
Abstract
The Blesa Formation is one of the richest formations of the Barremian of the Iberian Peninsula in terms of vertebrate biodiversity. In the La Cantalera-1 site more than 32 vertebrate taxa have been identified, but the only sauropod remains are three isolated teeth related to Euhelopodidae. In this paper, all the sauropod material studied so far in this formation is reviewed and new remains are described. The material comes from different sites in all three depositional sequences of the Blesa Formation. A new tooth, a fragmented cervical vertebra and a caudal vertebra are described in the La Cantalera-1 site and assigned tentatively to Euhelopodidae. One left tibia, one left ischium and one caudal vertebra have been recovered in different fossils sites in the Lower, Middle and Upper Blesa sequences respectively. These new sauropod remains are assigned to Titanosauriformes, the most common group of dinosaurs in the Barremian of the Iberian Peninsula. The isolated character of the fossils precludes more precise taxonomic attributions.
Resumen
La Formación Blesa es una de las formaciones con mayor biodiversidad de vertebrados del Barremiense de la península Ibérica. En el yacimiento de La Cantalera-1, donde se han identificado más de 32 taxones de vertebrados, los únicos restos de saurópodos son tres dientes aislados relacionados con Euhelopodidae. En este trabajo, se ha revisado todo el material de saurópodos estudiado hasta ahora en esta formación y se han descrito nuevos restos. Los fósiles provienen de diferentes yacimientos situados a lo largo de las tres secuencias de depósito de la Formación Blesa. En el yacimiento de La Cantalera-1 un nuevo diente, una vértebra cervical fragmentada y una vértebra caudal se han descrito y asignados tentativamente a Euhelopodidae. Una tibia izquierda, un isquion izquierdo y una vértebra caudal se han recuperado de varios afloramientos en las secuencias Inferior, Media y Superior respectivamente. Estos nuevos restos de saurópodos han sido asignados como Titanosauriformes, el grupo de dinosaurios más común en el Barremiense de la Península Ibérica.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Sauropod dinosaurs are the biggest terrestrial animals that have walked the Earth and their distribution and way of life have been the focus of attention for many researchers. They are a fundamental component of terrestrial ecosystems from the Lower Jurassic to their extinction at the end of the Cretaceous. The Iberian Peninsula has a significant record of sauropods in the Late Jurassic-Early Cretaceous transition. Among the sauropods described in Portugal and Spain are several species of turiasaurs: Zby atlanticus (Mateus et al., 2014), Losillasaurus giganteus (Casanovas et al., 2001) and Turiasaurus riodevensis (Royo-Torres et al., 2006); diplodocoids: Dinheirosaurus lourinhanensis (Bonaparte & Mateus, 1999; Mannion et al., 2012), camarasaurid: Lourinhasaurus alenquerensis (Antunes & Mateus, 2003; Lapparent & Zbyszewski, 1957; Mocho et al., 2014), basally branching macronarians: Aragosaurus ischiaticus (Sanz et al., 1987) and Titanosauriformes: Lusotitan atalaiensis (Antunes & Mateus, 2003; Mannion et al., 2013; Mocho et al., 2017a), Galvesaurus herreroi (Barco et al., 2005; Pérez-Pueyo et al., 2019; Sánchez-Hernández, 2005) and Oceanotitan dantasi (Mocho et al., 2019). In the Valanginian and Hauterivian no taxa have been described and the remains are very scarce. The fossil record improves in the Barremian, where three species of Titanosauriformes have been described, Tastavinsaurus sanzi in Teruel (Canudo et al., 2008), Europatitan eastwoodi in Burgos (Torcida Fernández-Baldor et al., 2017) and Soriatitan golmayensis in Soria (Royo-Torres et al., 2017a, 2017b) as well as the rebbachisaurid Demandasaurus darwini in Burgos (Torcida Fernández-Baldor et al., 2011). Despite the fragmentary character of the remains, the presence of sauropods in the early Barremian of the Iberian Peninsula is of great paleobiogeographical significance (typified by the rebbachisaurids) and evolutionary interest (notably the presence of early-branching somphospondylans). Canudo et al. (2002) described and assigned to the Asiatic clade Euhelopodidae two isolated teeth and fragments of one more. Euhelopodids have been recovered as early-branching titanosauriforms along the last two decades but Moore et al. (2020) shows that this group of Asiatic sauropods needs to be scrutinized.
The Iberian Range presents numerous outcrops of continental and transitional sediments with dinosaur remains from the lower Barremian (e.g., Aurell et al., 2018; García-Cobeña et al., 2022; Gasulla et al., 2015; Mocho et al., 2017c; Torcida Fernández-Baldor et al., 2017), so it seems an appropriate place to study the sauropod diversity of the lower Barremian in the Iberian Peninsula. One of the most interesting areas is the Oliete sub-basin in Teruel, located within the Mesozoic Maestrazgo Basin. The Blesa Formation represents the start of Cretaceous sedimentation (Barremian) in the Oliete sub-basin, which is the most northwestern sub-basin of the Maestrazgo Basin (Teruel). The dinosaur biodiversity of the Lower Blesa Sequence (Aurell et al., 2018) is relatively well known due to the La Cantalera-1 site, where hundreds of fossils have been recovered, most of them isolated teeth (Canudo et al., 2010). However, sauropod fossils are scarcer: two isolated teeth and fragments of one more have been described (Canudo et al., 2002). We here present a tooth, a fragmented cervical vertebra and a caudal vertebra tentatively assigned to Euhelopodidae indet. of this site. Until now, no sauropod remains have been found in the Middle and Upper Blesa Sequences (Aurell et al., 2018). The objective of this work is to analyse the diversity of sauropods from the Blesa Fm., compiling and reviewing all the previously published sauropod remains from this formation, and describing previously unpublished fossil remains from the three sequences.
2 Geographical and geological settings
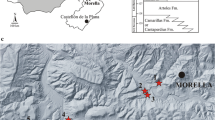
The Oliete sub-basin is located 100 km south of Zaragoza in northeast Spain. Geologically, the Oliete sub-basin is in the northwestern area of the Maestrazgo Basin, bounded by the Sierra de Arcos thrust system to the northeast and the Montalbán structural high to the south (Aurell et al., 2018) (Fig. 1). The Blesa Formation includes an up-to-150 m-thick siliciclastic-carbonate succession (Fig. 1c), which represents the earliest synrift sedimentary fill of the Oliete sub-basin (Salas et al., 2001; Soria et al., 1995). The Blesa Formation is subdivided into three depositional sequences: the Lower, Middle and Upper Blesa sequences (Aurell et al., 2018). The charophyte record is indicative of the early to mid-late Barremian, including the presence of Atopochara trivolvis triquetra identified in the Lower Blesa Sequence (Aurell et al., 2018). The overlying Alacón Fm. (Fig. 1b) has recently been reevaluated, and the Barremian-Aptian boundary is apparently lost due to a sedimentation gap between the Alacón and Forcall formations (García-Penas et al., 2022), which indicates that the depositional age of the upper sequence of the Blesa Fm. is earlier than the end of the Barremian.

Geographical and geological situation of the Oliete sub-basin (Teruel, Spain). a Mesozoic basins of the Iberian Peninsula. The square indicates the location of the Oliete sub-basin (modified from Medrano-Aguado et al., 2022). b Lower Cretaceous formations of the Oliete sub-basin (modified from Aurell et al., 2018). c Stratigraphic section of the Blesa Formation in Josa with the position of the different sauropod fossil sites (modified from Aurell et al., 2018). d Geological map of the Oliete sub-basin with the Blesa Formation (modified from Aurell et al., 2018). Stars indicate the sauropod fossil sites studied in this paper: 1: La Cantalera-1; 2: Los Planos; 3: Santa Cruz-1; 4: Camino San Blas
Since the 1990s, several vertebrate sites have been discovered among the outcrops of the Blesa Formation, but most of the work on them comes from the last decade (Alonso and Canudo, 2015; Alonso et al., 2018; Canudo et al., 2010; Holgado et al., 2019; Medrano-Aguado et al., 2022; Parrilla-Bel & Canudo, 2015). Four of these vertebrate fossil sites have yielded sauropod remains: La Cantalera-1 and Los Planos in the Lower Blesa Sequence, Santa Cruz-1 in the Middle Blesa Sequence and Camino San Blas in the Upper Blesa Sequence (Fig. 1c, d). Los Planos, Santa Cruz-1 and Camino San Blas are described here for the first time.
3 Palaeontological overview of the Blesa Formation
The Lower Blesa Sequence (LBS), deposited in continental environments, is formed by two well differentiated lithologies: red clays facies in the lower part and limestones and marls in the upper part. La Cantalera-1, one of the most significant vertebrate microfossil sites of the European Barremian in terms of diversity (Canudo et al., 2010), is located in an exceptional 50–80 m outcrop in the lower red clay part. These facies represent distal alluvial to palustrine mudflat deposits with local sheet-flood and debris-flow deposits (Aurell et al., 2018). Most vertebrate groups are represented in this site (in many cases as isolated teeth): ornithopod, ankylosaur, theropod and sauropod dinosaurs (Alonso & Canudo, 2016; Canudo et al., 2002, 2010; Gasca et al., 2014); goniopholidid, bernissartiid, atoposaurid and multiple ziphodont crocodylomorphs (Puértolas-Pascual et al., 2015); pterosaurs teeth, chelonian and other reptilian remains (Canudo et al., 2010); multituberculate mammalian teeth (Badiola et al., 2008); small bone fragments referred to lissamphibians; isolated vertebral centra of Osteichthyes; abundant eggshells fragments and three ootaxa (Canudo et al., 2010; Moreno-Azanza et al., 2014). Specifically, sauropod remains are scarce and correspond to isolated teeth (MPZ 97/464, MPZ 2001/206, Canudo et al., 2002; MPZ 97/465, Ruiz-Omeñaca, 2006; MPZ 2023/79, this work) and fragmented postcranial bones (MPZ 2023/80 and MPZ 2023/81). All these remains will be described herein.
A second fossiliferous locality with sauropod remains is the fossil site of Los Planos in the vicinity of Muniesa (Teruel). An isolated tibia is the only known bone discovered and collected. It was excavated and collected in the 1970s by Joaquín Colera Gascón, a teacher from Muniesa, who added this fossil to the private collection of the La Salle-Montemolín School. In 2019 the fossil was donated to the Museo de Ciencias Naturales de la Universidad de Zaragoza. The fossil comes from a Weald facies outcrop located north of the village of Alacón, lying over the Jurassic limestones that form the Mortero ravine, according to the “Carta Paleontológica de Aragón”. The Los Planos site is found near the northern limit of the Oliete sub-basin, where most of the Blesa Formation is covered by the Cenozoic fill of the Ebro Basin. Geologically it is located in the Lower Blesa Sequence. The site is represented by a small outcrop of the limestones and marls of the upper part of the Lower Blesa Sequence which correspond to distal alluvial to palustrine-lacustrine deposits. The isolated tibia is the first direct vertebrate fossil from these levels, since only a highly dinoturbed calcareous level with more than 50 dinosaur footprints has been identified to date in the locality of Obón (Aurell et al., 2018).
The Middle Blesa Sequence is formed by an alternation of oyster-rich marls and limestones deposited in a shallow restricted bay to coastal environment. Numerous fossil sites have been located in this sequence, but no paleontological site with a high concentration of fossils. Isolated bones from marine vertebrates are relatively common, including plesiosaur and crocodylomorphs remains (Parrilla-Bel & Canudo, 2015, 2018). The pterosaur species Iberodactylus andreui has also been described from a partial skull recovered in this sequence (Holgado et al., 2019). The oyster-rich facies bear microfossil accumulations where isolated teeth and fragmented bones belonging to Pycnodontiformes, Osteichthyes and small Chondrichthyes (Canudo et al., 2010) are abundant. Continental vertebrate remains are scarcer. The Santa Cruz-1 site was located and excavated in 2009 in a limestone level assigned to the Middle Blesa Sequence, near the top of the sequence. A left ischium of a sauropod (MPZ 2019/262) from Santa Cruz-1 represents the first dinosaurian material described in the Middle Blesa Sequence.
The Upper Blesa Sequence (UBS) is represented by distal alluvial to palustrine-lacustrine deposits with local marine influence (Aurell et al., 2018). Vertebrate remains were unknown in the Upper Sequence until the end of the 2010s. Barranco del Hocino-1, in the greyish marls/clays of the lower part of this sequence, is the best-studied palaeontological site from the UBS, but no sauropod remains have been recovered. The fossil association is dominated by a disarticulated ornithopod skeleton that could represents a new species (Medrano-Aguado et al., 2022). In addition, ankylosaur bones, theropod teeth (belonging to Spinosauridae and Carcharodontosauria) and scarce microvertebrate remains (Crocodylomorpha and Osteichthyes teeth), turtle shell fragments, coprolites and eggshells are also present (Alonso et al., 2016, 2018). The most basal levels of the UBS in Alacón are represented by 1–3 m-thick conglomerate facies with isolated vertebrate remains. MPZ 2022/726 is a sauropod vertebra donated by a resident of the village. The bone came from a conglomerate rock that served as a wall delimiting an olive plantation. This specimen came from a conglomeratic level that marks the basalmost part of the UBS.
Three other sites with abundant sauropod dinosaur material have been found in the palustrine-lacustrine limestone facies of the Middle and Upper Blesa Sequences, but excavation of these sites is still pending (Aurell et al., 2018).
4 Material and methods
The anatomical nomenclature used in this study follows the terminology common in other publications. The fossil bones are stored at the Museo de Ciencias Naturales de la Universidad de Zaragoza, under the label MPZ (Canudo, 2018). MPZ 2023/78 belongs to the “La Salle Montemolín” Collection. The new material studied is an isolated tooth (MPZ 2023/79), a cervical vertebra (MPZ 2023/80), two caudal vertebrae (MPZ 2023/81 and MPZ 2022/726), a left ischium (MPZ 2019/262) and a tibia (MPZ 2023/78). Mechanical and chemical methods were used for the preparation of the different fossils. The conservation of these specimens took place in the palaeontological laboratory of the University of Zaragoza and in Paleoymas S.L.
5 Systematic palaeontology
We have followed a stratigraphic order to describe this section of the work. We want to describe every element in their context, locating them in their respective depositional Sequence of the Blesa Formation, due to the possible differences in age (see Aurell et al., 2018, Fig. 11).
5.1 La Cantalera-1 site (Josa, Teruel), Lower Blesa Sequence, lower Barremian
5.1.1 Material
Four isolated teeth (MPZ 97/464, MPZ 2001/206, MPZ 97/465, MPZ 2023/79), one cervical vertebra (MPZ 2023/80) and one caudal vertebra (MPZ 2023/81).
-
DINOSAURIA Owen, 1841
-
SAURISCHIA Seeley, 1887
-
SAUROPODA Marsh, 1878
-
NEOSAUROPODA Bonaparte, 1986
-
TITANOSAURIFORMES Salgado et al., 1997
5.1.1.1 MPZ 2023/81
Description: MPZ 2023/81 is a middle-posterior caudal vertebra (Fig. 2). The centrum is well preserved, but the articular surfaces are eroded and the posterior one is broken. The beginning of the neural arch is preserved. The centrum is 112 mm long and the anterior articular surface is 63 mm wide and 51 mm high. The articular surfaces of the centrum are slightly concave, and the centrum is spool-shaped with both lateral and ventral surfaces that are anteroposteriorly concave. In the posterior part of the ventral surface there are three small foramina (Fig. 2d). The neural arch is located anteriorly in the centrum, and there is no evidence of transverse processes. In the broken part of the posterior surface the internal structure shows no evidence of any type of cavities such as camellae.

Caudal vertebra. MPZ 2023/81 in a dorsal view. b Left lateral view. c Right lateral view. d Ventral view. e Anterior view. f Posterior view. Abbreviations: f: foramina; na: neural arch
Discussion: The position of the neural arch in the anterior part of the centrum is a synapomorphy of Titanosauriformes (Salgado et al., 1997). The characters of MPZ 2023/81 are coincident with the other sauropods of the Lower Cretaceous of the Iberian Peninsula such as Soriatitan (Royo-Torres et al., 2017a, 2017b) and Tastavinsaurus (Canudo et al., 2008; Royo-Torres, 2009), as well as with Euhelopodidae, which is apparently present in the La Cantalera-1 site.
-
DINOSAURIA Owen, 1841
-
SAURISCHIA Seeley, 1887
-
SAUROPODA Marsh, 1878
-
NEOSAUROPODA Bonaparte, 1986
-
TITANOSAURIFORMES Salgado et al., 1997
-
SOMPHOSPONDYLI Wilson & Sereno, 1998
5.1.1.2 MPZ 2023/80
Description: MPZ 2023/80 is a poorly preserved and distorted opisthocoelus cervical vertebra (Fig. 3). Part of the neural arch is collapsed over the centrum and the preserved part of the centrum is slightly distorted laterally and dorsoventrally compressed. Anterior and posterior articulation surfaces are distinguishable. The right and ventral surfaces of the centrum are also preserved. The centrum is 230 mm long and the distance between the end of the parapophyses and the middle part of the centrum is 95 mm. MPZ 2023/80 has a well-developed sub-hemispherical anterior articulation and a concave posterior articulation. The anterior articulation is wider (72 mm) than high (46 mm preserved) and long (38 mm). The ventral side is fragmented but preserves part of a ventral keel (Fig. 3b, c). Anteroposteriorly the ventral side of the centrum is slightly concave. The anteroposterior length / height ratio of the posterior face (measuring the height from the base to the middle part of the articulation and doubling it due to the fragmented condition of the surface) is near to 3.83, but this figure should be treated with caution because the posterior face is incomplete. In the right lateral face of the centrum, the base of the pneumatic fossa is preserved. In the middle part of this face, the ventral part of the pleurocentral lamina is preserved (Fig. 3a) so the pleurocoel is probably divided into more than one chamber. The preserved parapophysis is situated ventrolaterally in the anterior part of the centrum, just behind the articular condyle and there is no evidence of a foramen or excavation in its dorsal surface. In the left ventral surface, between the start of the parapophysis and the anterior articular surface, there are two small ridges directed posteriorly with a soft groove in the middle. The internal structure corresponds to a camellate vertebra, composed of broken, thin, small bone laminae and small cavities filled with sediment that can be seen in the broken areas of the centrum.

Cervical vertebra. MPZ2023/80 in a dorsal view. b Ventral view. c Right lateral view. d Anterior view. e Posterior view. Abbreviations: cs: camellate structure; p: parapophyses; pcl: pleurocentral lamina; pl: pleurocoel: r: ridge; vk: ventral keel
Discussion: Strong opisthocoelus cervical vertebrae are associated with Eusauropoda as well as with some ceratosaurs and tetanurans theropods (Upchurch, 1995; Weishampel et al., 2004; Wilson & Sereno, 1998). MPZ 2023/80 is a strong opisthocoelus vertebra with the parapophysis situated in the anteroventral corner of the centrum, a common condition in cervical sauropod vertebrae. The presence of a pleurocentral lamina dividing the pleurocoel is a synapomorphy of Neosauropoda (Wilson & Sereno, 1998). The height-width ratio of the anterior cervical centrum is less than 1 whereas in Shunosaurus (McIntosh, 1990), Omeisaurus (He et al., 1988), Euhelopus and Mamenchisaurus it is 1.25 (Upchurch, 1998). MPZ 2023/80 is poorly preserved and distorted, so this ratio should be treated with caution. The presence of a well-developed ventral keel is common in basal sauropods, but it is greatly reduced or absent in most genera (Upchurch et al., 2004). The presence of a ventral keel has been described in some cervical vertebrae of various Early Cretaceous sauropods such as the Iberian Europatitan (Torcida Fernández-Baldor et al., 2017), Demandasaurus (Torcida Fernández-Baldor et al., 2011), and the Chinese somphospondylan Euhelopus. The presence of a soft ventral keel in cervical vertebrae seems to be undiagnostic in sauropods. The presence of camellae is shared with somphospondylan sauropods and is found in the phylogenetically unstable genus Euhelopus (within Somphospondyli sensu Wilson & Upchurch, 2009; Mannion et al., 2019). MPZ 2023/80 is a poorly preserved sauropod cervical vertebra that is difficult to assign to a genus or species. All the characters described in MPZ 2023/80 are characters present in titanosauriform taxa, especially in somphospondylans. This vertebra could belong to Euhelopus given its size and length and the presence of a pleurocentral lamina, soft ventral keel and camellate pneumaticity and this would be consistent with the presence of Euhelopus teeth in the La Cantalera-1 site (Canudo et al., 2002 and this work). The direction of the parapophysis seems to be less lateral than in the Asiatic taxon, but the preservation conditions, including dorsoventral compression, affect directly in the size, morphology, and direction of this structure and makes it difficult to determine the presence or absence of this character in MPZ 2023/80.
-
DINOSAURIA Owen, 1841
-
SAURISCHIA Seeley, 1887
-
SAUROPODA Marsh, 1878
-
NEOSAUROPODA Bonaparte, 1986
-
TITANOSAURIFORMES Salgado et al., 1997
-
EUHELOPODIDAE Romer, 1956
-
Euhelopodidae indet.
Description: Three isolated teeth were described by Canudo et al. (2002) and Ruiz-Omeñaca (2006). MPZ 97/464 (Fig. 4g) is a D-shaped semi-spatulate tooth with a cingular structure near the base of the crown on the lingual side (Canudo et al., 2002). MPZ 2001/206 is a rare sauropod tooth. It is described as an unformed tooth because its surface is completely smooth, without ornamentation and denticles. Finally, MPZ 97/465 is a fragment of the lingual face of a tooth with an ornamentation pattern similar to MPZ 97/464. These teeth were assigned to Euhelopodidae indet. (Canudo et al., 2002; Ruiz-Omeñaca, 2006).

Tooth. MPZ 2023/79 in a lingual view. b Labial view. c Apical view. d Distal view. e Medial view. f Distal-labial view, note the rounded-boss structure. MPZ 97/464 in g: lingual view, note that the rounded boss is partially worn. Abbreviations: alr: apicobasal lingual ridge; ps: possible pathological surface; rb: rounded boss; wf: wear facets
5.1.1.3 MPZ 2023/79
Description: MPZ 2023/79 is a complete sauropod tooth with part of the root preserved (Fig. 4). It is 16.7 mm high, with a maximum width of 9.4 mm mesiodistally and a width of 6.8 mm labiolingually measured in the part of the root preserved. The slenderness index (SI, maximum mesiodistal crown width divided by the apicobasal length, Upchurch, 1998) of the tooth is 1.77. Following the classification of Mocho et al. (2017b) for isolated sauropod teeth MPZ 2023/79 is a spatulate tooth with wrinkled enamel and a D-shaped cross-section. The enamel is preserved, and the tooth has a slightly worn section at the apex of the lingual surface (Fig. 4a, d). The labial face is strongly convex mesiodistally with a well-developed apicobasal bulge divided at the apex by a smooth central groove and bordered by two smooth grooves along the face. The lingual face is flat-to-convex mesiodistally and concave apicobasally with a smooth central apicobasal bulge. At the base of the crown the mesial edge is slightly concave in lingual view, whereas the distal edge is slightly convex. There are two leaf-shaped surfaces at the mesial and distal edge of the lingual face near the crown base, more developed at the distal edge, and a cingulum at the base of the crown in the lingual surface (Fig. 4d, e). The ridges that commonly separate these facets from the lingual face are smooth and at the mesial edge it is practically non-existent. There is a rounded boss over the leaf-shaped distal surface in the lingual face (Fig. 4f). The apex of the tooth is highly deflected toward the lingual face and has two small wear facets at the distal end of the apex, with the higher one more developed (Fig. 4a). The pattern of the enamel consists of an alternation of anastomosed crests and grooves that are mesiodistally organized. This pattern is present on the labial and lingual face and at the mesial edge. Near the apex, especially on the lingual face, and at the distal edge the enamel surface is smooth. In the middle of the labial face, the enamel has a strange surface, maybe related with a pathology during the formation of the tooth (Fig. 4b).
Discussion: The wear facets are located in the lingual face near the distal border. This position of the wear facet allows us tentatively to assign MPZ 2023/79 to a right maxillary tooth (Carballido & Pol, 2010; Wilson & Sereno, 1998). The higher curvature of the apex of this tooth in comparison it with those described by Canudo et al. (2002) could be related with a more posterior position of the tooth in the tooth row (Holwerda et al., 2015; Madsen et al., 1995; Osborn & Mook, 1921; Ostrom & McIntosh, 1966; Ouyang & Ye, 2002; Wilson & Upchurch, 2009). The enamel texture and the spatulate morphology of the tooth are characters that have been associated with the teeth of Camarasaurus and Euhelopus (e.g., Madsen et al., 1995; Mocho et al., 2017b; Ostrom & McIntosh, 1966; Wilson & Upchurch, 2009). Canudo et al. (2002) associated the sauropod teeth from the La Cantalera-1 site with Euhelopus on the basis of the rounded lingual bosses that are considered an autapomorphy of the group (Barret & Wang, 2007; Wilson, 2002; Wilson & Upchurch, 2009). The general spatulate morphology, the enamel structure and the presence of a rounded boss in the lingual face over the leaf-shaped distal surface of MPZ 2023/79 are similar to the Euhelopus teeth described by Canudo et al. (2002) from the same site. The SI of the tooth is 1.77. This value has been used to differentiate Camarasaurus from Euhelopus teeth by Mocho et al. (2017b), using the comparison made by Chure et al. (2010). According to these papers, the results obtained here are suggestive of Camarasaurus (1.57–2.47) rather than Euhelopus (2.20–3.33). Nevertheless, the SI range for Euhelopodidae in Chure et al. (2010) is wider (1.71–3.70), and the SI value of MPZ 2023/79 (1.77) falls within this range. The curvature of the apex, related with the possibly caudal position of the tooth in the tooth row, makes the tooth relatively short in comparison with other teeth in the tooth row and could explain this value far from the mean for Euhelopodidae. The general morphology, the enamel structure and the presence of the rounded boss allow us tentatively to assign this tooth to Euhelopodidae and differentiate it from the similar spatulate teeth of camarasaurids.
5.2 Los Planos (Muniesa, Teruel), Lower Blesa Sequence, Barremian
Material: MPZ 2023/78 is an incomplete left tibia (Fig. 5).
-
DINOSAURIA Owen, 1841
-
SAURISCHIA Seeley, 1887
-
SAUROPODA Marsh, 1878
-
NEOSAUROPODA Bonaparte, 1986
-
TITANOSAURIFORMES Salgado et al., 1997
-
Titanosauriformes indet.

Left tibia. MPZ 2023/78 in a anterior view. b Lateral view. c Medial view. d Posterior view. e Proximal view. f Distal view. Abbreviations: aap: astragalus ascendant process; cc: cnemial crest; fas: fibula articulation surface; gr: groove; lc: lateral condyle; lvc: lateroventral condyle; mps: medial process scar
Description: The tibia lacks the cnemial crest, some fragments of the middle of the shaft and the end of the ventral processes (see measurements in Table 1). These fragments were probably lost during the extraction of the bone or over the years due to the history of the fossil. The centre of the proximal surface is slightly concave (Fig. 5e) forming a groove between the lateral edge, which is preserved, and the eroded medial edge. The lateral edge of the proximal surface is straight and forms a small and rough pendant lip over the lateral face. Under this lip, in the lateral face, there is a roughly triangular surface for the attachment of the fibula. The scar of the cnemial crest is in the anterior surface and it continues along the anterolateral surface until near the middle of the shaft. Lateral to the cnemial crest scar, near the proximal end, there is a prominent circular tuberculum that could support the tibia-fibula articulation. The shaft is straight in lateral view, with the lateral surface flat and the medial one slightly concave. The shaft is transversely oval with the major axis oriented anteroposteriorly. The shaft is slightly twisted along its length. The distal surface has a heart-like outline with a groove between the two lateral and posterolateral condyles for the ascendant process of the astragalus (Fig. 5b). This surface is better developed lateromedially than anteroposteriorly and the lateral condyle seems to be bigger than the lateroventral one, but the end of both condyles is lost. The ventral surface of the tibia is rough, with holes and bulges (Fig. 5f). The surface where the medial process should be developed is lost.
Discussion: Although incomplete, the proximal surface is subcircular, a feature typically associated with Neosauropoda (Upchurch et al., 2004; Wilson & Sereno, 1998) and some non-neosauropods such as Lapparentosaurus (Bonaparte, 1986), Jobaria (Wilson, 2002) and Ferganosaurus (Alifanov & Averianov, 2003). This section is completely different from the transversely compressed proximal end observed in non-neosauropods such as Turiasaurus (Royo-Torres & Upchurch, 2012; Royo-Torres et al., 2006), Spinophorosaurus (Remes et al., 2009), Mamenchisaurus (Ouyang & Ye, 2002), and Chuanjiesaurus (Sekiya, 2011) and in some macronarians and deeply-nested titanosauriforms such as Euhelopus (Wiman, 1929, not significantly compressed), Sauroposeidon (D’Emic & Foreman, 2012), and Rapetosaurus (Rogers, 2009). The scar of the cnemial crest is located in the anterior surface, a condition usually associated with the anterolateral direction of the cnemial crest present in Titanosauriformes such as Tastavinsaurus, Ligabuesaurus (Bonaparte et al., 2006; Mannion et al., 2013) and Saltasaurus (Powell, 2003), and related with the embracing articulation between tibia and fibula (D’Emic, 2012). The straight but twisted tibia is shared with other members of Titanosauriformes (Canudo et al., 2008; Salgado et al., 1997). The distal end is expanded mediolaterally and compressed anteroposteriorly, a morphology typical of many titanosauriforms (Mannion & Calvo, 2011; Salgado et al., 1997; Upchurch et al., 2004). The robustness index of 0.27 according to the methodology of Wilson & Upchurch (2003) is similar to other early-branching titanosauriforms, but it should be treated with caution given the fragmented areas of MPZ 2023/78 that directly affect the measurements. The general morphology of the tibia resembles that of some titanosauriforms and is especially similar to Tastavinsaurus with its subcircular proximal end, the straight shaft, the heart-shaped distal end and the morphology of the ascendant process of the astragalus. In terms of size, MPZ 2023/78 is bigger and more robust than the Tastavinsaurus tibia. The absence of the cnemial crest prevents us from recognizing the two diagnostic crests of the divided cnemial crest of Tastavinsaurus in order to achieve a more precise understanding of the relationship between MPZ 2023/78 and this taxon. No tibia has been recovered for the other Barremian titanosauriforms of the Iberian Peninsula Europatitan and Soritatitan.
5.3 Santa Cruz-1 (Obón, Teruel), Middle Blesa Sequence, Barremian
Material: MPZ 2019/262 is a left ischium.
-
DINOSAURIA Owen, 1841
-
SAURISCHIA Seeley, 1887
-
SAUROPODA Marsh, 1878
-
NEOSAUROPODA Bonaparte, 1986
-
TITANOSAURIFORMES Salgado et al., 1997
-
Titanosauriformes indet.
Description: MPZ 2019/262 is a small and almost complete left ischium (Fig. 6). There is a strong predation mark in the lateral surface near the base of the ischial ramus (Fig. 6e). Another possible tooth mark is located anterior to this one. The edge of the pubic peduncle and the anteroventral edge of the ischial ramus are slightly broken due to their slenderness. The iliac peduncle and the end of the ischial ramus are not completely preserved. For measurements see Table 2. The iliac ramus is well differentiated and posteriorly projected, and it has a triangular cross-section in posterior view, being wider in the preserved proximal part. The lateral surface of the acetabulum is slightly concave and has an extremely thin margin, joining the iliac and pubic peduncles. The pubic peduncle is expanded dorsoventrally and its margin is extremely thin mediolaterally. The lateral surface of the main body is convex and the medial surface is concave. The preserved ischial ramus is approximately equal in length to the rest of the ischium. It has a triangular shape in cross-section with a rounded dorsal border. The ramus twists slightly, with the medioventral surface turning into its most ventral position. The ischiatic blade turns posteroventrally, forming a concave lateral outline for the posterior edge of the ischium. The distal part of the ischial ramus becomes slightly wider posteriorly, but the distal end is incomplete. At the end of the medial surface of the ischial ramus there is a symphysis for the attachment with the right ischium (Fig. 6c). From the lateral surface of the ramus emerges a strong ridge that finishes on the main body of the ischium, forming a wide and prominent tuberosity near the posterior edge of the ischium, at the level of the distal end of the pubic peduncle (Fig. 6b, e). This crest corresponds to the attachment of the flexor tibialis internus muscle (Borsuk-Bialynicka, 1977; Gorscak & O’Connor, 2019). The ridge-tuberosity union is pierced by a predation mark.

Left ischium. MPZ 2019/262 in a medial view; b lateral view; c posterior view; d anterior view. e Detail of the tuberculum and ridge. Abbreviations: ac: acetabulum; as: attachment surface; ip: iliac peduncle; ir: ischiatic ramus; pp: pubic peduncle; r: ridge; t: tuberculum; tm: tooth mark
Discussion: MPZ 2019/262 is one of the smallest sauropod ischia described from the Early Cretaceous of the Iberian Peninsula. The general morphology of MPZ 2019/262 is similar to the ischium of the Iberian Titanosauriformes: Europatitan (Torcida Fernández-Baldor et al., 2017), Soriatitan (Royo-Torres et al., 2017a, 2017b), and Tastavinsaurus (Canudo et al., 2008). The tuberosity is not bordered by a groove as occurs in Dicraeosaurus (Janensch, 1961), Camarasaurus (Ostrom & McIntosh, 1966), Aragosaurus (Royo-Torres et al., 2014), Lourinhasaurus (Mocho et al., 2014) and non-neosauropod eusauropods. A tuberosity raised above the flat surface of the ischium is a synapomorphy of titanosauriform sauropods (D’Emic, 2012). This tuberosity is more developed than in Tastavinsaurus, Europatitan, Soriatitan and Oceanotitan (Mocho et al., 2019). The morphology of the ischiatic blade, which is slender, with a slightly expanded distal end, and directed ventrally instead of posteriorly or posteroventrally is more similar to the Soriatitan ischium than to that of Tastanvinsaurus and Europatitan. The extreme slenderness of this ischium, especially in the acetabulum, pubic peduncle, and anteroventral margin, together with the strongly developed tuberosity, are characters that are not shared with other Iberian sauropods. Given the isolated state of the ischium and considering the characters described above we have decided to assign it to Titanosauriformes indet. but it could represent an undescribed taxon.
5.4 Camino San Blas (Alacón, Teruel). Upper Blesa Sequence. Barremian
Material: MPZ 2022/726 is a middle caudal vertebra.
-
DINOSAURIA Owen, 1841
-
SAURISCHIA Seeley, 1887
-
SAUROPODA Marsh, 1878
-
NEOSAUROPODA Bonaparte, 1986
-
TITANOSAURIFORMES Salgado et al., 1997
-
Titanosauriformes indet.
Description: MPZ 2022/726 is a middle-posterior caudal vertebra (Fig. 7). The centrum is broken, and the left posterior half and the neural arch are practically lost. The external surfaces of the union between the articular surfaces and the sides of the centrum are also eroded. The suture of the neural arch with the centrum is totally closed (Fig. 7c), which, together with the size of the centrum, indicates that MPZ 2022/726 might corresponds to an adult specimen. The vertebra is 170 mm long, 110 mm wide and 100 mm high. MPZ 2022/726 is amphicoelous, with both articular surfaces slightly concave. Anteroposteriorly the centrum is concave in both ventral and lateral surfaces and has a smooth ridge directed anteroposteriorly in the middle of the lateral surfaces. The internal structure of the centrum is spongy. The neural arch is located in an anterior position and lacks transverse processes. There is a smooth bulge in the lateral surface, indicative of the middle- caudal position of the vertebra.

Caudal vertebra. MPZ 2022/726 in a dorsal view. b Left lateral view. c Right lateral view. d Ventral view. Note that the convex right surface is due to the fracture of the posterior surface and not the real shape of the surface, which is concave (see a). e Anterior surface. f Posterior view. Abbreviations: nas: neural arch suture
Discussion: The general morphology of MPZ 2022/726 allows us to assign it to the clade Titanosauriformes, with the neural arch displaced to an anterior position (Canudo et al., 2008; Royo-Torres et al., 2017a, 2017b; Salgado et al., 1997). The size of the centrum is similar to those of the middle caudal vertebrae of Soriatitan (Royo-Torres et al., 2017a, 2017b) and probably to those of Europatitan (Torcida Fernández-Baldor et al., 2017), although no middle caudal vertebrae are preserved for the latter.
6 Discussion
The presence of a sauropod related to the Asiatic clade Euhelopodidae mentioned by Canudo et al. (2002) is consistent with the new remains herein described. MPZ 2023/79 is a new complete tooth that is very similar to the others assigned to Euhelopodidae (Canudo et al., 2002) but smaller and more curved than these, probably related with the posterior position of the tooth in the teeth row (Wilson & Upchurch, 2009). MPZ 2023/80 is a poorly preserved cervical vertebra. The camellate structure of the vertebra and the presence of the pleurocentral lamina suggest a possible member of a Somphospondyli or more derived sauropod (D’Emic, 2012; Wedel, 2003). The caudal vertebra MPZ 2023/81 has the neural arch in an anterior position, a feature typical of Titanosauriformes (Upchurch et al., 2004). We lack any evidence to demonstrate that the sauropod material of La Cantalera-1 belongs to the same individual or taxon. Most of the last 20 years papers that carried out sauropods’ taxonomic analyses place Euhelopodidae as a non-titanosaurian somphospondylan (i.e. Mannion et al., 2013; Mocho et al., 2019; Royo-Torres et al., 2017a, 2017b) which is congruent with the morphology and characters described in the above-mentioned material (the Titanosauriformes caudal vertebra, the Somphospondyli cervical vertebrae and the euhelopodid teeth) and could correspond with the same taxon. We propose this assignation because in this site several hundreds of sediments have been washed (see Canudo et al., 2010; Ruiz-Omeñaca, 2006) and hundreds of fossils have been recovered, but only the bones described in this paper could been assigned to sauropods. We prefer to tentatively assign all of this material to the clade Euhelopodidae instead of artificially creating diversity for a single fossil site. For these reasons, we tentatively assign MPZ 2023/80 and MPZ 2023/81 to Euhelopodidae indet. and they could represent the first postcranial elements of Euhelopodidae in the Iberian Peninsula.
MPZ 2023/78, MPZ 2019/262 and MPZ 2022/726 are attributed to Titanosauriformes indet. MPZ 2023/78 (Los Planos site) is an almost complete left tibia recovered 50 years ago but described for the first time in this paper. Of all the Barremian sauropods of the Iberian Peninsula, Tastavinsaurus sanzi is the only one whose tibia is preserved. There are close similarities between this taxon and MPZ 2023/78, but many of these features are also common in early-branching titanosauriforms. In addition, the cnemial crest of MPZ 2023/78 is lost, preventing us from comparing it with the cnemial crest diagnostic of Tastavinsaurus cnemial. MPZ 2023/78 is bigger than the two Tastavinsaurus sanzi tibia described (Canudo et al., 2008; Royo-Torres et al., 2012).
The slenderness of MPZ 2019/262 (Santa Cruz-1 site) is the most notable feature of this sauropod ischium. The general morphology is consistent with the ischia of basally-branching titanosauriforms. The presence of a well-developed tuberculum in the base of the ischial ramus is a remarkable character of this bone. Together with its extreme slenderness, it could indicate the presence of an undescribed taxon in the Oliete sub-basin.
The characteristic anterior displacement of the neural arch in the middle vertebrae of Titanosauriformes allows us to assign the first sauropod fossil from the Upper Blesa Sequence, MPZ 2022/726 (Camino San Blas site), to this group of sauropods. The size of the vertebra seems to be more closely related with larger titanosauriforms such as Europatitan or Soriatitan than with Tastavinsaurus.
Early-diverging titanosauriforms were the most common dinosaurs in the Barremian of the Iberian Peninsula (Canudo et al., 2008; Mocho et al., 2017c; Royo-Torres et al., 2012, 2017a, 2017b; Torcida Fernández-Baldor et al., 2017). In the Oliete sub-basin, only three isolated teeth had been described before this paper. In the light of the new material, we can assume that at least three different titanosauriforms taxa lived in this area: one taxon related with Euhelopodidae, the taxon that produced the ischium MPZ 2019/262, and a large taxon associated with the above-described tibia and caudal vertebra (see Table 3).
Despite the high diversity of sauropods in the Early Cretaceous of the Iberian Peninsula, including several taxa in the Jurassic-Cretaceous transition: Zby (Mateus et al., 2014), Losillasaurus (Casanovas et al., 2001), Turiasaurus (Royo-Torres et al., 2006), Dinheirosaurus (Bonaparte & Mateus, 1999; Mannion et al., 2012, 2013), Lourinhasaurus (Antunes & Mateus, 2003; Lapparent & Zbyszewski, 1957; Mocho et al., 2014), Aragosaurus (Sanz et al., 1987) Lusotitan (Antunes & Mateus, 2003; Mannion et al., 2013; Mocho et al., 2017a), Galvesaurus (Barco et al., 2005; Pérez-Pueyo et al., 2019; Sánchez-Hernández, 2005) and Oceanotitan (Mocho et al., 2019); and four in the Barremian-Aptian: Demandasaurus (Torcida Fernández-Baldor et al., 2011), Tastavinsaurus (Canudo et al., 2008), Europatitan (Torcida Fernández-Baldor et al., 2017) and Soriatitan (Royo-Torres et al., 2017a, 2017b); the remains of this group of dinosaurs are scarcer in the Blesa Formation. The small size and marginal condition of the Oliete sub-basin within the Maestrazgo Basin, surrounded by the Ebro, Ejulve and Montalbán highs, and the environment dominated by a marine influence, especially in the MBS and occasionally in the UBS (Aurell et al., 2018), could have restricted life in this area, making it a transit sub-basin for this group of dinosaurs. In addition, most of the fossil sites located to date in the Blesa Formation, are represented by isolated bones, which indicates that sauropods were present in the Oliete sub-basin, but that the taphonomy or fossilization conditions were unfavourable for preserving the remains of these big dinosaurs. Finally, palaeontological research in the Blesa Formation have just started, and only the La Cantalera-1 site is well known. Some isolated vertebrate remains are currently under study as well as the Barranco del Hocino-1 site for the UBS (Alonso et al., 2016, 2018; Medrano-Aguado et al., 2022), and these are a good example of the palaeontological potential of this formation. New discoveries could confirm or refute the scarcity of sauropod groups in this sub-basin in comparison with other faunas and with other areas, as well as providing new data on the implications of the Iberian Peninsula for the evolutionary history of this group of dinosaurs in a period of diversification, as the Barremian was for the successful clade Titanosauriformes.
7 Conclusions
In this paper we have reviewed and studied the sauropod remains of the Blesa Formation (Barremian). In the current state of knowledge, sauropod remains are scarcer in this formation than in other Barremian formations of the Iberian Peninsula where sauropod material is more common (i.e., Arcillas de Morella Fm. in the Galve and Morella sub-basins or the Castrillo de la Reina Fm. in the Cameros Basin). A working hypothesis for this could be the preference of Iberian sauropods for a particular environment that was not present or was scarce in the Oliete sub-basin, or it could be a result of taphonomic and fossilization processes.
New sauropod material from the La Cantalera-1 site has been described and assigned to Euhelopodidae indet. This represents the first postcranial material of this clade from the Barremian of the Iberian Peninsula. One tibia, one ischium and one caudal vertebra recovered in different sites in the Blesa Formation have been assigned to Titanosauriformes indet. The unusual morphology of the ischium could be related with a Titanosauriformes taxon distinct from the established ones. Titanosauriformes is the most common group of sauropods in the Barremian of the Iberian Peninsula, so their presence in the Oliete sub-basin is consistent with other evidence. There is no evidence of the presence of other sauropod groups described in the Lower Cretaceous of the Iberian Peninsula, such as diplodocimorphs or titanosaurians, in the Blesa Formation.
Data availability
All authors contributed to the study conception and design. All data supporting the findings of this study are available within this paper. The remains studied here are deposited in a public institution (Museo de Ciencias Naturales de la Universidad de Zaragoza).
Change history
09 November 2023
A Correction to this paper has been published: https://doi.org/10.1007/s41513-023-00223-9
References
Alifanov, V., & Averianov, A. O. (2003). Ferganasaurus verzilini, gen. et sp. nov., a new neosauropod (Dinosauria, Saurischia, Sauropoda) from the Middle Jurassic of Fergana Valley, Kirghizia. Journal of Vertebrate Paleontology, 23, 358–372.
Alonso, A., & Canudo, J. I. (2016). On the spinosaurid theropod teeth from the early Barremian (Early Cretaceous) Blesa Formation (Spain). Historical Biology, 28(6), 823–834.
Alonso, A., Gasca, J. M., Navarro-Lorbés, P., Núñez-Lahuerta, C., Galán, J., Parrilla-Bel, J., Rubio, C., & Canudo, J. I. (2016). La asociación faunística de Barranco del Hocino 1, un nuevo yacimiento de vertebrados del Barremiense (Cretácico Inferior) de Teruel. Cuadernos Del Museo Geominero, 20, 303–307.
Alonso, A., Gasca, J. M., Navarro-Lorbés, P., Rubio, C., & Canudo, J. I. (2018). A new contribution to our knowledge of the large-bodied theropods from the Barremian of the Iberian Peninsula: The “Barranco del Hocino” site (Spain). Journal of Iberian Geology, 44(1), 7–23. https://doi.org/10.1007/s41513-018-0051-9
Antunes, M. T., & Mateus, O. (2003). Dinosaurs of Portugal. Palevol, 2, 77–95.
Aurell, M., Soria, A. R., Bádenas, B., Liesa, C. L., Canudo, J. I., Gasca, J. M., Moreno-Azanza, M., Medrano-Aguado, E., & Meléndez, A. (2018). Barremian synrift depositional sequence evolution in continental to restricted bay environments with high diversity of vertebrates (Blesa Formation, Oliete sub-basin, NE Spain). Journal of Iberian Geology, 44(2), 285–308.
Badiola, A., Canudo, J. I., & Cuenca-Bescós, G. (2008). New multituberculate mammals from the Hauterivian/Barremian transition of Europe (Iberian Peninsula). Palaeontology, 51(6), 1455–1469.
Barco, J. L., Canudo, J. I., Cuenca-Bescós, G., & Ruiz-Omeñaca, J. I. (2005). Un nuevo dinosaurio saurópodo Galvesaurus herreroi gen. nov., sp. Nov., del tránsito Jurásico-Cretácico en Galve (Teruel, NE de España). Naturaleza Aragonesa, 15(4), e17.
Barret, P. M., & Wang, X. I. (2007). Basal titanosauriform (Dinosauria, Sauropoda) teeth from the Lower Cretaceous Yixian Formation of Liaoning Province, China. Palaeoworld, 16, 265–271.
Bonaparte, J. F. (1986). Les dinosaures (Carnosaures, Allosaurides, Sauropodes, Cetiosaurides) du Jurassique Moyen de Cerro Cóndor (Chubut, Argentine). Annales De Paléontologie, 72, 247–386.
Bonaparte, J. F., & Mateus, O. (1999). A new diplodocid, dinheirosaurus lourinhanensis gen. et sp. nov, from the Late Jurassic beds of Portugal: Introduction. Revista del Museo argentino de ciencias naturales Bernardino Rivadavia e Instituto nacional de investigación de las ciencias naturales, 5(2), 13.
Bonaparte, J. F., González Riga, B. J., & Apesteguía, S. (2006). Ligabuesaurus leanzai gen. et sp. nov. (Dinosauria, Sauropoda), a new titanosaur from the Lohan Cura Formation (Aptian, Lower Cretaceous) of Neuquén, Patagonia, Argentina. Cretaceous Research, 27, 364–376.
Borsuk-Bialynicka, M. (1977). A new camarasaurid sauropod Opisthocoelicaudia skarzynskii, gen. n., sp. n. from the Upper Cretaceous of Mongolia. Palaeontologia Polonica, 37, 1–64.
Canudo, J. I. (2018). The collection of type fossils of the Natural Science Museum of the University of Zaragoza (Spain). Geoheritage, 10, 385–392.
Canudo, J. I., Ruiz-Omeñaca, J. I., Barco, J. L., & Royo-Torres, R. (2002). ¿Saurópodos asiáticos en el Barremiense inferior (Cretácico Inferior) de España. Ameghiniana, 39(4), 443–452.
Canudo, J. I., Royo-Torres, R., & Cuenca-Bescós, G. (2008). A new sauropod: Tastavinsaurus sanzi gen. et sp. nov. from the Early Cretaceous (Aptian) of Spain. Journal of Vertebrate Paleontology, 28(3), 712–731.
Canudo, J. I., Gasca, J. M., Aurell, M., Badiola, A., Blain, H.-A., Cruzado- Caballero, P., Gómez-Fernández, D., Moreno-Azanza, M., Parrilla, J., Rabal, R., & Ruiz-Omeñaca, J. I. (2010). La Cantalera: An exceptional window onto the vertebrate biodiversity of the Hauterivian-Barremian transition in the Iberian Peninsula. Journal of Iberian Geology, 36(2), 205–224.
Carballido, J. L., & Pol, D. (2010). The dentition of Amygdalodon patagonicus (Dinosauria: Sauropoda) and the dental evolution in basal sauropods. Comptus Rendus Palevol, 9, 83–93.
Casanovas, M. L., Santafé, J. V., & Sanz, J. L. (2001). Losillasaurus giganteus, a new sauropod from the transitional Jurassic-Cretaceous of the Los Serranos basin (Valencia, Spain). Paleontologia y Evolució 32–33.
Chure, D., Britt, B. B., Whitlock, J. A., & Wilson, J. A. (2010). First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition. Die Naturwissenschaften, 97, 379–391.
D’Emic, M. D. (2012). The early evolution of titanosauriform sauropod dinosaurs. Zoological Journal of the Linnean Society, 166, 624–671.
D’Emic, M. D., & Foreman, B. Z. (2012). The beginning of the sauropod dinosaur hiatus in North America: Insights from the Lower Cretaceous Cloverly Formation of Wyoming. Journal of Vertebrate Paleontology, 32(4), 883–902.
García-Cobeña, J., Verdú, F. J., & Cobos, A. (2022). Abundance of large ornithopod dinosaurs in the El Castellar Formation (Hauterivian-Barremian, Lower Cretaceous) of the Peñagolosa sub-basin (Teruel, Spain). Journal of Iberian Geology. https://doi.org/10.1007/s41513-021-00185-w
García-Penas, A., Aurell, M., & Zamora, S. (2022). Progressive opening of a shallow-marine bay (Oliete Subbasin, Spain) and the record of possible eustatic fall events near the Barremian-Aptian boundary. Palaeogeography, Palaeoclimatology, Palaeoecology, 594, 110938.
Gasca, J. M., Canudo, J. I., & Moreno-Azanza, M. (2014). On the diversity of Iberian iguanodont dinosaurs: New fossils from the lower Barremian, Teruel province, Spain. Cretaceous Research, 50, 264–272. https://doi.org/10.1016/j.cretres.2014.05.009
Gasulla, J. M., Escaso, F., Narváez, I., Ortega, F., & Sanz, J. L. (2015). A new sail-backed styracosternan (Dinosauria: Ornithopoda) from the Early Cretaceous of Morella. Spain. PLoS One, 10(12), 1e27. https://doi.org/10.1371/journal.pone.0144167
Gorscak, E., & O’Connor, P.M. (2019). A new African Titanosaurian Sauropod Dinosaur from the middle Cretaceous Galula Formation (Mtuka Member), Rukwa Rift Basin, Southwestern Tanzania. PLoS ONE. 14(2):e0211412. https://doi.org/10.1371/journal.pone.0211412
He, X. L., Li, C., & Cai, K. J. (1988). The Middle Jurassic dinosaur fauna from Dashanpu, Zigong, Sichuan: Sauropod dinosaurs (2) Omeisaurus tianfuensis. Sichuan Publishing House of Science and Technology.
Holgado, B., Pêgas, R. V., Canudo, J. I., Fortuny, J., Rodrigues, T., Company, J., & Kellner, A. W. (2019). On a new crested pterodactyloid from the Early Cretaceous of the Iberian Peninsula and the radiation of the clade Anhangueria. Scientific Reports, 9(1), 1–10.
Holwerda, F. M., Pol, D., & Rauhut, O. W. M. (2015). Using dental enamel wrinkling to define sauropod tooth morphotypes from the Cañadón Asfalto Formation, Patagonia, Argentina. PLos One, 10, e0118100.
Janensch, W. (1961). Die Gliedmaszen und Gliedmaszengürtel der Sauropoden der Tendaguru-Schichten. Palaeontograph., 3, 177–235.
Lapparent, A. F., & Zbyszewski, G. (1957). Les dinosauriens du Portugal. Memórias dos Serviços Geológicos de Portugal, 2, 1–63.
Madsen, J. H., Jr., McIntosh, J. S., & Berman, D. S. (1995). Skull and atlas-axis complex of the Upper Jurassic sauropod Camarasaurus cope (Reptilia: Saurischia). Bulletin of the Carnegie Museum of Natural History, 31, 1–115.
Mannion, P. D., & Calvo, J. O. (2011). Anatomy of the basal titanosaur (Dinosauria, Sauropoda) Andesaurus delgadoi from the mid- Cretaceous (Albian-early Cenomanian) Río Limay Formation, Neuquén Province, Argentina: Implications for titanosaur systematics. Zoological Journal of the Linnean Society, 163, 155–181.
Mannion, P. D., Upchurch, P., Mateus, O., Barnes, R. N., & Jones, M. E. (2012). New information on the anatomy and systematic position of Dinheirosaurus lourinhanensis (Sauropoda: Diplodocoidea) from the Late Jurassic of Portugal, with a review of European diplodocoids. Journal of Systematic Palaeontology, 10(3), 521–551.
Mannion, P. D., Upchurch, P., Barnes, R. N., & Mateus, O. (2013). Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriformes. Zoological Journal of the Linnean Society, 168, 98–206.
Mannion, P. D., Upchurch, P., Jin, X., & Zheng, W. (2019). New information on the Cretaceous sauropod dinosaurs of Zhejiang Province, China: impact on Laurasian titanosauriform phylogeny and biogeography. Royal Society Open Science, 6(8), 191057.
Mateus, O., Mannion, P. D., & Upchurch, P. (2014). Zby atlanticus, a new turiasaurian sauropod (Dinosauria, Eusauropoda) from the Late Jurassic of Portugal. Journal of Vertebrate Paleontology, 34(3), 618–634.
McIntosh, J. S. (1990). Sauropoda. In D. B. Weishampel, P. Dodson, & H. Osmolska (Eds.), ’The Dinosauria (pp. 345–401). Berkeley and Los Angeles: University of California Press.
Medrano-Aguado, E., Parrilla-Bel, J., Gasca, J. M., Alonso, A., & Canudo, J. I. (2022). Ornithopod diversity in the Lower Cretaceous of Spain: New styracosternan remains from the Barremian of the Maestrazgo Basin (Teruel province, Spain). Cretaceous Research, 144, 105458. https://doi.org/10.1016/j.cretres.2022.105458
Mocho, P., Royo-Torres, R., & Ortega, F. (2014). Phylogenetic reassessment of Lourinhasaurus alenquerensis, a basal Macronaria (Sauropoda) from the Upper Jurassic of Portugal. Zoological Journal of the Linnean Society, 170(4), 875–916.
Mocho, P., Royo-Torres, R., & Ortega, F. (2017a). New data of the Portuguese brachiosaurid Lusotitan atalaiensis (Sobral Formation, Upper Jurassic). Historical Biology, 29(6), 789–817. https://doi.org/10.1080/08912963.2016.1247447
Mocho, P., Royo-Torres, R., Malafaia, E., Escaso, F., & Ortega, F. (2017b). Sauropod tooth morphotypes from the Upper Jurassic of the Lusitanian Basin (Portugal). Papers in Palaeontology. https://doi.org/10.1002/spp2.1075
Mocho, P., Pérez-García, A., Gasulla, J. M., & Ortega, F. (2017c). High sauropod diversity in the upper Barremian Arcillas de Morella Formation (Maestrat Basin, Spain) revealed by a systematic review of historical material. Journal of Iberian Geology, 43, 111–128.
Mocho, P., Royo-Torres, R., & Ortega, F. (2019). A new macronarian sauropod from the Upper Jurassic of Portugal. Journal of Vertebrate Paleontology. https://doi.org/10.1080/02724634.2019.1578782
Moreno-Azanza, M., Canudo, J. I., & Gasca, J. M. (2014). Unusual theropod eggshells from the Early Cretaceous Blesa formation of the Iberian range, Spain. Acta Paleontologica Polonica, 59(4), 843–854.
Osborn, H. F., & Mook, C. C. (1921). Camarasaurus, Amphicoelias, and other sauropods of Cope. Memoirs of the American Museum of Natural History, New Series, 3, 249–387.
Ostrom, J. H., & McIntosh, J. S. (1966). Marsh’s Dinosaurs: The Collection from Como Bluff (Vol. 1, p. 388). Yale University Press.
Ouyang, H., & Ye, Y. (2002). The First Mamenchisaurian Skeleton with Complete Skull, Mamenchisaurus youngi. Sichuan Publishing House of Science and Technology, Chengdu, China, 111 pp. [Chinese]
Parrilla-Bel, J., & Canudo, J. I. (2015). On the presence of plesiosaurs in the Blesa Formation (Barremian) in Teruel (Spain). Neues Jahrbuch Für Geologie Und Paläontologie Abhandlungen, 278(2), 213–227.
Parrilla-Bel, J., & Canudo, J. I. (2018). New longirostrine crocodylomorph remains from the Blesa Formation (Barremian) in the Iberian Peninsula (Spain). Journal of Iberian Geology, 44, 55–66.
Pérez-Pueyo, M., Canudo Sanagustín, J. I., Barco, J. L., & Moreno-Azanza, M. (2019). New contributions to the phylogenetic position of the sauropod Galvesaurus herreroi from the late Kimmeridgian-early Tithonian (Jurassic) of Teruel (Spain). Boletín Geológico y Minero, 130(3), 375–392.
Powell, J. E. (2003). Revision of South American titanosaurid dinosaurs: Palaeobiological, palaeobiogeographical and phylogenetic aspects. Records of the Queen Victoria Museum, 111, 1–173.
Remes, K., Ortega, F., Fierro, I., Joger, U., Kosma, R., Ferrer, J. M. M., Ide, O. A., & Maga, A. (2009). A new basal sauropod dinosaur from the Middle Jurassic of Niger and the early evolution of Sauropoda. PLoS ONE, 4, e6924.
Puértolas-Pascual, E., Rabal-Garcés, R. & Canudo, J. I. (2015). Exceptional crocodylomorph biodiversity of “La Cantalera” site (lower Barremian; Lower Cretaceous) in Teruel, Spain. Palaeontologia Electronica, 18.2.28A, 1–16. https://doi.org/10.26879/514
Rogers, K. C. (2009). The postcranial osteology of Rapetosaurus krausei (Sauropoda: Titanosauria) from the Late Cretaceous of Madagascar. Journal of Vertebrate Paleontology, 29(4), 1046–1086.
Royo-Torres, R. (2009). El saurópodo de Peñarroya de Tastavins. Instituto de Estudios Turolenses-Fundación Conjunto Paleontológico de Teruel-Dinópolis. Monografías Turolenses, 6, pp. 548
Royo-Torres, R., & Upchurch, P. (2012). The cranial anatomy of the sauropod Turiasaurus riodevensis and implications for its phylogenetic relationships. Journal of Systematic Palaeontology, 10, 553–583.
Royo-Torres, R., Cobos, A., & Alcalá, L. (2006). A giant European dinosaur and a new sauropod clade. Science, 314, 1925–1927.
Royo-Torres, R., Alcalá, L., & Cobos, A. (2012). A new specimen of the Cretaceous sauropod Tastavinsaurus sanzi from El Castellar (Teruel, Spain), and a phylogenetic analysis of the Laurasiformes. Cretaceous Research, 34, 61–83.
Royo-Torres, R., Upchurch, P., Mannion, P. D., Mas, R., Cobos, A., Gascó, F., Alcalá, L., & Sanz, J. L. (2014). The anatomy, phylogenetic relationships, and stratigraphic position of the Tithonian-Berriasian Spanish sauropod dinosaur Aragosaurus ischiaticus. Zoological Journal of the Linnean Society, 171(3), 623–655.
Royo-Torres, R., Fuentes, C., Meijide, M., Meijide-Fuentes, F., & Meijide-Fuentes, M. (2017a). A new Brachiosauridae sauropod dinosaur from the lower Cretaceous of Europe (Soria Province, Spain). Cretaceous Research, 80, 38–55.
Royo-Torres, R., Upchurch, P., Kirkland, J. I., DeBlieux, D. D., Foster, J. R., Cobos, A., & Alcalá, L. (2017b). Descendants of the Jurassic turiasaurs from Iberia found refuge in the Early Cretaceous of western USA. Scientific Reports, 7, 14311.
Ruiz-Omeñaca, J. I. (2006). Restos directos de dinosaurios (Saurischia, Ornithischia) en el Barremiense (Cretácico Inferior) de la Cordillera Ibérica en Aragón (Teruel, España). Ph.D. Thesis. Universidad de Zaragoza, Zaragoza, 439pp. (unpublished).
Salas, R., Guimerà, J., Mas, R., Martín-Closas, C., Meléndez, A., & Alonso, A. (2001). Evolution of the Mesozoic Central Iberian Rift System and its Cainozoic inversion (Iberian chain). Mémoires Du Muséum National D’histoire Naturelle, 186, 145–186.
Salgado, L., Coria, R. A., & Calvo, J. O. (1997). Evolution of titanosaurid sauropods: Phytogenetic analysis based on the postcranial evidence. Ameghiniana, 34(1), 3–32.
Sánchez-Hernández, B. (2005). Galveosaurus herreroi, a new sauropod dinosaur from Villar del Arzobispo Formation (Tithonian-Berriasian) of Spain. Zootaxa, 1034(1), 1–20.
Sanz, J. L., Buscalioni, A. D., Casanovas, M. L., & Santafé, J. V. (1987). Dinosaurios del Cretácico Inferior de Galve (Teruel, España). Estudios Geológicos Volumen Extraordinario, Galve Tremp, 45–64.
Sekiya, T. (2011). Re-examination of Chuanjiesaurus anaensis (Dinosauria: Sauropoda) from the Middle Jurassic Chuanjie Formation, Lufeng County, Yunnan Province, southwest China. Memoir of the Fukui Prefectural Dinosaur Museum, 10, 1–54.
Soria, A., Martín-Closas, C., Meléndez, A., Meléndez, M., & Aurell, M. (1995). Estratigrafía del Cretácico Inferior continental de la Cordillera Ibérica Central. Estudios Geológicos, 152(51), 141–152.
Torcida Fernández-Baldor, F., Canudo, J. I., Huerta, P., Montero, D., Suberbiola, X. P., & Salgado, L. (2011). Demandasaurus darwini, a new rebbachisaurid sauropod from the Early Cretaceous of the Iberian Peninsula. Acta Palaeontologica Polonica, 56(3), 535–552.
Torcida Fernández-Baldor, F., Canudo, J. I., Huerta, P., Moreno-Azanza, M., & Montero, D. (2017). Europatitan eastwoodi, a new sauropod from the lower Cretaceous of Iberia in the initial radiation of somphospondylans in Laurasia. PeerJ, 5, e3409.
Upchurch, P. (1995). Evolutionary history of sauropod dinosaurs. Philosophical Transactions of the Royal Society of London, B, 349, 365–390.
Upchurch, P. (1998). The phylogenetic relationships of sauropod dinosaurs. Zoological Journal of the Linnean Society, 124, 43–103.
Upchurch, P., Barrett, P. M., & Dodson, P. (2004). Sauropoda. In D. B. Weishampel, P. Dodson, & H. Osmólska (Eds.), The Dinosauria (2nd ed., pp. 259–324). Berkeley: University of California Press.
Wedel, M. J. (2003). The evolution of vertebral pneumaticity in sauropod dinosaurs. Journal of Vertebrate Paleontology, 23(2), 344–357. https://doi.org/10.1671/0272-4634(2003)023[0344:TEOVPI]2.0.CO;2
Weishampel, D. B., Dodson, P., & Osmólska, H. (Eds.). (2004). The Dinosauria. Berkeley: University of California Press.
Wilson, J. A. (2002). Sauropod dinosaur phylogeny: Critique and cladistic analysis. Zoological Journal of the Linnean Society, 136, 217–276.
Wilson, J. A., & Sereno, P. C. (1998). Early evolution and higher-level phylogeny of sauropod dinosaurs. Journal of Vertebrate Paleontology, 18(Suppl 2), 1–68.
Wilson, J. A., & Upchurch, P. (2003). A revision of Titanosaurus Lydekker (Dinosauria-Sauropoda), the first dinosaur genus with a ‘Gondwanan’ distribution. Journal of Systematic Palaeontology, 1(3), 125–160.
Wilson, J. A., & Upchurch, P. (2009). Redescription and reassessment of the phylogenetic affinities of Euhelopus zdanskyi (Dinosauria: Sauropoda) from the Late Jurassic or Early Cretaceous of China. Journal of Systematic Palaeontology, 7, 199–239.
Wiman, C. (1929). Die Kreide-Dinosaurier aus Shantung. Palaeontologia Sinica, Series C, 6, 1–67.
Acknowledgements
Juanita Alquézar Lázaro donated MPZ 2022/726 to the Museo de Ciencias Naturales de la Universidad de Zaragoza. The education, culture and sports department of the Aragón government let us consult the “Carta Paleontológica de Aragón”. I. Pérez helps with the photography of the fossils. R. Glasgow reviewed the text in English. Eduardo Medrano Aguado has a predoctoral contract financed by the Government of Aragón. Finally, we are grateful to Pedro Mocho, an anonymous reviewer, and the editor for their exhaustive comments, which have greatly enhanced the quality of this paper.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This work is funded by the Spanish Ministry of Science and Innovation (project PID2021-122612OB-I00) and by the Government of Aragón-ERDF (Group E18: Aragosaurus: Recursos Geológicos y Paleoambientales).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
The original online version of this article was revised: The captions of Fig. 2 and Fig. 4 have been swapped.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Medrano-Aguado, E., Parrilla-Bel, J. & Canudo, J.I. Palaeobiodiversity of Sauropoda in the Blesa Formation (Lower Cretaceous) at northeast Spain. J Iber Geol (2023). https://doi.org/10.1007/s41513-023-00219-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s41513-023-00219-5