermatol 129:2489–2496. https://doi.org/10.1038/jid.2009.106
Lee NY, Ibrahim O, Khetarpal S et al (2018) Dermal microflora restoration with ammonia-oxidizing bacteria Nitrosomonas eutropha in the treatment of keratosis pilaris: a randomized clinical trial. J Drugs Dermatol 17:285–288. https://doi.org/10.1001/jamadermatol.2014.2211
Legoux F, Bellet D, Daviaud C et al (2019) Microbial metabolites control the thymic development of mucosal-associated invariant T cells. Science 366:494–499. https://doi.org/10.1126/science.aaw2719
Lemoine L, Bayrambey D, Roloff A et al (2021) Commensal-related changes in the epidermal barrier function lead to alterations in the benzo[a]pyrene metabolite profile and its distribution in 3D skin. Mbio 12:e0122321. https://doi.org/10.1128/mBio.01223-21
Leung MHY, Tong X, Bastien P et al (2020) Changes of the human skin microbiota upon chronic exposure to polycyclic aromatic hydrocarbon pollutants. Microbiome 8:100. https://doi.org/10.1186/s40168-020-00874-1
Lew LC, Liong MT (2013) Bioactives from probiotics for dermal health: functions and benefits. J Appl Microbiol 114:1241–1253. https://doi.org/10.1111/jam.12137
Lew L-C, Gan C-Y, Liong M-T (2013) Dermal bioactives from lactobacilli and bifidobacteria. Ann Microbiol 63:1047–1055. https://doi.org/10.1007/s13213-012-0561-1
Li H, Goh BN, Teh WK et al (2018a) Skin commensal Malassezia globosa secreted protease attenuates Staphylococcus aureus biofilm formation. J Investig Dermatol 138:1137–1145. https://doi.org/10.1016/j.jid.2017.11.034
Li H, Yao Q, Mariscal AG et al (2018b) Epigenetic control of IL-23 expression in keratinocytes is important for chronic skin inflammation. Nat Commun 9:1420. https://doi.org/10.1038/s41467-018-03704-z
Li Z, Xia J, Jiang L et al (2021) Characterization of the human skin resistome and identification of two microbiota cutotypes. Microbiome 9:47. https://doi.org/10.1186/s40168-020-00995-7
Linehan JL, Harrison OJ, Han SJ et al (2018) Non-classical immunity controls microbiota impact on skin immunity and tissue repair. Cell 172:784–796.e18. https://doi.org/10.1016/j.cell.2017.12.033
Liu X, Zhang X, Zhang J et al (2020) Activation of aryl hydrocarbon receptor in Langerhans cells by a microbial metabolite of tryptophan negatively regulates skin inflammation. J Dermatol Sci 100:192–200. https://doi.org/10.1016/j.jdermsci.2020.10.004
Lo CW, Lai YK, Liu YT et al (2011) Staphylococcus aureus hijacks a skin commensal to intensify its virulence: immunization targeting β-hemolysin and CAMP factor. J Investig Dermatol 131:401–409. https://doi.org/10.1038/jid.2010.319
Loesche M, Gardner SE, Kalan L et al (2017) Temporal stability in chronic wound microbiota is associated with poor healing. J Investig Dermatol 137:237–244. https://doi.org/10.1016/j.jid.2016.08.009
Loomis KH, Wu SK, Ernlund A et al (2021) A mixed community of skin microbiome representatives influences cutaneous processes more than individual members. Microbiome 9:22. https://doi.org/10.1186/s40168-020-00963-1
Lousada MB, Lachnit T, Edelkamp J et al (2021) Exploring the human hair follicle microbiome. Br J Dermatol 184:802–815. https://doi.org/10.1111/bjd.19461
Lovászi M, Szegedi A, Zouboulis CC et al (2017) Sebaceous-immunobiology is orchestrated by sebum lipids. Dermatoendocrinol 9:e1375636. https://doi.org/10.1080/19381980.2017.1375636
Lugg JW, Ellis FP (1954) Some water-soluble vitamins in the sweat of tropically acclimatized European men. Br J Nutr 8:71–77. https://doi.org/10.1079/bjn19540011
Machowinski A, Krämer HJ, Hort W et al (2006) Pityriacitrin—a potent UV filter produced by Malassezia furfur and its effect on human skin microflora. Mycoses 49:388–392. https://doi.org/10.1111/j.1439-0507.2006.01265.x
Maclaughlin J, Holick MF (1985) Aging decreases the capacity of human skin to produce vitamin D3. J Clin Investig 76:1536–1538. https://doi.org/10.1172/jci112134
Mcginley KJ, Webster GF, Leyden JJ (1978) Regional variations of cutaneous propionibacteria. Appl Environ Microbiol 35:62–66. https://doi.org/10.1128/aem.35.1.62-66.1978
Mcgrath JA (2008) Filaggrin and the great epidermal barrier grief. Australas J Dermatol 49:67–73. https://doi.org/10.1111/j.1440-0960.2008.00443.x (quiz 73-4)
Midorikawa K, Ouhara K, Komatsuzawa H et al (2003) Staphylococcus aureus susceptibility to innate antimicrobial peptides, beta-defensins and CAP18, expressed by human keratinocytes. Infect Immun 71:3730–3739. https://doi.org/10.1128/iai.71.7.3730-3739.2003
Minshall C, Nadal J, Exley C (2014) Aluminium in human sweat. J Trace Elem Med Biol 28:87–88. https://doi.org/10.1016/j.jtemb.2013.10.002
Miskin JE, Farrell AM, Cunliffe WJ et al (1997) Propionibacterium acnes, a resident of lipid-rich human skin, produces a 33 kDa extracellular lipase encoded by gehA. Microbiology 143(Pt 5):1745–1755. https://doi.org/10.1099/00221287-143-5-1745
Moss CW, Wallace PL, Hollis DG et al (1988) Cultural and chemical characterization of CDC groups EO-2, M-5, and M-6, Moraxella (Moraxella) species, Oligella urethralis, Acinetobacter species, and Psychrobacter immobilis. J Clin Microbiol 26:484–492. https://doi.org/10.1128/jcm.26.3.484-492.1988
Müller DN, Wilck N, Haase S et al (2019) Sodium in the microenvironment regulates immune responses and tissue homeostasis. Nat Rev Immunol 19:243–254. https://doi.org/10.1038/s41577-018-0113-4
Murakami M (2011) Lipid mediators in life science. Exp Anim 60:7–20. https://doi.org/10.1538/expanim.60.7
Murakami M, Ohtake T, Dorschner RA et al (2002) Cathelicidin anti-microbial peptide expression in sweat, an innate defense system for the skin. J Investig Dermatol 119:1090–1095. https://doi.org/10.1046/j.1523-1747.2002.19507.x
Murphy B, Hoptroff M, Arnold D et al (2021) In-vivo impact of common cosmetic preservative systems in full formulation on the skin microbiome. PLoS ONE 16:e0254172. https://doi.org/10.1371/journal.pone.0254172
Myal Y, Robinson DB, Iwasiow B et al (1991) The prolactin-inducible protein (PIP/GCDFP-15) gene: cloning, structure and regulation. Mol Cell Endocrinol 80:165–175. https://doi.org/10.1016/0303-7207(91)90153-j
N’diaye A, Gannesen A, Borrel V et al (2017) Substance P and calcitonin gene-related peptide: key regulators of cutaneous microbiota homeostasis. Front Endocrinol 8:15. https://doi.org/10.3389/fendo.2017.00015
Naik S, Bouladoux N, Wilhelm C et al (2012) Compartmentalized control of skin immunity by resident commensals. Science 337:1115–1119. https://doi.org/10.1126/science.1225152
Naik S, Bouladoux N, Linehan JL et al (2015) Commensal-dendritic-cell interaction specifies a unique protective skin immune signature. Nature 520:104–108. https://doi.org/10.1038/nature14052
Nakatsuji T, Kao MC, Zhang L et al (2010) Sebum free fatty acids enhance the innate immune defense of human sebocytes by upregulating beta-defensin-2 expression. J Investig Dermatol 130:985–994. https://doi.org/10.1038/jid.2009.384
Nakatsuji T, Chen TH, Narala S et al (2017) Antimicrobials from human skin commensal bacteria protect against Staphylococcus aureus and are deficient in atopic dermatitis. Sci Transl Med. https://doi.org/10.1126/scitranslmed.aah4680
Nakatsuji T, Chen TH, Butcher AM et al (2018) A commensal strain of Staphylococcus epidermidis protects against skin neoplasia. Sci Adv 4:eaao4502. https://doi.org/10.1126/sciadv.aao4502
Natsch A, Gfeller H, Gygax P et al (2003) A specific bacterial aminoacylase cleaves odorant precursors secreted in the human axilla. J Biol Chem 278:5718–5727. https://doi.org/10.1074/jbc.M210142200
Nichols J, Miller AT Jr (1948) Excretion of adrenal corticoids in the sweat. Proc Soc Exp Biol Med 69:448. https://doi.org/10.3181/00379727-69-16751
Nicolaides N (1974) Skin lipids: their biochemical uniqueness. Science 186:19–26. https://doi.org/10.1126/science.186.4158.19
Nistor A, Bowden G, Blanchard A et al (2009) Influence of mouse prolactin-inducible protein in saliva on the aggregation of oral bacteria. Oral Microbiol Immunol 24:510–513. https://doi.org/10.1111/j.1399-302X.2009.00543.x
Nithya S, Radhika T, Jeddy N (2015) Loricrin—an overview. J Oral Maxillofac Pathol 19:64–68. https://doi.org/10.4103/0973-029x.157204
Nizet V, Ohtake T, Lauth X et al (2001) Innate antimicrobial peptide protects the skin from invasive bacterial infection. Nature 414:454–457. https://doi.org/10.1038/35106587
Notay M, Saric-Bosanac S, Vaughn AR et al (2020) The use of topical Nitrosomonas eutropha for cosmetic improvement of facial wrinkles. J Cosmet Dermatol 19:689–693. https://doi.org/10.1111/jocd.13060
Nunome Y, Tsuda T, Kitagawa K (2010) Determination of fatty acids in human sweat during fasting using GC/MS. Anal Sci 26:917–919. https://doi.org/10.2116/analsci.26.917
O’neill AM, Nakatsuji T, Hayachi A et al (2020) Identification of a human skin commensal bacterium that selectively kills cutibacterium acnes. J Investig Dermatol 140:1619–28.e2. https://doi.org/10.1016/j.jid.2019.12.026
Oh J, Byrd AL, Deming C et al (2014) Biogeography and individuality shape function in the human skin metagenome. Nature 514:59–64. https://doi.org/10.1038/nature13786
Ohsawa K, Watanabe T, Matsukawa R et al (1984) The possible role of squalene and its peroxide of the sebum in the occurrence of sunburn and protection from the damage caused by U.V. irradiation. J Toxicol Sci 9:151–159. https://doi.org/10.2131/jts.9.151
Ong JS, Taylor TD, Yong CC et al (2020) Lactobacillus plantarum USM8613 aids in wound healing and suppresses Staphylococcus aureus infection at wound sites. Probiotics Antimicrob Proteins 12:125–137. https://doi.org/10.1007/s12602-018-9505-9
Ono E, Murota H, Mori Y et al (2018) Sweat glucose and GLUT2 expression in atopic dermatitis: Implication for clinical manifestation and treatment. PLoS ONE 13:e0195960. https://doi.org/10.1371/journal.pone.0195960
Paharik AE, Parlet CP, Chung N et al (2017) Coagulase-negative staphylococcal strain prevents Staphylococcus aureus colonization and skin infection by blocking quorum sensing. Cell Host Microbe 22:746–56.e5. https://doi.org/10.1016/j.chom.2017.11.001
Pappas A (2009) Epidermal surface lipids. Dermatoendocrinol 1:72–76. https://doi.org/10.4161/derm.1.2.7811
Park JS, Lee EJ, Lee JC et al (2007) Anti-inflammatory effects of short chain fatty acids in IFN-gamma-stimulated RAW 264.7 murine macrophage cells: involvement of NF-kappaB and ERK signaling pathways. Int Immunopharmacol 7:70–77. https://doi.org/10.1016/j.intimp.2006.08.015
Park JH, Park GT, Cho IH et al (2011) An antimicrobial protein, lactoferrin exists in the sweat: proteomic analysis of sweat. Exp Dermatol 20:369–371. https://doi.org/10.1111/j.1600-0625.2010.01218.x
Petry T, Bury D, Fautz R et al (2017) Review of data on the dermal penetration of mineral oils and waxes used in cosmetic applications. Toxicol Lett 280:70–78. https://doi.org/10.1016/j.toxlet.2017.07.899
Picardo M, Ottaviani M, Camera E et al (2009) Sebaceous gland lipids. Dermatoendocrinology 1:68–71. https://doi.org/10.4161/derm.1.2.8472
Pinto D, Ciardiello T, Franzoni M et al (2021) Effect of commonly used cosmetic preservatives on skin resident microflora dynamics. Sci Rep 11:8695. https://doi.org/10.1038/s41598-021-88072-3
Pistone D, Meroni G, Panelli S et al (2021) A journey on the skin microbiome: pitfalls and opportunities. Int J Mol Sci. https://doi.org/10.3390/ijms22189846
Pochi PE, Strauss JS, Downing DT (1979) Age-related changes in sebaceous gland activity. J Investig Dermatol 73:108–111. https://doi.org/10.1111/1523-1747.ep12532792
Postler TS, Ghosh S (2017) Understanding the holobiont: how microbial metabolites affect human health and shape the immune system. Cell Metab 26:110–130. https://doi.org/10.1016/j.cmet.2017.05.008
Prescott SL, Larcombe DL, Logan AC et al (2017) The skin microbiome: impact of modern environments on skin ecology, barrier integrity, and systemic immune programming. World Allergy Organ J 10:29. https://doi.org/10.1186/s40413-017-0160-5
Preti G, Wysocki CJ, Barnhart KT et al (2003) Male axillary extracts contain pheromones that affect pulsatile secretion of luteinizing hormone and mood in women recipients. Biol Reprod 68:2107–2113. https://doi.org/10.1095/biolreprod.102.008268
Pullar JM, Carr AC, Vissers MCM (2017) The roles of vitamin C in skin health. Nutrients. https://doi.org/10.3390/nu9080866
Ramsey MM, Freire MO, Gabrilska RA et al (2016) Staphylococcus aureus shifts toward commensalism in response to Corynebacterium species. Front Microbiol 7:1230. https://doi.org/10.3389/fmicb.2016.01230
Reithmayer K, Meyer KC, Kleditzsch P et al (2009) Human hair follicle epithelium has an antimicrobial defence system that includes the inducible antimicrobial peptide psoriasin (S100A7) and RNase 7. Br J Dermatol 161:78–89. https://doi.org/10.1111/j.1365-2133.2009.09154.x
Ridaura VK, Bouladoux N, Claesen J et al (2018) Contextual control of skin immunity and inflammation by Corynebacterium. J Exp Med 215:785–799. https://doi.org/10.1084/jem.20171079
Rosenberg E, Koren O, Reshef L et al (2007) The role of microorganisms in coral health, disease and evolution. Nat Rev Microbiol 5:355–362. https://doi.org/10.1038/nrmicro1635
Roth RR, James WD (1988) Microbial ecology of the skin. Annu Rev Microbiol 42:441–464. https://doi.org/10.1146/annurev.mi.42.100188.002301
Roux PF, Oddos T, Stamatas G (2022) Deciphering the role of skin surface microbiome in skin health: an integrative multiomics approach reveals three distinct metabolite-microbe clusters. J Investig Dermatol 142:469–479.e5. https://doi.org/10.1016/j.jid.2021.07.159
Salminen S, Collado MC, Endo A et al (2021) The International Scientific Association of Probiotics and Prebiotics (ISAPP) consensus statement on the definition and scope of postbiotics. Nat Rev Gastroenterol Hepatol 18:649–667. https://doi.org/10.1038/s41575-021-00440-6
Sanchez Rodriguez R, Pauli ML, Neuhaus IM et al (2014) Memory regulatory T cells reside in human skin. J Clin Investig 124:1027–1036. https://doi.org/10.1172/jci72932
Sánchez-Suárez J, Coy-Barrera E, Villamil L et al (2020) Streptomyces-derived metabolites with potential photoprotective properties—a systematic literature review and meta-analysis on the reported chemodiversity. Molecules. https://doi.org/10.3390/molecules25143221
Sandby-Møller J, Poulsen T, Wulf HC (2003) Epidermal thickness at different body sites: relationship to age, gender, pigmentation, blood content, skin type and smoking habits. Acta Derm Venereol 83:410–413. https://doi.org/10.1080/00015550310015419
Sanford JA, Zhang LJ, Williams MR et al (2016) Inhibition of HDAC8 and HDAC9 by microbial short-chain fatty acids breaks immune tolerance of the epidermis to TLR ligands. Sci Immunol. https://doi.org/10.1126/sciimmunol.aah4609
Sanford JA, O’neill AM, Zouboulis CC et al (2019) Short-chain fatty acids from cutibacterium acnes activate both a canonical and epigenetic inflammatory response in human sebocytes. J Immunol 202:1767–1776. https://doi.org/10.4049/jimmunol.1800893
Sardana K, Garg VK (2010) An observational study of methionine-bound zinc with antioxidants for mild to moderate acne vulgaris. Dermatol Ther 23:411–418. https://doi.org/10.1111/j.1529-8019.2010.01342.x
Sargent F, Robinson P, Johnson R (1944) Water-soluble vitamins in sweat. J Biol Chem 153:285–294
Sato K (1977) The physiology, pharmacology, and biochemistry of the eccrine sweat gland. Rev Physiol Biochem Pharmacol 79:51–131. https://doi.org/10.1007/BFb0037089
Sato K, Sato F (1990) Na+, K+, H+, Cl−, and Ca2+ concentrations in cystic fibrosis eccrine sweat in vivo and in vitro. J Lab Clin Med 115:504–511
Sato K, Kang WH, Saga K et al (1989a) Biology of sweat glands and their disorders. I. Normal sweat gland function. J Am Acad Dermatol 20:537–563. https://doi.org/10.1016/s0190-9622(89)70063-3
Sato K, Kang WH, Saga K et al (1989b) Biology of sweat glands and their disorders. II. Disorders of sweat gland function. J Am Acad Dermatol 20:713–726. https://doi.org/10.1016/s0190-9622(89)70081-5
Scharschmidt TC, Fischbach MA (2013) What lives on our skin: ecology, genomics and therapeutic opportunities of the skin microbiome. Drug Discov Today Dis Mech. https://doi.org/10.1016/j.ddmec.2012.12.003
Scharschmidt TC, Vasquez KS, Truong HA et al (2015) A wave of regulatory T cells into neonatal skin mediates tolerance to commensal microbes. Immunity 43:1011–1021. https://doi.org/10.1016/j.immuni.2015.10.016
Scharschmidt TC, Vasquez KS, Pauli ML et al (2017) Commensal microbes and hair follicle morphogenesis coordinately drive Treg migration into neonatal skin. Cell Host Microbe 21:467–477.e5. https://doi.org/10.1016/j.chom.2017.03.001
Schenkels LC, Walgreen-Weterings E, Oomen LC et al (1997) In vivo binding of the salivary glycoprotein EP-GP (identical to GCDFP-15) to oral and non-oral bacteria detection and identification of EP-GP binding species. Biol Chem 378:83–88. https://doi.org/10.1515/bchm.1997.378.2.83
Schittek B, Hipfel R, Sauer B et al (2001) Dermcidin: a novel human antibiotic peptide secreted by sweat glands. Nat Immunol 2:1133–1137. https://doi.org/10.1038/ni732
Schwarz A, Bruhs A, Schwarz T (2017) The short-chain fatty acid sodium butyrate functions as a regulator of the skin immune system. J Investig Dermatol 137:855–864. https://doi.org/10.1016/j.jid.2016.11.014
Sears ME, Kerr KJ, Bray RI (2012) Arsenic, cadmium, lead, and mercury in sweat: a systematic review. J Environ Public Health 2012:184745. https://doi.org/10.1155/2012/184745
Serag A, Shakkour Z, Halboup AM et al (2021) Sweat metabolome and proteome: recent trends in analytical advances and potential biological functions. J Proteomics 246:104310. https://doi.org/10.1016/j.jprot.2021.104310
Solano F (2020) Metabolism and functions of amino acids in the skin. Adv Exp Med Biol 1265:187–199. https://doi.org/10.1007/978-3-030-45328-2_11
Sørensen OE, Follin P, Johnsen AH et al (2001) Human cathelicidin, hCAP-18, is processed to the antimicrobial peptide LL-37 by extracellular cleavage with proteinase 3. Blood 97:3951–3959. https://doi.org/10.1182/blood.v97.12.3951
Souak D, Barreau M, Courtois A et al (2021) Challenging cosmetic innovation: the skin microbiota and probiotics protect the skin from UV-induced damage. Microorganisms. https://doi.org/10.3390/microorganisms9050936
Staudinger T, Pipal A, Redl B (2011) Molecular analysis of the prevalent microbiota of human male and female forehead skin compared to forearm skin and the influence of make-up. J Appl Microbiol 110:1381–1389. https://doi.org/10.1111/j.1365-2672.2011.04991.x
Suffia IJ, Reckling SK, Piccirillo CA et al (2006) Infected site-restricted Foxp3+ natural regulatory T cells are specific for microbial antigens. J Exp Med 203:777–788. https://doi.org/10.1084/jem.20052056
Takahashi T, Yamasaki K (2020) Psoriasis and antimicrobial peptides. Int J Mol Sci. https://doi.org/10.3390/ijms21186791
Takken W, Kline DL (1989) Carbon dioxide and 1-octen-3-ol as mosquito attractants. J Am Mosq Control Assoc 5:311–316
Tauch A, Burkovski A (2015) Molecular armory or niche factors: virulence determinants of Corynebacterium species. FEMS Microbiol Lett 362:fnv185. https://doi.org/10.1093/femsle/fnv185
Thiboutot D (2004) Regulation of human sebaceous glands. J Investig Dermatol 123:1–12. https://doi.org/10.1111/j.1523-1747.2004.t01-2-.x
Thody AJ, Shuster S (1989) Control and function of sebaceous glands. Physiol Rev 69:383–416. https://doi.org/10.1152/physrev.1989.69.2.383
Thomas RM, Jobin C (2020) Microbiota in pancreatic health and disease: the next frontier in microbiome research. Nat Rev Gastroenterol Hepatol 17:53–64. https://doi.org/10.1038/s41575-019-0242-7
Tilles G (2014) Acne pathogenesis: history of concepts. Dermatology 229:1–46. https://doi.org/10.1159/000364860
Timm CM, Loomis K, Stone W et al (2020) Isolation and characterization of diverse microbial representatives from the human skin microbiome. Microbiome 8:58. https://doi.org/10.1186/s40168-020-00831-y
Tözsér J, Berta A (1998) Lactate dehydrogenase activity in pathological human tears obtained with glass capillaries correlates with the albumin content. Int Ophthalmol 22:289–292. https://doi.org/10.1023/a:1006378613666
Traisaeng S, Herr DR, Kao HJ et al (2019) A derivative of butyric acid, the fermentation metabolite of Staphylococcus epidermidis, inhibits the growth of a Staphylococcus aureus strain isolated from atopic dermatitis patients. Toxins. https://doi.org/10.3390/toxins11060311
Treiner E, Duban L, Bahram S et al (2003) Selection of evolutionarily conserved mucosal-associated invariant T cells by MR1. Nature 422:164–169. https://doi.org/10.1038/nature01433
Unno M, Cho O, Sugita T (2017) Inhibition of Propionibacterium acnes lipase activity by the antifungal agent ketoconazole. Microbiol Immunol 61:42–44. https://doi.org/10.1111/1348-0421.12464
Vallhov H, Johansson C, Veerman RE et al (2020) Extracellular vesicles released from the skin commensal yeast Malassezia sympodialis activate human primary keratinocytes. Front Cell Infect Microbiol 10:6. https://doi.org/10.3389/fcimb.2020.00006
Van Der Beek EJ (1991) Vitamin supplementation and physical exercise performance. J Sports Sci 9:77–89
Van Loosdrecht MC, Lyklema J, Norde W et al (1990) Influence of interfaces on microbial activity. Microbiol Rev 54:75–87. https://doi.org/10.1128/mr.54.1.75-87.1990
Vardhan M, Flaminio Z, Sapru S et al (2019) The microbiome, malignant fungating wounds, and palliative care. Front Cell Infect Microbiol 9:373. https://doi.org/10.3389/fcimb.2019.00373
Vinolo MA, Rodrigues HG, Nachbar RT et al (2011) Regulation of inflammation by short chain fatty acids. Nutrients 3:858–876. https://doi.org/10.3390/nu3100858
Wang Y, Kuo S, Shu M et al (2014) Staphylococcus epidermidis in the human skin microbiome mediates fermentation to inhibit the growth of Propionibacterium acnes: implications of probiotics in acne vulgaris. Appl Microbiol Biotechnol 98:411–424. https://doi.org/10.1007/s00253-013-5394-8
Wang Y, Wu Y, Wang Y et al (2017a) Antioxidant properties of probiotic bacteria. Nutrients. https://doi.org/10.3390/nu9050521
Wang Z, Mascarenhas N, Eckmann L et al (2017b) Skin microbiome promotes mast cell maturation by triggering stem cell factor production in keratinocytes. J Allergy Clin Immunol 139:1205–1216.e6. https://doi.org/10.1016/j.jaci.2016.09.019
Wang Q, Cui S, Zhou L et al (2019a) Effect of cosmetic chemical preservatives on resident flora isolated from healthy facial skin. J Cosmet Dermatol 18:652–658. https://doi.org/10.1111/jocd.12822
Wang X, Zhou H, Chen D et al (2019b) Whole-genome sequencing reveals a prolonged and persistent intrahospital transmission of Corynebacterium striatum, an emerging multidrug-resistant pathogen. J Clin Microbiol. https://doi.org/10.1128/jcm.00683-19
Wang G, Sweren E, Liu H et al (2021) Bacteria induce skin regeneration via IL-1β signaling. Cell Host Microbe 29:777–791.e6. https://doi.org/10.1016/j.chom.2021.03.003
Wanke I, Steffen H, Christ C et al (2011) Skin commensals amplify the innate immune response to pathogens by activation of distinct signaling pathways. J Investig Dermatol 131:382–390. https://doi.org/10.1038/jid.2010.328
Watabe A, Sugawara T, Kikuchi K et al (2013) Sweat constitutes several natural moisturizing factors, lactate, urea, sodium, and potassium. J Dermatol Sci 72:177–182. https://doi.org/10.1016/j.jdermsci.2013.06.005
Watanabe S, Kano R, Sato H et al (2001) The effects of Malassezia yeasts on cytokine production by human keratinocytes. J Investig Dermatol 116:769–773. https://doi.org/10.1046/j.1523-1747.2001.01321.x
Wei Q, Li Z, Gu Z et al (2022) Shotgun metagenomic sequencing reveals skin microbial variability from different facial sites. Front Microbiol. https://doi.org/10.3389/fmicb.2022.933189
Wikler JR, Janssen N, Bruynzeel DP et al (1990) The effect of UV-light on pityrosporum yeasts: ultrastructural changes and inhibition of growth. Acta Derm Venereol 70:69–71
Williams GC (1996) Adaptation and natural selection. Princeton University Press, Princeton
Williams MR, Costa SK, Zaramela LS et al (2019) Quorum sensing between bacterial species on the skin protects against epidermal injury in atopic dermatitis. Sci Transl Med. https://doi.org/10.1126/scitranslmed.aat8329
Wisecaver JH, Alexander WG, King SB et al (2016) Dynamic evolution of nitric oxide detoxifying flavohemoglobins, a family of single-protein metabolic modules in bacteria and eukaryotes. Mol Biol Evol 33:1979–1987. https://doi.org/10.1093/molbev/msw073
Wollenberg MS, Claesen J, Escapa IF et al (2014) Propionibacterium-produced coproporphyrin III induces Staphylococcus aureus aggregation and biofilm formation. Mbio 5:e01286-14. https://doi.org/10.1128/mBio.01286-14
Wondrak GT, Jacobson MK, Jacobson EL (2006) Endogenous UVA-photosensitizers: mediators of skin photodamage and novel targets for skin photoprotection. Photochem Photobiol Sci 5:215–237. https://doi.org/10.1039/b504573h
Wu G, Zhao H, Li C et al (2015) Genus-wide comparative genomics of malassezia delineates its phylogeny, physiology, and niche adaptation on human skin. PLoS Genet 11:e1005614. https://doi.org/10.1371/journal.pgen.1005614
Xia X, Li Z, Liu K et al (2016) Staphylococcal LTA-induced miR-143 inhibits propionibacterium acnes-mediated inflammatory response in skin. J Investig Dermatol 136:621–630. https://doi.org/10.1016/j.jid.2015.12.024
Xu M, Lu H, Lee YH et al (2018) An interleukin-25-mediated autoregulatory circuit in keratinocytes plays a pivotal role in psoriatic skin inflammation. Immunity 48:787–98.e4. https://doi.org/10.1016/j.immuni.2018.03.019
Yamane T, Morioka Y, Kitaura Y et al (2018) Branched-chain amino acids regulate type I tropocollagen and type III tropocollagen syntheses via modulation of mTOR in the skin. Biosci Biotechnol Biochem 82:611–615. https://doi.org/10.1080/09168451.2017.1386084
Yamasaki K, Schauber J, Coda A et al (2006) Kallikrein-mediated proteolysis regulates the antimicrobial effects of cathelicidins in skin. FASEB J 20:2068–2080. https://doi.org/10.1096/fj.06-6075com
Yang ES, Tan J, Eells S et al (2010) Body site colonization in patients with community-associated methicillin-resistant Staphylococcus aureus and other types of S. aureus skin infections. Clin Microbiol Infect 16:425–431. https://doi.org/10.1111/j.1469-0691.2009.02836.x
Yokozeki H, Hibino T, Takemura T et al (1991) Cysteine proteinase inhibitor in eccrine sweat is derived from sweat gland. Am J Physiol 260:R314–R320. https://doi.org/10.1152/ajpregu.1991.260.2.R314
Youn SH, Choi CW, Choi JW et al (2013) The skin surface pH and its different influence on the development of acne lesion according to gender and age. Skin Res Technol 19:131–136. https://doi.org/10.1111/srt.12023
Yu Y, Prassas I, Muytjens CM et al (2017) Proteomic and peptidomic analysis of human sweat with emphasis on proteolysis. J Proteomics 155:40–48. https://doi.org/10.1016/j.jprot.2017.01.005
Yu J, Luo Y, Zhu Z et al (2019) A tryptophan metabolite of the skin microbiota attenuates inflammation in patients with atopic dermatitis through the aryl hydrocarbon receptor. J Allergy Clin Immunol 143:2108–19.e12. https://doi.org/10.1016/j.jaci.2018.11.036
Zhang YJ, Han Y, Sun YZ et al (2019) Extracellular vesicles derived from Malassezia furfur stimulate IL-6 production in keratinocytes as demonstrated in in vitro and in vivo models. J Dermatol Sci 93:168–175. https://doi.org/10.1016/j.jdermsci.2019.03.001
Zhang H, Zhu Y, Liu Z et al (2022) A volatile from the skin microbiota of flavivirus-infected hosts promotes mosquito attractiveness. Cell 185:2510–2522.e16. https://doi.org/10.1016/j.cell.2022.05.016
Zheng Y, Hunt RL, Villaruz AE et al (2022) Commensal Staphylococcus epidermidis contributes to skin barrier homeostasis by generating protective ceramides. Cell Host Microbe 30:301–13.e9. https://doi.org/10.1016/j.chom.2022.01.004
Zipperer A, Konnerth MC, Laux C et al (2016) Human commensals producing a novel antibiotic impair pathogen colonization. Nature 535:511–516. https://doi.org/10.1038/nature18634
Acknowledgements
This work was supported by the Shanghai Municipal Science and Technology Major Project (2017SHZDZX01), the CAMS Innovation Fund for Medical Sciences (2019-I2M-5-066), the 111 Project (B13016), and a startup grant from the Greater Bay Area Institute of Precision Medicine (Guangzhou), Fudan University to JX.
Author information
Authors and Affiliations
Contributions
Original draft preparation: HC, JX and QZ; Review and editing: HC, JX, JW, QZ, CD; Scientific supervision: JX, JW, JK. All authors have read and agreed to this version of the manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no conflict.
Consent to Participate
Not applicable.
Consent for Publication
Not applicable.
Ethical Approval
Not applicable.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/
About this article

Cite this article
Chen, H., Zhao, Q., Zhong, Q. et al. Skin Microbiome, Metabolome and Skin Phenome, from the Perspectives of Skin as an Ecosystem. Phenomics 2, 363–382 (2022). https://doi.org/10.1007/s43657-022-00073-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s43657-022-00073-y