
Abstract
African elephants (Loxodonta africana) are increasingly exposed to high levels of human disturbance and are threatened by poaching and human–elephant conflict. As anthropogenic pressures continue to increase, both inside and outside protected areas, understanding elephant behavioural responses to human activity is required for future conservation management. Here, we use bycatch data from camera trap surveys to provide inferences on elephant habitat use and temporal activity in Kasungu National Park (KNP), Malawi. The KNP elephant population has declined by ~ 95% since the late 1970s, primarily because of intensive poaching, and information on elephant ecology and behaviour can assist in the species’ recovery. Using occupancy modelling, we show that proximity to water is the primary driver of elephant habitat use in KNP, with sites closer to water having a positive effect on elephant site use. Our occupancy results suggest that elephants do not avoid sites of higher human activity, while results from temporal activity models show that elephants avoid peak times of human activity and exhibit primarily nocturnal behaviour when using the KNP road network. As key park infrastructure is located near permanent water sources, elephant spatiotemporal behaviour may represent a trade-off between resource utilisation and anthropogenic-risk factors, with temporal partitioning used to reduce encounter rates. Increased law enforcement activity around permanent water sources could help to protect the KNP elephant population during the dry season. Our findings highlight that camera trap bycatch data can be a useful tool for the conservation management of threatened species beyond the initial scope of research.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
African elephants (Loxodonta africana; hereafter elephant) play an integral role in ecosystem processes (Kerley and Landman 2006; Ripple et al. 2015; Coverdale et al. 2016) and can provide economic and social benefits through wildlife-based tourism (Naidoo et al. 2016; Ripple et al. 2016). Nevertheless, elephants have experienced significant range contractions and population declines, which have accelerated over the last two decades (Wittemyer et al. 2014; Chase et al. 2016). Unsustainable illegal hunting (Wittemyer et al. 2014; Hauenstein et al. 2019), habitat loss (Ripple et al. 2015; Wall et al. 2021) and human-elephant conflict (Di Minin et al. 2021) are key drivers of elephant population decline across Africa. As anthropogenic pressures continue to increase across protected areas and surrounding land (Jones et al. 2018a), it is predicted that the drivers of elephant population decline will intensify and risk further localised extirpations (Ripple et al. 2015; Wall et al. 2021).
Anthropogenic disturbance, both inside and outside protected areas, can have implications for animal behaviour, human-wildlife conflict and population demographics (Nyhus 2016; Di Marco et al. 2018; Suraci et al. 2019). Areas of higher human activity can often represent a complex trade-off for animals, as heightened activity in these areas can increase encounter rates with humans but also provide access to high-quality food resources and potentially reduced levels of interspecific competition (Brown et al. 1999; Ogutu et al. 2014; Bastille-Rousseau et al. 2020). As a result, animals may adopt spatiotemporal responses in human-dominated areas, whereby they adjust their use of space and/or time to minimise the risk of human interaction whilst maintaining access to resources (Gunn et al. 2014; Gaynor et al. 2018). Owing to their flexible activity patterns, advanced cognitive ability, and tendency to utilise anthropogenic resources, elephants are a model species for investigating spatiotemporal responses to human disturbance (Gaynor et al. 2018). As 82% of protected areas containing elephants are adjacent to areas with significant human pressures (Di Minin et al. 2021), understanding elephant ecology and the species’ behavioural responses to anthropogenic activity is important for future conservation and management efforts (Bastille-Rousseau et al. 2020; Wall et al. 2021).
Malawi, in south-central Africa, exemplifies the conservation issues facing both elephants and wider biodiversity. The human population in Malawi is predicted to double by 2038 (World Bank 2020), and with ~ 60% of the total land area already used for agricultural purposes (FAO 2019), protected areas are becoming increasingly isolated and targeted for bushmeat hunting and habitat alteration (Gondwe et al. 2019; van Velden et al. 2020). The national elephant population in Malawi has declined by ~ 21% over a twenty-year period, from approximately 1651 individuals in 1995 to an estimated 1307 individuals in 2015 (Thouless et al. 2016), although in some areas population declines have been more severe (NEAPM 2015). For example, between 1978 and 2003, aerial counts in Kasungu National Park (KNP), Malawi, revealed that the elephant population had declined by approximately 95% (Bhima et al. 2003). The elephant population decline in KNP is primarily a result of unsustainable illegal hunting and encroachment into park buffer zones increasing human-elephant conflict (Bhima et al. 2003; NEAPM 2015). The drivers of elephant population decline in KNP are synonymous with the causes of elephant decline throughout protected areas in Malawi (NEAPM 2015). However, most Malawian protected areas lack detailed ecological information to inform elephant management strategies and aid population recovery (NEAPM 2015; Sievert et al. 2022). Consequently, information on elephant behaviour and ecology in KNP can potentially inform wider management strategies in areas that are currently data deficient.
Camera trap studies and bycatch data on non-target species are increasingly used to answer ecological questions beyond the initial scope of research (Caravaggi et al. 2017; Edwards et al. 2018). As camera trap surveys are often employed to collect data on elusive species that are found at low densities (e.g., leopard Panthera pardus), the amount of bycatch data for non-target species can often outweigh that of target animals and provide a valuable source of data for species of conservation concern (Edwards et al. 2018). In this study, we use bycatch data from large carnivore camera trap surveys (Davis et al. 2021a, b) to investigate elephant habitat use and spatiotemporal responses to human activity in KNP. We apply occupancy and temporal activity models to determine the impact of a range of environmental and anthropogenic effects on elephant site use and use these data to inform conservation management in KNP. Elephants in KNP have experienced high rates of illegal hunting and have previously been found to alter their spatiotemporal behaviour in areas of increased human activity (Gaynor et al. 2018; Ihwagi et al. 2018), therefore, we predict that elephants will display spatial avoidance of areas of higher human activity and exhibit primarily nocturnal behaviour to minimise interaction with humans.
Materials and methods
Study area
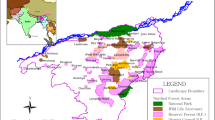
Kasungu National Park is a 2316 km2 legally protected area located in the Central Region of Malawi and forms an international border with Zambia along its western boundary (Fig. 1). KNP is predominantly covered by closed canopy miombo woodland, comprising Brachystegia and Julbernardia spp., interspersed with seasonally wet grasslands and isolated rocky inselbergs. Elevation ranges between 1000 and 1500 m and the mean annual rainfall is 780 mm, which primarily falls between November and April (Bhima et al. 2003). During the dry season (May–October) the average daily temperature is 25.6 °C, with sunrise between 05:30 and 06:10 and sunset between 17:27 and 17:43.

Map showing a the location and key features of KNP, with the inset map showing the location of KNP within Malawi (shaded), b camera trap locations for the 2017 survey, c camera trap locations for the 2018 survey and d the forest cover (%) layer for KNP used in occupancy modelling
Illegal hunting pressure has led to widespread declines in the elephant and large mammal populations in KNP (Munthali and Mkanda 2002; Bhima et al. 2003; Davis et al. 2021a). The KNP elephant population has declined from an estimated 2000 individuals in the late 1970s (Bell 1978 in Bhima et al. 2003) to a reported low of 40 individuals in 2014 (Macpherson 2015). A poaching spike in the early 1980s caused a significant decline in the KNP elephant population and this trend has continued through to the present day, with poaching incidents recorded during this study. A low scout density (1 scout/52.6 km2), a lack of adequate resources and difficulties policing the international border are considered key reasons for the ongoing poaching issues in KNP (NEAPM 2015). Poaching pressure can alter elephant population structure and demographics, leading to large aggregations as a risk avoidance strategy (Abe 1994) or skewing group sizes and sex ratios (Jones et al. 2018b; Mkuburo et al. 2020). However, elephants in KNP are still observed in normal family units and bull groups (Macpherson 2015).
Kasungu National Park is bordered by communal land in both Malawi and Zambia, with the surrounding land used primarily for subsistence farming and tobacco production (Munthali and Mkanda 2002; NEAPM 2015). Encroachment and agricultural expansion are regarded as significant threats to the KNP elephant population, with the potential to increase human-elephant conflict levels and reduce available habitat (NEAPM 2015). No human settlements, besides national park authorities (operating from eleven scout camps inside the park), are permanently based in KNP, and trophy hunting is not permitted in the park. Park roads are used for management, tourism, and research purposes, with no public traffic through the park. As a result of reduced animal densities, the presence of tsetse flies, and limited park infrastructure, tourist numbers are low in KNP. Elephants exhibit both avoidance and aggressive behaviour in response to vehicles in KNP, particularly in areas further from human settlements, as continued poaching pressure, human-elephant conflict in surrounding areas, and low tourism rates have limited habituation levels.
Camera trap surveys
We used bycatch data from annual camera trap surveys (2016–2018) designed to estimate large carnivore density in KNP (see Davis et al. 2021a, b). Due to the low sample size for elephant captures in 2016, we only used the 2017 and 2018 datasets for our study. Camera trap surveys were conducted once a year, with camera trap stations (2017, n = 50; 2018, n = 25) deployed for 120 days (23rd June–20th October) and 90 days (27th June–25th September) in 2017 and 2018, respectively. Roads and major trails were prioritised for camera placement to maximise the rate of photographic captures for target species, as large carnivores frequently travel along road networks (Davis et al. 2021a, b). We deployed one camera per station, and stations were checked regularly to maintain camera function and data collection. We used a combination of white-flash (Cuddeback Models C and F; Cuddeback Inc., Green Bay, WI, USA) and infrared (Bushnell Trophy Cam HD; Bushnell Corporation, Overland Park, KS, USA) camera models.
Occupancy modelling
Occupancy design
Occupancy models use replicated detection/non-detection data to produce unbiased estimates of occupancy (Ψ; the probability of a site being occupied by a species) and detection (p; probability of detecting a species) (MacKenzie et al. 2017). By incorporating covariates of interest, occupancy models can account for heterogeneity in Ψ and p, allowing researchers to make inferences about factors driving habitat use by species (MacKenzie et al. 2017). We developed a binary matrix of detection/non-detection capture histories for elephants at each camera trap station in KNP, where “1” indicated an elephant detection during a sampling occasion and “0” indicated no detection. Sampling occasions were pooled into 10 day intervals to account for a limited sample size and reduce non-detections. We removed one camera trap station from analyses due to camera malfunction. Occupancy models assume temporal and geographic closure, whereby sites are closed to changes in occupancy and detection histories between sites are independent (MacKenzie et al. 2002). As our camera trap survey length was a maximum of 120 days, and elephants are a highly mobile species with an average home range of 172.9 km2 (Wall et al. 2021), we relaxed the closure assumptions of occupancy modelling by interpreting occupancy (Ψ) as the probability of site use, rather than the proportion of area occupied (MacKenzie et al. 2017).
Detection covariates
Incorporating site-specific and/or observation-specific detection parameters into occupancy models can account for heterogeneity in detection probability (MacKenzie et al. 2017). We identified four covariates thought to potentially influence elephant detection probability between sampled sites: camera trap effort, camera model, year sampled and trail type. As camera trap effort, defined here as the number of days a camera was active during a sampling occasion (minimum = 0, maximum = 10), can affect species detection probability (e.g., Wevers et al. 2021) we included it as an observational covariate. Camera models and flash types can differ in their detection range and trigger speed, which can impact detection probability (Apps and McNutt 2018). The two Cuddeback camera models we deployed (Models C and F) have identical trigger speeds and were therefore pooled together, resulting in two camera type categories (infrared, Bushnell Trophy Cam; white flash, Cuddeback Models) for the camera model covariate. The year in which a camera was deployed (2017 or 2018) was included as a site-specific covariate. Although pooling data across years relaxes the basic assumption of temporal closure in occupancy studies (MacKenzie et al. 2017), we reasoned that annual variation in detection probability would be minimal over the timeframe of the study. This approach has been used successfully in other occupancy studies to account for temporal variation in detectability (e.g., Wevers et al. 2021). Non-random camera placement (e.g., targeted placement on roads) is known to influence species detectability (Cusack et al. 2015). As camera traps were deployed on roads and trails in KNP we included a trail type site covariate, comprising three categories: primary road (two-way government-maintained dirt road), secondary road (single-track dirt road) and game trail (paths created by animals).
Site use covariates
Based on evidence from previous studies, we selected six covariates that could predict elephant site use across KNP: distance to water (km), human activity (average number of human captures per station), tree cover (%), habitat quality (vegetation index), distance to scout camp (km), and distance to park border (km). Distance to water was incorporated as an indicator of habitat use based on species-specific requirements for water (Chamaillé-Jammes et al. 2007; Abraham et al. 2021). KNP has no buffer zone and human settlements often begin at the park boundary (Munthali and Mkanda 2002). Encroachment and habitat loss to agricultural expansion have been identified as key threats to the KNP elephant population (NEAPM 2015). Therefore, distance to park border was included as a measure of human activity indicating the potential for human-elephant conflict. Elephants can perceive differences in risk and discriminate between human threats (McComb et al. 2014; Goldenberg et al. 2017). For example, elephants may aggregate in areas closer to ranger posts, as the increased ranger presence acts as a deterrent to poachers, whilst elephants will adapt their behavioural responses to acoustic cues from human groups that pose varying threat levels (McComb et al. 2014; Beale et al. 2018). We included the distance to scout camp covariate as a proxy for habitat ‘safety’, reasoning that camera stations closer to scout camps would provide greater refuge from poaching pressure (Beale et al. 2018). For all distance-based covariates, the Euclidian distance (km) between each camera trap and the chosen feature was extracted in QGIS v.3.20.2 (QGIS Development Team 2020).
To account for the effects of human activity on elephant behaviour and distribution (Buij et al. 2007; Gaynor et al. 2018), we calculated the average number of human captures per day per station by dividing the total number of human captures (defined as any human activity on foot or in a vehicle by park staff, researchers or tourists) per camera trap station, by the number of active trap nights per station. The use of human images from camera trap data followed the ethical code of conduct outlined by Sharma et al. (2020). Permission to use camera traps was obtained from the appropriate government department and no data on individuals or activities were recorded or shared without their prior consent (Sharma et al. 2020).
Spatial variability in vegetation cover and habitat quality can affect elephant distribution (Harris et al. 2008; Chibeya et al. 2021). We used the MODIS Terra Vegetation Continuous Fields dataset (MOD44B; Dimiceli et al. 2015) to measure the percentage cover of tree-vegetation (henceforth tree cover), as a metric for woody vegetation. Habitat quality was measured using the Normalized Difference Vegetation Index (NDVI), a frequently used proxy for primary productivity and habitat ‘greenness’ (Pettorelli et al. 2011). NDVI measures the difference between near-infrared (which vegetation strongly reflects) and red light (which vegetation absorbs) from remote sensing data, resulting in a value between − 1 and 1, where positive values indicate higher habitat productivity (Pettorelli et al. 2011). We calculated NDVI from the surface-reflectance bands of the MODIS terra satellite (MOD13Q1; Didan 2015). Both MODIS vegetation layers had a resolution of 250 m × 250 m and we extracted values for each cell that contained a camera trap station.
Occupancy analyses
We analysed presence-absence camera trap data in a single-season, single-species occupancy model, using the package ‘unmarked’ v1.1.1 (Fiske and Chandler 2011) in the R environment (version 4.1.1; R Core Team 2021). We followed a two-step modelling approach, by first identifying the detection covariates that best explained heterogeneity in detection probability (p) before modelling occupancy variables (MacKenzie et al. 2017). First, we fitted single-season occupancy models, where occupancy (Ψ) remained constant (using the most parametrised model of non-correlated covariates), and p varied by either a single covariate or all possible combinations of detection covariates. We limited candidate models to a maximum of two detection covariates to avoid overparameterisation. Then, using model selection, we identified the detection model with the lowest AIC value and retained these detection covariates for testing the significance of occupancy covariates (Karanth et al. 2011). Following this, we modelled the probability of site use (Ψ), using only the top-ranked detection covariates identified in the previous stage, and all possible combinations (additive only, no interactions) of site use covariates. We used a maximum of three site use covariates per single model, using the rule of thumb of 15–25 observations per predictor variable (Henschel et al. 2016). We standardised site covariates to a mean of zero and a standard deviation of one using the R-package ‘vegan’ (Oksanen et al. 2020) and tested for collinearity using Pearson’s correlation test, excluding covariates from the same model if correlated at r > 0.7 (Dormann et al. 2013).
We used model selection to determine the top model by comparison of AIC values and retained all models considered to have strong support (ΔAIC < 2) in the final model set (Burnham and Anderson 2002). From the final model set, we applied model averaging to obtain average β-coefficient estimates and determine the importance and direction of site use covariates. To assess the relative contribution of each covariate in explaining elephant occupancy in KNP, we calculated the summed model weights (Σw) of all site use covariates in the final model set. This metric can be used to assess the overall level of support for a given variable (Burnham and Anderson 2002). Values can range from zero to one, where a value of one indicates that a covariate is present in all models, and variables with high summed model weights are considered more important in explaining heterogeneity in site use (Henschel et al. 2016). Model averaging and summed model weights were calculated using the “MuMIn” package (Barton 2020). Model fit and overdispersion (ĉ > 1.1) were evaluated using a goodness-of-fit test with Pearson’s chi-square statistic and 10,000 parametric bootstraps (MacKenzie and Bailey 2004).
Temporal overlap
We used the time and date stamps from photographic captures to describe the daily activity levels (percentage of time spent active over the 24 h daily cycle) and temporal overlap of elephants and humans in KNP. We followed the criteria of Gaynor et al. (2018) to define independent elephant capture events. Human activity was defined as any vehicle or person on foot passing a camera trap. Time of day was converted to solar time (i.e., adjusted according to sunrise and sunset) using the ‘solartime’ function in the package “activity” v.1.3. (Rowcliffe 2019). After testing for differences between survey years, to ensure no bias between individual years, we combined the 2017 and 2018 datasets for the final temporal analyses. Before conducting our temporal analyses, we performed a Hermans-Rasson test on temporal data (Landler et al. 2019), using the package “CircMLE” (Fitak and Johnsen 2017), to determine whether activity patterns were significantly different to a random distribution over the circadian cycle (Havmøller et al. 2020).
Daily activity levels were estimated using the Kernel circular density function (Ridout and Linkie 2009; Rowcliffe et al. 2014). We used the overlap coefficient (Δ), which uses a scale from 0 (no overlap) to 1 (total overlap), to quantify the degree of overlap between elephant and human activity. Following Meredith and Ridout (2016), we used the Δ4 estimator for the overlap coefficient as both sample sizes were > 75 observations. Ninety-five percent confidence intervals were calculated for Δ, using 10,000 smoothed bootstrap repetitions for each distribution. Kernel density estimation curves were produced using the R-package ‘overlap’ v.0.3.2 (Meredith and Ridout 2014). We used the function compareCkern() in the package “activity” v.1.3. (Rowcliffe 2019) and 10,000 bootstrap samples to test for significant differences between the two activity curves. The compareCkern() function tests the probability that two activity curves come from the same distribution (Rowcliffe 2019; Havmøller et al. 2020).
Results
A survey effort of 4707 camera-trap nights at 74 camera stations resulted in 145 elephant capture events. Elephants were captured at 41% of camera stations (n = 30 stations). Using 10 day sampling occasions resulted in a combined total of 490 occasions at 74 stations (average: 6.62 occasions per station), with elephants recorded on 91 sampling occasions.
Occupancy
Detection covariates Effort + Trail best explained elephant detection probability (Table 1). Camera trap effort (number of trap nights per sampling occasion) positively influenced elephant detection probability (βeffort = 0.24 ± SE 0.12). Elephant detectability was higher on secondary roads (βsecondary = 2.77 ± SE 1.04) and trails (βtrail = 0.61 ± SE 1.50), when compared to primary roads.
Three models had substantial support (< 2 ΔAIC) for elephant occupancy (Table 2). Distance to water was the most important predictor explaining elephant site use (βwater = − 2.22 ± SE 0.61; Σw = 1.00), with elephant occupancy declining at sites further from water (Fig. 2). Tree cover (%) (Σw = 0.68), and human activity (Σw = 0.38) were both present in the final candidate models, but only distance to water had a significant effect on elephant site use (Table 3). A goodness-of-fit test on the top-ranked model in the final candidate set did not indicate evidence of overdispersion (ĉ = 0.96) or lack of fit (p = 0.41).

The effect of distance to water (km) on the expected probability of elephant site use (Ψ) in Kasungu National Park, Malawi. Predicted Ψ was based on the top-ranked occupancy model. The grey shaded area represents 95% confidence intervals
Temporal activity
We observed a temporal overlap average of Δ = 0.34 (0.28–0.40) between elephants and humans. Elephant use of the diel cycle was statistically different compared to human activity (p < 0.001; Fig. 3). Elephant use of the KNP road network showed peaks after sunset (18:00) and before sunrise (04:00–06:00), with 83% of elephant photographic captures during nocturnal hours (18:00–06:00). Human activity was predominantly between the hours of 06:00 and 17:00, peaking between 10:00 and 16:00. Overall activity (proportion of time spent active over the diel cycle) was estimated as 0.43 (SE = 0.06) for elephants and 0.49 (SE = 0.01) for humans, with the Hermans-Rasson test indicating that both species had activity patterns that were significantly different from random (p < 0.001 for both).

Diel distribution and coefficient of overlap (Δ) of elephant and human activity on roads in Kasungu National Park, Malawi. The grey region shows the overlap between the two distributions. The coefficient of overlap value is presented, along with 95% confidence intervals
Discussion
Understanding the factors influencing elephant ecology and their behavioural responses to human disturbance can help mitigate elephant population decline and inform conservation and management (Fritz 2017). Here, using a combination of occupancy and temporal activity models, we examined the drivers of elephant habitat use and temporal activity in KNP, a Malawian protected area that has experienced an historical decline in elephant population numbers (Bhima et al. 2003). During the dry season, proximity to water was the key driver of elephant site use in KNP. Contrary to our prediction, elephants exhibited no spatial response to areas of increased human activity or areas of higher potential risk (park borders). Instead, our findings suggest that elephants in KNP adjust their temporal activity, exhibiting primarily nocturnal behaviour around the KNP road network, potentially to minimise encounters with humans.
Our occupancy modelling results corroborate existing work suggesting that elephant site use is driven by the availability of water (Chamaillé-Jammes et al. 2007; Harris et al. 2008; Abraham et al. 2021). Elephants need to drink frequently and prefer to travel short distances (Harris et al. 2008; Thaker et al. 2019), therefore, increased elephant site use close to water is expected in KNP. The predicted probability of elephant site use declined by ~ 50% at distances more than 3 km from water sources. These findings suggest that, during the dry season, increased law enforcement activity around permanent water areas may help to reduce poaching pressure on the KNP elephant population. Riparian areas in KNP are comprised of nutrient-rich vegetation and are frequented by large herbivores (Bhima et al. 2003), while detection rates for threatened carnivore species were also found to increase at sites closer to water in KNP (Davis et al. 2021b). As a result, increased law enforcement activity around water sources may also benefit herbivore and large carnivore populations.
In this study, increased human activity had no significant effect on elephant site use in KNP. However, elephant detection was significantly higher on secondary roads and trails than primary roads, suggesting elephants utilise areas of reduced vehicle traffic. In addition, elephant and human activity patterns were significantly different, with elephants displaying primarily nocturnal behaviour when using the KNP road network. The nocturnal activity patterns of elephants, when using human infrastructure, may suggest avoidance of anthropogenic activity in KNP, as elephant activity often peaks in the early morning and late afternoon (e.g., Shannon et al. 2008; Kasozi et al. 2022). Increased nocturnal activity could also be a response to thermal stress, as elephants are known to shift their activity peaks to cooler times of the day to aid thermoregulation and move over longer distances (Kinahan et al. 2007; Thaker et al. 2019). The observed temporal activity pattern may allow elephants to utilise the KNP road network as movement corridors (Gaynor et al. 2018), travelling over longer distances whilst it is cooler and minimising interaction with humans.
Our temporal activity results support findings from Gorongosa National Park (GNP), Mozambique, where elephants exhibited increased crepuscular and nocturnal behaviour when using roads (Gaynor et al. 2018). In our study, 83% of elephant activity on roads occurred during nocturnal hours, in accordance with Okita-Ouma et al. (2021) who found that 78% of elephant railway crossings took place at night. Similarly, Wadey et al. (2018) observed that Asian elephants (Elephas maximus) crossed roads more frequently at night. In accordance with these previous findings, KNP elephants appear to reduce their use of roads during times of peak vehicle traffic (Gaynor et al. 2018). Elephants can show behavioural responses to vehicle disturbance, including increased conspecific- and human-directed aggression (Szott et al. 2019a, b) and physiological stress (Szott et al. 2019a, b; Oduor et al. 2020), which can have long-term consequences for reproduction and individual fitness. As elephants in KNP often exhibit stress behaviour around vehicles and can be aggressive towards humans, it is recommended that speed limits and guidelines on minimum safe viewing distances are introduced and enforced in KNP to reduce disturbance levels for elephants.
The lack of spatial avoidance exhibited by elephants in areas of higher human activity may be due to the presence of permanent water near key park infrastructure (i.e., park headquarters and scout camps) and park borders. Previous studies have indicated that elephants can differentiate between human risks (McComb et al. 2014) and preferentially use areas of increased safety from poaching pressure, such as areas close to scout camps (Beale et al. 2018). Our results suggest that elephant site use in the dry season is not driven by risk avoidance, as neither the distance to park border nor scout camp covariates influenced the probability of elephant site use. As such, we reason that water is the driving factor influencing habitat selection. Elephant site use during the dry season may represent a trade-off between resource access and human-risk factors. Similar trade-offs between resource access and avoidance of human activity have been observed in other elephant populations. For example, Bastille-Rousseau et al. (2020) found that elephants in the Laikipia/Samburu ecosystem, Kenya, prioritised water and forage access over avoidance of human settlements. By adjusting their temporal activity and limiting their use of park infrastructure (i.e., roads) to times of reduced anthropogenic activity, elephants in KNP can utilise key resources whilst minimising direct contact with people.
Elephants in KNP showed no spatial response to areas of anthropogenic activity, despite the historic influence of human predation (i.e., ivory poaching) on the local population (Bhima et al. 2003). Our results suggest that proximity to the KNP boundary has no effect on elephant site use, evidenced by the absence of this covariate in our final set of candidate models, and elephants are likely to be found in areas of increased human activity. This may have implications for human-elephant conflict (HEC), as elephants move closer to the park boundary the possibility of crop raiding increases, particularly during the dry season when crop raiding incidents often peak in elephant populations (e.g., King et al. 2017). Elephant site use near park borders may also be related to the presence of the KNP river network, which runs parallel to the park’s eastern boundary. Although water sources are widespread within KNP (Macpherson 2020), prolonged periods of drought, which are predicted to increase with climate change (Warnatzsch and Reay 2019), may increase elephant movements outside of the park and impact levels of HEC. Efforts to install fencing along a 40 km section of the eastern perimeter of KNP (IFAW 2020), where communities have experienced conflict with elephants, could address potential HEC issues. However, recent studies have indicated that fencing can shift areas of HEC (Osipova et al. 2018). Continued monitoring of elephant movements and HEC would be useful for HEC management strategies in KNP.
We acknowledge that the placement of camera traps on the KNP road network may have limited our ability to predict drivers of elephant site use, as elephants may reduce their use of the road network to avoid vehicles (e.g., Blake et al. 2008). However, Gaynor et al. (2018) found that cameras placed on the road network in GNP detected similar overall levels of elephant activity compared to camera grids deployed off roads. Therefore, we believe our results are representative of elephant behaviour in KNP during the dry season. As the availability of surface water and food production increases during the wet season, elephants may disperse into less human-utilised areas (Martin et al. 2010). Investigating the drivers of habitat use and elephant behaviour in anthropogenic areas between seasons would be beneficial for law enforcement and management. Camera trap surveys are difficult in KNP during the wet season, as road access is limited and vegetation obscures camera detections (Davis et al. 2021a, b). Consequently, wet season monitoring of elephant behaviour in KNP is not feasible using camera traps. Future studies would benefit from satellite collaring elephants in KNP, providing fine-scale data on elephant movements and improving our knowledge of elephant distribution in the wet season. Initial elephant collaring efforts in Vwaza Marsh Wildlife Reserve and Namizimu Forest Reserve, Malawi, have highlighted the benefits of collaring for investigating transboundary movements and potential areas of HEC (Sievert et al. 2022).
Our study has added to the growing body of literature highlighting the benefits of camera trap bycatch data for providing additional ecological information on non-target species (Edwards et al. 2018; Gaynor et al. 2018; Williams et al. 2021). Funding limitations, time budgets and the sparsity of resources often restricts the ability of conservation practitioners to make informed conservation management decisions (e.g., Lindsey et al. 2014). As such, utilising camera trap datasets and the additional data collected on species of conservation concern can be a useful tool for conservation practitioners. Whilst our study provides a small-scale example from one protected area, previous studies have demonstrated that these methods can be applied to camera trap datasets at a national and international level (Scotson et al. 2017a, b; Williams et al. 2021). We advocate the wider use of camera trap bycatch data to investigate ecological questions outside the initial scope of research. However, researchers must ensure that the sampling design and modelling approach undertaken are appropriate for the non-target species and analytical framework being considered (Hardouin et al. 2021). As the use of camera traps continues to expand across Africa (Agha et al. 2018), the possibilities for large-scale collaborations and the use of bycatch data to produce meaningful insights for a wide range of taxa will increase. By improving the ease of data sharing and following best practices for data management, bycatch data can be a useful tool for conservation management on a broad spatial scale (Scotson et al. 2017a, b).
Availability of data and material
The datasets generated and analysed during the current study are available from the corresponding author on reasonable request.
Code availability
The code supporting the current study is available from the corresponding author on request.
References
Abe EL (1994) Behavioural ecology of elephant survivors in Queen Elizabeth National Park, Uganda. Dissertation, University of Cambridge
Abraham JO, Goldberg ER, Botha J, Staver AC (2021) Heterogeneity in African savanna elephant distributions and their impacts on trees in Kruger National Park, South Africa. Ecol Evol 11:5624–5634
Agha M, Batter T, Bolas EC, Collins AC, Gomes da Rocha D, Monteza-Moreno CM, Preckler-Quisquater S, Sollmann R (2018) A review of wildlife camera trapping trends across Africa. Afr J Ecol 56:694–701
Apps P, McNutt JW (2018) Are camera traps fit for purpose? A rigorous, reproducible and realistic test of camera trap performance. Afr J Ecol 56:710–720
Barton K (2020) MuMIn: Multi-Model Inference. Retrieved from: http://cran.r-project.org/web/packages/MuMIn/ index.html
Bastille-Rousseau G, Wall J, Douglas-Hamilton I, Lesowapir B, Loloju B, Mwangi N, Wittemyer G (2020) Landscape-scale habitat response of African elephants shows strong selection for foraging opportunities in a human dominated ecosystem. Ecography 43:149–160
Beale CM, Hauenstein S, Mduma S, Frederick H, Jones T, Bracebridge C, Maliti H, Kija H, Kohi EM (2018) Spatial analysis of aerial survey data reveals correlates of elephant carcasses within a heavily poached ecosystem. Biol Conserv 218:258–267
Bhima R, Howard J, Nyanyale S (2003) The status of elephants in Kasungu National Park, Malawi. Pachyderm 35:31–36
Blake S, Deem SL, Strindberg S, Maisels F, Momont L, Isia IB, Douglas-Hamilton I, Karesh WB, Kock MD (2008) Roadless wilderness area determines forest elephant movements in the Congo Basin. PLoS ONE 3:e3546
Brown JS, Laundré JW, Gurung M (1999) The ecology of fear: optimal foraging, game theory, and trophic interactions. J Mamm 80:385–399
Buij R, McShea WJ, Campbell P, Lee ME, Dallmeier F, Guimondou S, Mackaga L, Guisseougou N, Mboumba S, Hines JE, Nichols JD, Alonso A (2007) Patch-occupancy models indicate human activity as major determinant of forest elephant Loxodonta cyclotis seasonal distribution in an industrial corridor in Gabon. Biol Conserv 135:189–201
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach, 2nd edn. Springer, New York, NY
Caravaggi A, Banks PB, Burton AC, Finlay CM, Haswell PM, Hayward MW, Rowcliffe MJ, Wood MD (2017) A review of camera trapping for conservation behaviour research. Remote Sens Ecol Conserv 3:109–122
Chamaillé-Jammes S, Valeix M, Fritz H (2007) Managing heterogeneity in elephant distribution: interactions between elephant population density and surface-water availability. J Appl Ecol 44:625–633
Chase MJ, Schlossberg S, Griffin CR, Bouché PJC, Djene SW, Elkan PW, Ferreira S, Grossman F, Kohi EM, Landen K, Omondi P, Peltier A, Selier SAJ, Sutcliffe R (2016) Continent-wide survey reveals massive decline in African savannah elephants. PeerJ 4:e2354
Chibeya D, Wood H, Cousins S, Carter K, Nyirenda MA, Maseka H (2021) How do African elephants utilize the landscape during wet season? A habitat connectivity analysis for Sioma Ngwezi landscape in Zambia. Ecol Evol 11:14916–14931
Coverdale TC, Kartzinel TR, Grabowski KL, Shriver RK, Hassan AA, Goheen JR, Palmer TM, Pringle RM (2016) Elephants in the understory: opposing direct and indirect effects of consumption and ecosystem engineering by megaherbivores. Ecol 97:3219–3230
Cusack JJ, Dickman AJ, Rowcliffe JM, Carbone C, Macdonald DW, Coulson T (2015) Random versus game trail-based camera trap placement strategy for monitoring terrestrial mammal communities. PLoS ONE 10:e0126373
Davis RS, Stone EL, Gentle LK, Mgoola W, Uzal A, Yarnell RW (2021a) Spatial partial identity model reveals low densities of leopard and spotted hyaena in a miombo woodland. J Zool 313:43–53
Davis RS, Yarnell RW, Gentle LK, Uzal A, Mgoola W, Stone EL (2021b) Prey availability and intraguild competition regulate the spatiotemporal dynamics of a modified large carnivore guild. Ecol Evol 11:7890–7904
Di Marco M, Venter O, Possingham HP, Watson JE (2018) Changes in human footprint drive changes in species extinction risk. Nat Comms 9:1–9
Di Minin E, Slotow R, Fink C, Bauer H, Packer C (2021) A pan-African spatial assessment of human conflicts with lions and elephants. Nat Comms 12:1–10
Didan K (2015) MOD13Q1 MODIS/Terra Vegetation Indices 16 Day L3 Global 250m SIN Grid V006. NASA EOSDIS Land Proc DAAC. https://doi.org/10.5067/MODIS/MOD13Q1.006
Dimiceli C, Carroll M, Sohlberg R, Kim D, Kelly M, Townshend J (2015) MOD44B MODIS/Terra Vegetation Continuous Fields Yearly L3 Global 250m SIN Grid v006. NASA EOSDIS Land Processes DAAC. Accessed 10/11/2021 from 10.5067/MODIS/MOD44B.006.
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G, García Marquéz JR, Gruber B, Lafourcade B, Leitão PJ, Münkemüller T, McClean C, Osborne PE, Reineking B, Schröder B, Skidmore AK, Zurell D, Lautenbach S (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:27–46
Edwards S, Cooper S, Uiseb K, Hayward M, Wachter B, Melzheimer J (2018) Making the most of by-catch data: assessing the feasibility of utilising non-target camera trap data for occupancy modelling of a large felid. Afr J Ecol 56:885–894
Fiske IJ, Chandler RB (2011) Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. J Statistical Softw 43:1–23
Fitak RR, Johnsen S (2017) Bringing the analysis of animal orientation data full circle: model-based approaches with maximum likelihood. J Exp Biol 220:3878–3882
Food and Agriculture Organization of the United Nations (2019) Country Profiles: Malawi. FAO, Rome, Italy. Available at: https://www.fao.org/countryprofiles/index/en/?iso3=MWI Accessed: [11/02/2022]
Fritz H (2017) Long-term field studies of elephants: understanding the ecology and conservation of a long-lived ecosystem engineer. J Mamm 98:603–611
Gaynor KM, Branco PS, Long RA, Gonçalves DD, Granli PK, Poole JH (2018) Effects of human settlement and roads on diel activity patterns of elephants (Loxodonta africana). Afr J Ecol 56:872–881
Goldenberg SZ, Douglas-Hamilton I, Daballen D, Wittemyer G (2017) Challenges of using behavior to monitor anthropogenic impacts on wildlife: a case study on illegal killing of African elephants. Anim Conserv 20:215–224
Gondwe MF, Cho MA, Chirwa PW, Geldenhuys CJ (2019) Land use land cover change and the comparative impact of co-management and government-management on the forest cover in Malawi (1999–2018). J Land Use Sci 14:281–305
Gunn J, Hawkins D, Barnes RF, Mofulu F, Grant RA, Norton GW (2014) The influence of lunar cycles on crop-raiding elephants; evidence for risk avoidance. Afr J Ecol 52:129–137
Hardouin M, Searle CE, Strampelli P, Smit J, Dickman A, Lobora AL, Rowcliffe JM (2021) Density responses of lesser-studied carnivores to habitat and management strategies in southern Tanzania’s Ruaha-Rungwa landscape. PLoS ONE 16:e0242293
Harris GM, Russell GJ, van Aarde RI, Pimm SL (2008) Rules of habitat use by elephants Loxodonta africana in southern Africa: insights for regional management. Oryx 42:66–75
Hauenstein S, Kshatriya M, Blanc J, Dormann CF, Beale CM (2019) African elephant poaching rates correlate with local poverty, national corruption and global ivory price. Nat Comms 10:1–9
Havmøller RW, Jacobsen NS, Scharff N, Rovero F, Zimmermann F (2020) Assessing the activity pattern overlap among leopards (Panthera pardus), potential prey and competitors in a complex landscape in Tanzania. J Zool 311:175–182
Henschel P, Petracca LS, Hunter LT, Kiki M, Sewadé C, Tehou A, Robinson HS (2016) Determinants of distribution patterns and management needs in a critically endangered lion Panthera leo population. Front Ecol Evol 4:110
Ihwagi FW, Thouless C, Wang T, Skidmore AK, Omondi P, Douglas-Hamilton I (2018) Night-day speed ratio of elephants as indicator of poaching levels. Ecol Indic 84:38–44
International Fund for Animal Welfare (2020) Rehabilitation of Kasungu National Park underway. Retrieved from: https://www.ifaw.org/africa/press-releases/kasungu-national-park-rehabilitation
Jones KR, Venter O, Fuller RA, Allan JR, Maxwell SL, Negret PJ, Watson JE (2018a) One-third of global protected land is under intense human pressure. Science 360:788–791
Jones T, Cusack JJ, Pozo RA, Smit J, Mkuburo L, Baran P, Lobora AL, Mduma S, Foley C (2018b) Age structure as an indicator of poaching pressure: Insights from rapid assessments of elephant populations across space and time. Ecol Indic 88:115–125
Karanth KU, Gopalaswamy AM, Kumar NS, Vaidyanathan S, Nichols JD, MacKenzie DI (2011) Monitoring carnivore populations at the landscape scale: occupancy modelling of tigers from sign surveys. J Appl Ecol 48:1048–1056
Kasozi H, Moll RJ, Kityo RM, Montgomery RA (2022) Phylogeny is a stronger predictor of activity than allometry in an African mammal community. Biol J Linnean Soc 135:599–609
Kerley GI, Landman M (2006) The impacts of elephants on biodiversity in the Eastern Cape Subtropical Thickets: elephant conservation. S Afr J Sci 102:395–402
Kinahan AA, Inge-moller R, Bateman PW, Kotze A, Scantlebury M (2007) Body temperature daily rhythm adaptations in African savanna elephants (Loxodonta africana). Physiol Behav 92:560–565
King LE, Lala F, Nzumu H, Mwambingu E, Douglas-Hamilton I (2017) Beehive fences as a multidimensional conflict-mitigation tool for farmers coexisting with elephants. Conserv Biol 31:743–752
Landler L, Ruxton GD, Malkemper EP (2019) The Hermans-Rasson test as a powerful alternative to the Rayleigh test for circular statistics in biology. BMC Ecol 19:1–8
Lindsey PA, Nyirenda VR, Barnes JI, Becker MS, McRobb R, Tambling CJ, Taylor WA, Watsont’Sas-Rolfes M FG (2014) Underperformance of African protected area networks and the case for new conservation models: insights from Zambia. PLoS ONE 9:e94109
MacKenzie DI, Bailey LL (2004) Assessing the fit of site-occupancy models. J Agric Biol Environ Stat 9:300–318
MacKenzie DI, Nichols JD, Lachman GB, Droege S, Royle JA, Langtimm CA (2002) Estimating site occupancy rates when detection probabilities are less than one. Ecol 83:2248–2255
MacKenzie DI, Nichols JD, Royle JA, Pollock KH, Bailey LL, Hines JE (2017) Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Academic Press, Oxford, UK
Macpherson D (2015) Report on an aerial wildlife census of Kasungu National Park, Malawi, October 2014. Cluny Wildlife Trust, Namitete, Malawi
Macpherson D (2020) Report on an Aerial Wildlife Census of Kasungu National Park, Malawi—September/October 2020. Cluny Wildlife Trust, Namitete, Malawi
Martin J, Chamaillé-Jammes S, Nichols JD, Fritz H, Hines JE, Fonnesbeck CJ, MacKenzie DI, Bailey LL (2010) Simultaneous modeling of habitat suitability, occupancy, and relative abundance: African elephants in Zimbabwe. Ecol Appl 20:1173–1182
McComb K, Shannon G, Sayialel KN, Moss C (2014) Elephants can determine ethnicity, gender, and age from acoustic cues in human voices. PNAS 111:5433–5438
Meredith M, Ridout M (2014) Overview of the overlap package. R. Project. 1–9. Retrieved from: https://cran.r-project.org/web/packages/overl ap/vignettes/overlap.pdf
Meredith M, Ridout M (2016) Package ‘Overlap’—Estimates of Coefficient of Overlapping for Animal Activity Patterns. Retrieved from: https://cran.r-project.org/web/packa ges/overlap/index.html
Mkuburo L, Nahonyo C, Smit J, Jones T, Kohi E (2020) Investigation of the effect of poaching on African elephant (Loxodonta africana) group size and composition in Ruaha National Park. Tanzania Sci African 9:e00490
Munthali SM, Mkanda FX (2002) The plight of Malawi’s wildlife: is trans-location of animals the solution? Biodivers Conserv 11:751–768
Naidoo R, Fisher B, Manica A, Balmford A (2016) Estimating economic losses to tourism in Africa from the illegal killing of elephants. Nat Comms 7:1–9
NEAPM (2015) National elephant action plan for Malawi. Department of National Parks and Wildlife Malawi, Lilongwe, Malawi
Nyhus PJ (2016) Human–wildlife conflict and coexistence. Annu Rev Environ Resour 41:143–171
Oduor S, Brown J, Macharia GM, Boisseau N, Murray S, Obade P (2020) Differing physiological and behavioral responses to anthropogenic factors between resident and non-resident African elephants at Mpala Ranch, Laikipia County. Kenya Peerj 8:e10010
Ogutu JO, Reid RS, Piepho HP, Hobbs NT, Rainy ME, Kruska R, Worden JS, Nyabenge M (2014) Large herbivore responses to surface water and land use in an East African savanna: implications for conservation and human-wildlife conflicts. Biodivers Conserv 23:573–596
Okita-Ouma B, Koskei M, Tiller L, Lala F, King L, Moller R, Amin R, Douglas-Hamilton I (2021) Effectiveness of wildlife underpasses and culverts in connecting elephant habitats: a case study of new railway through Kenya’s Tsavo National Parks. Afr J Ecol 59:624–640
Oksanen J, Guillaume Blanchet F, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) vegan: Community Ecology Package. R package version 2.5–7. https://CRAN.R-project.org/package=vegan
Osipova L, Okello MM, Njumbi SJ, Ngene S, Western D, Hayward MW, Balkenhol N (2018) Fencing solves human-wildlife conflict locally but shifts problems elsewhere: a case study using functional connectivity modelling of the African elephant. J Appl Ecol 55:2673–2684
Pettorelli N, Ryan S, Mueller T, Bunnefeld N, Jędrzejewska B, Lima M, Kausrud K (2011) The Normalized difference vegetation index (NDVI): unforeseen successes in animal ecology. Clim Res 46:15–27
QGIS Development Team (2020) QGIS Geographic Information System. Open-Source Geospatial Foundation. Retrieved from: http://qgis.org
R Core Development Team (2021) R: A Language and Environment for Statistical Computing. Retrieved from: http://www.r-project.org
Ridout MS, Linkie M (2009) Estimating overlap of daily activity patterns from camera trap data. J Agric Biol Environ Stat 14:322–337
Ripple WJ, Newsome TM, Wolf C, Dirzo R, Everatt KT, Galetti M, Hayward MW, Kerley GIH, Levi T, Lindsey PA, Macdonald DW, Malhi Y, Painter LE, Sandom CJ, Terborgh J, van Valkenburgh B (2015) Collapse of the world’s largest herbivores. Sci Adv 1:e1400103
Ripple WJ, Chapron G, López-Bao JV, Durant SM, Macdonald DW, Lindsey PA, Zhang L (2016) Saving the world’s terrestrial megafauna. Bioscience 66:807–812
Rowcliffe JM, Kays R, Kranstauber B, Carbone C, Jansen PA (2014) Quantifying levels of animal activity using camera trap data. Methods Ecol Evol 5:1170–1179
Rowcliffe JM (2019) Package ‘Activity’: Animal Activity Statistics, version 1.2. 2019. Retrieved from: https://cran.r-project.org/web/packages/activity/index.html
Scotson L, Fredriksson G, Ngoprasert D, Wong WM, Fieberg J (2017a) Projecting range-wide sun bear population trends using tree cover and camera-trap bycatch data. PLoS ONE 12:e0185336
Scotson L, Johnston LR, Iannarilli F, Wearn OR, Mohd-Azlan J, Wong WM, Gray TNE, Dinata Y, Suzuki A, Willard CE, Frechette J, Loken B, Steinmetz R, Moßbrucker AM, Clements GR, Fieberg J (2017b) Best practices and software for the management and sharing of camera trap data for small and large scales studies. Remote Sens Ecol Conserv 3:158–172
Shannon G, Page BR, Mackey RL, Duffy KJ, Slotow R (2008) Activity budgets and sexual segregation in African elephants (Loxodonta africana). J Mamm 89:467–476
Sharma K, Fiechter M, George T, Young J, Alexander JS, Bijoor A, Suryawanshi K, Mishra C (2020) Conservation and people: towards an ethical code of conduct for the use of camera traps in wildlife research. Ecol Solut Evidence 1:e12033
Sievert O, Evans K, Mgoola W, Harwood A (2022) Early insights into the movements of Malawi’s transboundary elephants. Afr J Ecol. https://doi.org/10.1111/aje.12984
Suraci JP, Clinchy M, Zanette LY, Wilmers CC (2019) Fear of humans as apex predators has landscape-scale impacts from mountain lions to mice. Ecol Lett 22:1578–1586
Szott ID, Pretorius Y, Ganswindt A, Koyama NF (2019a) Physiological stress response of African elephants to wildlife tourism in Madikwe Game Reserve, South Africa. Wildl Res 47:34–43
Szott ID, Pretorius Y, Koyama NF (2019b) Behavioural changes in African elephants in response to wildlife tourism. J Zool 308:164–174
Thaker M, Gupte PR, Prins HHT, Slotow R, Vanak AT (2019) Fine-scale tracking of ambient temperature and movement reveals shuttling behavior of elephants to water. Front Ecol Evol 7:4
Thouless CR, Dublin HT, Blanc JJ, Skinner DP, Daniel TE, Taylor RD, Maisels F, Frederick HL, Bouché P (2016) African Elephant Status Report 2016: an update from the African Elephant Database. Occasional Paper Series of the IUCN Species Survival Commission (60). IUCN/SSC Africa Elephant Specialist Group.
van Velden JL, Wilson K, Lindsey PA, McCallum H, Moyo BH, Biggs D (2020) Bushmeat hunting and consumption is a pervasive issue in African savannahs: insights from four protected areas in Malawi. Biodivers Conserv 29:1443–1464
Wadey J, Beyer HL, Saaban S, Othman N, Leimgruber P, Campos-Arceiz A (2018) Why did the elephant cross the road? The complex response of wild elephants to a major road in Peninsular Malaysia. Biol Conserv 218:91–98
Wall J, Wittemyer G, Klinkenberg B, LeMay V, Blake S, Strindberg S, Henley M, Vollrath F, Maisels F, Ferwerda J, Douglas-Hamilton I (2021) Human footprint and protected areas shape elephant range across Africa. Curr Biol 31:2437–2445
Warnatzsch EA, Reay DS (2019) Temperature and precipitation change in Malawi: evaluation of CORDEX-Africa climate simulations for climate change impact assessments and adaptation planning. Sci Total Environ 654:378–392
Wevers J, Beenaerts N, Casaer J, Zimmermann F, Artois T, Fattebert J (2021) Modelling species distribution from camera trap by-catch using a scale-optimized occupancy approach. Remote Sens Ecol Conserv 7:534–549
Williams KS, Pitman RT, Mann GK, Whittington-Jones G, Comley J, Williams ST, Hill RA, Balme GA, Parker DM (2021) Utilizing bycatch camera-trap data for broad-scale occupancy and conservation: a case study of the brown hyaena Parahyaena brunnea. Oryx 55:216–226
Wittemyer G, Northrup JM, Blanc J, Douglas-Hamilton I, Omondi P, Burnham KP (2014) Illegal killing for ivory drives global decline in African elephants. Proc Natl Acad Sci 111:13117–13121
World Bank Group (2020) Malawi Overview. World Bank Group, Washington DC, USA. Available at: https://www.worldbank.org/en/country/malawi/overview Accessed: [11/02/2022]
Acknowledgements
This study was conducted in collaboration with Carnivore Research Malawi (CRM), Conservation Research Africa (CRA), and the Department of National Parks and Wildlife Malawi (DNPW). We thank DNPW and the Kasungu National Park management team and scouts for their continued collaboration and support; all the volunteers and staff at CRM for assisting with data collection and field support; and CRM for use of equipment and logistical support. This work was funded by Nottingham Trent University, Conservation Research Africa, Oklahoma Zoo, and Idea Wild. We are grateful to the anonymous reviewers for their valuable feedback.
Funding
Nottingham Trent University,Carnivore Research Malawi,Oklahoma Zoo,Idea Wild
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
Ethical approval was granted by Nottingham Trent University and Department of National Parks and Wildlife Malawi.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling editor: Francesco Ferretti.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Davis, R.S., Gentle, L.K., Mgoola, W.O. et al. Using camera trap bycatch data to assess habitat use and the influence of human activity on African elephants (Loxodonta africana) in Kasungu National Park, Malawi. Mamm Biol 103, 121–132 (2023). https://doi.org/10.1007/s42991-022-00330-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42991-022-00330-7