
Abstract
Combined application of biochar with fertilizers has been used to increase soil fertility and crop yield. However, the coupling mechanisms through which biochar improves crop yield at field scale and the time span over which biochar affects carbon and nitrogen transformation and crop yield are still little known. In this study, a long-term field trial (2013–2019) was performed in brown soil planting maize. Six treatments were designed: CK—control; NPK—application of chemical fertilizers; C1PK—low biochar without nitrogen fertilizer; C1NPK, C2NPK and C3NPK—biochar at 1.5, 3 and 6 t ha−1, respectively, combined with chemical fertilizers. Results showed that the δ15N value in the topsoil of 0–20 cm layer in the C3NPK treatment reached a peak of 291 ‰ at the third year (2018), and demonstrated a peak of 402 ‰ in the NPK treatment in the initial isotope trial in 2016. Synchronously, SOC was not affected until the third to fourth year after biochar addition, and resulted in a significant increase in total N of 2.4 kg N ha−1 in 2019 in C3NPK treatment. During the entire experiment, the 15N recovery rates of 74–80% were observed highest in the C2NPK and C3NPK treatments, resulting in an annual increase in yields significantly. The lowest subsoil δ15N values ranged from 66‰ to 107‰, and the 15N residual rate would take 70 years for a complete decay to 0.001% in the C3NPK. Our findings suggest that biochar compound fertilizers can increase C stability and N retention in soil and improve N uptake by maize, while the loss of N was minimized. Biochars, therefore, may have an important potential for improving the agroecosystem and ecological balance.
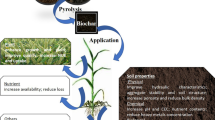
Graphic abstract

Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Nitrogen (N) is an essential element for plant growth in terrestrial ecosystems. The N cycle in soil is modulated by microorganisms decomposing soil organic matter (SOM), with a potential to synchronize N supply and plant N demand (Liang et al. 2013a, b). During this process, more energy substance with high C/N ratios lead to an increase in N immobilization and a decrease in N mineralization (Lehmann et al. 2003), for example, soil dissolved organic carbon (DOC) undergoes fast turnover; whereas, relatively stable pyrogenic carbon and aromatic hydrocarbon (PyC) have slow turnover (Pan et al. 2019). Initially, the bioavailable N (NH4+/NO3−) seems to be decreased (Kopacek et al. 2013). However, the microbes in soil preferentially decompose fresh plant litter over “old” SOM (Cui et al. 2017; Liang et al. 2017), thus slowing down the rate of SOM turnover as a whole (Pan et al. 2019). In other words, compared to the ‘fresh’ organic matter, the ‘old’ SOM may provide a more stable and persistent N source for plant uptake, thus avoiding losses, especially in the presence of biochar (Cui et al. 2017). Although the effect of aging process of biochar on SOM may indirectly affect the nitrogen cycle, however, detailed investigation of the coupling of soil/biochar-carbon and nitrogen is still lacking; their interactive mechanisms are virtually unknown, and the regulatory role of biochar in the interaction process is not fully elucidated in the presence or absence of plants (Weng et al. 2015).
Biochar effectively holds soil nutrients and promotes nutrient absorption in crops by stimulating microbial and enzyme activities. Biochar has well-developed pore structure and high biochemical stability (Han et al. 2020), as well as high sorption and redox capacities caused by its huge specific surface area, thus participating in the competition of mineral surface sites (Shi et al. 2019; Wang et al. 2020b). Additionally, biochar also plays a regulatory role in activating or inactivating nutrients, hormones or toxins by affecting alkaline functional groups (Novak et al. 2019). Lehmann et al. (2003) reported that the indirect effects of biochar on soil amendment and nutrient cycling are more significant than the direct effects of biochar itself, similarly to SOM which is considered to be the micro-domain framework coordinating the balance between solid, liquid and gaseous phases in soil (Pan et al. 2019). However, the biochar addition alone has few significant yield-improving effects (Li et al. 2019).
To promote a modern and sustainable agricultural/environmental technology inspired by an ancient practice, Chen et al. (2019) put forward the concept of “Straw Biochar Returning” and developed the application technology in the whole agriculture-industry chain. In the past decade, combining biochar and various fertilizers, such as compound fertilizers, organic fertilizers and bio-fertilizers, to improve plant growth and agricultural production attracted substantial interest (Jones et al. 2012; Yu et al. 2018; Chen et al. 2019). The commercial production of biochar–fertilizer mixtures has been achieved in China (Wang et al. 2018b), and some long-term field trial studies have been reported (Madari et al. 2017; Yu et al. 2018; Li et al. 2019, 2020b). Nevertheless, most of the previous studies were conducted in short term (< 12 months) by pot experiments (Song et al. 2018) and simulation experiments (He et al. 2020) with different soil types, crop species and biochar properties, application rates and methods (Liu et al. 2019; Wu et al. 2019), thus leading to some controversial conclusions regarding its positive and negative effects on crop yield, carbon and nitrogen priming (Mukherjee et al. 2016a; Liu et al. 2019). There are no detailed theoretical studies on the field spatio-temporal scale at present (Meng et al. 2019).
In this study, the arable brown soils from the long-term field experiment sites were used to explore the cycling of N and C by labeled biochar-13C and fertilizer-15N. The biochar was applied with the same amount of maize straw return annually. Based on the 7-year field experiment with successive biochar application combined with N fertilizer, it was hypothesized that the synergistic effects of biochar were demonstrated by increasing N retention in soil and N uptake by maize, while the loss of N was minimized. This study provides a theoretical basis for applying new eco-friendly biochar-based fertilizer for improving the agroecosystem to achieve both green agriculture and ecological balance.
2 Materials and methods
2.1 Field experimental site
The long-term fertilizer trial on brown soil was located at the experimental station (40°48′ N and 123°33′ E) in a semi-humid region of Shenhe district, Shenyang of Liaoning Province, Northeast China. The site has the typical continental monsoon climate with mean annual temperature of 7.0–8.1 ℃, precipitation of 574–684 mm and the frost-free period of 147–180 days. The tested soil belongs to brown earth of Alfisols derived from loess with the soil texture of clay loam (48% sand, 29% silt and 23% clay), with 9.87 g kg−1 total SOC, 0.90 g kg−1 total N, 112.65 mg kg−1 alkali hydrolysable N, 16.30 mg kg−1 available P, 109.90 mg kg−1 available K, 14.34 cmol kg−1 cation exchange capacity (CEC), pH (H2O) 6.00, 1.30 g cm−3 bulk density, 49% soil total porosity, and 27% field capacity at 0–20 cm depth in 2013 (Han et al. 2017).
2.2 Treatments
This experiment is a part of a long-term field trial established in April 2013 using a randomized block design. The experiment includes fifteen different treatments in three replicates, with each plot measuring 3.6 m × 7.0 m (25.5 m2). In this study, six treatments were chosen (Fig. 1a, Table 1): CK—the treatment without application of biochar and fertilizer; NPK—the treatment with application of nitrogen–phosphorus–potassium using urea (N 46.3%), single superphosphate (P2O5 16%), and muriate of potash (K2O 60%) as chemical fertilizers; C1PK—the treatment with low rate of biochar (1.5 t ha−1), phosphorus and potassium (without nitrogen); and the three treatments, C1NPK, C2NPK and C3NPK, with low biochar at 1.5 t ha−1, moderate biochar at 3 t ha−1 and high biochar at 6 t ha−1, respectively, and combined the same amounts of mineral fertilizer. The biochar and fertilizer were applied annually (from 2013 to 2019) and mixed well with the topsoil (0–20 cm layer) by a rotavator before sowing. Maize (Zea mays L.) cultivar Dongdan 6531 was sown on April 25 and harvested on October 1 annually. As shown in Table 1, the biochar applied at 1.5, 3 and 6 t ha−1 is equal to the amounts of 4.5, 9 and 18 t ha−1 of maize straw returned annually. The biochar was made by pyrolysing maize straw of less than 2 cm in length and heating at 450 ℃ over 30 min, with 490.0 g kg−1 total C, 14.4 g kg−1 total N, 8.5 g kg−1 total P, 32.0 g kg−1 total K, pH (H2O) 10.4, 26.9 m2 g−1 BET surface area, 0.0425 cm3 g−1 pore volume, and 7.1 nm mean pore size (Han et al. 2017).

The field distribution of experiment treatments (2013) (a) and isotope experiments (2016) (b, c): e size dimension of elevation view and top view of PVC (polyvinyl chloride) bottomless box, g, h soil digging and entombing hierarchically, f applying biochar and chemical fertilizers in the ratio of per hectare, and mixing well with the topsoil (0–20 cm) by hand, d biochar–soil ‘co-aggregation’ phenomenon during 7 years successive application and tillage
The isotope experiments (Fig. 1b–h) were established in 2016. A set of PVC (polyvinyl chloride) bottomless boxes were fixed on the ridging of chosen plots before seeding, the area of PVC micro-plots was 0.333 m2 (60 cm × 55 cm) with a burial depth of 50 cm. The experimental plots were supplied with urea (15N atom% = 10.16%) at the same relative rate as in the whole experiment, which was mixed well with the topsoil (0–20 cm) by hand, then planting two maize seeds in each hole was performed to ensure only two seedlings remained in each PVC plot in which the same amounts of unlabeled N and unlabeled biochar had been applied every year since 2017 and 2018 (Table 1). To allow field operations, the plant space and ridge space of 27 cm × 60 cm were kept both inside and outside of PVC plots (Fig. 1e). The biochar (13C atom% = 1.20%) derived from 13C-labeled maize straw pyrolysed at 450 ℃ was added to the C1PK and C1NPK micro-plots in 2017, as described in supporting information (Table S1).
2.3 Sampling and analyses
For each harvest from 2013 to 2019, the aboveground maize plants were harvested from both inside and outside of PVC plots, and only the maize stubble remained in field. After weighing, a few root branches were selected to determine the isotope ratio before returning to the PVC plots. The aboveground plants were dissected into stem, leaf, grain, and corncob, and then dried in an oven at 60 ℃ for 72 h. We did not weigh the root and corncob in 2016 due to sample losses. The isotope soil samples and bulk density in the 0–20 cm and 20–40 cm depths were collected carefully to avoid contamination and dried at room temperature. Visible biochar, plant debris and roots were hand-picked and removed from all samples by the same researcher to avoid a bias in data, and all samples were ground by a ball mill and passed through 0.15 mm mesh. Isotope abundance ratios and the contents of total N and C in both soil and plant materials were determined by isotope ratio mass spectrometry coupled with an elemental analyzer (EA-IRMS, Germany).
Total N and total C concentration in plant and soil materials, δ15N/δ13C values, and mass data of plant and soil were used to calculate the annual 15N tracer retention and the turnover rate constant in different sources of carbon using the following equations:
where δ‰ represents the isotope abundance values expressed as δ‰13C or δ‰15N relative to, respectively, Pee Dee Belemnite (PDB) (Rst = 0.0112372) or the atmospheric air nitrogen (Rst = 0.0036765), Atom% means the percentage of rare isotope with respect to the total amount of the element, Rsa and Rst are the ratios of rare isotope versus prevalent isotope (13C/12C or 15N/14N) in the sample (Rsa) and the standard materials (Rst).
where 15Nrec is both the nitrogen recovery rate (%) in maize plant and the nitrogen residue rate (%) in soil; \({\text{Sample}}\left( {N \times {\text{Atom\% excess}}} \right)\) is the product of multiply nitrogen content by atom percent excess in sample depending on the plant biomass and the soil bulk density; \({\text{Fertilizer}}\left( {N \times {\text{Atom\% excess}}} \right)\) is the product of multiply 15N-urea fertilizer application amount by atom percent excess.
where f is the percentage (%) of exogenous carbon in any soil organic fraction after a duration of the labeled experiment; δend is the δ13C value in the soil samples at the final experiment stage; δinitial is the δ13C value in the soil samples without exogenous carbon addition at the beginning of the experiment; δinput is the δ13C value in the applied biochar.
where k (year−1) denotes the turnover rate constant of the carbon fractions, t indicates the labeled experiment duration (year), and the reciprocal of k represents the mean residence time (MRT) (year) (Dorodnikov et al. 2011).
Nitrogen retention rate was fitted to a simple exponential decay function, where y is the 15N retention rate (%) of the initial 15N input of 630.7 mg per plot (0.333 m2) at time x, N0 denotes the initial maximum retention rate, λ is the exponential decay rate constant (year−1), and r is the mean residence time (year).
2.4 Statistical analysis
One-way ANOVA and the Duncan’s multiple range tests were performed to observe the variation in δ15N/δ13C values, 15N recovery rates and crop yields in different treatments and inter-annually. The independent-sample t-test comparison was conducted to compare the turnover rate constant of carbon fractions in biochar application with or without N fertilizer. All analyses were performed using SPSS Statistics version 19.0 (IBM Corporation, Armonk, New York, USA.). The figures were drawn by OriginPro 2018 software (Origin Lab Corporation, Northampton, Massachusetts, USA.). The significant differences were defined at α = 0.05.
3 Results and discussion
3.1 Improved N-use efficiency and retention
Generally, exogenous glucose application to soil stimulates microbial activity and promotes N immobilization (Fiorentino et al. 2019). However, in this study, the immobilization of fertilizer-supplied N as well as the unimodal peak were delayed in the topsoil of the C3NPK treatment (Fig. 3). In addition, C3NPK improved accumulation of labeled and non-labeled N in harvested maize in 2016 (initial trial stage) and also increased maize yield every year (Table 3a, Fig. 2d). Given a large amount of N absorbed by crops (Fig. 2 and Table 3a), we recorded decreased residual soil inorganic N (SIN) in 2015 (Han et al. 2017) and also decreased 15N immobilization in 2016 (Fig. 3). These were also similar to the results reported by Li et al. (2019); however, the findings presented here appear to be different from the explanations of Song et al. (2018). Fiorentino et al. (2019) also thought that biochar might partially buffer the immobilization of inorganic N derived from both native soil and fertilizer. Hence, biochar could counteract the immobilization of NH4+ caused by the increased microbial activity. Simultaneously, the N immobilization rates were inhibited by plants (He et al. 2020); the loss of microbial necromass N was also a major source to ensure the priority use of N for crop absorption (Wang et al. 2020a). However, this phenomena of offset effect disappeared with the extension of time because the C2NPK and C3NPK treatments significantly improved N retention in the fourth corn planting season (Figs. 3, 5a) (P < 0.05). The peak of the curve of C2NPK and C1NPK was, respectively, in 2016 and 2017 because of the relatively smaller delay or buffer capacity compared to the C3NPK treatment. Additionally, the C2NPK treatment increased total N retention in subsoil from 2018 but no significant difference was observed (Fig. 3) (P > 0.05). This is of great significance to the nitrogen supply to crops from the native soil nitrogen pool (Table 3a) that is supplemented every year by exogenous nitrogen. What we found was similar to the “revisited mental model”- the importance of soil and crop residue N turnover (Yan et al. 2020). Our data also showed a short-lived N enrichment effect in different organs of maize in 2 years (2016–2017), but this effect gradually faded in subsequent years, becoming non-significant in leaves and grain in 2018 (Fig. 2a–c).

δ15N mean values of root, stem, leaf, grain, corncob and grain yields of maize in different years. The small letters denote significant differences at the 5% level, ns, no significant difference

δ15N mean values of two layers of soil all sampled at every harvest season from 2016 to 2019. The soil background value was sampled in 2015 before the isotope experiment establish
Just as the result showed above, high rate of biochar buffered the inorganic N immobilization by microbes at the beginning of the experiment, but ultimately, biochar resulted in increased nitrogen retention in soil (Fig. 3). Our analyses suggest a possible mechanism reliant on a transient sorptive capacity of biochar increasing maize uptake of N from the fertilizer and native soil pools (Table 3a). This result was consistent with the field trials with grasses, but different from the maize pot trials conducted by Jones et al. (2012), who reported severe negative effects on maize (Jones et al. 2012) and wheat growth (Li et al. 2019) with excessive addition of biochar at 50 (maize) and 40 t ha−1 (wheat). On the other hand, application of biochar carrying electron transfer sites (Kopacek et al. 2013), high C:N ratio (Table 2) and active energy substrate (labile biochar-C components) may stimulate growth of microorganisms that can degrade recalcitrant SOM, thus acquiring additional N when readily available N in the soil is relatively low (Nelissen et al. 2012). However, the transient stimulation was dominated by the provision of favorable microbial habitats (Weng et al. 2017; Zheng et al. 2018), and resulted in stable C and N accumulation in the microbial biomass through entombing effect (Liang et al. 2017, 2019; Wang et al. 2020a). This transition effect (associated with biochar aging) lasted more than 3 years according to our study result, which was in accordance with the investigation of field-aged biochar spatial niches (Quilliam et al. 2013). Surprisingly, biochar applied at 10 t ha−1 only once in 2012 showed a delayed, yet short-lived effect, in 2 years, but this effect gradually faded in subsequent years (Griffin et al. 2017). Annually applied biochar at lower rates (< 6 t ha−1 year−1) in our study and also in the report by Nan et al. (2020) suggested applying biochar in alternate years would be a more cost-effective strategy (Wang et al. 2018b), but this suggestion would need to be tested in the further studies.
In this study, it is noteworthy that the effect of favorable nitrogen regulation was not triggered by the biochar nitrogen, but was modulated by the combined application of biochar and N fertilizer. Despite the high total nitrogen content in biochar produced from wheat straw (Fiorentino et al. 2019) or maize straw (Table S1), the available nitrogen is likely to be tiny (Fiorentino et al. 2019), and the amounts of N adsorption by both fresh and field-aged woody biochar were relatively small (Jones et al. 2012). As shown in our previous study (Han et al. 2017) and in Fig. S1–S5, whether we calculate biochar nitrogen or not, combined application of biochar improved N-use efficiency compared to the control, suggesting that the contribution of nitrogen from biochar to the crop uptake might be almost negligible. Therefore, the indirect effects of biochar on soil amendment and nutrient cycling would be of great importance (Lehmann et al. 2003). A recent local-scale study also showed that the physico-chemical properties of soil might be more important than microbial enzyme activity in controlling C and N pools (Li et al. 2020a). Thus, we speculated that the stability, resilience and bio-availability (accessibility) of SOM were important in synchronization between plant growth and soil fertility.
3.2 New carbon input and SOM turnover
In a long-term experiment, the dominant factors influencing biochar stability are the environmental variables such as water availability and temperature together with biotic factors such as roots and microorganisms. The abiotic oxidation is a requisite process for biological oxidation (e.g., mineralization of SOC), which then undergoes the process of biological mineralization, termed ‘New Regulatory Gate Hypothesis’ by Brookes et al. (2017). In contrast to the climate parameters of the study site in Major et al. (2010), our study region has low annual precipitation and high silt and clay content in subsoil (An et al. 2015). Therefore, the erosion and leaching rates in this study were lower (also meaning that any biochar loss was minimal or absent), especially in the PVC micro-plots with border barrier of 10 cm in height; hence, biochar mineralization would not be underestimated as reported by Jiang et al. (2016). Consequently, 7–11% of total CO2 respired originated from biochar mineralization under the N addition (Jiang et al. 2016).
Our study clearly showed that compared to the control, the turnover rate constant of biochar-C (k1) was significantly decreased when N was added in the first year (2017) (Table 3b) (*, P < 0.05). In addition, the native soil C turnover rate constant of k2 (C1PK) was less than k2 (C1NPK) (Table 3b) (*, P < 0.05), and the total soil C content in the NPK and C1NPK treatments significantly decreased (P < 0.05), especially in subsoil (20–40 cm) (Table 2). These findings suggested that the treatment with N addition stimulated the mineralization of the native soil carbon (Tables 2, 3b), although field-aged biochar lowered SOC mineralization by 5.5% (Weng et al., 2017). The aging process influences decomposition and mineralization of biochar, thus adding biochar might be a quick and direct way to improve SOM (Haumaier and Zech 1995; Chen et al. 2019; Han et al. 2020). However, some studies did not show any significant changes in total soil C (Neff et al. 2002), and even reported a decrease in the amount of fine roots (Kopacek et al. 2013), although N addition increased aboveground biomass of crop. This report was similar to our investigation, whereby the total soil C concentration decreased in topsoil, and was lowered by 50% in subsoil (Table 2) (P < 0.05), although both the aboveground and root biomass increased (data not shown) compared to the plots without N addition. Nevertheless, the high addition of biochar prevented a loss of the total carbon in the topsoil, and SOC was not affected until the third; to fourth year (Table 2), which were similar to the results of the five-years field trial (Madari et al. 2017), although the subsoil C was still very low (Table 2), which was related to slow replenishment.
The transient stimulation effects from the microbial activity have been reported (Luo et al. 2017; Nguyen et al. 2018). Here, we proposed a hypothesis that a possible reason was the ecological niches substitution effect (Fig. 4) from direct physical mixing of biochar with soil (Han et al. 2020), as previously reported in biochar’s spatial niches metabolism (Quilliam et al. 2013) and ‘new microbiological niche’ (Luo et al. 2017). The potential readily mineralized carbon was actually generated from the ‘young’ native soil carbon derived from lignin and cellulose (Neff et al. 2002). The lignin-rich biochar (Major et al. 2010) under low pyrolysing temperature has a high similarity to native soil organic matter components (Haumaier and Zech 1995), potentially resulting in substitution/replacement of the ‘young’ C from SOM pool by aging biochar particles containing microbial residue C and N (Zheng et al. 2018), and thus protecting the soil silt–clay aggregates as reported in our previous study (Chen et al. 2020). Biochar was more beneficial to SOC accumulation compared to maize straw application (Chen et al. 2020), depending on the physical and chemical protection (Dorodnikov et al. 2011; Weng et al. 2017; Zheng et al. 2018; Han et al. 2020). In our study (Table 3b), N addition lowered the k values of biochar-C (*, P < 0.05), but there was no difference in topsoil in the second year (*, P > 0.05). However, the biochar particles in subsoil demonstrated a strong stability and slowly responded to the exogenous N due to the ‘co-aggregation’ effect (Fig. 4), implying that the biochar aging in subsoil with a high clay content and oxidation resistance was slower (**, P < 0.01). In contrast, the residual biochar in the topsoil was much easier to oxidize and accelerate humification under a long-term exposure in the presence of oxygen and adequate temperature and nutrient supply. In the BC-rich Amazon Anthrosol or Terra Preta de Indios (TPI) soil (Liang et al. 2013a, b), the oxidation of external surface of BC particles in subsoil layer was inevitably increased over millennia. In the present study, after a short period of 3 years for stabilization of soil microbiome, an enhanced increase in soil C with biochar aging might indicate a long-term dominant C storage mechanism, similarly to the result that 50% of the initial C would be sequestered (Nan et al. 2020), which was faster than the biological decomposition pathway dependent on the microbial entombing effect (< 10–20% of the initial C after 5–10 years) (Lehmann et al. 2003; Liang et al. 2017).

Conceptual view of fertilizer-N (urea) and biochar combined application inducing the turnover of SOC and N accessibility to crop uptake (nitrogen supply ‘capacity’), increasing the soil C stability and N retention ‘capacity’, and reducing nitrogen loss ‘capacity’. In addition, the low rate (< 6 t ha−1 year−1) biochar application strategy will have a ‘capacity’ of cost reduction in future. Note that the ‘four capacity’ above-mentioned is the meanings of ‘four win’ for agricultural promotion; number 1, 3, and 4 was an example of the large aggregates (> 2 mm), micro-aggregates (0.053–0.25 mm), and silt–clay aggregates (< 0.053 mm) from topsoil, respectively number 2 was the magnification view of small aggregates (0.25–2 mm); number 5 of red particles was an example of microorganism colonization; number 6 was the magnification view of ‘co-aggregation’ involving different sized soil aggregates–microbes (Fungi/Bacteria)–biochar–roots; The red (potential and actual events denoted by dotted and solid arrows) and blue arrows denote available C, N and necromass C, N, released from and immobilized to the SOM, respectively. NPK mineral fertilizer for nitrogen–phosphorus–potassium, F fungi, B bacteria, DOC dissolved organic carbon, SIN soil inorganic nitrogen, SOC soil organic carbon
During the long-term alteration and ecological selection as reported by Feng et al. (2020), the SOC loss is unlikely to subside after 5 years because microbial community was restructured fundamentally (Feng et al. 2020). It remains to be investigated whether these effects promote the new microbial colonization, and better explain the adaptive mechanism to utilize fresh C and N. However, based on the existing observations, we surmised there is an ecological niches substitution effect in the presence of biochar, although the quantitative aspects of microbial shifts (e.g., fungi/bacteria or K‐/r‐strategists) as influenced by the interactions between abiotic and biotic factors remain largely unknown. Similarly, the thresholds associated with N limitation and C limitation regulating the processes in the biochar-amended dryland cropping systems have yet to be ascertained. Based on our general conceptual hypothesis presented in Fig. 4, there is a dynamic balance effect of abiotic and biotic ‘Regulation Gate’ switching to regulate the acquisition of available C and N, and the nutrient limitation may likewise depend on the ratio of fungi versus bacteria (F/B). Both of these effects are dominated by the quantity (concentration) and quality (C:N) of the nutritional input (Qiu et al. 2016) and energy (Liang et al. 2020), and also regulated by the temporal and spatial dynamics of soil microbial activity (Brookes et al. 2017) in dependence on the biodiversity and homeostasis maintenance (Yu et al. 2018). As previously reported in a 10-year field trial, B9+1NPK had the same α-diversity indices as the B9NPK and NPK treatments (Nguyen et al. 2018). Wang et al. (2020a) also reported that the necromass C and N of bacteria, fungi and actinomycetes showed a similar pattern of decomposition and stabilization, contributing significantly to the SOM pool, as influenced by soil spatial heterogeneity (Yu et al. 2018), land use intensity (Liang et al. 2017) and nitrogen additions (Neff et al. 2002). Moreover, F/B ratio may have either positive (Oladele et al. 2019) or negative (Song et al. 2018; Yu et al. 2018) responses to biochar addition, and further work is required.
3.3 Dynamics of biochar effects is associated with increased N bio-availability and stability as well as minimized losses
As shown in Table 3, after isotope trial for 1 year, N addition increased the turnover rate of the native soil C, while biochar addition increased the bio-availability of fertilizer-N and native soil N. Indeed, biochar addition initially induced positive priming in our study and the other reports. For instance, the soil CO2 fluxes were maximal after rice seeding (Oladele et al. 2019), triggering positive priming in the period 0–62 days, but turning negative afterwards (62–388 days) (Weng et al. 2015). This also may result from the ‘co-metabolism’ of carbon and nitrogen, thus improving N synchronization, and increasing nutrient use efficiency in maize growth. As shown in our study, compared to the NPK and C1NPK treatments, the N accumulation amounts (Table 3a) and the annual plant recovery rates of 15N in the C2NPK and C3NPK were significantly higher (P < 0.05).
Arguably, this interaction of biochar and C/N is influenced not only by the chemical factors (Fiorentino et al. 2019), but physical factors as well because gross desorption rates would be very low when NH4+ and NO3− were bound strongly to the (field-aged) biochar (Mukherjee et al. 2016b, c; Li et al. 2019), and the reactive sites were blocked over time (Durenkamp et al. 2010). Therefore, we inferred that the balance between the microbial priming effect (PE) and entombing effect (EE) as well as ecological niches substitution would regulate the stability and the size of soil C (Liang et al. 2017) and N pools (Liang et al. 2019; Wang et al. 2020a). After 2 years of the isotope trial (Table 3b), the turnover rates of carbon from both native soil and biochar decreased and showed a low response to the exogenous N addition, suggesting that C together with N was protected in the soil aggregates (‘co-aggregation’) through ecological niches substitution (Fig. 4) and in the microbial entombing effect (Liang et al. 2017). These findings also suggested formation of new organic matter, especially in the presence of high biochar application rate (6 t ha−1). With prolonged biochar aging (after 3 years) (Fig. 3), the SOM reached a new equilibrium between the immobilization and mineralization, resulting in more retention in soil of N derived from fertilizer and crop residues (Fig. 2b, c and Fig. 3). These findings were likely associated with increasing microbial biomass without altering microbial community structure (Yu et al. 2018). Griffin et al. (2017) also reported that the potential mineralizable N may decrease with biochar field aging after 3 years. However, compared to the ‘fresh’ organic matter, the ‘old’ SOM may provide a more stable and persistent N source for plant uptake, thus avoiding losses, especially in the presence of biochar (Cui et al. 2017). As shown in our study, the highest recovery rates of N in the C2NPK treatment ranged from 74 to 80% (Fig. 5a), but non-significantly compared to other treatments in first two years. However in the third year (2018), the total recovery rate of N increased significantly by 77% (P < 0.05) at 3 and 6 t ha−1 biochar applications because of a low risk of N losses compared to the C1NPK and NPK treatments. Similarly, ecosystem switch (Liang et al. 2019) from intense agriculture to moderate forest was accompanied by increased amounts of SOC through the ‘ex vivo’ pathway (Liang et al. 2017, 2019) and the greater abiotic factor contribution. Our investigation suggested that the C3NPK was associated with an increase in net C and N storage by possibly enhancing the effect of niches substitution greater than the sum of PE plus EE (Fig. 4), which had synergies in improving soil fertility and increasing maize grain yield.

a Accumulative recovery rate of plants-soil systems (0–40 cm) and their total values. b Decay functions fitted to observed δ15N values of soil (0–40 cm) under four gradient biochar application. The model suggests that it will take about 70 years to tend to the background δ15N values of + 6.4‰ and + 11.7‰ (showed in Fig. 2, but estimated 0.001% of the initial 15N input here) in topsoil and subsoil, respectively, observed before tracer application
According to the previous study, the combined application of biochar with N significantly decreased the N-loss risk because of strong uptake, the ratio of UNH4 (uptake rate)/INH4 (immobilization rate) reached 374 in the presence of crops (He et al. 2020), thus supporting our findings (Figs. 2, 5a, and Table 3a) that biochar addition increased bio-availability of N for plant uptake. In this study, we used a simple exponential decay function fitted to the isotope data to show the rates of residual 15N in soil (Fig. 5b). In the case of the C3NPK treatment, it was predicted that decaying to the background δ15N values of + 6.4‰ and + 11.7‰ as shown in Fig. 2 would take about 70 years (Fig. 5b), depending on the measured data of soil 15N in the topsoil and subsoil before tracer application (the total residual rate of decaying to 0, which was estimated by 0.001% here). These findings were similar to the previous reports (Sebilo et al. 2013) based on tracing the fate of nitrate fertilizer after three decades of sole N application (the average annual fertilization amount of 135 kg N ha−1 year−1). This is likely caused by excessive nitrogen application (increasing the N-loss risk), and by the negative effects on C sequestration because the balance is tilted in favor of microbial priming effect and ‘N saturation’ (Kopacek et al. 2013), suggesting that it might decrease N losses or buffer N saturation in the presence of biochar. With an increase in biochar application rates, there was a significant decrease in decaying constant λ from the NPK to the C3NPK treatment (Fig. 5b) (from 1/2.26 to 1/7.81) (P < 0.05), revealing another direct evidence of decreasing the risk of N losses. Additionally, the 15N content in subsoil (20–40 cm layer) in the C3NPK treatment was also very low during the entire experiment because of the low leaching of N (Fig. 3), which was the same as the results in the previous report (Li et al. 2019; Lehmann et al. 2003), with the reduced leaching risk after several millennia black carbon being aged in archeological Anthrosols from Central Amazonia, the nutrient availability was higher.
4 Conclusions
Using the long-term field experiment by labeled biochar-13C and fertilizer-15N, we found that the high rate of biochar application (6 t ha−1 year−1) prevented the loss of total C in topsoil and buffered the immobilization of inorganic N derived both from native soil and fertilizer. As a consequence of these long-term positive effects, there were an annual increase in the maize yields significantly. The synergistic effects of biochar were confirmed by increasing C stability and N retention in soil and improving N uptake by maize, while the loss of N was minimized. Our investigation indicates a fact that the long-term effect of the biochar’s stimulation and retention cannot be observed in the short-term already reported works. Given that the greater significance of carbon–nitrogen coupling effects on soil organic matter than a single element, we suggest that the detailed study are still needed in future, and better explain the adaptive mechanism of microbial colonization in soil/biochar-carbon and nitrogen coupling system especially in the presence of plants. Biochars, therefore, should be specifically designed or selected for a more cost-effective strategy than overuse to bring out its potential.
Data availability
The original data can be obtained from the authors upon reasonable request. And the supplementary material related to this article can be found online.
References
An TT, Schaeffer S, Zhuang J, Radosevich M, Li S, Li H, Pei J, Wang JK (2015) Dynamics and distribution of 13C-labeled straw carbon by microorganisms as affected by soil fertility levels in the Black Soil region of Northeast China. Biol Fertil Soils 51:605–613. https://doi.org/10.1007/s00374-015-1006-3
Brookes P, Chen Y, Chen L, Qiu G, Luo Y, Xu J (2017) Is the rate of mineralization of soil organic carbon under microbiological control? Soil Biol Biochem. https://doi.org/10.1016/j.soilbio.2017.05.003
Chen WF, Meng J, Han XR, Lan Y, Zhang WM (2019) Past, present, and future of biochar. Biochar 1:75–87. https://doi.org/10.1007/s42773-019-00008-3
Chen K, Peng J, Li J, Yang Q, Zhan X, Liu N, Han X (2020) Stabilization of soil aggregate and organic matter under the application of three organic resources and biochar-based compound fertilizer. J Soils Sediments. https://doi.org/10.1007/s11368-020-02693-1
Cui J, Ge T, Kuzyakov Y, Nie M, Fang C, Tang B, Zhou C (2017) Interactions between biochar and litter priming: A three-source 14C and δ13C partitioning study. Soil Biol Biochem 104:49–58. https://doi.org/10.1016/j.soilbio.2016.10.014
Dorodnikov M, Kuzyakov Y, Fangmeier A, Wiesenberg GLB (2011) C and N in soil organic matter density fractions under elevated atmospheric CO2: turnover vs. stabilization. Soil Biol Biochem 43:579–589. https://doi.org/10.1016/j.soilbio.2010.11.026
Durenkamp M, Luo Y, Brookes PC (2010) Impact of black carbon addition to soil on the determination of soil microbial biomass by fumigation extraction. Soil Biol Biochem 42:2026–2029. https://doi.org/10.1016/j.soilbio.2010.07.016
Feng JJ, Wang C, Lei J, Yang Y, Yan Q, Zhou X, Tao X, Ning D, Yuan M, Qin Y, Shi Z, Guo X, He Z, Van Nostrand J, Wu L, Bracho-Garillo R, Penton C, Coles JR, Konstantinidis K, Zhou J (2020) Warming-induced permafrost thaw exacerbates tundra soil carbon decomposition mediated by microbial community. Microbiome. https://doi.org/10.1186/s40168-019-0778-3
Fiorentino N, Sánchez-Monedero MA, Lehmann J, Enders A, Fagnano M, Cayuela ML (2019) Interactive priming of soil N transformations from combining biochar and urea inputs: a 15N isotope tracer study. Soil Biol Biochem 131:166–175. https://doi.org/10.1016/j.soilbio.2019.01.005
Griffin DE, Wang D, Parikh SJ, Scow KM (2017) Short-lived effects of walnut shell biochar on soils and crop yields in a long-term field experiment. Agr Ecosyst Environ 236:21–29. https://doi.org/10.1016/j.agee.2016.11.002
Han XR, Ge YF, Li N, Peng J, Fan Y, Gao TY (2017) Effects of continuous application of biochar on soil physic-chemical properties and nitrogen use efficiency. J Shenyang Agric Univ 48:392–398. http://en.cnki.com.cn/Article_en/CJFDTotal-SYNY201704002.htm (in Chinese with English abstract)
Han LF, Sun K, Yang Y, Xia X, Li F, Yang Z, Xing B (2020) Biochar’s stability and effect on the content, composition and turnover of soil organic carbon. Geoderma 364:114184. https://doi.org/10.1016/j.geoderma.2020.114184
Haumaier L, Zech W (1995) Black carbon—possible source of highly aromatic components of soil humic acids. Org Geochem 23:191–196. https://doi.org/10.1016/0146-6380(95)00003-W
He XX, Chi Q, Cai Z, Cheng Y, Zhang J, Müller C (2020) 15N tracing studies including plant N uptake processes provide new insights on gross N transformations in soil-plant systems. Soil Biol Biochem 141:107666. https://doi.org/10.1016/j.soilbio.2019.107666
Jiang XY, Haddix M, Cotrufo MF (2016) Interactions between biochar and soil organic carbon decomposition: effects of nitrogen and low molecular weight carbon compound addition. Soil Biol Biochem 100:92–101. https://doi.org/10.1016/j.soilbio.2016.05.020
Jones DL, Rousk J, Edwards-Jones G, DeLuca TH, Murphy DV (2012) Biochar-mediated changes in soil quality and plant growth in a three year field trial. Soil Biol Biochem 45:113–124. https://doi.org/10.1016/j.soilbio.2011.10.012
Kopacek J, Cosby BJ, Evans CD, Hruska J, Moldan F, Oulehle F, Santruckova H, Tahovska K, Wright RF (2013) Nitrogen, organic carbon and sulphur cycling in terrestrial ecosystems: linking nitrogen saturation to carbon limitation of soil microbial processes. Biogeochemistry 115:33–51. https://doi.org/10.1007/s10533-013-9892-7
Lehmann J, Silva Junior J, Steiner C, Nehls T, Zech W, Glaser B (2003) Nutrient availability and leaching in an archaeological Anthrosol and a Ferralsol of the Central Amazon basin: fertilizer, manure and charcoal amendments. Plant Soil 249:343–357. https://doi.org/10.1023/A:1022833116184
Li SL, Wang S, Shangguan Z (2019) Combined biochar and nitrogen fertilization at appropriate rates could balance the leaching and availability of soil inorganic nitrogen. Agr Ecosyst Environ 276:21–30. https://doi.org/10.1016/j.agee.2019.02.013
Li JQ, Nie M, Pendall E (2020a) Soil physico-chemical properties are more important than microbial diversity and enzyme activity in controlling carbon and nitrogen stocks near Sydney. Australia Geoderma 366:114201. https://doi.org/10.1016/j.geoderma.2020.114201
Li SL, Wang S, Fan M, Wu Y, Shangguan Z (2020b) Interactions between biochar and nitrogen impact soil carbon mineralization and the microbial community. Soil Tillage Res 196:104437. https://doi.org/10.1016/j.still.2019.104437
Liang B, Zhao W, Yang X, Zhou J (2013a) Fate of nitrogen-15 as influenced by soil and nutrient management history in a 19-year wheat–maize experiment. Field Crops Res 144:126–134. https://doi.org/10.1016/j.fcr.2012.12.007
Liang BQ, Wang C-H, Solomon D, Kinyangi J, Luizão F, Wirick S, Skjemstad J, Lehmann J (2013b) Oxidation is key for black carbon surface functionality and nutrient retention in amazon anthrosols. Br J Env Clim Change 3:9–23. https://doi.org/10.9734/BJECC/2013/2267
Liang C, Schimel JP, Jastrow JD (2017) The importance of anabolism in microbial control over soil carbon storage. Nature Microbiol 2:17105. https://doi.org/10.1038/nmicrobiol.2017.105
Liang C, Amelung W, Lehmann J, Kästner M (2019) Quantitative assessment of microbial necromass contribution to soil organic matter. Glob Change Biol 25:3578–3590. https://doi.org/10.1111/gcb.14781
Liang C, Kaestner M, Joergensen R (2020) Microbial necromass on the rise: the growing focus on its role in soil organic matter development. Soil Biol Biochem. https://doi.org/10.1016/j.soilbio.2020.108000
Liu Q, Liu B, Zhang Y, Hu T, Lin Z, Liu G, Wang X, Ma J, Wang H, Jin H, Ambus P, Amonette JE, Xie Z (2019) Biochar application as a tool to decrease soil nitrogen losses (NH3 volatilization, N2O emissions, and N leaching) from croplands: options and mitigation strength in a global perspective. Glob Change Biol 25(6):2077–2093. https://doi.org/10.1111/gcb.14613
Luo Y, Zang H, Yu Z, Chen Z, Gunina A, Kuzyakov Y, Xu J, Zhang K, Brookes PC (2017) Priming effects in biochar enriched soils using a three-source-partitioning approach: 14C labelling and 13C natural abundance. Soil Biol Biochem 106:28–35. https://doi.org/10.1016/j.soilbio.2016.12.006
Madari BE, Silva MAS, Carvalho MTM, Maia AHN, Petter FA, Santos JLS, Tsai SM, Leal WGO, Zeviani WM (2017) Properties of a sandy clay loam Haplic Ferralsol and soybean grain yield in a five-year field trial as affected by biochar amendment. Geoderma 305:100–112. https://doi.org/10.1016/j.geoderma.2017.05.029
Major J, Lehmann J, Rondon M, Goodale C (2010) Fate of soil-applied black carbon: downward migration, leaching and soil respiration. Glob Change Biol 16:1366–1379. https://doi.org/10.1111/j.1365-2486.2009.02044.x
Meng J, He T, Sanganyado E, Lan Y, Zhang W, Han X, Chen W (2019) Development of the straw biochar returning concept in China. Biochar 1:139–149. https://doi.org/10.1007/s42773-019-00019-0
Mukherjee S, Weihermueller L, Tappe W, Vereecken H, Burauel P (2016a) Microbial respiration of biochar- and digestate-based mixtures. Biol Fertil Soils 52:151–164. https://doi.org/10.1007/s00374-015-1060-x
Mukherjee S, Weihermüller L, Tappe W, Hofmann D, Köppchen S, Laabs V, Vereecken H, Burauel P (2016b) Sorption–desorption behaviour of bentazone, boscalid and pyrimethanil in biochar and digestate based soil mixtures for biopurification systems. Sci Total Environ 559:63–73. https://doi.org/10.1016/j.scitotenv.2016.03.145
Mukherjee S, Tappe W, Weihermueller L, Hofmann D, Köppchen S, Laabs V, Schroeder T, Vereecken H, Burauel P (2016c) Dissipation of bentazone, pyrimethanil and boscalid in biochar and digestate based soil mixtures for biopurification systems. Sci Total Environ 544:192–202. https://doi.org/10.1016/j.scitotenv.2015.11.111
Nan Q, Wang C, Wang H, Yi Q, Liang B, Xu J, Wu W (2020) Biochar drives microbially-mediated rice production by increasing soil carbon. J Hazard Mater 387:121680. https://doi.org/10.1016/j.jhazmat.2019.121680
Neff J, Townsend A, Gleixner G, Lehman S, Turnbull J, Bowman W (2002) Variable effects of nitrogen additions on the stability and turnover of soil C. Nature 419:915–917. https://doi.org/10.1038/nature01136
Nelissen V, Rütting T, Huygens D, Staelens J, Ruysschaert G, Boeckx P (2012) Maize biochars accelerate short-term soil nitrogen dynamics in a loamy sand soil. Soil Biol Biochem 55:20–27. https://doi.org/10.1016/j.soilbio.2012.05.019
Nguyen TTN, Wallace HM, Xu C-Y, Zwieten L, Weng ZH, Xu Z, Che R, Tahmasbian I, Hu H-W, Bai SH (2018) The effects of short term, long term and reapplication of biochar on soil bacteria. Sci Total Environ 636:142–151. https://doi.org/10.1016/j.scitotenv.2018.04.278
Novak JM, Ippolito JA, Watts DW, Sigua GC, Ducey TF, Johnson MG (2019) Biochar compost blends facilitate switchgrass growth in mine soils by reducing Cd and Zn bioavailability. Biochar 1:97–114. https://doi.org/10.1007/s42773-019-00004-7
Oladele S, Adeyemo A, Adegaiye A, Awodun M (2019) Effects of biochar amendment and nitrogen fertilization on soil microbial biomass pools in an Alfisol under rain-fed rice cultivation. Biochar 1:163–176. https://doi.org/10.1007/s42773-019-00017-2
Pan GX, Ding Y, Chen S, Sun J, Feng X, Zhang C, Marios D, Zheng J, Zhang X, Cheng K (2019) Exploring the nature of soil organic matter from humic substances isolation to SOMics of molecular assemblage. Adv Earth Sci 34(5):451–470. https://doi.org/10.11867/j.issn.1001-8166.2019.05.0451.(inChinesewithEnglishabstract)
Qiu QY, Wu L, Ouyang Z, Li B, Xu Y, Wu S, Gregorich EG (2016) Priming effect of maize residue and urea N on soil organic matter changes with time. Appl Soil Ecol 100:65–74. https://doi.org/10.1016/j.apsoil.2015.11.016
Quilliam RS, Glanville HC, Wade SC, Jones DL (2013) Life in the ’charosphere’—does biochar in agricultural soil provide a significant habitat for microorganisms? Soil Biol Biochem 65:287–293. https://doi.org/10.1016/j.soilbio.2013.06.004
Sebilo M, Mayer B, Nicolardot B, Pinay G, Mariotti A (2013) Long-term fate of nitrate fertilizer in agricultural soils. Proc Natl Acad Sci 110:18185–18189. https://doi.org/10.1073/pnas.1305372110
Shi W, Ju Y, Bian R, Li L, Joseph S, Mitchell D, Munroe P, Taherymoosavi S, Pan G (2019) Biochar bound urea boosts plant growth and reduces nitrogen leaching. Sci Total Environ 701:134424. https://doi.org/10.1016/j.scitotenv.2019.134424
Song DL, Tang J, Xi X, Zhang S, Liang G, Zhou W, Wang X (2018) Responses of soil nutrients and microbial activities to additions of maize straw biochar and chemical fertilization in a calcareous soil. Eur J Soil Biol 84:1–10. https://doi.org/10.1016/j.ejsobi.2017.11.003
Wang C, Liu D, Bai E (2018a) Decreasing soil microbial diversity is associated with decreasing microbial biomass under nitrogen addition. Soil Biol Biochem 120:126–133. https://doi.org/10.1016/j.soilbio.2018.02.003
Wang L, Li L, Cheng K, Ji C, Yue Q, Bian R, Pan G (2018b) An assessment of emergy, energy, and cost-benefits of grain production over 6 years following a biochar amendment in a rice paddy from China. Environ Sci Pollut Res 25:9683–9696. https://doi.org/10.1007/s11356-018-1245-6
Wang C, Wang X, Pei G, Xia Z, Peng B, Sun L, Wang J, Gao D, Chen S, Liu D, Dai W, Jiang P, Fang Y, Liang C, Wu N, Bai E (2020a) Stabilization of microbial residues in soil organic matter after two years of decomposition. Soil Biol Biochem 141:107687. https://doi.org/10.1016/j.soilbio.2019.107687
Wang T, Tian Z, Tunlid A, Persson P (2020b) Nitrogen acquisition from mineral-associated proteins by an ectomycorrhizal fungus. New Phytologist n/a. https://doi.org/10.1111/nph.16596
Weng Z, Van Zwieten L, Singh BP, Kimber S, Morris S, Cowie A, Macdonald LM (2015) Plant-biochar interactions drive the negative priming of soil organic carbon in an annual ryegrass field system. Soil Biol Biochem 90:111–121. https://doi.org/10.1016/j.soilbio.2015.08.005
Weng Z, Van Zwieten L, Singh BP, Tavakkoli E, Joseph S, Macdonald L, Rose T, Rose M, Kimber S, Morris S, Cozzolino D, Araujo J, Archanjo B, Cowie A (2017) Biochar built soil carbon over a decade by stabilizing rhizodeposits. Nature Clim Change. https://doi.org/10.1038/nclimate3276
Wu P, Ata-Ul-Karim ST, Singh BP, Wang H, Wu T, Liu C, Fang G, Zhou D, Wang Y, Chen W (2019) A scientometric review of biochar research in the past 20 years (1998–2018). Biochar 1:23–43. https://doi.org/10.1007/s42773-019-00002-9
Yan M, Pan GX, Lavallee JM, Conant RT (2020) Rethinking sources of nitrogen to cereal crops. Glob Change Biol 26:191–199. https://doi.org/10.1111/gcb.14908
Yu L, Yu M, Lu X, Tang C, Liu X, Brookes PC, Xu J (2018) Combined application of biochar and nitrogen fertilizer benefits nitrogen retention in the rhizosphere of soybean by increasing microbial biomass but not altering microbial community structure. Sci Total Environ 640–641:1221–1230. https://doi.org/10.1016/j.scitotenv.2018.06.018
Zheng H, Wang X, Luo X, Wang Z, Xing B (2018) Biochar-induced negative carbon mineralization priming effects in a coastal wetland soil: Roles of soil aggregation and microbial modulation. Sci Total Environ 610–611:951–960. https://doi.org/10.1016/j.scitotenv.2017.08.166
Acknowledgments
We would like to thank Yinfeng Ge and Tianyi Gao for the management of the field operations. And also thank Muhammad Imran Ashraf (Sustainable Development Study Centre, Government College University, Lahore, Punjab, Pakistan) for revision. We would like to express our gratitude to EditSprings (https://www.editsprings.com/) for the expert linguistic services provided. This work was supported by the Natural Science Foundation of China (31972511), and the National Key Research and Development Program of China (No. 2018YFD0300308; 2017YFD0300700).
Author information
Authors and Affiliations
Contributions
Conceptualization: XH; methodology: JP, XH; formal analysis and investigation: JP, NL, KC, JY, XZ, PL, NL; writing-original draft preparation: JP; writing-review and editing: PL, NL; funding acquisition: XH, NL; resources: XH, NL; supervision: NL, JY. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peng, J., Han, X., Li, N. et al. Combined application of biochar with fertilizer promotes nitrogen uptake in maize by increasing nitrogen retention in soil. Biochar 3, 367–379 (2021). https://doi.org/10.1007/s42773-021-00090-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s42773-021-00090-6