
Abstract
Specialist ecosystems have developed on small areas (< 1 ha) of alkaline (pH 7–10) soil-free disturbed substrates in semiarid areas of southern New Zealand, where rare endemic plants are being outcompeted by adventive species as soil develops. We have conducted a pilot study for soil and weed removal at seven sites to increasing habitat availability for the rare plants. Six sites are on impermeable clay-altered schist where evaporation of marine aerosols in rain has formed localised NaCl-rich encrustations. These bare substrates host endemic salt-tolerant plants (halophytes), especially where electrical conductivity (EC) exceeds 1 mS/cm and excludes most weeds. One site is on limestone and associated erosional residues, where endemic calciflora have evolved on substrates that have pH of 7–8 and variable EC. Soil and weeds were removed to a depth of ~ 0.5 m, and the study plots were monitored for changes in substrate salinity, pH and plant colonisation for up to 1.5 years. The most successful saline plot, an abandoned mining area, had initial halophyte colonisation after 4 months, and after a year, the halophytes had achieved > 50% substrate coverage. The other saline plots also had lesser halophyte colonisation and more weed incursion. Bare limestone talus that had weeds allowed successful calciphyte germination. Parts of the limestone site where weeds and soil were removed had no calciphyte colonisation but significant weed development after 6 months. Our results suggest that soil removal is a viable management option for enhancing ecosystems that require bare substrates, preferably with deeper (~ 1 m) excavation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Rehabilitation of disturbed land is an issue of increasing importance, especially for mine sites, agricultural areas and some urban settings, as growing human populations become more environmentally concerned (Banning et al. 2011; Meuser 2012; Clewell and Aronson 2013; Bolan et al. 2017; Hancock et al. 2020). One of the most common approaches to rehabilitation is addition of soil to the disturbed surface areas, either from pre-disturbance stockpiles, importation of soil from elsewhere, or addition of organic material such as biosolids (Sere et al. 2008; Banning et al. 2011; Meuser 2012; Munshower 2018; Hancock et al. 2020). This approach can be successful at establishing a vegetation cover and limiting erosional sediment runoff (Meuser 2012; Munshower 2018; Hancock et al. 2020). Consequently, this approach has become the principal strategy for rehabilitation of disturbed land in most settings (Banning et al. 2011; Meuser 2012; Munshower 2018). Even low-fertility dryland environments can be rehabilitated by enhancing the biological components of the surface zone (Mengual et al. 2014; Weber et al. 2016; Chock et al. 2019).
Despite the widespread successes of this rehabilitation strategy, it typically leads to a uniform landscape and associated vegetation, with limited biodiversity and variable long term (decades) success (Hobbs et al. 2009; Tropek et al. 2012; Prach and Tolvanen 2016). Consequently, some researchers have proposed rehabilitation strategies that involve natural processes without soil intervention, to encourage greater biodiversity in the end result (Bradshaw 2000; Hodacova and Prach 2003; Prach and Hobbs 2008; Tropek et al. 2012, 2013; Prach and Tolvanen 2016; Sebelikova et al. 2016). Further, it has been suggested that excessive intervention during rehabilitation of disturbed land does not support sustainable revegetation like natural rehabilitation processes, which should be encouraged rather than thwarted (Holliday and Johnson 1979; Bizoux et al. 2004; Valente et al. 2012; Tropek et al. 2012, 2013; Anawar 2015; Haigh 2018). Hence, greater variation of rehabilitation strategies may be desirable to enhance biodiversity and achieve a sustainable result (Valente et al. 2012; Tropek et al. 2012, 2013; Doley and Audet 2013; Prach and Tolvanen 2016). This is particularly relevant for rehabilitation of land that has distinctive substrate chemistry, that may host specifically adapted and tolerant ecosystems (e.g. Valente et al. 2012; Alday et al. 2016), and that is the basis for the current study (Rufaut et al. 2018).

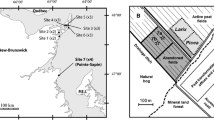
Location, regional geology and regional climate for the sites in this study. a Topographic hillshade image of the South Island of New Zealand. b Annual rainfall map for the southern South Island (NIWA 2021) showing the principal features relevant to the rain shadow and the studied sites. c Geological map for sites developed in this study on clay-altered basement unconformity zone. d Sketch cross section through the Kurow limestone site described in this study
In this study, we document pilot trials of a novel strategy for enhancement of geochemically distinctive soil-free substrates that have been exposed by natural and human-related physical processes (Fig. 2a–d). Our strategy involves establishment of bare ground, mainly by removal of soil that has been accumulating naturally on small areas of these distinctive geological substrates in association with adventive weed species colonisation (Fig. 2a–d). This apparently contrary approach has the advantage of facilitating the survival, growth and surface coverage of existing specialist endemic vegetation (halophytes and calciflora) that would otherwise be excluded from these areas by competition. We focus on the underlying geological and geochemical nature of the specialist substrates and the initial endemic plant colonisation (Fig. 2a–d). We then document the preliminary successes and failures of specific trials of our soil removal strategy in enhancing local indigenous biodiversity (Fig. 2e, f).

Natural site photographs and cross-section sketches to show the rationale for study, involving soil removal to enhance rare plant habitats. a Photograph of a halophytic salt pan with encroaching soil and weeds. b Photograph of a limestone outcrop and erosional debris cover, with calciflora being crowded out by encroaching weeds. c Sketch cross section through a typical schist basement site and erosional pan, showing strategy to enhance halophyte habitat. d Sketch cross section through limestone outcrop and talus, with strategy to enhance calciflora habitat. e Close view of clay-altered schist substrate from which soil and weeds had been removed, after 6 months, with rapidly colonising halophytes (Atriplex buchananii). f Close view of weeded limestone and talus debris after 6 months, with bare ground maintained but only exotic weeds sprouting at that time
General setting and rationale
Climate, geology and mineralogy
A prominent rain shadow has developed to the east of actively rising mountains (Fig. 1b; Chamberlain et al. 1999). Abundant rain (up to 12 m/year) falls on the western side of the mountains as prevailing westerly winds rise at the mountain barrier (Fig. 1b), and warm dry fȍhn-style winds descend on the eastern side to contribute to high evaporation in the rain shadow (Chamberlain et al. 1999; Craw et al. 2013). Consequently, the rain shadow area, the focus of this study, is a region of low erosion rates with widespread formation of evaporative salts on and in surface deposits (Craw et al. 2013, 2022). Rain events in the rain shadow are distributed throughout the year, with no distinct wet and dry seasons (Fig. 3a). However, surface evaporation rates are strongly seasonal, with low rates in winter and high rates in summer (Fig. 3b).

General characterisation of study sites, substrates and substrate water end-members, compiled and summarised from Craw et al. (2022) and references therein. a Daily rainfall data for the rainshadow area in Fig. 1c for the time including the period of this study (NIWA 2021). b Monthly evapotranspiration for 2021, at a site ~ 30 km NE of schist sites (Fig. 1c; ORC 2021). c Contrasting EC of Ca-bicarbonate and NaCl waters. Model NaCl–EC relationship calculated from empirical Marion–Babcock equation (Sposito 2016). d EC variations over time for the transect at the Conroys reference site (Fig. 1c). e EC and pH data for limestone-hosted calciflora substrates, as determined from field slurry measurements (5 min) and laboratory leachates (6 weeks). f Comparison of pH and EC of typical Na-Cl saline substrates (Rogers et al. 2000) and schist basement Ca-carbonate groundwaters in the rain shadow area (Fig. 1b, c), compared to typical regional soils. Generalised pH values for dissolution of calcite and Na-carbonate minerals, and evaporation of seawater (Babel and Schreiber 2014) are indicated. g Combined pH and EC data for clay-altered schist sites developed in this study in the area in Fig. 1c, showing broad site differences
The basement rock for the whole studied area is Mesozoic (~ 200 Ma) schist (Fig. 1a; Table 1; Turnbull 2000; Forsyth 2001). A veneer of Cenozoic (70 Ma–present) sedimentary rocks was deposited on this basement, and the basement and sedimentary veneer have been deformed, uplifted and differentially eroded by late Cenozoic tectonics (Jackson et al. 1996; Landis et al. 2008). Groundwater-driven water–rock interaction occurred along the regional unconformity between basement and sedimentary veneer, resulting in variable and locally intense clay alteration of the basement rocks immediately beneath the unconformity (Chamberlain et al. 1999; Craw et al. 2013, 2023). The sedimentary veneer in the centre of the schist belt (Fig. 1a, c) consists of Miocene nonmarine deposits, including gold-bearing conglomerates, whereas marine sediments including some prominent limestone predominate on the flanks of the belt (Fig. 1a, d).
Remnants of Cenozoic sediments and the underlying clay-altered unconformity zone are preserved on the lower slopes of many ranges, at valley margins (Fig. 1c, d), and include the sites involved in this study (Fig. 1c, d). Mineralogy of all the substrates in this study is dominated by that of the schist basement which formed the source for overlying debris and Cenozoic sediments (Table 1). Clay-altered schist basement and its erosional products contain abundant kaolinite and only minor calcite, whereas the limestone is dominated by calcite with subordinate detrital silicates (Table 1). These rock-sourced minerals are variably augmented by surficial evaporative precipitates from marine aerosols and shallow groundwater dissolution processes (Fig. 2c, d; Table 1).
Rationale for study
Small areas (hectares) of bare, soil-free geological substrates that were exposed in the rain shadow were impermeable to rain water drainage so that evaporative salts form from marine aerosols with variable amounts of dissolved components from the underlying rocks (Craw et al. 2022). Schist-derived clay surfaces have been colonised through the late Cenozoic by endemic salt-tolerant plants (halophytes; Allen and McIntosh 1997; Rogers et al. 2000; Rufaut et al. 2018) that likely originated from coastal populations. Likewise, limestone decomposition and localised erosion in the rain shadow has yielded bare, soil-free substrates on which endemic limestone-tolerant plants (calciflora) have evolved in the late Cenozoic (Rogers et al. 2018; Heenan and McGlone 2019).
The halophyte and calciflora ecosystems are made up of low-growing (< 10 cm high) species that thrive on their geochemically distinct substrates and dynamic surficial environments with active erosion and redeposition of their soil-free substrates (Fig. 2c, d; Supplementary Fig. 1). Halophytes occupy both salt pans and associated drainage features and it is the former that our study has concentrated on. Incursion of high-biomass adventive plant species is being facilitated by rain-washed and wind-blown sediment coatings on the specialised substrates (Fig. 2a–d). This process of encroachment onto the bare substrates is creating new soil cover and stabilising surface environments from active erosion. Essentially, these processes are crowding out the halophytes and calciflora that are now restricted to soil-free micro-habitats in a wider weedy matrix. Consequently, these endemic species are now facing landscape-wide extinction in these inland areas (Allen and McIntosh 1997; Miller and Duncan 2003; Rogers et al. 2018; Rufaut et al. 2018).
The aim of this project is to conduct a pilot study in which the encroaching soil and adventive weeds are removed, in the hope that re-exposure of the original bare substrates with dynamic surface processes can be re-established to favour natural expansion of the specialised endemic flora (Fig. 2c, d). No chemical control was conducted. Specific questions addressed in the study are: (a) can soil removal and related bare substrate re-establishment facilitate expansion of dryland areas occupied by endemic halophytes and calciphytes; (b) what is the time scale for recovery of the distinctive surface chemistry after disturbance; and (c) how is this recovery affected by seasonal variations in that chemistry. Because this study was testing a new concept in general, we sought wide diversity of site substrates and topography, rather than aiming for replication of specific parameters that would accompany a more focused study.
Dynamic soil-free substrates and endemic plants
The most actively eroding substrate surfaces remain unvegetated, but debris derived from erosion, typically accumulating at the bottom of eroding slopes, can be colonised by halophytes or calciflora (Fig. 2a–d; Supplementary Fig. 1; Rufaut et al. 2018). Inland halophytes are particularly resilient in environments where repeated erosional disturbance redistributes saline substrate, including partial or even complete burial by rain-washed clay (Supplementary Fig. 1; Rufaut et al. 2018). Calciflora colonise limestone areas in active limestone talus aprons and residual quartz sands in localised rock solution cavities and joints (Fig. 2b, d; Supplementary Fig. 1; Rogers et al. 2018). Both ecosystem types are drought-tolerant, with halophytes establishing on dry impermeable clay-rich crusts and calciflora establishing on permeable limestone debris and in fractures in adjacent outcrops (Fig. 2a–d; Supplementary Fig. 1).
The halophytes are not obligate salt-dwellers (Cheeseman 2015), and the most important environmental criterion is exclusion of high-biomass competing species. Even formation of a biocrust on saline bare substrates can exclude some halophytes (Supplementary Fig. 1). Most adventive plant species can be excluded from halophyte habitat by NaCl-dominated salinity equivalent to EC of > 1 mS/cm (Table 1; Fig. 3c, d; Rufaut et al. 2018) and this may assist on limestone as well (Fig. 3e). Elevated pH is also a factor that may discourage competing species and most halophyte sites have pH > 8. Halophytes can tolerate pH > 10 that arises where Na-carbonate is relatively abundant (Fig. 3f; Table 1; Craw et al. 2022), and this high pH may help to exclude competing species as well. We focused on two particular rare halophyte species that are still relatively widespread on salt pans: the herb Buchanan orache Atriplex buchananii and the salt grass Puccinellia raroflorens. Similarly, the calciflora typically occurs on sites with pH ~ 8 and they may be tolerant of localised evaporative NaCl-dominated salinity as well (Table 1). We used the presence of the rare forb, Chaerophyllum basicola, as an indicator for limestone habitat conditions. As for halophytes, the absence of high-biomass competing species in a dynamic physical environment is critical to the survival of the calciflora (Miller and Duncan 2003; Rogers et al. 2018).
Despite the apparent inhospitality of soil-free substrates on which halophytes and calciflora can live, there are adequate macronutrients in the rocks from which the substrates are derived (Table 1). Potassium is abundant in weathering muscovite, especially in basement-derived substrates, and muscovite is also a source for nitrogen (Table 1). Phosphorus occurs in apatite in schist and schist-derived debris, and this dissolves only slowly in the surficial environment but groundwater-mobilised phosphate also occurs as more soluble vivianite (Table 1). Francolite is a more soluble form of apatite and is an important potential phosphate source from the Cenozoic limestone sequence (Hutton and Seelye 1942). Other major cations are readily available from basement, limestone and/or marine aerosol-derived salts (Table 1).
Methods
Studied sites
Six study plots on clay-altered basement rocks were cleared of soil and weeds in areas where halophyte communities still occur. The initial four of these plots were developed in winter 2020 in the Chapman Road area (Fig. 1c), near to where a scientific reserve for halophytes has been created on land that was disturbed by historical gold mining (Druzbicka et al. 2015; Rufaut et al. 2018; Craw et al. 2023). The following autumn, a fifth plot was developed at Galloway, and a sixth at Crawford Hills (Fig. 1c), also in the vicinity of halophyte communities. Limestone outcrops near Kurow (Fig. 1a, d) were cleared of soil and weeds to provide more geological and chemical diversity of substrates in the study site, and a different set of plant species (calciflora). One section of limestone surface was completely stripped of soil and weeds, and depressions were back-filled with nearby limestone talus material extracted from below 40 cm depth to exclude weed seeds. Some additional areas were weeded by hand around established calciflora (Supplementary Fig. 6).
Soil and weed removal
Site development and subsequent observations occurred over 16 months, from winter (August) 2020 to summer 2021–22 and did not disturb existing specialist flora. Development strategy evolved over that time on the basis of observations at the first sites at Chapman Road. Development of each site was controlled by the unique features of the site, with the principal aim to maximise bare ground that would exclude weeds most effectively. Site soil clearance was done by hand with spades and trowels, up to a depth of 0.5 m dependent on the amount accumulated on top of the desirable geological surface at individual sites. Extra schist-derived clay was excavated and distributed over one Chapman Road site, and extra limestone-derived sandy debris was distributed at the limestone site. At the Crawford site, a tractor-mounted bucket was used to scape several square metres of particularly deep soil build up. The sites were all prepared with firmly packed substrates that sloped gently to facilitate surface water runoff. All of the above-ground adventive plant biomass was removed, with the majority of roots, but any endemic species were specifically avoided.
Developed plots were deliberately irregular in shape outline, with diffuse margins, to maintain a natural appearance and link in with existing specialist plant species locations. Sites were revisited every 1–3 months to monitor surface conditions and any plant colonisation and/or regrowth. The schist clay sites were monitored monthly over the transition from winter to early summer 2021 to document the effects of seasonal changes in substrate chemistry. Follow-up observational visits later in 2022 monitored specialist plant growth and weed incursion.
Site characterisation
The nature of soil-free substrates has been characterised in the vicinity of halophytes in the general areas of this study by previous workers (Table 1; Druzbicka et al. 2015; Law et al. 2016). These data, including substrate leachate chemistry, augmented by our own more recent observations, are used to provide mineralogical, geochemical and physical contexts for the specific test plots created in this study, as outlined in following sections. This previous work has shown that substrate pH and salinity (as determined by electrical conductivity, EC) are the most significant habitat parameters (Fig. 3c–f). These parameters are strongly affected by the mineralogy of the substrates, especially the more soluble evaporative minerals (Fig. 3c, f; Table 1; Craw et al. 2022). In particular, the alkaline pH of the substrates is mainly controlled by halite, calcite and/or Na-carbonates, and EC is mainly controlled by halite (Fig. 3c, f). Ephemeral surface waters are sodic, reflecting the abundance of evaporative halite and alteration of schist albite (Table 1; Craw et al. 2022). In contrast, shallow groundwaters in schist basement and schist-derived debris with less-intense clay alteration are dominated by Ca-bicarbonate dissolved from calcite (Fig. 3f; Craw et al. 2022).
The EC and pH of saline study sites were determined with an Oakton portable meter with separate probes for EC and pH. Probes were calibrated with manufacturer-supplied standard solutions. Samples were collected from sites and analysed in a field building at 18–20 °C. About 20 mL of solids was combined with 40 mL of distilled water and stirred thoroughly before measurements were taken after 4–5 min. At each clay-altered schist site, the EC and pH data were collected periodically from taped transects across the bare ground through the middle of study plots. When plant colonisation had occurred, the EC and pH of substrate beneath well-established specimens were collected to augment these transects. Resultant data are summarised in Fig. 3f.
Limestone substrates were characterised in the field by the same pH and EC methods (Fig. 3e). In addition, a set of eight representative limestone substrates was leached in a laboratory for 6 weeks to obtain a longer-term estimate of chemical variations (Fig. 3e). Leachates were decanted from 200 mL of substrate that was soaked in 2 L of distilled water and analysed by methods described by Craw et al. (2022). Limestone leachates, like the shallow groundwaters in schist basement, have chemistry dominated by Ca-bicarbonate ions (Fig. 3f; Craw et al. 2022), with EC > 1 mS/cm driven by evaporative halite (Fig. 3e).
Mineral identification for salt encrustations was done with X-ray diffraction (XRD) and/or scanning electron microscopy with energy-dispersive analytical facility (SEM–EDX) by methods described in more detail by Druzbicka et al. (2015) and Law et al. (2016). Geochemical modelling of potential mineral precipitates from evaporation of surface waters was carried out with Geochemists Workbench software (www.gwb.com), as documented by Craw et al. (2022).
Reference EC and pH values for target ecosystems
Previous work on sites with halophytes has indicated that an EC value of > 1 mS/cm and pH 7–10 is typical of halophytic ecosystems (above; Law et al. 2016; Rufaut et al. 2018). Hence establishment of these threshold values was a desired result in our pilot studies (Fig. 3c–g), and these values were used for reference at each site. In our on-going monitoring of the study sites, we included adjacent existing soils at both ends of our transects for comparison to the developed bare substrates. All the developed substrates were highly inhomogeneous at the cm scale. Variable micro-effects of mineralogy, permeability and surface and sub-surface water flow were also complexly inter-related, so statistical assumptions on sample independence were not met and statistical evaluations were not carried out.
The effects of different seasons on evaporative salt presence and distribution, and consequently the substrate EC of saline substrates, was unknown before this study. Hence, we established a reference site, Conroys, in the general vicinity of the developed sites (Fig. 1c; Supplementary Fig. 2) to quantify seasonal changes in EC as a result of the seasonal changes in evaporation rates (Fig. 3b). This reference site has bare substrate that was formed during excavation in 2013 for a fence line across an area of clay-altered schist (Supplementary Fig. 2). The site has subsequently developed an impermeable surface with evaporative salts and scattered halophytes have colonised parts of it, although trampling by grazing animals affect their distribution. Substrate EC and pH were measured each metre along a 10 m transect on a monthly basis from winter to summer 2021 (Fig. 3d; Supplementary Fig. 2) in parallel with some of the soil removal site monitoring. The EC ranged from ~ 1 to 10 mS/cm in winter, with a rapid increase in salinity to 10–20 mS/cm in spring (Fig. 3d). The pH of the site is broadly uniform, from pH 7 to 8 (Fig. 3g). Evaporative salts were not visible in winter, but were widespread in early summer.
Limestone substrates at our site were dominated by calcite and no Na-carbonates have been detected (Table 1) so pH of 7–8.5 was typical, compared to regional soils that are generally more acidic (Fig. 3e, f), and this weakly alkaline pH is a useful reference for calciflora habitat. The effects of EC on calciflora ecosystems is poorly known, but dissolution of calcite results in a maximum EC of ~ 1 mS/cm (Fig. 3c, e), and this is a desirable reference target. Locally higher EC values in the limestone substrates result from dissolution of evaporative marine aerosols, especially halite (Fig. 3c, e, f) and this elevated EC helps to limit incursion of weeds. Because of the general uniformity of pH and EC of the limestone substrates compared to the clay-altered schist, fewer monitoring samples were collected.
Results at soil removal sites
Chapman Road white clay site
This site (Fig. 1c) is located at the margin of a prominent historic gold mine sluicing area (Druzbicka et al. 2015), and was also mined for gold. The developed plot is at the foot of a shallow landslide of white schist-derived kaolinitic clay material with some intermixed solid basement debris and had abundant halite-dominated salt encrustations. Some soil and weeds were removed from the site, but most preparation involved addition of a layer (up to 30 cm) of white clay to the surface so that no soil remained exposed (Supplementary Fig. 3). The plot was developed so that a small number of existing halophytes (A. buchananii; Puccinellia raroflorens) were linked into the margins. Site development involved random redistribution of existing salt encrustations. Subsequent results from chemical monitoring along a transect across the site (Fig. 4a) show that the constructed site was highly variable over time and space (Fig. 5a–c). However, substrate EC was between 1 and 10 mS/cm at times, and pH was commonly between 8 and 9 (Figs. 3g, 5a–c).

Drone-generated photomosaics of the two largest sites developed on clay-altered schist basement in the Chapman Road area in August 2020, with transect lines for periodic measurement of EC and pH (Fig. 5). Distributions of colonising halophytes and salt encrustations in September 2021 are indicated. a White clay-rich schist was deposited over a site cleared of weeds and most soil. b Weeds and soil were removed from indicated area to expose hard brown clay-altered schist debris

EC and pH variations at Chapman Road sites, with targeted thresholds (EC > 1 mS/cm; pH > 7) indicated with dashed black lines. a, b EC and pH for transect over time for white clay site. c Spot EC and pH data for ecozones mapped at white and brown sites (Fig. 4a, b) in September 2021. d, e EC and pH for transect over time for brown clay site. f EC and pH for the eastern sites
After 4 months, halophyte establishment by A. buchananii was well under way, covering at least 20% of the area (Fig. 2e). Rainfall through spring had consolidated the new surface to leave armouring quartz fragments and A. buchananii plants were established in intervening clay (Supplementary Fig. 3). After 7 months, halophyte colonisation was even further advanced, with A. buchananii on > 50% of the study plot (Supplementary Fig. 3). After 13 months, in September 2021, almost all the site had some coverage by A. buchananii, with the exception of a steep cliff at the top of the slope, and one patch on the lower eastern side (Fig. 4a). Spot samples taken from ecozones mapped in September 2021 (Fig. 4a) had similar EC and pH to the transect (Fig. 5c), although the zone with no halophytes or weeds had lower EC than the halophyte-bearing zones (Fig. 5c). Despite the generally low-moderate EC along the monitoring transect and within the ecozones In general, numerous salt encrustations had developed on the upper parts of the site by September 2021 (Fig. 4a). These encrustations occurred as small patches (~ 10–30 cm across), and as a 10–30 cm wide zone across the slope (Fig. 4a), in places where sub-surface seepages of water had evaporated. In August 2022, most of the developed site was covered in A. buchananii with only minor extra weed incursions.
Chapman Road brown clay-rich substrate
This site, located 200 m east of the above white clay site, was also sluiced for gold, leaving a sloping surface of brown variably oxidised clay-rich debris derived from the underlying schist basement (Fig. 4b; Supplementary Fig. 4). Halite-dominated salt encrustations were less abundant than at the white clay site (above), and were concentrated towards the base of the slope. Most of the site had developed a post-mining thin (10–50 cm) covering of soil with adventive plant species. Small exposed remnants of the brown clay-altered substrate material hosted scattered halophytes, with abundant A. buchananii and P. raroflorens below the site. Our plot development included removal of the soil to expose impermeable basement and compacted basement-derived material (Fig. 4b; Supplementary Fig. 4). In contrast to the white site, the EC across the exposed site was relatively low (mostly 0.1–1 mS/cm) with some higher EC spots at the time of plot development and remained relatively low (Fig. 5c). The pH was also generally lower than the white site, ranging from ~ 5.5 to 8 throughout the study period (Fig. 5d). Seasonal changes of EC and pH were highly variable, but the EC only rarely rose above 1 mS/cm reference level on the monitoring transect (Figs. 3g, 5d, e).
Scattered halophytes (A. buchananii) had colonised some of the lower parts of the plot after 4 months (December 2020; Supplementary Fig. 4), although they covered only a small proportion overall (< 10%). Some patches of A. buchananii were well established in March 2021 (Supplementary Fig. 4), but site coverage remained < 20% overall. However, after 13 months by September 2021, there were scattered A. buchananii plants over ~ 40% of the plot (Fig. 4b). By that time, some small (~ 10 cm) salt patches had formed near site margins (Fig. 4b). However, the upper portion of the site remained essentially free of salt and halophytes (Fig. 4b). After 2 years, in August 2022, A. buchananii coverage had progressively extended up the developed plot and only the upper 15% was still bare. However, there had also been some expansion of weeds (Thymus vulgaris) on the lower portions. Spot samples from the ecozones mapped in September 2021 showed that the EC and pH were similar between the ecozones and similar to the monitoring transect (Fig. 5c–e).
Chapman Road eastern sites
Two small sites were developed in close proximity (~ 20 m) in August 2020, on white clay that had been extensively disturbed by anthropogenic activities, including gold mining and associated drainage construction. Most of this clay had been excavated from a nearby (5 m) channel. The sites initially had small patches (2–5 m2) of bare ground, surrounded by abundant invasive couch grass, Elymus repens, and reduced cover of A. buchananii. The grass and up to 30 cm of underlying soil and clay were removed. Nevertheless, these small sites proved to be prone to couch grass re-establishment and had to be redeveloped during the course of the study to a greater depth, 50–60 cm (July 2021). The sites retained relatively high salinity and pH (Fig. 5f) compared to the white and brown sites farther west (Fig. 5a–e). Some native, salt-tolerant grass species, Puccinellia stricta, developed around the margins of the study plots during spring of 2021, and minor colonisation by A. buchananii had occurred by early summer. However, couch grass remained a problem throughout this study and much of this weed incursion occurred from remnants of grass roots that were inevitably missed during excavations. Hence, despite the relatively high EC and pH at these sites, weed incursion effectively negated the development activities.
High pH sites at Galloway and Crawford
These two sites (Fig. 1c; Supplementary Fig. 5) were developed on clay-altered basement that has extensive but discontinuous soil-free exposures over several square kilometres. The exposures were not created by mining and natural erosion coupled with high salinity has maintained many open saline pans despite slow encroachment of adventive species. In addition, both these sites are distinctive in that many of the areas of bare ground have persistently high pH ~ 10 (Fig. 3f) irrespective of the EC.
A Galloway site was developed on land adjacent to the broader area of natural saline pans. This site had been disturbed by roading and fence construction, with minor mining in a nearby gully (Supplementary Fig. 5). Steep outcrops of brown clay-altered schist were coated with a thin veneer of angular quartz pebbles, locally encrusted with halite, Na-carbonate and Na-sulphate, and extensive halophyte colonisation had occurred (Supplementary Fig. 5). Halophytes (A. buchananii; P. raroflorens) had become preferentially established on exposures of saline clay, rather than the armouring quartz pebbles. Encroaching weeds at the site had trapped wind-blown silt to form a new soil coating and this combination was displacing halophytes. For this study, in winter 2021, an area of weeds and soil was removed, as well as some adjacent quartz pebble-armoured bare ground, in an area that included outcrop and a downstream clay pan (Supplementary Fig. 5). A monitoring transect down this site showed low EC (< 1 mS/cm) in winter, but this rose in portions of the site in spring (September 2021; Fig. 6a). The pH of the site was highest on the soil-free outcrop and decreased on the constructed clay pan at the base (Fig. 6b). A patch of pre-existing A. buchananii on the outcrop had expanded on to the cleared substrate after 6 months (Supplementary Fig. 5). The first halophytes (P. raroflorens) appeared on the clay pan by September 2021 (Supplementary Fig. 5), and both A. buchananii and P. raroflorens had colonised some edges of the lower pan by early summer. In August 2022, A. buchananii was abundant across much of the site and P. raroflorens was well-established at the base of the site. However, there had also been extensive colonisation by the exotic salt-tolerant weed Plantago coronopus, especially at the base of the site.

EC and pH data over time for the transects down the soil removal sites at the high-pH Galloway (a and b) and Crawford (c and d) localities (Fig. 1c; Supplementary Fig. 5). Targeted EC threshold (> 1 mS/cm) is indicated with dashed black lines
The nearby site on Crawford Hills (Fig. 1c) contained naturally exposed bare ground that was dominated by almost-flat discontinuous white kaolinitic schist-derived clay pans separated by large areas of adventive species (Supplementary Fig. 5). Adventive weeds and their soil were removed to create a trial plot for this study that connected a number of pre-existing small bare patches (Supplementary Fig. 5). A monitoring transect down the plot spanned the constructed clay pan and adjacent variably weedy pans. The EC of the site remained low (~ 1 mS/cm) during winter, but the site became rapidly saline in the spring, with abundant surficial salt encrustations including halite, Na-sulphates and Na-carbonates. The EC almost consistently exceeded the 1 mS/cm reference level and was generally ~ 10 mS/cm or even higher (Fig. 6c). The pH of the bare substrate remained broadly uniform, near 10 (Fig. 6d). The first germinating halophytes (A. buchananii; P. raroflorens) appeared, with abundant evaporative salts, by September 2021 (Supplementary Fig. 5). However, most of the site remained bare of halophytes and weeds into summer 2021. On an August 2022 visit, most of the developed site was still bare, but numerous patches of A. buchananii and some P. raroflorens had become established and were sprouting new extensions to these patches.
Cenozoic limestone
The Cenozoic limestone at Kurow (Fig. 1b, d) contains abundant shells and detrital minerals, especially quartz sand (Table 1). A large (200 m × 50 m) limestone outcrop has a series of plateaus on its upper surface, from which all other Cenozoic sediments have been stripped. Weathering of this limestone has created talus slopes of limestone debris and residual quartz sand beneath outcrops and in depressions within the outcrops (Fig. 2d; Supplementary Figs. 1, 6). Outcrop surfaces are fluted and irregular, with abundant solution cavities that are variably filled with residual quartz sand (Fig. 2b; Supplementary Figs. 1, 6).
Existing calciflora were established in these depressions and joint cracks, rooted in sandy weathering residue, as well as in talus on the margins of the outcrop (Fig. 2b; Supplementary Figs. 1, 6). However, encroachment by adventive weeds, especially the grass species Festuca rubra over the last 15 years, has reduced the micro-habitat range of the calciflora (Supplementary Fig. 6). Establishment of invasive grass has led to development of thick (up to 30 cm) organic-rich soil that fills cavities on the limestone plateau and forms a cap over the talus (Supplementary Fig. 6).
Soil removal trials to form bare ground were begun in April–May 2021 on a plateau that no longer retained any endemic plant species, although they occurred on adjacent plateaus. The encroaching soil and vegetation was peeled back to re-expose the surface depressions of 2 m × 2 m area and the depressions were partially back-filled with residual sand transported from unvegetated sandy talus deposits (Supplementary Fig. 6). This treatment was similar to that at the white clay site at Chapman Road (Supplementary Fig. 3). Additional trials were conducted for soil and weed removal around established calciflora (Supplementary Fig. 6), with excavations locally extending to 50 cm depth to ensure removal of weed roots. This latter activity was a development on earlier weeding attempts, with shallow excavation, that failed to restrict weed regrowth. The weeded areas were developed in a similar manner to the Galloway saline site (Supplementary Fig. 5).
After 6 months, the bare nature of the soil removal trial plot had been partially maintained, with some sprouting of adventive Bromus grass species, with minor F. rubra. (Fig. 2f; Supplementary Fig. 6). The EC remained low (~ 0.2 mS/cm) and pH was consistently ~ 8 (Fig. 2f; Supplementary Fig. 6). At least some of the sprouting weeds had developed from residual rooted material clinging into cracks within the limestone substrate. However, the trials with deep weeding around established calciflora were more successful as the calciflora were thriving (Supplementary Fig. 6), and some new calciflora germination was occurring. Only minor weed sprouting had occurred at these latter sites.
Discussion
Physical and chemical controls on plant colonisation
The halophytes and calciflora tolerate, or even require, on-going disturbance within dynamic soil-free environments (Fig. 2a–d; Supplementary Fig. 1). Such disturbance can prevent formation of biocrust, for example, which stabilises bare surfaces and facilitates incursion of other plant species (Mengual et al. 2014; Weber et al. 2016; Chock et al. 2019). Some anthropogenic activities, including pastoral agriculture and historical mining, create surface disturbances that can lead to localised erosion in rain events and these activities appear to enhance soil-free areas. The surficial disturbances created by soil removal in this pilot study therefore had primarily positive effects on these soil-free ecosystems. Our most successful plots, the white clay site at Chapman Road and Crawford Hills (Fig. 4a; Supplementary Fig. 5), were where most surface disturbance occurred including some physical transport of schist-derived clay around the site. Despite this extreme disturbance, halophytes rapidly became established, salinity rose and weeds were largely excluded during the monitoring period (Figs. 4a, 5a, 7a, b). Observations in this study suggest that gently sloping surfaces that channel surface water runoff down clay-rich surfaces are the most effective at redevloping high salinity and high pH after soil removal. However, we have not investigated more substantial intervention with surface and sub-surface hydrology to try to enhance supply of dissolved salts.

Summary of the principal successful soil-free sites developed in this study. a White schist-derived clay distributed over a Chapman Road site after removal of weeds and soil in winter 2020. b Close view of the Chapman substrate in winter 2022, showing extensive halophyte colonisation (light brown patches). c Crawford site during development in winter 2021. d Close view of Crawford substrate in winter 2022, with mainly bare substrate and some sprouting halophytes. e Deep weeding disturbance of limestone talus, done in winter 2021, with incipient sprouting of weeds. f Weeded limestone talus below established calciflora, in summer 2022, with new sprouting calciflora (circled) and negligible weed incursion
Relatively high Na levels in soils (e.g. exchangeable sodium percentage, ESP or sodium adsorption ratio, SAR) are considered to be a negative feature in agricultural settings, primarily because the abundant Na, typically adsorbed to clay particles, affects soil structure and commonly leads to subsoil impermeability (Sposito 2016). Our trial sites on schist-derived clay have essentially the opposite situation: it is the soil-free substrate impermeability that leads to the high Na that appears as Na-bearing evaporative mineral precipitates, commonly with formation of Na-carbonate as well as halite (Fig. 3e; cf Datta et al. 2002). Such alkaline pH may assist, along with high salinity, in maintaining chemically extreme environments that favour halophytes over adventive species (e.g. Galloway, Crawford sites; Supplementary Fig. 5). The Crawford site in particular was one of the most successful at excluding weeds from bare ground throughout the study, even though halophyte colonisation has been slower than the white clay at Chapman Road (Fig. 7a–d).
Soil-free limestone-related substrates also contain adequate nutrients for plant establishment, albeit in some different minerals from the basement (Table 1). In contrast to the schist clay substrates, however, the limestone substrates have relatively low Na and have calcite-mediated pH near 8 (Table 1; Fig. 3e). Hence, the chemical environment provided by limestone for calciflora, while moderately alkaline, is less challenging for adventive species. Establishment of calciflora on the soil-free limestone environments is at least partially controlled by their tolerance for physically dynamic environments, in contrast to their potential competitors (Fig. 2b, d; Supplementary Fig. 6). Deep weeding and soil removal around established calciflora, with exposure of rough, bare and irregularly permeable limestone debris proved to be the most successful intervention strategy in this study (Fig. 7e, f).
Rates of colonisation and weed incursion
Our trial plots have shown that significant halophyte colonisation can occur on a time scale of months, provided suitable bare substrates are provided. Colonisation was faster than anticipated from known reproduction cycles and suggests vegetative means or the existence of nearby viable seed, both of which were washed into the trial plots after rain events. While coverage of bare substrates by halophytes is seldom, if ever, complete, the bare substrates we have created constitute enhanced significant refugia for these rare endemic plants, from which they can spread to surrounding areas if bare substrates become available. The rare halophyte Lepidium kirkii did not colonise any of the saline sites we developed because the only seed sources were distant (kilometres) from our sites. This species is remarkably tolerant of surface disturbance (Supplementary Fig. 1d) and deliberate translocation to a site of soil removal may be a plausible conservation strategy. Rapid germination of calciflora seeds occurred near to parent plants on weeded substrates. However, longer-distance calciflora seed migration (tens of metres) did not occur during our study, although suitable bare substrates were maintained by our treatment processes (Fig. 2b, d, f).
Our results show that from the plant survival perspective, and associated enhanced biodiversity, soil removal and re-establishment of bare substrates is a viable land management tool where specialised plants and substrates are available. Based on our experience in this study, removal of at least 1 m of soil and underlying substrate is likely to be most effective, as this will remove most weed rootlets and seed bank and slow the re-establishment of weeds. Our hand-operated soil removal was too limited in places to complete this as effectively as necessary and several of our sites were less successful as a result, especially the eastern Chapman Road sites. Minor mechanical excavation at the Crawford site in this study was most effective at re-creating weed-free substrate, at least in the short term (Supplementary Fig. 5). If the concept tested in this study was to be scaled up to larger areas, mechanical removal of weeds and soil would be necessary and is a practical approach.
What is not yet clear is the time scale on which bare substrates can be maintained without intensive management to prevent incursion of a soil layer and/or other plants. Adventive plants and new soil development appear to be intimately linked, in that wind-blown or water-washed sediment on bare substrates can host adventive species, and the presence of those plants encourages entrapment of more sediment, leading to formation of new soil and exclusion of halophytes or calciflora. In particular, exotic salt-tolerant species such as the herb Plantago coronopus (Koyro 2006) and exotic grasses (e.g. Elymus and Puccinellia spp.) can become established in saline bare substrates almost as quickly as the endemic halophytes (1–2 years) and trap sediment to allow incursion of less salt-tolerant species. Similarly, the exotic grass Bromus sp. and Festuca rubra appear to colonise limestone pavements and associated residual deposits and exclude endemic calciflora, on a time scale of 2–10 years. Residual roots and especially seeds from such exotic grasses may remain a regeneration problem on bare limestone-related sites after soil removal and backfilling with talus material (Supplementary Fig. 6; Rydgren et al. 2016). In a scaled-up soil removal operation, mechanical removal of up to 1 m of surficial material to remove roots of weeds, and even then it may be necessary to use herbicide at times prevent re-incursion.
Limited historic (since 2000) high-resolution aerial photographic coverage suggests that on a 10–20 year time scale, there has been significant shrinkage (5–20 m scale) of areas of exposed bare substrates on clay-altered schist and on limestone outcrops. These generalised aerial observations support anecdotal observations from local farmers and researchers. However, the model example we describe from Conroys reference site in this study Supplementary Fig. 2) shows no sign of new soil development or weed infestation after 8 years. Shrinkage of areas of bare substrate in medium to long term (decades) results from natural processes, and in the pre-human environment they would have occurred at similar rates (Craw et al. 2013). In the pre-human environment, new areas of bare substrate would have developed periodically in a dynamic surficial environment (Craw et al. 2013). Colonisation of these new soil-free sites by endemic halophytes or calciflora from the existing but shrinking sites permitted the species to evolve and survive through the late Cenozoic. In the present human-dominated environment, bare substrates are relatively rare and short-lived because of a tendency to rehabilitate them with soil cover and/or irrigation. Irrigation in particular is an increasing threat to the persistence of halophyte communities outside reserves.
Soil-free rehabilitation in other settings
The endemic plants and their relationships to soil-free substrates in this study are specific to southern New Zealand and are not necessarily directly applicable elsewhere in the world. However, the approach of soil removal and formation or expansion of bare substrates as a management strategy to enhance biodiversity during rehabilitation of disturbed lands can be applied elsewhere if suitable plant–substrate relationships can be defined. Our study has shown that there are adequate nutrients for plant establishment in common rock substrates and their eroded debris (Table 1) and similar nutrient availability is indicated by spontaneous vegetation establishment of plants on soil-free historic abandoned disturbed land (Hodacova and Prach 2003; Prach and Hobbs 2008; Valente et al. 2012). Further, bare surfaces created by recreational activity had enhanced biodiversity outcomes during restoration of sand pits (Rehounkova et al. 2016). Direct planting of specialist species that are tolerant of acid, alkalinity, metals, or salts into geochemically distinctive soil-free substrates such as mine tailings or waste rock piles (Bizoux et al. 2004; Valente et al. 2012; Parraga-Aguadoet al. 2013; Xie and van Zyl 2020) is a similar strategy to soil removal as described herein as long as these plantings can be maintained without subsequent soil development and vegetation succession.
Observations of natural (spontaneous) rehabilitation processes show that endemic plants can colonise bare substrates with a wide range of compositions, especially at historic mine sites (Prach and Hobbs 2008; Valente et al. 2012; Tropek et al. 2012, 2013; Doley and Audet 2013). In most situations, the colonising vegetation facilitates development of new soil cover by intercepting fine sediments and accumulation of leaf litter (Valente et al. 2012; Tropek et al. 2012, 2013; Tarvainen and Tolvanen 2016). In addition, colonisation by pioneer species facilitates later establishment of other species (Alday et al. 2016). While these processes may be considered desirable in many situations, they lead to relative uniformity of landscape and lower biodiversity. Hence, the soil removal strategy described herein is one way to retain some specialist plants that rely on particular bare substrates in a wider landscape of established grasses and weeds, even though most of an area may undergo rehabilitation to vegetated soil.
Conclusions
Our pilot study has provided evidence that soil and weed removal to re-establish bare substrates, at least on a small scale, can enhance habitat availability for specialised ecosystems on chemically distinctive disturbed substrates, thereby increasing the spatial range of local biodiversity. In our study, we have investigated colonisation success by rare endemic salt-tolerant (halophyte) plants on clay-rich rock substrates and also rare endemic calciflora on limestone substrates. Our observations at a reference site which was excavated 8 years previously show that clay-rich impermeable substrates can attain sufficiently high salinities from surface evaporation during all seasons to limit weed incursion and permit establishment of halophytes. At that reference site, there is a well-defined increase in salinity of substrates during spring and early summer as regional evapotranspiration increases.
Soil-free clay-rich sites that were prepared for this study had ~ 0.5 m of surface removal and reworking. After this activity, none of the clay-rich sites developed uniform high salinity in the time scale of our observations (maximum 16 months), but some patches of high salinity did develop locally. Our most successful saline site (Fig. 7a, b) is located in an area that was extensively disturbed by historical gold mining and the substrate prepared for this study is especially clay-rich. This cleared site was extensively colonised by halophytes over the first year of our observations, despite the irregularly evolving salinity. However, some incursion of weeds occurred as well, in less saline portions. Other saline sites from which soil was removed had variable amounts of halophyte colonisation, weed incursion and proportions of remaining bare ground. Weed incursion is expected to decrease as a result of rising salinity although they may not be completely eliminated. Two of the saline sites became rapidly highly alkaline which further inhibited weed incursion, especially at the site with the consistently highest salinity (EC up to 10 mS/cm) and highest pH (~ 10) (Fig. 7c, d).
Full soil clearance and backfilling with talus at the limestone site was not successful in attracting calciphyte colonisation, although the bare substrates were maintained by our treatments with only minor weed sprouting. The most successful strategy on limestone was deep hand weeding of coarse talus areas around established calciflora, which permitted rapid germination of new plants (Fig. 7e, f). These observations, combined with those at the saline sites, suggest that excavation deeper than 0.5 m would have been more successful at removing traces of weeds and exposing substrate for colonisation by the specialist plants on the resulting disturbed ground.
Data availability
All data for this study are included in the paper, the Supplementary File, and in cited references.
References
Alday JG, Zaldívar P, Torroba-Balmori P, Fernández-Santos B, Martínez- Ruiz C (2016) Natural forest expansion on reclaimed coalmines in Northern Spain: the role of native shrubs as suitable microsites. Environ Sci Pollut Res 23:13606–13616
Allen RB, McIntosh PD (1997) Guidelines for conservation of salt pans in Central Otago. Science for Conservation 49, Department of Conservation, Wellington, New Zealand
Anawar HM (2015) Sustainable rehabilitation of mining waste and acid mine drainage using geochemistry, mine type, mineralogy, texture, ore extraction and climate knowledge. J Environ Manag 158:111–121
Babel M, Schreiber BC (2014) Geochemistry of evaporites and evolution of seawater. Chapter 9.17. In: Holland HD, Turekian KK (eds) Treatise on geochemistry, 2nd edn. Elsevier Science, New York, pp 483–560 (ISBN 978-0-08-098300-4)
Banning NC, Lalor BM, Grigg AH, Phillips IR, Colquhoun IJ, Jones DL, Murphy DV (2011) Rehabilitated mine-site management, soil health and climate change. Chapter 13 soil health and climate change, soil biology, vol 29. Springer, Berlin, pp 287–314. https://doi.org/10.1007/978-3-642-20256-8-13
Bizoux JP, Brevers F, Meerts P, Graitson E, Mahy G (2004) Ecology and conservation of Belgian populations of Viola calaminaria, a metallophyte with a restricted geographic distribution. Belg J Bot 137:91–104
Bolan NS, Kirkham MB, Ok YS (2017) Spoil to soil: mine site rehabilitation and revegetation. CRC Press, Boca Raton
Bradshaw A (2000) The use of natural processes in reclamation—advantages and difficulties. Landsc Urban Plan 51:89–100
Chamberlain CP, Poage MA, Craw D, Reynolds RC (1999) Topographic development of the Southern Alps recorded by the isotopic composition of authigenic clay minerals, South Island, New Zealand. Chem Geol 155:279–294
Cheeseman JM (2015) The evolution of halophytes, glycophytes and crops, and its implications for food security under saline conditions. New Phytol 206:557–570
Chock T, Antoninka AJ, Faist AM, Bowker MA, Belnap J, Barger NN (2019) Responses of biological soil crusts to rehabilitation strategies. Jarid Environ 163:77–85
Clewell AF, Aronson J (2013) Ecological restoration: principles, values, and structure of an emerging profession, 2nd edn. Island Press, Washington, DC (ISSN 2945-5391)
Craw D, Rufaut C (2017) Geochemical and mineralogical controls on mine tailings rehabilitation and vegetation, Otago Schist, New Zealand. NZ J Geol Geophys 60:176–187
Craw D, Druzbicka J, Rufaut C, Waters J (2013) Geological controls on paleo-environmental change in a tectonic rain shadow, southern New Zealand. Palaeogeogr Palaeoclimatol Palaeoecol 370:103–116
Craw D, Rufaut C, Pillai D (2022) Geological controls on evolution of evaporative precipitates on soil-free substrates and ecosystems, southern New Zealand. Sci Total Environ 849:157792
Craw D, Rufaut C, Pillai D (2023) Evolution of alkaline mine drainage and unique biodiversity on soil-free mine substrates, southern New Zealand. Mine Water Environ 42:3–23. https://doi.org/10.1007/s10230-023-00913-x
Datta S, Thibault Y, Fyfe WS, Powell MA, Hart BR, Martin RR, Triphy S (2002) Occurrence of trona in alkaline soils of the Indo-Gangetic Plains of Uttar Pradesh (U.P.), India. Episodes 25:236–239
Doley D, Audet P (2013) Adopting novel ecosystems as suitable rehabilitation alternatives for former mine sites. Ecol Processes 2:22
Druzbicka J, Rufaut C, Craw D (2015) Evaporative mine water controls on natural revegetation of placer gold mines, southern New Zealand. Mine Water Environ 34:375–387
Forsyth PJ (2001) Geology of the Waitaki area. Institute of Geological and Nuclear Sciences 1:250,000 Geological Map 19. Institute of Geological & Nuclear Sciences Ltd, Lower Hutt
Haigh M (2018) Building a cradle for nature: a paradigm for environmental reconstruction. In: Prasad MNV, Favas P, Maiti SK (eds) Bio-geotechnologies for mine site rehabilitation. Elsevier, Amsterdam, pp 593–615 (ISBN 978-0-12-812986-9)
Hancock GR, Martin Duque JF, Willgoose GR (2020) Mining rehabilitation—using geomorphology to engineer ecologically sustainable landscapes for highly disturbed lands. Ecol Eng 155:105836
Heenan PB, McGlone MS (2019) Cenozoic formation and colonisation history of the New Zealand vascular flora based on molecular clock dating of the plastid rbcL gene. NZ J Bot 57:204–226
Hobbs RJ, Higgs E, Harris JA (2009) Novel ecosystems: implications for conservation and restoration. Trends Ecol Evol 24:599–605
Hodacova D, Prach K (2003) Spoil heaps from brown coal mining: technical reclamation versus spontaneous revegetation. Restor Ecol 11:385–391
Holliday R, Johnson M (1979) The contribution of derelict mineral and industrial sites to the conservation of rare plants in the United Kingdom. Environ Geochem Health 1:1–7
Hutton CO, Seelye FT (1942) Francolite, a carbonate-apatite from Milburn, Otago. Trans R Soc NZ 72:191–198
Jackson J, Norris RJ, Youngson JH (1996) The structural evolution of active fault and fold systems in Central Otago, New Zealand: evidence revealed by drainage patterns. J Struct Geol 18:217–234
Koyro H-W (2006) Effect of salinity on growth, photosynthesis, water relations and solute composition of the potential cash crop halophyte Plantago coronopus (L.). Environ Exp Bot 56:136–146
Landis CA, Campbell HJ, Begg JG, Mildenhall DC, Paterson AM, Trewick SA (2008) The Waipounamu erosion surface: Questioning the antiquity of the New Zealand land surface and terrestrial fauna and flora. Geol Mag 145:173–197
Law S, Rufaut C, Lilly K, Craw D (2016) Geology, evaporative salt accumulation and geoecology at Springvale historic gold mine, Central Otago, New Zealand. N Z J Geol Geophys 59:382–395
Mengual C, Schoebitz M, Azcon R, Roldan A (2014) Microbial inoculants and organic amendment improves plant establishment and soil rehabilitation under semiarid conditions. J Environ Manag 134:1–7
Meuser H (2012) Soil remediation and rehabilitation: treatment of contaminated and disturbed land. Springer Science and Business Media, Berlin (E-ISSN 2215-1702)
Miller AL, Duncan RP (2003) Extrinsic and intrinsic controls on the distribution of the critically endangered cress, Ischnocarpus exilis (Brassicaceae). Biol Conserv 110:153–160
Munshower FF (2018) Practical handbook of disturbed land revegetation. CRC Press, Boca Raton
NIWA (2021) National Institute and Water and atmospheric research websites. https://www.niwa.co.nz/climate; https://www.cliflo.niwa.co.nz. Accessed Feb 2022
ORC (2021) Otago Regional Council, monthly evapotranspiration, lauder site. www.orc.govt.nz/managing-our-environment/land/land-monitoring. Accessed Feb 2022
Parraga-Aguado I, Gonzalez-Alcaraz MN, Alvarez-Rogel J, Jimenez-Carceles FJ, Conesa HM (2013) The importance of edaphic niches and pioneer plant species succession for the phytomanagement of mine tailings. Environ Pollut 176:134–143
Prach K, Hobbs RJ (2008) Spontaneous succession versus technical reclamation in the restoration of disturbed sites. Restor Ecol 16:363–366
Prach K, Tolvanen A (2016) How can we restore biodiversity and ecosystem services in mining and industrial sites? Environ Sci Pollut Res 23:13587–13590
Rehounkova K, Cizek L, Rehounek J, Sebelíková L, Tropek L, Lencova K, Bogusch P, Marhoul P, Maca J (2016) Additional disturbances as a beneficial tool for restoration of post-mining sites: a multi-taxa approach. Environ Sci Pollut Res 23:13745–13753
Rogers G, Hewitt A, Wilson JB (2000) Ecosystem-based conservation strategy for Central Otago’s saline patches. Science for Conservation 166, Department of Conservation, Wellington
Rogers GM, Courtney SP, Heenan PB (2018) The calcicolous vascular flora of New Zealand: life forms, taxonomy, biogeography and conservation status. Science for Conservation 331, Department of Conservation, Wellington, New Zealand
Rufaut CG, Craw D, Law S, Druzbicka J (2018) Conservation of saline patches in Central Otago needs better recognition of physical processes to secure future habitats. N Z J Bot 56:115–126
Rydgren K, Auestad I, HamreL N, Hagen D, Rosef L, Skjerdal G (2016) Long-term persistence of seeded grass species: an unwanted side-effect of ecological restoration. Environ Sci Pollut Res 23:13591–13597
Sebelikova L, Rehounkova K, Prach K (2016) Spontaneous revegetation vs. forestry reclamation in post-mining sand pits. Environ Sci Pollut Res 23:13598–13605
Sere G, Schwartz C, Ouvrard S, Sauvage C, Renat J-C, Morel JL (2008) Soil construction: a step for ecological reclamation of derelict lands. J Soils Sed 8:130–136
Sposito G (2016) The chemistry of soils, 3rd edn. Oxford University Press, Oxford (ISBN:9780190630881)
Tarvainen O, Tolvanen A (2016) Healing the wounds in the landscape: reclaiming gravel roads in conservation areas. Environ Sci Pollut Res 23:13732–13744
Tropek R, Kadlec T, Hejda M, Kocarek P, Skuhrovec J, Malenovsky I, Vodka S, Spitzer L, Banar P, Konvicka M (2012) Technical reclamations are wasting the conservation potential of post-mining sites. a case study of black coal spoil dumps. Ecol Eng 43:13–18
Tropek R, Hejda M, Kadlec T, Spitzer L (2013) Local and landscape factors affecting communities of plants and diurnal Lepidoptera in black coal spoil heaps: Implications for restoration management. Ecol Eng 57:252–260
Turnbull IM (2000) Geology of the Wakatipu area. Institute of Geological & Nuclear Sciences 1:250,000 geological map 18. Institute of Geological & Nuclear Sciences Ltd, Lower Hutt
Valente T, Gomes P, Pamplona J, Luisa de la Torre M (2012) Natural stabilization of mine waste dumps: evolution of the vegetation cover in distinctive geochemical and mineralogical environments. J Geochem Expl 123:152–161
Weber B, Budel B, Belnap J (2016) biological soil crusts: an organizing principle in drylands. Springer International Publishing, AG, Cham (ISBN 978-3-319-30214-0)
Xie L, van Zyl D (2020) Distinguishing reclamation, revegetation and phytoremediation, and the importance of geochemical processes in the reclamation of sulfidic mine tailings: a review. Chemosphere 252:126446
Acknowledgements
We thank Euan Hind of Otago Regional Council for his help. Central Otago District Council staff provided helpful logistical support. Discussions with Rob and Kate Wardle, Clement Lagrue, Ellery Mayhence and Sasha Roselli on site management were useful for planning purposes. We gratefully acknowledge land access and enthusiastic support provided by Roger and Wendy Browne, Anna Robinson, Nick Loughnan and Faith Gray. Volunteer assistance from members of the Haehaeata Natural Heritage Trust helped to get this project started.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions This research was funded by Otago Regional Council and University of Otago.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception, design, site selection and data gathering. CR and DP developed the ecological components of the study and DC developed the geochemical and geological components. DP collected regular monitoring data. DC produced the first draft with direct input from CR. Subsequent drafts were compiled by DC and revised in accordance with comments by CR and DP. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rufaut, C., Pillai, D. & Craw, D. Enhancement of alkaline saline soil-free bare substrates and specialist ecosystems, southern New Zealand. Environ Earth Sci 82, 440 (2023). https://doi.org/10.1007/s12665-023-11145-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12665-023-11145-1