
Abstract
Somatostatin and its receptors are expressed in the spinal cord, but the functional roles of the peptide remain unknown. In this study, we examined the colokinetic effect of somatostatin in the spinal defecation center in anesthetized rats. Intrathecal application of somatostatin into the lumbo-sacral cord caused propulsive contractions of the colorectum. However, somatostatin administered intravenously or intrathecally to the thoracic cord failed to enhance colorectal motility. Transection of the thoracic cord had no significant impact on the colokinetic action of somatostatin. The enhancement of colorectal motility by intrathecal administration of somatostatin was abolished by severing the pelvic nerves. Our results demonstrate that somatostatin acting on the spinal defecation center causes propulsive motility of the colorectum in rats. Considering that somatostatin is involved in nociceptive signal transmission in the spinal cord, our results provide a rational explanation for the concurrent appearance of chronic abdominal pain and colonic motility disorders in IBS patients.
Similar content being viewed by others
Introduction
Somatostatin, known as somatotrophin-releasing inhibitory factor (SRIF), was originally discovered as a hypothalamic neuroendocrine hormone that potently inhibits the secretion of growth hormone from the anterior pituitary gland [1]. Somatostatin is also produced throughout the central nervous system and in peripheral organs such as the pancreas and gastrointestinal tracts [2, 3]. Somatostatin binds to five subtypes (sst1-5) of receptors belonging to the family of G protein-coupled receptors [3]. Activation of somatostatin receptors mainly leads to inhibition of intracellular adenylate cyclase via Gi [2, 4, 5].
Somatostatin and some of its receptors are also expressed in the spinal cord [4, 6]. Endogenous somatostatin of the spinal cord is derived from various neurons including primary afferent neurons, spinal interneurons and descending neurons from some regions of the brain [7–10]. Abundant somatostatin-immunoreactive nerve terminals project to the spinal dorsal horn, where somatostatin is involved in inhibitory regulation of nociceptive signal transmission [6, 9, 11, 12]. Spinal somatostatin-immunoreactive nerve terminals as well as neurons immunoreactive to its receptors have also been detected in the intermediolateral cell column [6, 9, 13]. Since the intermediolateral cell column consists of a mass of preganglionic neuronal cells of the autonomic nerves, spinal somatostatin would be involved in regulation of the autonomic nervous system at the spinal level.
In our previous study, we demonstrated that intrathecal administration of ghrelin, a peptide acting on the growth hormone secretagogue receptor 1a (GHSR1a), at the L6–S1 spinal level causes strong propulsive contractions of the rat colorectum [14]. The effect of ghrelin on colorectal motility is brought about by activation of GHSR1a-expressing neurons of the intermediolateral cell column at the lumbo-sacral spinal cord, where the spinal defecation center is located [15, 16]. Recently, we have tried to reveal the characteristics of ghrelin-sensitive neurons of the lumbo-sacral defecation center by comparing them with the characteristics of NPY-containing hypothalamic ghrelin-sensitive neurons [17]. Although our results demonstrated that the characteristics of ghrelin-sensitive neurons are different from those of hypothalamic neurons, the precise characteristics of ghrelin-sensitive neurons in the spinal defecation center remain to be clarified.
In the hypothalamus, somatostatin exerts an effect opposite to that of ghrelin and reduces growth hormone release [1, 18]. In addition, ghrelin-sensitive neurons are directly inhibited by somatostatin in rat arcuate neurons in vitro [19]. Thus, we hypothesized that somatostatin may compete with the colokinetic effect of ghrelin in the spinal defecation center. To test the hypothesis, we investigated whether intrathecally administered somatostatin inhibited colorectal motility. Contrary to our expectation, somatostatin intrathecally administered into the L6–S1 region of the spinal cord enhanced colorectal motility. Thus, in the present study, we investigated the mechanisms of the effect of somatostatin on colorectal motility.
Materials and methods
Animals
Male Sprague-Dawley rats (Japan SLC, Inc., Shizuoka, Japan) were used. The rats were maintained in plastic cages at 22 °C with a 12:12 h light:dark cycle (light on 07:00–19:00 h), and they were supplied with both laboratory chow (MF, Oriental Yeast Co., Ltd., Tokyo, Japan) and water ad libitum prior to experiments. The experimental procedures were performed according to the guidelines for the care and use of laboratory animals approved by the Animal Care and Use Committee of Gifu University (Permission Nos. 14100, 15094). The total number of animals used in this study was 45.
Recording of colorectal motility
The procedures for recording colorectal motility were based on those described previously [14, 20]. Rats (8–10 weeks old) were sedated with ketamine hydrochloride (50 mg/kg, intramuscular injection) and anesthetized with alpha-chloralose (60 mg/kg, into the tail vein). The femoral artery was cannulated, and anesthesia was maintained by intra-arterial infusion of alpha-chloralose (10–20 mg/kg/h) combined with ketamine hydrochloride (3–5 mg/kg/h) in 0.9% saline. The arterial cannula was connected to a pressure transducer for monitoring arterial blood pressure. Body temperature was maintained at 36–37 °C by a heating blanket (Homeothermic Blanket System, Harvard Apparatus, Holliston, MA, USA) throughout experiments. At the completion of the experiments, rats were immediately killed by intraperitoneal injection of a lethal dose of sodium pentobarbitone (100 mg/kg) while they were still under anesthesia.
The colorectum of each anesthetized rat was cannulated and tied around both in the region of the distal colon behind the bladder and at the anus, and then the body wall was closed around the oral cannula. The oral cannula was connected to a Mariotte bottle filled with warm saline kept at 37 °C, and the aboral cannula was connected to a pressure transducer and a fluid outlet through a one-way valve for measurements of intraluminal pressure of the colorectum and expelled fluid volume, respectively. The basal level of intraluminal pressure was maintained at 4–6 mmHg by adjusting the heights of the Mariotte bottle and outlet tube. Expelled fluid from the aboral cannula was collected in a cylinder positioned beneath the fluid outlet and measured with a force transducer.
When the spinal cord was transected at T4 during the recording session, the region of the T3 and T4 vertebrae was exposed by laminectomy, and the spinal cord was transected with micro scissors [21]. In some series of experiments, the parasympathetic pelvic nerves or sympathetic nerves (colonic and hypogastric nerves) were bilaterally cut before the colorectal cannulation [22, 23]. After the surgical operation for recording colorectal motility, the rats were kept for about 1 h to allow the basal colorectal motility and blood pressure to stabilize.
Administration of somatostatin
For intrathecal application of somatostatin to the thoracic or lumbo-sacral spinal cord, a 30-gauge needle connected to a polyethylene tube was inserted between the T3 and T4 or L1 and L2 vertebrae from the dorsal surface. The L1–L2 vertebrae correspond to spinal cord level L6–S1 in the rat. The cannula was secured in place with instant adhesive (Aron Alpha Extra; Toagosei, Co., Ltd., Tokyo, Japan) to create a tight seal at the point of cannulation. There was no cerebrospinal fluid leak. For intravenous injection of somatostatin, the femoral vein was cannulated.
Reagents
The following compounds were used: alpha-chloralose (Nacalai Tesque, Inc., Kyoto, Japan), ketamine hydrochloride (Daiichi Sankyo Co., Ltd., Tokyo, Japan) and somatostatin (Peptide Institute, Osaka, Japan). Somatostatin was dissolved in distilled water and stored in a freezer. Alpha-chloralose was solubilized with 10% 2-hydroxypropyl-beta-cyclodextrin (Wako Pure Chemical Industries, Ltd., Osaka, Japan) and then made up with 0.9% saline for infusion.
Statistics
Data are expressed as mean ± SD. We checked the normality of variance of our data by using the chi-square test for goodness of fit and checked the homogeneity of variance of our data by using the F test. According to the distribution of variance, either the paired or unpaired, two-tailed Student’s t test or Wilcoxon signed rank sum test was used for statistical analysis between two groups. Multiple comparisons were performed by the Steel-Dwass test (Figs. 3, 5). Multiple comparisons between related groups were performed by two-way ANOVA followed by Scheffé’s F test (Fig. 2b, c). P values <0.05 were considered to be statistically significant. Somatostatin-induced responses were quantified using data obtained for an initial period of 20 min after the beginning of their appearance. When responses did not appear, data calculation was started 5 min after drug administration. The data shown in Fig. 2d were analyzed for each 10-min period after somatostatin injection until 60 min to demonstrate time-dependent change in the frequency of contractions. For counting the number of contractions, we used the LabChart software (ADInstruments, NSW, Australia). All pressure increases of more than 2 mmHg above baseline were included. Changes of blood pressure were calculated as the rate of change corresponding to the mean arterial blood pressure in a 10-min period prior to somatostatin injection.
Results
Effects of intrathecal injection of somatostatin on colorectal motility
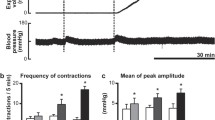
Spontaneous brief and small rises in intraluminal pressure without accompanying fluid output from the anal cannula were recorded in a resting state before intrathecal application. When 0.9% saline was injected into the lumbo-sacral spinal cord as a control, there was no significant difference in either the expelled volume or frequency of contractions before and after saline injection (n = 5, Figs. 1, 2b, c). Subsequent injection of 0.6 nmol somatostatin transiently enhanced colorectal motility and significantly increased the expelled fluid volume compared with their respective vehicle controls (n = 5, Fig. 2a, b). Frequency of colorectal contractions was unchanged after the injection (n = 5, Fig. 2c). A higher dose of somatostatin (6 nmol) caused strong contractions of the colorectum (Figs. 1, 2a). Increases in intraluminal pressure of more than 2 mmHg above baseline, which were accompanied by expulsion of fluid from the aboral cannula, were continuously observed for nearly 1 h (54.6 ± 30.2 min; n = 5). The expelled fluid volume and frequency of colorectal contractions were significantly increased compared with their respective vehicle controls (n = 5, Fig. 2b, c). Enhancement of the frequency was evident until 30 min after intrathecal administration of 6 nmol somatostatin (Fig. 2d). Blood pressure was transiently increased by intrathecal injection of somatostatin at the L6–S1 level (n = 5, Figs. 1a, 3e). In two of the five rats, a slight decrease in blood pressure was observed prior to the increase (Fig. 1a).

Responses of the colorectum to intrathecal somatostatin applied to the lumbo-sacral spinal cord. Representative recording traces of intraluminal pressure change (upper), expelled liquid volume (middle) and blood pressure (lower) before and after intrathecal application of somatostatin (6 nmol) are shown. Vehicle denotes time of 0.9% saline (10 µL) injection as a control

Quantitative analysis of the effect of somatostatin applied to the lumbo-sacral spinal cord. a Representative recording traces of intraluminal pressure change before and after intrathecal administration of somatostatin at a dose of 0.6 (upper) or 6 nmol (lower). Bar graphs summarize b expelled liquid volume (ml/20 min) and c frequency of contractions (contractions/20 min) before saline injection, after saline injection and after somatostatin (0.6 or 6 nmol) injection. Each value represents the mean ± SD (n = 5). *Significantly different between two groups (P < 0.05). d Time-dependent changes in frequency of contractions before and after injection of 6 nmol somatostatin (n = 5) are shown

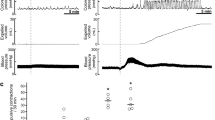
Effect of thoracic intrathecal or intravenous administration of somatostatin on colorectal motility. Representative recording traces of intraluminal pressure change (upper) and blood pressure (lower) before and after a thoracic intrathecal and b intravenous application of somatostatin (6 nmol) are shown. In both administration routes, somatostatin failed to enhance colorectal motility. Bar graphs summarize c expelled liquid volume (ml/20 min) and d frequency of contractions (contractions/20 min) after saline injection and after somatostatin (6 nmol) injection. Each value represents the mean ± SD (n = 5). Blood pressure was greatly increased by somatostatin injected into the thoracic spinal cord but not by somatostatin injected into the vein. e Quantitative data of increase of blood pressure caused by somatostatin (6 nmol) injection into the lumbo-sacral spinal cord, thoracic spinal cord and vein (n = 5). *Significantly different between two groups (P < 0.05)
Effects of thoracic or intravenous administration of somatostatin on colorectal motility
To verify the action site of intrathecally injected somatostatin, the peptide was administered into the thoracic spinal cord. Intrathecal administration of somatostatin at the T3–4 level failed to enhance colorectal motility (Fig. 3a, c, d). In addition, no changes in colorectal motility were observed when somatostatin was administered intravenously (Fig. 3b–d). Thoracic injection, but not intravenous injection, of somatostatin greatly increased blood pressure (Fig. 3a, b, e). An initial drop of blood pressure was also observed in all of the rats injected intravenously and in three of the five rats injected into the thoracic cord.
Effects of T4 spinal transection on the somatostatin-induced enhancement of colorectal motility
To examine whether activity of the supraspinal regions is essential for the action of somatostatin, the thoracic spinal cord was transected at the T4 level. T4 transection did not affect spontaneous colorectal contractions (Fig. 4b). One hour after transection of the thoracic spinal cord, 6 nmol of somatostatin was injected into the spinal cord at the L6–S1 level. As shown in Fig. 4a, somatostatin caused marked propulsive motility of the colorectum, the parameters (expelled volume and frequency of contractions) of which were comparable to those observed in rats without transection of the thoracic spinal cord (Fig. 4c). In rats in which the thoracic spinal cord was transected at the T4 level, blood pressure did not increase after administration of somatostatin into the L6–S1 spinal cord (data not shown).

Colokinetic effect of somatostatin after surgical transection of the thoracic spinal cord at T4. a Representative recording traces of intraluminal pressure change (upper) and expelled liquid volume (lower) before and after injection of somatostatin (6 nmol) under the condition of neural disconnection between the brain and spinal defecation center by transection of the thoracic spinal cord at T4 level are shown. b Bar graphs summarize expelled liquid volume (ml/20 min, left panel) and frequency of contractions (contractions/20 min, right panel) before and after T4 transection. Somatostatin caused propulsive contractions of the colorectum after T4 transection. c Bar graphs summarize expelled liquid volume (ml/20 min, left panel) and frequency of contractions (contractions/20 min, right panel) in control rats and T4-transected rats. Each value represents the mean ± SD (n = 5). *Significantly different between two groups (P < 0.05)
Effects of surgical nerve severing on the somatostatin-induced enhancement of colorectal motility
To identify the neural pathway that mediates the action of somatostatin, effects of nerve transection were examined. Bilateral transection of the lumbar colonic and hypogastric nerves did not affect the somatostatin-induced enhancement of colorectal motility (Fig. 5a, c). On the other hand, when the pelvic nerves were bilaterally transected, intrathecal administration of somatostatin failed to increase the frequency of contractions, amplitude of colorectal pressure changes or expelled volume (Fig. 5b, c).

Colokinetic effect of somatostatin after surgical nerve severing. Representative recording traces of intraluminal pressure change (upper) and expelled liquid volume (lower) in response to intrathecal injection of somatostatin (6 nmol) after bilaterally severing a the lumbar colonic and hypogastric nerves or b the pelvic nerves are shown. Somatostatin-induced enhancement of colorectal motility was observed after severing sympathetic nerves but not after cutting the pelvic nerves. c Bar graphs summarize the expelled liquid volume (ml/20 min, left panel) and frequency of contractions (contractions/20 min, right panel) in control rats, in rats with sympathetic nerves cut (SN cutting) and in rats with pelvic nerves cut (PN cutting). Each value represents the mean ± SD (n = 5). *Significantly different between two groups (P < 0.05)
Discussion
The present study was undertaken to examine the possible inhibitory effect of somatostatin on colorectal motility in the lumbo-sacral spinal defecation center. Contrary to our expectation, intrathecal application of somatostatin to the L6–S1 region of the spinal cord enhanced colorectal motility. The major findings regarding the mechanisms of action are (1) intravenous and/or intrathoracic administration of somatostatin had no effect on colorectal motility, (2) somatostatin enhanced colorectal motility without neural interaction with the supraspinal region and (3) somatostatin-induced enhancement of colorectal motility was abolished after cutting the pelvic nerves but not after severing sympathetic nerves. These findings indicate that somatostatin enhances colorectal motility because of activation of parasympathetic preganglionic neurons of the pelvic nerves in the spinal defecation center.
Since the intrathecally injected somatostatin would be delivered to the defecation center in the brain stem through cerebrospinal fluid circulation, neurons located in supraspinal regions could be the action site of somatostatin. In addition, receptors expressed in the enteric nervous system of the gastrointestinal tract would be responsible for the action of somatostatin. In the present work, however, somatostatin had no effect when the peptide was applied to the thoracic cord, closer to the brain stem than the L6–S1 level. Also, intravenously applied somatostatin, which can directly act on the enteric nervous system, had no effect on colorectal motility. Thus, it is unlikely that stimulation of colorectal motility by intrathecal administration of somatostatin at the L6–S1 level is due to its diffusion to sites remote from the lumbo-sacral defecation center, such as the supraspinal defecation center, thoracic spinal cord and periphery.
The lumbo-sacral defecation center is controlled by descending facilitatory and inhibitory pathways from supraspinal regions [24]. Since somatostatin is known to be a neurotransmitter of primary afferent fibers in the spinal cord [7], it is possible that the neurons activated in response to intrathecal application of somatostatin belong to ascending pathways from the spinal cord to the brain. If this is the case, transmission of signals to the supraspinal defecation center would be essential to exert the colokinetic action. However, intrathecally injected somatostatin at the L6-S1 level caused propulsive motility of the colorectum even after transection of the thoracic spinal cord at the T4 level (Fig. 4a). Accordingly, intrathecally administered somatostatin can directly activate outflow pathways at the spinal defecation center without interaction with the brain.
Somatostatin receptors have been classified into five subtypes (sst1-5) that belong to the family of G protein-coupled receptors [3]. Stimulation of somatostatin receptors mainly results via Gi in suppression of adenylate cyclase [2, 4, 5]. In agreement with this, electrophysiologic studies have demonstrated that somatostatin suppresses neuronal activity in the locus coeruleus, periaqueductal gray and hypothalamic arcuate nucleus [19, 25, 26]. Accordingly, it is natural to assume that somatostatin suppresses neuronal activity also in the spinal cord, and therefore the peptide would directly suppress the activity of sympathetic preganglionic neurons. Suppression of sympathetic nerve activity can lead to enhancement of colorectal motility by removing tonic inhibitory influence [23, 27]. However, this seems unlikely since surgical cutting of the sympathetic lumbar colonic and hypogastric nerves failed to block the effect of somatostatin (Fig. 5a). Alternatively, we found that severing the parasympathetic pelvic nerve prevents the colokinetic effect of somatostatin (Fig. 5b). Taken together, the results suggest that somatostatin activates sacral parasympathetic preganglionic neurons indirectly via inhibitory interneurons. In support of this, the sacral intermediolateral cell column, which consists of a mass of preganglionic neuronal cells of the parasympathetic nerves, was shown to be densely innervated by somatostatin-immunoreactive terminals in rats [6, 9, 13].
In our previous study, we demonstrated that intrathecally injected noradrenaline or dopamine caused propulsive contractions of the colorectum via activation of the lumbo-sacral spinal defecation center [28, 29]. In the spinal cord, noradrenaline and dopamine derived from the supraspinal regions modulate noxious signal transmission, which is called descending pain inhibition [30, 31]. Based on these facts, we hypothesized that descending pain inhibitory pathways control not only pain but also the defecation reflex [32, 33]. Similar to these monoamine transmitters, some of the endogenous somatostatin in the spinal cord is derived from the supraspinal regions, including the raphe pallidus, raphe obscurus, A5 and the hypothalamus [9, 10]. Considering that A5 is the main origin of descending pain inhibitory pathways, it is possible that somatostatin and noradrenaline act as co-transmitters in the spinal defecation center.
Remarkably, blood pressure significantly increased when somatostatin was administered in the thoracic or lumbo-sacral spinal cord (Fig. 3e). Since intravenous administration of somatostatin did not increase blood pressure, somatostatin would primarily act in the central nervous system. It has been demonstrated that intracerebroventricular administration of somatostatin increases mean arterial blood pressure by increasing the plasma concentration of vasopressin in rats [34, 35]. It is therefore possible that somatostatin administered into the spinal cord promotes vasopressin release directly (i.e., diffusion to vasopressin neurons) or indirectly (i.e., activation of neurons that make synaptic contact with the vasopressin neurons). In agreement with this idea, somatostatin administered into the L6–S1 spinal cord failed to increase blood pressure after transection of the thoracic spinal cord. It has also been reported that somatostatin would evoke hypotension if the peptide acts on bulbospinal neurons in the rostral ventrolateral medulla by reducing sympathetic vasomotor tone [36]. This mechanism might be related to the initial drop of blood pressure observed in some rats in the present study. Further study is needed to clarify the precise mechanism underlying spinal somatostatin-induced increase in blood pressure.
Recently, interest has been shown in the relationship between pain and bowel dysfunction. Irritable bowel syndrome (IBS) is a functional disorder of the gastrointestinal tract characterized by abdominal pain or discomfort in the absence of a structural basis for pathogenesis [37]. The symptoms regarding noxious sensation usually appear in combination with disturbed bowel habits [38, 39]. However, the etiologic relationship among these symptoms remains unclear. In IBS patients, it has been reported that the visceral sensory threshold to noxious stimuli such as colorectal distention decreases, leading to visceral hypersensitivity [40, 41]. Somatostatin is known as a neurotransmitter associated with inhibition of noxious signaling in the spinal cord [42–44]. Actually, somatostatin hyperpolarized dorsal horn neurons and caused reduction or abolition of spontaneous firing in a rat spinal cord slice preparation [42]. Furthermore, intrathecal administration of somatostatin exerted antinociceptive effects in vivo in rats [43] and humans [44]. In this study, we showed that spinal somatostatin might be involved in the regulation of colorectal motility. Considering these findings, diminution or dysfunction of the spinal somatostatin system is a possible pathologic mechanism connecting the abdominal pain and disturbed bowel habits in IBS patients. Indeed, somatostatin-immunoreactive cells of the rat lumbar dorsal root ganglia markedly increased after surgical removal of the adrenal glands, suggesting that adrenal steroid hormones reduce the synthesis of somatostatin in dorsal root ganglion cells [45]. Since stress activates the HPA axis and stimulates release of adrenal steroid hormones, it is possible that stress-induced release of steroid hormones suppresses somatostatin synthesis and disrupts the inhibition of noxious signaling in the spinal dorsal horn, leading to visceral hypersensitivity. Therefore, stress-induced disorder of somatostatin function in the spinal cord could cause both abdominal pain and disorder of colorectal motility in IBS patients.
In summary, we have shown that somatostatin acting on the lumbo-sacral defecation center causes propulsive motility of the colorectum in rats. Considering that somatostatin is a neurotransmitter associated with noxious signaling in the spinal cord, our results provide a rationale for the concurrent appearance of chronic abdominal pain and colonic motility disorder in IBS patients. Our results also provide a new insight into the underlying mechanisms of central defecation control and suggest a novel etiology of IBS.
References
Brazeau P, Vale W, Burgus R, Ling N, Butcher M, Rivier J, Guillemin R (1973) Hypothalamic polypeptide that inhibits the secretion of immunoreactive pituitary growth hormone. Science 179:77–79
Patel YC (1999) Somatostatin and its receptor family. Front Neuroendocrinol 20:157–198
Olias G, Viollet C, Kusserow H, Epelbaum J, Meyerhof W (2004) Regulation and function of somatostatin receptors. J Neurochem 89:1057–1091
Møller LN, Stidsen CE, Hartmann B, Holst JJ (2003) Somatostatin receptors. Biochim Biophys Acta 1616:1–84
Yang SK, Chen C (2007) Involvement of somatostatin receptor subtypes in membrane ion channel modification by somatostatin in pituitary somatotropes. Clin Exp Pharmacol Physiol 34:1221–1227
Schulz S, Pauli SU, Schulz S, Händel M, Dietzmann K, Firsching R, Höllt V (2000) Immunohistochemical determination of five somatostatin receptors in meningioma reveals frequent overexpression of somatostatin receptor subtype sst2A. Clin Cancer Res 6:1865–1874
Hökfelt T, Elde R, Johansson O, Luft R, Nilsson G, Arimura A (1976) Immunohistochemical evidence for separate populations of somatostatin-containing and substance P-containing primary afferent neurons in the rat. Neuroscience 1:131–136
Gutierrez-Mecinas M, Furuta T, Watanabe M, Todd AJ (2016) A quantitative study of neurochemically defined excitatory interneuron populations in laminae I-III of the mouse spinal cord. Mol Pain. doi:10.1177/1744806916629065
Krisch B (1981) Somatostatin-immunoreactive fiber projections into the brain stem and the spinal cord of the rat. Cell Tissue Res 217:531–552
Strack AM, Sawyer WB, Platt KB, Loewy AD (1989) CNS cell groups regulating the sympathetic outflow to adrenal gland as revealed by transneuronal cell body labeling with pseudorabies virus. Brain Res 491:274–296
Todd AJ (2010) Neuronal circuitry for pain processing in the dorsal horn. Nat Rev Neurosci 11:823–836
Braz J, Solorzano C, Wang X, Basbaum AI (2014) Transmitting pain and itch messages: a contemporary view of the spinal cord circuits that generate gate control. Neuron 82:522–536
Schrøder HD (1984) Somatostatin in the caudal spinal cord: an immunohistochemical study of the spinal centers involved in the innervation of pelvic organs. J Comp Neurol 223:400–414
Shimizu Y, Chang EC, Shafton AD, Ferens DM, Sanger GJ, Witherington J, Furness JB (2006) Evidence that stimulation of ghrelin receptors in the spinal cord initiates propulsive activity in the colon of the rat. J Physiol 576:329–338
Hirayama H, Shiina T, Shima T, Kuramoto H, Takewaki T, Furness JB, Shimizu Y (2010) Contrasting effects of ghrelin and des-acyl ghrelin on the lumbo-sacral defecation center and regulation of colorectal motility in rats. Neurogastroenterol Motil 22:1124–1131
Ferens DM, Yin L, Bron R, Hunne B, Ohashi-Doi K, Kitchener PD, Sanger GJ, Witherington J, Shimizu Y, Furness JB (2010) Functional and in situ hybridization evidence that preganglionic sympathetic vasoconstrictor neurons express ghrelin receptors. Neuroscience 166:671–679
Naitou K, Shiina T, Sugita R, Nakamori H, Shimizu Y (2015) Characterization of ghrelin-sensitive neurons in the lumbosacral defecation center in rats. Neurogastroenterol Motil 27:147–155
Kojima M, Hosoda H, Date Y, Nakazato M, Matsuo H, Kangawa K (1992) Ghrelin is a growth-hormone-releasing acylated peptide from stomach. Nature 402:656–660
Mori K, Kim J, Sasaki K (2010) Electrophysiological effect of ghrelin and somatostatin on rat hypothalamic arcuate neurons in vitro. Peptides 31:1139–1145
Bogeski G, Shafton AD, Kitchener PD, Ferens DM, Furness JB (2005) A quantitative approach to recording peristaltic activity from segments of rat small intestine in vivo. Neurogastroenterol Motil 17:262–272
Shima T, Shiina T, Naitou K, Nakamori H, Shimizu Y (2014) Functional roles of capsaicin-sensitive intrinsic neural circuit in the regulation of esophageal peristalsis in rats: in vivo studies using a novel method. Am J Physiol Gastrointest Liver Physiol 306:G811–818
Naitou K, Mamerto TP, Pustovit RV, Callaghan B, Rivera LR, Chan AJ, Ringuet MT, Pietra C, Furness JB (2015) Site and mechanism of the colokinetic action of the ghrelin receptor agonist, HM01. Neurogastroenterol Motil 27:1764–1771
Takaki M, Neya T, Nakayama S (1980) Sympathetic activity in the recto-rectal reflex of the guinea pig. Pflugers Arch 388:45–52
Takaki M, Neya T, Nakayama S (1985) Pelvic afferent reflex control of rectal motility and lumbar colonic efferent discharge mediated by the pontine sympatho-inhibitory region in guinea pigs. Pflugers Arch 403:164–169
Chessell IP, Black MD, Feniuk W, Humphrey PP (1996) Operational characteristics of somatostatin receptors mediating inhibitory actions on rat locus coeruleus neurones. Br J Pharmacol 117:1673–1678
Connor M, Bagley EE, Mitchell VA, Ingram SL, Christie MJ, Humphrey PP, Vaughan CW (2004) Cellular actions of somatostatin on rat periaqueductal grey neurons in vitro. Br J Pharmacol 142:1273–1280
Yamanouchi M, Shimatani H, Kadowaki M, Yoneda S, Nakagawa T, Fujii H, Takaki M (2002) Integrative control of rectoanal reflex in guinea pigs through lumbar colonic nerves. Am J Physiol Gastrointest Liver Physiol 283:G148–156
Naitou K, Shiina T, Kato K, Nakamori H, Sano Y, Shimizu Y (2015) Colokinetic effect of noradrenaline in the spinal defecation center: implication for motility disorders. Sci Rep 5:12623. doi:10.1038/srep12623
Naitou K, Nakamori H, Shiina T, Ikeda A, Nozue Y, Sano Y, Yokoyama T, Yamamoto Y, Yamada A, Akimoto N, Furue H, Shimizu Y (2016) Stimulation of dopamine D2-like receptors in the lumbosacral defaecation centre causes propulsive colorectal contractions in rats. J Physiol 594:4339–4350
Millan MJ (2002) Descending control of pain. Prog Neurobiol 66:355–474
Pertovaara A (2006) Noradrenergic pain modulation. Prog Neurobiol 80:53–83
Shiina T, Naitou K, Nakamori H, Sano Y, Ikeda A, Hirayama H, Shimizu Y (2015) Roles of peptides and amines in the regulation of the colorectal motility via the spinal cord. J Physiol Sci 65:S32
Naitou K, Nakamori H, Sano Y, Shiina T, Shimizu Y (2016) Descending adrenergic pathways activate the spinal defecation center in rats. J Physiol Sci 66:S83
Rettig R, Geist R, Sauer U, Rohmeiss P, Unger T (1989) Central effects of somatostatin: pressor response, AVP release, and sympathoinhibition. Am J Physiol 257:R588–R594
Brown MR (1988) Somatostatin-28 effects on central nervous system regulation of vasopressin secretion and blood pressure. Neuroendocrinology 47:556–562
Burke PG, Li Q, Costin ML, McMullan S, Pilowsky PM, Goodchild AK (2008) Somatostatin 2A receptor-expressing presympathetic neurons in the rostral ventrolateral medulla maintain blood pressure. Hypertension 52:1127–1133
Drossman DA (2006) The functional gastrointestinal disorders and the Rome III process. Gastroenterology 130:1377–1390
Longstreth GF, Thompson WG, Chey WD, Houghton LA, Mearin F, Spiller RC (2006) Functional bowel disorder. Gastroenterology 130:1480–1491
Posserud I, Syrous A, Lindström L, Tack J, Abrahamsson H, Simrén M (2007) Altered rectal perception in irritable bowel syndrome is associated with symptom severity. Gastroenterology 133:1113–1123
Ritchie J (1973) Pain from distension of the pelvic colon by inflating a balloon in the irritable colon syndrome. Gut 14:125–132
Mertz H (2003) Review article: visceral hypersensitivity. Aliment Pharmacol Ther 17:623–633
Coveñas R, DeLeón M, Chadi G, Cintra A, Gustafsson JA, Narvaez JA, Fuxe K (1994) Adrenalectomy increases the number of substance P and somatostatin immunoreactive nerve cells in the rat lumbar dorsal root ganglia. Brain Res 640:352–356
Murase K, Nedeljkov V, Randić M (1982) The actions of neuropeptides on dorsal horn neurons in the rat spinal cord slice preparation: an intracellular study. Brain Res 234:170–176
Chapman V, Dickenson AH (1992) The effects of sandostatin and somatostatin on nociceptive transmission in the dorsal horn of the rat spinal cord. Neuropeptides 23:147–152
Mollenholt P, Rawal N, Gordh T Jr, Olsson Y (1994) Intrathecal and epidural somatostatin for patients with cancer. Analgesic effects and postmortem neuropathologic investigations of spinal cord and nerve roots. Anesthesiology 81:534–542
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Funding
This research was supported in part by Grants-in-Aid for Scientific Research (KAKENHI) from the Ministry of Education, Culture, Sports, Science, and Technology of Japan (JP26292164). This research was also supported in part by Grant-in-Aid for JSPS Research Fellow (JP16J03278). KN was supported by Research Fellowships for Young Scientists from the Japan Society for the Promotion of Science. The authors declare no competing financial interests.
Conflict of interest
The authors declare that they have no conflict of interest.
About this article

Cite this article
Naitou, K., Shiina, T., Nakamori, H. et al. Colokinetic effect of somatostatin in the spinal defecation center in rats. J Physiol Sci 68, 243–251 (2018). https://doi.org/10.1007/s12576-017-0524-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12576-017-0524-1