
Abstract
• Key message
Natural regeneration of P. abies (L.) H. Karst. may reach high densities in lower mountain elevations. The highest densities were found in sites with moderate light availability, with low pH, and not near the riverbank. However, age-height classes differed in the predicted magnitude of response, but were consistent in response directions. Mosses and understory species typical of coniferous forests were positively correlated with regeneration density.
• Context
Norway spruce Picea abies (L.) H. Karst. in Central Europe is at risk under climate change scenarios, particularly in mountain regions. Little is known about the impact of environmental factors on the natural regeneration of P. abies in low-elevation mountain forests.
• Aims
We aimed to assess impacts of distance from the riverbank, soil pH, and light availability on natural P. abies regeneration. We hypothesized that (1) natural P. abies regeneration would depend on light availability and soil pH and (2) there are understory plant species which may indicate the microsites suitable for natural regeneration of P. abies.
• Methods
The study was conducted in the Stołowe Mountains National Park (SW Poland, 600–800 m a.s.l.). We established 160 study plots (25 m2) for natural regeneration, light availability, soil pH, and understory vegetation assessment.
• Results
The highest densities of natural regeneration of P. abies were observed in sites with moderate light availability (0.1–0.2 of open sky) and low pH (3.5–4.5), and located relatively far from the riverbank. Cover of 22 understory plant species were correlated with natural P. abies regeneration densities, mostly positively.
• Conclusion
Different stages of natural regeneration of P. abies revealed different regeneration niches. Most understory plant species (bryophytes and herbs typical of coniferous forests) do not compete with natural regeneration of P. abies.
Similar content being viewed by others


1 Introduction
Natural regeneration is a crucial process responsible for generational replacement in forest ecosystems (Baraloto et al. 2005; Beckage et al. 2005; Dech et al. 2008). Young regeneration is especially vulnerable to limiting abiotic factors, for instance frost or drought (Niinemets and Valladares 2006; Eckstein et al. 2011; Jagodziński et al. 2017), as well as to herbivory (de Chantal and Granström 2007; Iszkuło et al. 2014; Bodziarczyk et al. 2017), leading to low survival rates. Another important factor is competition with understory herbaceous species (Lorimer et al. 1994; Emborg 1998; Chmura and Sierka 2007), as well as competition within the regeneration layer, both intra- and interspecific (Janzen 1970; Robakowski and Bielinis 2011; Kawaletz et al. 2014; Martínez-García et al. 2015). For that reason, assessment of natural regeneration is crucial both for understanding dynamics of forest species composition and for developing forest management strategies.
Norway spruce Picea abies (L.) H. Karst occurs in boreal and mountain forests, but also occupies temperate forests with poor soils. The largest P. abies trees can reach heights of 50–60 m and diameter at breast height (dbh) of 150 cm. Its range in Europe (both natural and introduced) covers an area from the Ural Mountains in the east to Great Britain in the west and from the Alps and Pyrenees in the south to the timberline in Scandinavia in the north. In the Alps, P. abies reaches its maximum elevation of 2400 m a.s.l. (Caudullo et al. 2016). P. abies is a shade-tolerant tree species (4.45 on the 0–5 scale of Niinemets and Valladares 2006), with high moisture requirements (Ellenberg 1988; Niinemets and Valladares 2006). The latter implies low tolerance not only for drought but also for waterlogging (Niinemets and Valladares 2006). P. abies has been widely cultivated since the nineteenth century, as a fast-growing tree producing valuable timber. For that reason, extensive areas of mountain beech and fir forests have been replaced by P. abies plantations (Felton et al. 2010; Caudullo et al. 2016; Naudts et al. 2016).
Future dynamics and occurrence of P. abies in Central Europe seem to be at risk, due to increased frequency and intensity of bark beetle infestations (Kučerová et al. 2008; Bastl et al. 2009; Müller et al. 2010; Seidl et al. 2014), which result from climate change and decreased water availability (Seidl et al. 2011; Gutowski and Jaroszewicz 2015; Grodzki 2016; Dyderski et al. 2018b). This is especially important due to the high transpiration rate of P. abies (Cienciala et al. 1992). Uncertainty about P. abies persistence in forest ecosystems is currently a subject of broad discussions in forest ecology and nature conservation (Brzeziecki et al. 2016; Jaroszewicz et al. 2017). For that reason, most of the papers regarding its natural regeneration are focused on post-disturbance areas (e.g., Kučerová et al. 2008; Pröll et al. 2015; Zeppenfeld et al. 2015). Miina and Saksa (2013) elaborated a model of natural regeneration based on silvicultural treatments and soil properties, but not accounting for light availability. Vacek et al. (2017) studied P. abies regeneration in forests, but their research was focused on natural regeneration of Fagus sylvatica and data about P. abies were only a background for the study. Holeksa et al. (2007) analyzed patterns of natural regeneration of P. abies in natural forests, dominated by P. abies, and Jonášová and Prach (2004) studied its patterns in semi-natural P. abies forests after a bark beetle outbreak. There is a lack of studies focused on natural regeneration of P. abies in secondary forests, where the species is growing outside of sites where it would dominate the potential natural vegetation.
We aimed to assess impacts of distance from the riverbank, soil pH, and indirect light availability on natural regeneration of P. abies in low-elevation mountain forests in a river valley. We hypothesized that (1) natural regeneration of P. abies would be most abundant in sites with the highest light availability and the lowest soil pH and nearest to the riverbank and (2) there are understory plant species which may indicate the microsites suitable for natural regeneration of P. abies.
2 Materials and methods
2.1 Study area
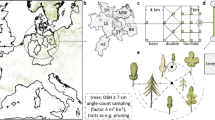
We conducted our study in the Czerwona Woda River Valley in the Stołowe Mountains National Park (SW Poland; 50.47° N, 16.35° E). The Czerwona Woda River is the longest river in the Stołowe Mountains National Park (13.4 km long), and its width ranges from less than 0.5 m in the upper course to ca. 3 m in the lower course. The river valley covers an area of 91.2 ha (as confined by clear topographic features), and elevations range from ca. 800 m a.s.l (source) to 600 m a.s.l (border of the national park). Within the study area, most of the river course flows at a 115° azimuth (EES), and over 90% of study plots were located on flat sites (slopes less than 5°). For that reason, our study did not account for microtopography. Mean annual temperature in 1951–2000 was 7.1 °C (Kłodzko, ca. 20 km from the study site; Trouet and Van Oldenborgh 2013) and mean annual precipitation measured at seven stations (years 1976–2005, stations: Pasterka, Słoszów, Lewin Kłodzki, Chocieszów, Kudowa Zdrój, Polanica Zdrój, and Gajów) was 773 mm (Tarka et al. 2011). The dominant bedrock types in the study area are turonian marls and sandstones (Migoń et al. 2011). Soils in the upper course of the river are haplic cambisols and haplic luvisols; in the middle course, histic gleysols; and in the lower course, gleyic fluvisols and gleyic cambisols (Kabała et al. 2011). Forests of the Stołowe Mountains National Park are mostly transformed by human activity, via replacement of natural, broadleaved forests by coniferous monocultures. Most of the forests are dominated by P. abies, with an admixture of Acer pseudoplatanus, Alnus incana, Betula pendula, Fagus sylvatica, Larix decidua, Larix kaempferi, and Sorbus aucuparia. The most frequent ages of forest stands in the Czerwona Woda River Valley range from 80 to 120 years.
Our preliminary vegetation survey revealed that within the Czerwona Woda River Valley, forests dominated by P. abies cover an area of 67.6 ha (73.4% of the study area), while the coverage of other forest types is only 0.4 ha (Pielech et al. 2018; Wierzcholska et al. 2018). Other forest ecosystems (remnants of riparian forests) compose ca. 1%, while the remainder of the area is covered by meadows or grasslands. The P. abies forests are diverse and represented by different plant communities, including Picea abies-Avenella flexuosa (51.8 ha—56.8%), Vaccinio uliginosi-Piceetum abietis Schubert 1972 (14.8 ha—16.2%), and Equiseto sylvatici-Piceetum abietis (0.4 ha—0.4%). Understories of these forests are dominated by Vaccinium myrtillus, Calamagrostis villosa, Avenella flexuosa, and Oxalis acetosella in the herb layer and Polytrichastrum formosum, Dicranum scoparium, and Pleurozium schreberi in the moss layer.
According to forest structure measurements (all trees > 1.3 m height) conducted in nine plots (0.1 ha each), the basal area of forest stands ranged from 23.9 to 74.9 m2 ha−1 (average 51.6 ± 5.8 m2 ha−1), the mean tree-stand height ranged from 20.0 to 27.8 m (average 25.4 ± 0.9 m), the top height (heights of the 10% of trees with the largest diameters) ranged from 16.7 to 31.7 m (average 26.7 ± 1.7 m), the tree-stand density ranged from 710 to 6830 trees ha−1 (average 2145 ± 733 trees ha−1), and the tree-stand volume ranged from 303.9 to 977.6 m3 ha−1 (average 606.4 ± 83.2 m3 ha−1).
2.2 Data collection
The study was conducted in early September 2017, when current-year natural regeneration was completely developed and canopy cover was maximal (in conditions of minimal light availability). For assessment of natural regeneration, we used a set of study sites, located at 100-m intervals along a 9-km segment (excluding non-forest areas) of the river course. Within each study site, we established two square 25-m2 study plots—the first was established 1 m from the riverbank and the second at a distance from 5 to 50 m from the riverbank (measured using a tape measure). Distance between plots varied from 1 to 45 m (mean 17.1 ± 1.1 m) to ensure that two plots were not pseudoreplications and to account for vegetation diversity within the study area. This system is not ideal, due to non-systematic design of distances from the river, which limits inference about the influence of this parameter. To prevent subjectivity in locating plots, we decided to always establish the first plot at a fixed distance from the riverbank (i.e., 1 m) and the second plot in a different vegetation patch. We determined the differences by visual estimation of dominant understory plant species, which indicate environmental conditions and microsites (Ellenberg and Leuschner 2010). To avoid the observer effect, one person did all the estimations for all study plots. In total, 160 study plots were established along the river (Dyderski et al. 2018a).
In each plot, we investigated only natural regeneration of P. abies within three age-height classes (Jagodziński et al. 2017): seedlings (plants germinated in the current year), short saplings (plants ≥ 1 year old and < 0.5 m tall), and tall saplings (plants 0.5–1.3 m tall). Data on other woody plant species (which rarely occurred) was collected but not analyzed. We also listed understory species and assessed their abundance using a modified, nine-degree Braun-Blanquet cover scale. Vascular plant nomenclature follows the Euro+Med PlantBase (Euro+Med 2006); mosses, Ochyra et al. (2003); and liverworts, Szweykowski (2006). Phytosociological nomenclature for plant associations follows Chytrý (2013). We also measured topsoil (upper 5–10 cm of soil) pH in distilled water solution after 24 h, using an electronic pH meter. We treated soil pH as a proxy for soil fertility (Bigelow and Canham 2002). As a proxy for light availability, we used diffuse non-interceptance (DIFN; fraction of the open sky) obtained by a LAI-2200 plant canopy analyzer (Li-Cor Inc., Lincoln, NE, USA) 50 cm above the forest floor (understory height). For each study plot, we obtained a mean from ten samples. DIFN is literally the ratio of photon flux densities captured by five lenses in an open area (control) and beneath the forest canopy (measurement). The lenses are oriented at 7, 23, 38, 53, and 68° angles to characterize light interception at different heights. DIFN is a direct measurement of the amount of indirect light availability and serves as a common proxy for light availability for plants (Stenberg et al. 1994; Machado and Reich 1999; Knight et al. 2008; Mueller et al. 2016). Although DIFN may be less informative for some researchers, it can be recalculated into leaf area index (LAI, m2 m−2) using the formula LAI = 1.9092*DIFN−0.2528 (R2 of the linearized form = 0.82). However, this proxy does not account for either direct light or photosynthetically active radiation, as the LAI-2200 registers only ultraviolet to blue wavelengths (< 490 nm), which are also important for plant growth (e.g., Ozolinčius et al. 1996; Curt et al. 2005; Löf et al. 2005).
In the study area, soil pH ranged from 3.40 to 6.21, with an average of 4.03 ± 0.04 (lower quartile 3.75, upper quartile 4.22); DIFN ranged from 0.003 to 0.399, with an average of 0.047 ± 0.004 (lower quartile 0.017, upper quartile 0.064). LAI ranged from 2.408 to 8.292 m2 m−2, with an average of 4.723 ± 0.093 m2 m−2 (lower quartile 3.821, upper quartile 5.369). Distance to the riverbank ranged from 1 to 50 m, with an average of 8 ± 1 m (lower quartile 1 m, upper quartile 15 m; Dyderski et al. 2018a).
2.3 Data analysis
We conducted all analyses using R software (R Core Team 2017). All mean values are followed by the standard error (SE). Prior to modeling, we log-transformed dependent variables (natural regeneration densities) and we centered and scaled independent variables (DIFN, soil pH, and distance from the riverbank) using the base::scale() function. These transformations were used to stabilize variance, increase normality of distributions, and overcome problems with different magnitudes of variables. To account for a high number of zero values, we used hurdle models implemented in the pscl package (Zeileis et al. 2008). These models are combinations of two parts: the first part of the model predicts the probability of a non-zero value of the parameter studied, and the second predicts the value using an adequate distribution type. For natural regeneration density, we used the Poisson distribution (with log link function) for count model coefficients and binomial distribution (with logit link function) for zero-inflation model coefficients. We developed these models using the pscl::zeroinfl() function (Zeileis et al. 2008). To predict the proportion of each age-height class in the natural P. abies regeneration, we used zero- and one-inflated beta regression models implemented in the gamlss package (Rigby and Stasinopoulos 2005). This type of model allows for separate estimation of numerous distribution types by separate estimation of estimated value μ, variance σ, and two distribution shape parameters: ν and τ (Ospina and Ferrari 2010). In the case of zero- and one-inflated models, we used beta distributions with a logit link function for μ and σ estimation and the log function for ν and τ estimation. This procedure was implemented in the gamlss::gamlss() function with family = BEINF argument (Rigby and Stasinopoulos 2005). For each age-height class, we started from a model with three potential predictors (DIFN, distance to the riverbank, and soil pH) and we tried to reduce the variable number to decrease Akaike’s information criterion (AIC). The model with the lowest AIC was chosen as the final model.
For visualization of relationships between environmental parameters studied (soil pH, DIFN, and distance from the river), natural P. abies regeneration density, and understory vegetation, we used detrended correspondence analysis (DCA; Hill and Gauch 1980). This technique of unconstrained ordination reveals the main gradients in vegetation. DCA was performed using the vegan::decorana() function (Oksanen et al. 2018). To explain correlations of gradients with measured parameters, we used the vegan::envfit() function (Oksanen et al. 2018). We used DCA instead of other ordination techniques due to long gradients (> 3 SD units). To identify the significant relationships between cover of understory plant species and natural P. abies regeneration, we used Spearman’s rank correlation, as abundance was recorded in an unequal interval scale. These analyses were carried out only for understory plant species which occurred in at least ten plots. We did not adjust p values for multiple hypothesis testing, as such adjustments (e.g., Bonferroni correction) in cases of multiple species testing may cause artificial rejection of biologically important results and each chi-squared test for a particular species should be treated as a separate case (Moran 2003).
3 Results
3.1 Variability of natural regeneration densities
Natural regeneration of P. abies was found in 119 of 160 study plots (Dyderski et al. 2018a). Seedlings were present in 67 study plots, short saplings in 104, and tall saplings in 57. Density of seedlings ranged from 0 to 41,200 ind. ha−1, with an average of 1695.2 ± 405.6 ind. ha−1 (lower quartile 0, upper quartile 900). Densities of short and tall saplings ranged from 0 to 94,000 ind. ha−1 (average 6862.0 ± 1021.4, lower quartile 0, upper quartile 8800) and from 0 to 34,800 ind. ha−1 (average 1180.0 ± 322.2, lower quartile 0, upper quartile 400), respectively. No correlations among densities of particular age-height classes were found, with the exception of positive correlation between seedlings and tall saplings (p < 0.001, r2 = 0.12, y = 689.54 + 0.29x). The proportion of short saplings was negatively correlated with the proportion of seedlings (p = 0.001, r2 = 0.05, y = 0.5214 − 0.3451x) and tall saplings (p = 0.037, r2 = 0.02, y = 0.4951 − 0.2782x), and the proportion of seedlings was negatively correlated with the proportion of tall seedlings (p = 0.025, r2 = 0.02, y = 0.1915 − 0.2154x).
3.2 Impact of resource availability on regeneration density and structure of age-height classes
Zero-inflated Poisson models of natural regeneration densities explained low amounts of variance in seedlings, short saplings, and tall saplings (Table 1). In the model of seedling density, the statistically significant parameters were distance from the riverbank and soil pH: seedling density increased with increasing distance from the riverbank and decreased with increasing soil pH (Fig. 1). Short sapling density decreased with increasing soil pH and increased with increasing DIFN and distance from the riverbank. Tall sapling density increased with increasing distance from the riverbank but decreased with increasing DIFN and soil pH.

Relationships between density of natural regeneration of Picea abies and three environmental variables for three age-height classes: seedlings (germinated in the current year), short saplings (≥ 1 year old, < 50 cm height), and tall saplings (50–130 cm height). Lines represent fitted generalized linear models with Poisson distribution for visualization of general trends. See Table 1 for zero-inflated regression models and their parameters. Note log-scale on the y axis
Analysis of proportions among age-height classes revealed that seedling proportions depended only on distance from the riverbank (Table 2). This relationship was negative (Fig. 2). In the cases of short and tall saplings, final models included only a relationship with soil pH. The proportion of short-sapling regeneration decreased with increasing pH while the proportion of tall saplings increased.

Relationships between proportion of age-height classes of natural regeneration of Picea abies and three environmental variables. Classes: seedlings (germinated in the current year), short saplings (≥ 1 year old, < 50 cm height), and tall saplings (50–130 cm height). Lines represent fitted generalized linear models with binomial distribution for visualization of general trends. See Table 2 for zero- and one-inflated regression models and their parameters
3.3 Relationships with understory vegetation
Understory vegetation was composed of 136 taxa of plants, and 36 of them occurred in more than ten plots. Species composition of understory vegetation was organized mainly along a soil pH gradient (Fig. 3; Table 3), while the importance of other factors was lower. The second gradient (DCA2) was correlated with seedling density, whereas densities of short saplings were correlated with DCA1. Density of tall saplings was not correlated with either gradient. Abundance of 22 species was statistically significantly correlated with natural regeneration density of P. abies (Table 4). Most of the species were positively associated with density of natural P. abies regeneration, especially species typical of natural, acidophilous coniferous forests (Vaccinio-Piceetea class) and other bryophytes. In the latter group, the exceptions were species typical of more fertile forests: Atrichum undulatum and Mnium hornum. The highest correlations were with Dicranum polysetum for seedlings (0.391), with Vaccinium myrtillus for short saplings (0.594), and with Avenella flexuosa for tall saplings (0.371).

Detrended correspondence analysis (DCA) of understory vegetation. Points represent species; labels (four first letters from genus and species names) are provided for species occurring in at least ten plots. Eigenvalues of axes: DCA1 = 0.4458 and DCA2 = 0.2157. Fitness of vectors representing natural P. abies regeneration density and environmental parameters studied are presented in Table 3
4 Discussion
4.1 Study limitations
The limited amount of variance explained by our models seems to be the most important drawback of this study. This reflects the high variability of natural regeneration and suggests that other factors are also involved in environmental controls of P. abies regeneration, e.g., deer browsing, which is difficult to account for in models using small study plots. However, P. abies is rarely browsed by ungulates (Pröll et al. 2015; Bodziarczyk et al. 2017; Vacek 2017); thus, the lack of deer browsing proxy in the analyses should not have undermined the results. Another factor may be microsite heterogeneity, e.g., local hollows, microrelief, and logs. Local hollows were microsites colonized mainly by sphagna, which are correlated with natural regeneration density of P. abies (Table 2) and indicators of increased moisture (Rydin et al. 2006). Microrelief modifies height growth by impacts on water retention and exposure to soil erosion (Jonášová and Prach 2004; Vacek et al. 2017) as well as light availability Czortek et al. (2018a). Logs are also suitable regeneration niches, due to their ability of water storage (Holeksa et al. 2007; Chećko et al. 2015). Although spatial distribution of microsites was not accounted for in the models, due to systematic sampling, we may assume that their effects were randomly distributed within the data, or—at least—not affected by an observer’s preferences.
Although our study represents a typical empirical analysis of site-specific observations, our conclusions can be applied to other sites. In Central Europe, there are a lot of P. abies forests growing in low elevations outside the species’ ecological optimum (Ellenberg 1988; Caudullo et al. 2016). These sites have usually been under long-term forest management, which influences soil conditions. We provided measurements of conditions in which the study was conducted, which allows our models to be applied in different conditions. Our study was not an experiment and some unknown factors could have influenced the results (see paragraph above). However, due to the high number of study sites characterized in a systematic way and low possibility of systematic biases, we may assume high confidence for transferability of our results (Schweiger et al. 2016; Bhatta et al. 2012). The least transferable conclusions are related to distance from the riverbank, as its ecological effects are strongly dependent on river morphology. For that reason, this part of the conclusions is the most site specific.
Another disadvantage of our study might be connected with the type of light availability measurements. Because we used DIFN (Machado and Reich 1999), we only evaluated indirect light, not regarding which part of the photon flux was photosynthetically active. We also did not measure direct irradiation, which could also influence the growth of saplings. Regarding models explaining proportions of age-height classes in P. abies regeneration, the best-fit model was for short saplings, which were the most well represented. Despite zero- and one-inflation accounting, low abundance of seedlings and tall saplings influenced model quality. Although our results came from a relatively long river valley, extrapolation of the conclusions has to account for site conditions such as climate, elevation, and soil properties.
4.2 Impact of resource availability
Our study revealed the importance of resource availability for natural regeneration of P. abies. Light availability was of the highest importance for tall saplings, which confirmed the observations of Dovčiak et al. (2008). The persistence of young P. abies in deep shade strongly influences its morphology and leads to “umbrella” forms (Metslaid et al. 2007). Short saplings depended less on DIFN than tall saplings. P. abies is a late-successional, shade-tolerant species (Niinemets and Valladares 2006) with seedlings usually exhibiting higher shade tolerance than mature trees because of a more favorable ratio of carbohydrate-producing vs. carbohydrate-consuming biomass. Thus, light availability has no importance for seedlings, which germinate in all possible sites, although only a small proportion of them are able to survive. Probably for that reason, DIFN had no influence on proportions of age-height classes. A similar trend was reported by Jonášová and Prach (2004), who found similar amounts of natural P. abies regeneration in both uncleared and cleared forest patches. Our previous study (Jagodziński et al. 2017) on natural regeneration of pioneer species (Pinus sylvestris and Betula pubescens) also revealed that seedlings differ in ecological requirements from short saplings. Metslaid et al. (2005) also found that further development stages (i.e., saplings over 1.3 m height) are more prone to competition.
Soil pH affected the density of all age-height classes. Moreover, regression coefficients for soil pH increased with age-height classes and there was a negative trend. Dovčiak et al. (2008) also found a negative relationship between soil pH and young sapling density and a positive relationship for taller saplings. Miina and Saksa (2013) also found lower densities of natural P. abies regeneration in more fertile site types (Oxalis-Maianthemum and Oxalis-Myrtillus) than in less fertile ones (Vaccinium and Calluna). In contrast, in the lowlands of northern Poland, Szydlarski and Modrzyński (2015) found natural regeneration of P. abies more frequently in more fertile sites of mixed-deciduous forests than in mixed-coniferous forests, typical of P. abies. As P. abies is a species typical of higher latitudes and elevations (Caudullo et al. 2016), it is a better competitor in poorer sites. For that reason, low pH is unfavorable for most of P. abies competitors. Soil pH also had low impacts on proportions of tall saplings and short saplings. Higher proportion of short saplings was connected with lower numbers of seedlings and tall saplings. This may indicate suitable sites for regeneration, but not for growth of saplings. In the case of tall saplings, this may be an artifact caused by a low number of plots with numerous tall saplings.
Our study also revealed a low but positive impact of distance from the riverbank on P. abies regeneration density. This result seems to contradict the low drought resistance of P. abies (Ellenberg and Leuschner 2010; Gutowski and Jaroszewicz 2015; Caudullo et al. 2016) and its better performance in wetland sites during bark beetle outbreaks (Jaroszewicz et al. 2017). However, P. abies is also prone to waterlogging (Niinemets and Valladares 2006). This effect may also be connected with higher abundance of understory plants typical of more fertile sites near the riverbank. These species are better competitors, and the nearest neighborhood is a local refugium for them, similar to river valleys in urban ecosystems (Dyderski et al. 2017). Moreover, in the study area, there are local hollows with small peatlands, which are not in the nearest closure to the bank (Pielech et al. 2018). These hollows are usually covered by mosses indicating high moisture (e.g., Polytrichum commune, Sphagnum fallax, Sphagnum girgensohnii) and typical of acidic, poor wetlands (Gunnarsson et al. 2002; Rydin and Jeglum 2006; Wierzcholska et al. 2018), which shows compound effects of soil moisture and acidity. In the age of global changes and predicted contraction of the P. abies range (Sykes et al. 1996; Hanewinkel et al. 2013; Dyderski et al. 2018a), our results indicate conditions of local refugia, where P. abies may still regenerate in a non-natural elevational belt. However, further growth of naturally regenerated P. abies will require control of competition (Metslaid et al. 2005).
4.3 Relationships with understory
Our study revealed mostly positive relationships between natural regeneration of P. abies and understory species cover. Similarly, Dovčiak et al. (2008) also reported positive association of natural regeneration of P. abies and moss and Vaccinium cover, but they found a negative association with grass cover. This may be explained by different grass species—in our study, dominant grasses were Avenella flexuosa and Calamagrostis villosa, non-clump-forming species with relatively thin leaves. In the study of Dovčiak et al. (2008), there were mostly clump-forming species. Negative effects of grasses on natural P. abies regeneration were also found in other studies (Jonášová and Prach 2004; Pröll et al. 2015). Although Jonášová and Prach (2004) found that V. myrtillus cover was negatively correlated with natural P. abies density, the positive correlation between P. abies regeneration density and boreal species cover may confirm its predicted dynamic tendencies.
In conditions of low soil fertility and high acidity, competition, which is typical of more fertile sites, is lower (Tilman 1986). In our study, natural regeneration density of P. abies was negatively correlated with the cover of species typical of deciduous temperate forests (Carpino-Fagetea sylvaticae class), which usually grow on more fertile soils (Ellenberg 1988; Mucina et al. 2016). These species are effective competitors, due to high specific leaf area (Jagodziński et al. 2016), one of the adaptations for light acquisition (Westoby 1998). This speculation is in line with the study of Holeksa et al. (2007), who found increasing density of P. abies saplings with increasing elevation, which is also a proxy for decreasing competition in mountain plant communities (Czortek et al. 2018b).
5 Conclusions
Our study highlighted that in lower mountain sites, natural regeneration of P. abies may reach high densities (higher than plantation norms in Poland—4500 ind. ha−1), despite the fact that P. abies forest is not a potential natural vegetation type on these sites. This indicates that natural regeneration may be an alternative to P. abies planting. In Central Europe, natural low-elevation mountain forests are usually replaced by P. abies plantations. P. abies is capable of self-replacement here, and this fact should be accounted for in cases when the next generation of forest would be coniferous (as a consequence of existing management) or broadleaved (restoration). We found the highest densities of natural regeneration of P. abies on sites with moderate (0.1–0.2) light availability and low pH (3.5–4.5). However, these parameters describe the regeneration niche for short saplings of P. abies rather than conditions suitable for tall saplings and further growth of this species. In the study area, most understory species were typical of poor forest sites and thus did not compete with P. abies regeneration. The exceptions were species typical of more fertile forests, showing affiliation of young P. abies regeneration to the less fertile sites.
Our study showed that in less fertile sites in lower mountain elevations, replacement of mature P. abies may be achieved using natural regeneration. However, for successful regeneration, one needs to maintain moderate canopy closure. If thinning is too intensive, understory herbs may develop excessively and decrease abundance and survival of P. abies seedlings and saplings. These results suggest that maintaining moderate canopy closure may be suitable for natural P. abies regeneration in low-elevation mountain belts.
Data availability
The datasets generated and analyzed during the current study are available in the figshare repository (Dyderski et al. 2018a). The datasets were not peer reviewed.
References
Baraloto C, Goldberg DE, Bonal D (2005) Performance trade-offs among tropical tree seedlings in contrasting microhabitats. Ecology 86:2461–2472. https://doi.org/10.1890/04-1956
Bastl M, Štechová T, Prach K (2009) Effect of disturbance on the vegetation of peat bogs with Pinus rotundata in the Třeboň Basin, Czech Republic. Preslia 81:105–117
Beckage B, Lavine M, Clark JS (2005) Survival of tree seedlings across space and time: estimates from long-term count data. J Ecol 93:1177–1184. https://doi.org/10.1111/j.1365-2745.2005.01053.x
Bhatta KP, Chaudhary RP, Vetaas OR (2012) A comparison of systematic versus stratified-random sampling design for gradient analyses: a case study in subalpine Himalaya, Nepal. Phytocoenologia 42:191–202. https://doi.org/10.1127/0340-269X/2012/0042-0519
Bigelow SW, Canham CD (2002) Community organization of tree species along soil gradients in a north-eastern USA forest. J Ecol 90:188–200. https://doi.org/10.1046/j.0022-0477.2001.00655.x
Bodziarczyk J, Zwijacz-Kozica T, Gazda A, Szewczyk J, Frączek M, Zięba A, Szwagrzyk J (2017) Species composition, elevation, and former management type affect browsing pressure on forest regeneration in the Tatra National Park. For Res Pap 78:238–247. https://doi.org/10.1515/frp-2017-0026
Brzeziecki B, Pommerening A, Miścicki S, Drozdowski S, Żybura H (2016) A common lack of demographic equilibrium among tree species in Białowieża National Park (NE Poland): evidence from long-term plots. J Veg Sci 27:460–469. https://doi.org/10.1111/jvs.12369
Caudullo G, Tinner W, de Rigo D (2016) Picea abies in Europe: distribution, habitat, usage and threats. In: San-Miguel-Ayanz J, de Rigo D, Caudullo G, Houston Durrant T, Mauri A (eds) European atlas of forest tree species. Publication Office of the European Union, Luxembourg, pp 114–116
de Chantal M, Granström A (2007) Aggregations of dead wood after wildfire act as browsing refugia for seedlings of Populus tremula and Salix caprea. For Ecol Manag 250:3–8. https://doi.org/10.1016/j.foreco.2007.03.035
Chećko E, Jaroszewicz B, Olejniczak K, Kwiatkowska-Falińska AJ (2015) The importance of coarse woody debris for vascular plants in temperate mixed deciduous forests. Can J For Res 45:1154–1163. https://doi.org/10.1139/cjfr-2014-0473
Chmura D, Sierka E (2007) The invasibility of deciduous forest communities after disturbance: a case study of Carex brizoides and Impatiens parviflora invasion. For Ecol Manag 242:487–495. https://doi.org/10.1016/j.foreco.2007.01.083
Chytrý M (2013) Vegetace České republiky 4. Lesní a křovinná vegetace/Vegetation of the Czech Republic 4. Forest and scrub vegetation. Academia, Praha
Cienciala E, Lindroth A, Čermák J, Hällgren J-E, Kučera J (1992) Assessment of transpiration estimates for Picea abies trees during a growing season. Trees 6:121–127. https://doi.org/10.1007/BF00202427
Curt T, Coll L, Prévosto B, Balandier P, Kunstler G (2005) Plasticity in growth, biomass allocation and root morphology in beech seedlings as induced by irradiance and herbaceous competition. Ann For Sci 62:51–60. https://doi.org/10.1051/forest:2004092
Czortek P, Delimat A, Dyderski MK, Zięba A, Jagodziński AM, Jaroszewicz B (2018b) Climate change, tourism and historical grazing influence the distribution of Carex lachenalii Schkuhr—a rare arctic-alpine species in the Tatra Mts. Sci Total Environ 618:1628–1637. https://doi.org/10.1016/j.scitotenv.2017.10.001
Czortek P, Ratyńska H, Dyderski MK, Jagodziński AM, Orczewska A, Jaroszewicz B (2018a) Cessation of livestock grazing and windthrow drive a shift in plant species composition in the Western Tatra Mts. Tuexenia 37:177–196. https://doi.org/10.14471/2018.38.008
Dech JP, Robinson LM, Nosko P (2008) Understorey plant community characteristics and natural hardwood regeneration under three partial harvest treatments applied in a northern red oak (Quercus rubra L.) stand in the Great Lakes-St. Lawrence forest region of Canada. For Ecol Manag 256:760–773. https://doi.org/10.1016/j.foreco.2008.05.033
Dovčiak M, Hrivnák R, Ujházy K, Gömöry D (2008) Seed rain and environmental controls on invasion of Picea abies into grassland. Plant Ecol 194:135–148. https://doi.org/10.1007/s11258-007-9280-2
Dyderski MK, Tyborski J, Jagodziński AM (2017) The utility of ancient forest indicator species in urban environments: a case study from Poznań, Poland. Urban For Urban Green 27:76–83. https://doi.org/10.1016/j.ufug.2017.06.016
Dyderski M K, Gazda A, Hachułka M, Horodecki P, Kałucka I, Kamczyc J, Malicki M, Pielech R, Smoczyk M, Skorupski M, Wierzcholska S, Jagodziński AM (2018a). Picea abies natural regeneration dataset. V1. figshare. [Dataset]. https://doi.org/10.6084/m9.figshare.6683447
Dyderski MK, Paź S, Frelich LE, Jagodziński AM (2018b) How much does climate change threaten European forest tree species distributions? Glob Change Biol 24:1150–1163. https://doi.org/10.1111/gcb.13925
Eckstein J, Leuschner HH, Bauerochse A (2011) Mid-Holocene pine woodland phases and mire development—significance of dendroecological data from subfossil trees from northwest Germany. J Veg Sci 22:781–794. https://doi.org/10.1111/j.1654-1103.2011.01283.x
Ellenberg H (1988) Vegetation ecology of Central Europe. Cambridge University Press, Cambridge
Ellenberg H, Leuschner C (2010) Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht. UTB, Stuttgart
Emborg J (1998) Understorey light conditions and regeneration with respect to the structural dynamics of a near-natural temperate deciduous forest in Denmark. For Ecol Manag 106:83–95. https://doi.org/10.1016/S0378-1127(97)00299-5
Euro+Med (2006) The Euro+Med PlantBase—the information resource for Euro-Mediterranean plant diversity. http://ww2.bgbm.org/EuroPlusMed/. Accessed 29 Dec 2017
Felton A, Lindbladh M, Brunet J, Fritz Ö (2010) Replacing coniferous monocultures with mixed-species production stands: an assessment of the potential benefits for forest biodiversity in northern Europe. For Ecol Manag 260:939–947. https://doi.org/10.1016/j.foreco.2010.06.011
Grodzki W (2016) Mass outbreaks of the spruce bark beetle Ips typographus in the context of the controversies around the Bialowieza primeval forest. For Res Pap 77:324–331. https://doi.org/10.1515/frp-2016-0033
Gunnarsson U, Malmer N, Rydin H (2002) Dynamics or constancy in Sphagnum dominated mire ecosystems? A 40-year study. Ecography 25:685–704. https://doi.org/10.1034/j.1600-0587.2002.250605.x
Gutowski JM, Jaroszewicz B (2015) Zmiany udziału świerka pospolitego w drzewostanach Puszczy Białowieskiej w kontekście dynamiki liczebności kornika drukarza Ips typographus. In: Wikło A (ed) Stan ekosystemów leśnych Puszczy Białowieskiej. Ogólnopolska Konferencja Naukowa Ministerstwa Środowiska i Generalnej Dyrekcji Lasów Państwowych. Warszawa, 28 października 2015. Centrum Informacyjne Lasów Państwowych, Warszawa, pp 87–108
Hanewinkel M, Cullmann DA, Schelhaas M-J, Nabuurs G-J, Zimmermann NE (2013) Climate change may cause severe loss in the economic value of European forest land. Nat Clim Chang 3:203–207. https://doi.org/10.1038/nclimate1687
Hill MO, Gauch HG (1980) Detrended correspondence analysis: an improved ordination technique. Vegetatio 42:47–58. https://doi.org/10.1007/BF00048870
Holeksa J, Saniga M, Szwagrzyk J, Dziedzic T, Ferenc S, Wodka M (2007) Altitudinal variability of stand structure and regeneration in the subalpine spruce forests of the Pol’ana biosphere reserve, Central Slovakia. Eur J For Res 126:303–313. https://doi.org/10.1007/s10342-006-0149-z
Iszkuło G, Nowak-Dyjeta K, Sekiewicz M (2014) Influence of initial light intensity and deer browsing on Taxus baccata saplings: a six years field study. Dendrobiology 71:93–99. https://doi.org/10.12657/denbio.071.009
Jagodziński AM, Dyderski MK, Rawlik K, Kątna B (2016) Seasonal variability of biomass, total leaf area and specific leaf area of forest understory herbs reflects their life strategies. For Ecol Manag 374:71–81. https://doi.org/10.1016/j.foreco.2016.04.050
Jagodziński AM, Horodecki P, Rawlik K, Dyderski MK (2017) Do understorey or overstorey traits drive tree encroachment on a drained raised bog? Plant Biol 19:571–583. https://doi.org/10.1111/plb.12569
Janzen DH (1970) Herbivores and the number of tree species in tropical forests. Am Nat 104:501–528
Jaroszewicz B, Bobiec A, Eycott AE (2017) Lack of demographic equilibrium indicates natural, large-scale forest dynamics, not a problematic forest conservation policy—a reply to Brzeziecki et al. J Veg Sci 28:218–222. https://doi.org/10.1111/jvs.12458
Jonášová M, Prach K (2004) Central-European mountain spruce (Picea abies (L.) Karst.) forests: regeneration of tree species after a bark beetle outbreak. Ecol Eng 23:15–27. https://doi.org/10.1016/j.ecoleng.2004.06.010
Kabała C, Chodak T, Bogacz A, Łabaz B, Jezierski P, Kaszubkiewicz B, Glina B (2011) Przestrzenne zróżnicowanie gleb i siedlisk Parku Narodowego Gór Stołowych. In: Chodak T, Kabała C, Kaszubkiewicz B, Migoń P, Wojewoda J (eds) Geoekologiczne warunki środowiska przyrodniczego Parku Narodowego Gór Stołowych: (monografia), WIND. Wrocław, pp 141–168
Kawaletz H, Mölder I, Annighöfer P, Terwei A, Zerbe S, Ammer C (2014) Back to the roots: how do seedlings of native tree species react to the competition by exotic species? Ann For Sci 71:337–347. https://doi.org/10.1007/s13595-013-0347-z
Knight KS, Oleksyn J, Jagodzinski AM, Reich PB, Kasprowicz M (2008) Overstorey tree species regulate colonization by native and exotic plants: a source of positive relationships between understorey diversity and invasibility. Divers Distrib 14:666–675. https://doi.org/10.1111/j.1472-4642.2008.00468.x
Kučerová A, Rektoris L, Štechová T, Bastl M (2008) Disturbances on a wooded raised bog—how windthrow, bark beetle and fire affect vegetation and soil water quality? Folia Geobot 43:49–67
Löf M, Bolte A, Welander NT (2005) Interacting effects of irradiance and water stress on dry weight and biomass partitioning in Fagus sylvatica seedlings. Scand J For Res 20:322–328. https://doi.org/10.1080/02827580500201593
Lorimer CG, Chapman JW, Lambert WD (1994) Tall understorey vegetation as a factor in the poor development of oak seedlings beneath mature stands. J Ecol 82:227–237. https://doi.org/10.2307/2261291
Machado J-L, Reich PB (1999) Evaluation of several measures of canopy openness as predictors of photosynthetic photon flux density in deeply shaded conifer-dominated forest understory. Can J For Res 29:1438–1444. https://doi.org/10.1139/x99-102
Martínez-García LB, Pietrangelo O, Antunes PM (2015) Parent tree distance-dependent recruitment limitation of native and exotic invasive seedlings in urban forests. Urban Ecosyst 19:969–981. https://doi.org/10.1007/s11252-015-0507-z
Metslaid M, Ilisson T, Vicente M, Nikinmaa E, Jõgiste K (2005) Growth of advance regeneration of Norway spruce after clear-cutting. Tree Physiol 25:793–801
Metslaid M, Jõgiste K, Nikinmaa E, Moser WK, Porcar-Castell A (2007) Tree variables related to growth response and acclimation of advance regeneration of Norway spruce and other coniferous species after release. For Ecol Manag 250:56–63. https://doi.org/10.1016/j.foreco.2007.03.009
Migoń P, Latocha A, Parzóch K, Kasprzak M, Owczarek P, Witek M, Pawlik Ł (2011) Współczesny system morfogenetyczny Gór Stołowych. In: Chodak T, Kabała C, Kaszubkiewicz J, Migoń P, Wojewoda J (eds) Geoekologiczne warunki środowiska przyrodniczego Parku Narodowego Gór Stołowych: (monografia), WIND. Wrocław, pp 1–52
Miina J, Saksa T (2013) Predicting establishment of tree seedlings in regeneration areas of Picea abies in southern Finland. Balt For 19:187–200
Moran MD (2003) Arguments for rejecting the sequential Bonferroni in ecological studies. Oikos 100:403–405. https://doi.org/10.1034/j.1600-0706.2003.12010.x
Mucina L, Bültmann H, Dierßen K, Theurillat J-P, Raus T, Čarni A, Šumberová K, Willner W, Dengler J, García RG, Chytrý M, Hájek M, Di Pietro R, Iakushenko D, Pallas J, Daniëls FJA, Bergmeier E, Santos Guerra A, Ermakov N, Valachovič M, Schaminée JHJ, Lysenko T, Didukh YP, Pignatti S, Rodwell JS, Capelo J, Weber HE, Solomeshch A, Dimopoulos P, Aguiar C, Hennekens SM, Tichý L (2016) Vegetation of Europe: hierarchical floristic classification system of vascular plant, bryophyte, lichen, and algal communities. Appl Veg Sci 19:3–264. https://doi.org/10.1111/avsc.12257
Mueller KE, Eisenhauer N, Reich PB, Hobbie SE, Chadwick OA, Chorover J, Dobies T, Hale CM, Jagodziński AM, Kałucka I, Kasprowicz M, Kieliszewska-Rokicka B, Modrzyński J, Rożen A, Skorupski M, Sobczyk Ł, Stasińska M, Trocha LK, Weiner J, Wierzbicka A, Oleksyn J (2016) Light, earthworms, and soil resources as predictors of diversity of 10 soil invertebrate groups across monocultures of 14 tree species. Soil Biol Biochem 92:184–198. https://doi.org/10.1016/j.soilbio.2015.10.010
Müller J, Noss RF, Bussler H, Brandl R (2010) Learning from a “benign neglect strategy” in a national park: response of saproxylic beetles to dead wood accumulation. Biol Conserv 143:2559–2569. https://doi.org/10.1016/j.biocon.2010.06.024
Naudts K, Chen Y, McGrath MJ, Ryder J, Valade A, Otto J, Luyssaert S (2016) Europe’s forest management did not mitigate climate warming. Science 351:597–600. https://doi.org/10.1126/science.aad7270
Niinemets Ü, Valladares F (2006) Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol Monogr 76:521–547. https://doi.org/10.1890/0012-9615(2006)076[0521:TTSDAW]2.0.CO;2
Ochyra R, Bednarek-Ochyra H, Żarnowiec J (2003) Census catalogue of Polish mosses. Biodiversity of Poland Vol. 3. W. Szafer Institute of Botany, Institute of Nature Conservation, Polish Academy of Sciences, Kraków
Oksanen J, Blanchet FG, Kindt R, Legendre P, Michin PR, O’Hara RB, Simpson GL, Solymos P, Henry M, Stevens H, Wagner H (2018) “vegan” 2.3.3.—community ecology package
Ospina R, Ferrari SLP (2010) Inflated beta distributions. Stat Pap 51:111–126. https://doi.org/10.1007/s00362-008-0125-4
Ozolinčius R, Mikšys V, Stakénas V (1996) Above-ground phytomass and light regime in Norway spruce stands planted with different initial density. Biomass Bioenergy 11:201–206
Pielech R, Malicki M, Smoczyk M, Jagodziński AM, Dyderski MK, Horodecki P, Wierzcholska S, Skorupski M, Kamczyc J, Kałucka I, Hachułka M, Gazda A (2018) Plant communities of the Czerwona Woda River valley (Stołowe Mountains National Park). For Res Pap 79:181–197. https://doi.org/10.2478/frp-2018-0019
Pröll G, Darabant A, Gratzer G, Katzensteiner K (2015) Unfavourable microsites, competing vegetation and browsing restrict post-disturbance tree regeneration on extreme sites in the northern calcareous Alps. Eur J For Res 134:293–308. https://doi.org/10.1007/s10342-014-0851-1
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rigby RA, Stasinopoulos DM (2005) Generalized additive models for location, scale and shape. J R Stat Soc Ser C Appl Stat 54:507–554. https://doi.org/10.1111/j.1467-9876.2005.00510.x
Robakowski P, Bielinis E (2011) Competition between sessile oak (Quercus petraea) and black cherry (Padus serotina): dynamics of seedlings growth. Pol J Ecol 59:297–306
Rydin H, Jeglum JK (2006) The biology of peatlands. Oxford University Press
Rydin H, Gunnarsson U, Sundberg S (2006) The role of Sphagnum in peatland development and persistence. In: Wieder RK, Vitt DH (eds) Boreal peatland ecosystems. Springer, Berlin Heidelberg, pp 47–65
Schweiger AH, Irl SDH, Steinbauer MJ, Dengler J, Beierkuhnlein C (2016) Optimizing sampling approaches along ecological gradients. Methods Ecol Evol 7:463–471. https://doi.org/10.1111/2041-210X.12495
Seidl R, Schelhaas M-J, Lexer MJ (2011) Unraveling the drivers of intensifying forest disturbance regimes in Europe. Glob Change Biol 17:2842–2852. https://doi.org/10.1111/j.1365-2486.2011.02452.x
Seidl R, Schelhaas M-J, Rammer W, Verkerk PJ (2014) Increasing forest disturbances in Europe and their impact on carbon storage. Nat Clim Chang 4:806–810. https://doi.org/10.1038/nclimate2318
Stenberg P, Linder S, Smolander H, Flower-Ellis J (1994) Performance of the LAI-2000 plant canopy analyzer in estimating leaf area index of some Scots pine stands. Tree Physiol 14:981–995
Sykes MT, Prentice IC, Cramer W (1996) A bioclimatic model for the potential distributions of north European tree species under present and future climates. J Biogeogr 23:203–233
Szweykowski J (2006) An annotated checklist of Polish liverworts and hornworts. Biodiversity of Poland. Vol. 4. W. Szafer Institute of Botany, Institute of Nature Conservation, Polish Academy of Sciences, Kraków
Szydlarski M, Modrzyński J (2015) Increase of natural regeneration area of Norway spruce (Picea abies L. Karst.) in the Kaszuby Lake District during the decade 2002–2012. For Res Pap 76:66–72. https://doi.org/10.1515/frp-2015-0007
Tarka R, Buczyński S, Modelska M, Olichwer T, Staśko P (2011) Hydrogeologia obszaru Parku Narodowego Gór Stołowych. In: Chodak T, Kabała C, Kaszubkiewicz B, Migoń P, Wojewoda J (eds) Geoekologiczne warunki środowiska przyrodniczego Parku Narodowego Gór Stołowych: (monografia), WIND. Wrocław, pp 113–140
Tilman D (1986) Resources, competition and the dynamics of plant communities. In: Crawley M (ed) Plant ecology. Blackwell Scientific Publications, Oxford, pp 51–75
Trouet V, Van Oldenborgh GJ (2013) KNMI climate explorer: a web-based research tool for high-resolution paleoclimatology. Tree-Ring Res 69:3–13. https://doi.org/10.3959/1536-1098-69.1.3
Vacek Z (2017) Structure and dynamics of spruce-beech-fir forests in nature reserves of the Orlické hory Mts. in relation to ungulate game. Cent Eur Fore J 63:23–34. https://doi.org/10.1515/forj-2017-0006
Vacek Z, Bulušek D, Vacek S, Hejcmanová P, Remeš J, Bílek L, Štefančík I (2017) Effect of microrelief and vegetation cover on natural regeneration in European beech forests in Krkonoše national parks (Czech Republic, Poland). Aust J For Sci 134:75–96
Westoby M (1998) A leaf-height-seed (LHS) plant ecology strategy scheme. Plant Soil 199:213–227. https://doi.org/10.1023/A:1004327224729
Wierzcholska S, Dyderski MK, Pielech R, Gazda A, Smoczyk M, Malicki M, Horodecki P, Kamczyc J, Skorupski M, Hachułka M, Kałucka I, Jagodziński AM (2018) Natural forest remnants as refugia for bryophyte diversity in a transformed mountain river valley landscape. Sci Total Environ 640–641:954–964. https://doi.org/10.1016/j.scitotenv.2018.05.340
Zeileis A, Kleiber C, Jackman S (2008) Regression models for count data in R. J Stat Soft 27:1–21. https://doi.org/10.18637/jss.v027.i08
Zeppenfeld T, Svoboda M, DeRose RJ, Heurich M, Müller J, Čížková P, Starý M, Bače R, Donato DC (2015) Response of mountain Picea abies forests to stand-replacing bark beetle outbreaks: neighbourhood effects lead to self-replacement. J Appl Ecol 52:1402–1411. https://doi.org/10.1111/1365-2664.12504
Acknowledgements
We are thankful to Mrs. Natalia Czapiewska, Mrs. Michalina Grabowska, Mrs. Natalia Smarul, Mr. Karol Tomczak, and Mrs. Patrycja Wajer for their assistance during the fieldwork. We also would like to thank Dr. Lee E. Frelich (Department of Forest Resources, University of Minnesota, USA) for linguistic revision of the manuscript. We are grateful to two anonymous reviewers for their useful remarks.
Funding
The study was financially supported by the Polish Forest Fund, General Directorate of State Forests, Warsaw, Poland (research project: “Recognition and assessment of the current state of natural habitats in the vicinity of Czerwona Woda River”).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Before establishing the study sites, the authors obtained approval from the Director of the Stołowe Mountains National Park.
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Andreas Bolte
Contribution of the co-authors AMJ conceived the idea and developed the methodology with the collaboration of AG, MH, PH, ILK, JK, RP, and MSk. AMJ acquired funds for the implementation of the research and coordinated the research project. MKD led the data collection in the field and analyzed the data. AG, MM, MSm, and RP performed the vegetation surveys and SW determined the identities of bryophyte species. AG, PH, JK, MSk, and SW reviewed the literature. MKD and AMJ led the manuscript preparation with substantial contributions from all the co-authors. All of the authors contributed substantially to the methodology development, data collection, and critical revisions of the manuscript. All of the authors approved the final version of the manuscript submitted.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Dyderski, M.K., Gazda, A., Hachułka, M. et al. Impacts of soil conditions and light availability on natural regeneration of Norway spruce Picea abies (L.) H. Karst. in low-elevation mountain forests. Annals of Forest Science 75, 91 (2018). https://doi.org/10.1007/s13595-018-0775-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s13595-018-0775-x