
Abstract
Palynological studies of lacustrine sediments in Eocene maar craters on the Sprendlinger Horst (Hesse, SW Germany) are complemented by 26 core samples from Groß-Zimmern. In addition, diatoms have been studied to assess palaeolimnological conditions. The palynomorph assemblage proves a middle Eocene age more or less coeval to the upper part of the nearby lacustrine succession at Messel. The core includes 33 m of massive to finely laminated bituminous shale abruptly following the underlying breccia of the diatreme filling. The lacustrine deposits are characterised by frequent mass movements and redeposition probably due to tectonic activity in combination with syn- and postsedimentary subsidence in the lake basin. Together with a diatom assemblage characteristic for shallow to moderately deep water, this indicates that the core was drilled in the area of a marginal syncline rather than in the centre of the basin. Freshwater conditions with low nutrient levels were inferred for Lake Groß-Zimmern from diatoms and coccal green algae. Despite the strong disturbance of the sedimentary record, results of cluster analysis and non-metric multidimensional scaling (NMDS) of the palynomorph assemblages can be compared to the undisturbed succession from Messel. This includes successional stages of the azonal vegetation in the crater area during an initial and early recolonisation phase as well as in the zonal vegetation, which are related to the reoccupation of the crater area by a thermophilic forest. Furthermore, slight changes of the climate can be inferred.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
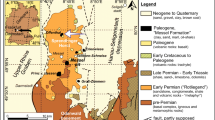
As northern extension of the Hercynian Odenwald basement in SW Germany, the Sprendlinger Horst flanks the Upper Rhine Graben (URG) to the northeast (Fig. 1). The URG is the central part of the European Cenozoic rift system, which trends from the North Sea to the Mediterranean (e.g. Ziegler 1994; Schumacher 2002; Dezes et al. 2004; Reinhold et al. 2016). Early Eocene extensional tectonic activity in the area of the future URG was accompanied by pre-rift volcanism (Feciakova et al. 2007) that led to the formation of maars and pull-apart basins on the Sprendlinger Horst (Backhaus and Rahnama-Rad 1991; Jacoby et al. 2000; Derer et al. 2005). At least half a dozen of these small, isolated basins are filled by lacustrine sediments of Eocene age (e.g. Harms et al. 1999; Felder et al. 2001; Felder and Harms 2004).

Following gravimetric surveys (Jacoby et al. 2001), research cores have been drilled in some of these basins since the end of the 1990s in order to unveil their origin and the character of their lacustrine filling (Felder et al. 2001; Felder and Harms 2004). It could be demonstrated by reaching down into volcanic rocks, mainly volcanic breccias typical for maar-diatreme volcanoes (Lorenz 2000), that the lacustrine records of Lake Messel (Felder and Harms 2004), Lake Offenthal and Lake Groß-Zimmern (Felder et al. 2001) are fillings of maar structures resulting from phreatomagmatic activity. Only for Lake Prinz von Hessen a small pull-apart basin has been suggested (Felder et al. 2001; Felder 2002; Moshayedi et al. 2018) that may have developed due to the reactivation of a fault zone initial to the URG formation (Mezger et al. 2013).
According to their Eocene age and lithological similarities to the well-studied succession at Messel with its excellently preserved fossil assemblage (Gruber and Micklich 2007; Smith et al. 2018), the lacustrine fillings of the other basins were also informally assigned to the “Messel Formation” of Weber and Hofman (1982) by Felder et al. (2001) which is predominantly characterised by bituminous claystones that are often finely laminated and commonly called “oilshale” (Felder et al. 2001).
Maar lake sediments are excellent archives for palaeoenvironmental and palaeoclimatic information (Sabel et al. 2005). Therefore, the Sprendlinger Horst offers the rare opportunity for studying almost coeval and nearby fillings of Eocene maar lakes allowing detailed insight into the vegetation of a limited area within continental Europe during the Paleogene greenhouse. Previous studies already focusing on quantitative high-resolution palynology have been published for the maar lakes of Messel (e.g. Lenz et al. 2007a, 2015, 2017; Lenz and Wilde 2018) and Offenthal (Moshayedi et al. 2020). Our present palynological study of the lacustrine succession from the maar lake of Groß-Zimmern provides a further window into the Paleogene vegetation on the Sprendlinger Horst, even in a highly disturbed succession, by applying multivariate statistical methods such as cluster analysis and non-metric multidimensional scaling (NMDS).
The lacustrine succession at Groß-Zimmern
With a diameter of 250 - 400 m, the maar lake of Groß-Zimmern was significantly smaller than that of Messel, for which a diameter of 650 - 1100 m can be assumed, but similar to the maar lake at Offenthal which had a diameter of about 400 m (Felder et al. 2001). Accordingly, the preserved thicknesses of the lacustrine fillings in the two small basins are comparable (33 m at Groß-Zimmern and 29 m at Offenthal; Fig. 2), while the preserved thickness of the lacustrine filling at Messel is 229 m (Fig. 2; Felder and Harms 2004).

A characteristic sequence of lithological units for maar diatremes (maar lithozones or MLZ; Pirrung 1998), can be recognised in all of the Eocene maar structures on the Sprendlinger Horst (Fig. 2; Felder et al. 2001; Felder and Harms 2004). While MLZ A (diatreme breccia) and MLZ B (syneruptive breccia) mainly comprise the volcanic filling of the maar diatremes, MLZ C includes the first lacustrine sediments with a characteristic succession of partly graded sand and gravel at a dm- to m-scale including wallrock debris (Pirrung 1998; Pirrung et al. 2003). At Messel and Offenthal MLZ C is well preserved indicating a holomictic lake phase (Fig. 2; Lenz et al. 2007a; Moshayedi et al. 2020). However, such deposits are missing in the lacustrine record of the core at Groß-Zimmern. Instead, MLZ C is here characterised by grain-supported breccias between 58.5 and 33 m in the core (Fig. 2; Felder et al. 2001). At 33 m, finely laminated diatom-rich oil-shales including a relatively high proportion of silt and fine sand (Fig. 3; Felder et al. 2001; Felder 2002) start abruptly without transition to the underlying breccia. The deposition of oilshale is characteristic for MLZ D, which includes the transition from event to background sedimentation (Fig. 2; Pirrung 1998; Pirrung et al. 2003). It can therefore be divided in different subzones: While MLZ D1 is characterised by the gradual onset of first oilshale layers within finely bedded or laminated organic-poor silts, MLZ D2 mainly includes laminated to massive oilshale (Pirrung et al. 2003). Therefore, the upper 33 m of the record at Groß-Zimmern most probably can be assigned to MLZ D1 and 2. They are in the focus of our study and characterised by severely disturbed oilshale. Representative parts of the lacustrine succession in the core are presented in Fig. 4, whereas the complete photo documentation can be found in Online Resource 1.

Lacustrine succession of core Groß-Zimmern showing palynozones (PZ) as used in this paper. The position of core images as provided on Figure 4 is indicated

Selected images of the core illustrating the lacustrine record of Groß-Zimmern and its lithozones. The position of the respective palynological samples is indicated by red dots. The images are based on photographs of FIS/HLUG (S. Borges, M. Reich, W. Schiller and M. Stryj) from the core B/97-BK 8. a 10281.pcd-10287.pcd; b 10270.pcd-10276.pcd; c 10247.pcd-10250.pcd; d 10245.pcd-10246.pcd; e 10228.pcd-10234.pcd; f 10209.pcd-10215.pcd
The oilshale up to a depth of 27.35 m is strongly disturbed and characterised by slumping, indicating redeposition of unconsolidated material (Figs. 4a, b). Furthermore, intraclasts of finely laminated oilshale within massive oilshale are typical for this part of the core. In the following part, between 27.35 m and 26.30 m, oilshale is missing (Fig. 3). Instead, a graded layer of medium gravel to sand was deposited, in which intraclasts of various lithologies are embedded. Oilshale reappears above up to a depth of 15.95 m. However, the sediment is still characterised by debris and turbidity flows of graded sands containing intraclasts (Fig. 3). Structures related to slumping are common and the bedding of the oilshale sometimes appears almost vertical (Fig. 4d). Redeposition of oilshale can also be observed in the succeeding part of the core up to a depth of 4.5 m (Fig. 4e). However, disturbance of the sediment is not as strong as in the lower part of the core. The top of the lacustrine sequence is formed by slightly dipping, finely laminated oilshale (Fig. 4f).
The silting-up stage of maar lakes which corresponds to MLZ D3 and MLZ E is characterised by the deposition of peat (Pirrung 1998; Pirrung et al. 2003) which has not been found at Groß-Zimmern. It has either not been deposited or was eroded later (Felder et al. 2001). Instead, the succession is topped by Quaternary sediments at a depth of c. 4.5 m (Fig. 3).
Material and methods
The scientific core B/97-BK 8 was drilled in 1997 in the supposed centre of the basin at Groß-Zimmern (49°52'55.73" N, 8°47'30.10" E) and reached a depth of 116.30 m. The lacustrine succession is limited to the uppermost 33 m of the core (Fig. 2). The lithological description of the fresh core (Fig. 3) has been published by Felder et al. (2001).
Sampling and sample processing for palynological analysis
For palynological analysis of the lacustrine sediments between 33.90 m and 4.90 m of the Groß-Zimmern core, 26 samples were selected at approximately 1 m intervals. For sample processing the microwave digestion technology has been used. Digestion was done by a CEM Corporation Microwave MARS VI system that is equipped with a 360° turn-table that holds up to twelve 100-ml closed pressure Teflon (PFA) vessels of the EasyPrepTM family. They allow gases to escape if the internal pressure becomes too high while the temperature in the vessels is controlled during the entire preparation time. The MARS VI system allows to specifically adjust the power (=temperature) if necessary, which prevents the temperature from rising too quickly or exceeding the target value. All chemicals used were distributed by dispensers with a fixed ml setting only allowing for the addition of certain amounts of chemicals into the desired sample vessels. This ensures uniform and comparable sample procession.
Samples were crushed into small fragments and homogenised. A standard sample size of 4 g was used to minimise variations of reaction parameters such as sample surface area (Ellin and McLean 1994). Together with the sample material, 10 ml of 45% hydrofluoric acid (HF) were filled into a digestion vessel. Six vessels were used at the same time for digestion in the microwave system. The sealed vessels were treated by a standardised process with 15 minutes of heating at 110 °C using 1000-watt power and 45 minutes of cooling. Afterwards, the residue was mixed with 0.5 g of boric acid (H3BO3) to neutralise excess HF thereby improving safety for sample handling (Wilson et al. 2006). The residue in the vessels was then heated again for 5 minutes to 160° C by applying a power of 1000 watt. Following another cooling period of 45 minutes, the residues could be transferred into larger containers, which were filled with demineralised water and regularly decanted until neutralisation. After sieving through a 10 μm mesh screen, the residues were briefly oxidised by applying hydrogen peroxide (H2O2). Subsequently, the residues were sieved again, mixed with 5% potassium hydroxide (KOH) solution, and heated to 80 °C for 10 minutes in the MARS VI system. At the end of sample processing, the remaining residue was provided with a small amount of 35% hydrochloric acid (HCl) to solve remaining fluorides. After final sieving, the residues were transferred into crimp-top vials and 2 drops of 35% HCl were added to provide a slightly acidic environment that prevents fungal infestation.
For each sample permanent slides have been produced by mixing the residue with glycerin jelly. Slides and residues are housed at the Senckenberg Research Institute und Natural History Museum, Frankfurt am Main, Germany.
Quantitative palynological analysis
Palynological analysis was undertaken by using a transmitted light microscope (Olympus BX40). To obtain a representative dataset that can be used for multivariate statistical analysis between 140 and 338 individual palynomorphs per sample were identified and counted at ×400 magnification. Only the lowermost sample at 33.90 m provided an insignificant number of palynomorphs. Therefore, this sample has been specifically highlighted in the pollen diagram and was excluded from statistical analyses. Between 10 and 20% of the palynomorphs in the samples could not be identified and were counted as “Varia” which is similar to the other more or less coeval maar lake fillings of Messel (Lenz and Wilde 2018) and Offenthal (Moshayedi et al. 2020). Palynomorphs were identified using the systematic-taxonomic studies of Thomson and Pflug (1953), Krutzsch (1970a), Thiele-Pfeiffer (1988), and Lenz (2005) (see complete list of taxa and raw data in Online Resources 2 - 4).
For the graphic presentation of the abundance values in percentages PanPlot 11.04 (Alfred Wegener Institut für Polare und Marine Forschung, PANGEA) has been used. In the pollen diagram, the taxa are arranged according to their weighted average value in relation to depth using the WA regression of ter Braak and Looman (1995) and software C2 1.7.6 (Juggins 2007). The structured arrangement of taxa allows to identify the major patterns of compositional variation in relation to depth as well as possible steps in the evolution of the vegetation (Janssen and Birks 1994). For calculation of percentage values pollen and spores were summed up to 100%, whereas algal remains (Botryococcus, Ovoidites) were recorded as additional percentages in percent of the total pollen sum.
Statistical analysis (Cluster analysis and Non-Metric Multidimensional Scaling)
Statistical analyses of the palynological data serve for an objective zonation of the pollen diagram and revealing compositional differences and ecological trends in the vegetation in the vicinity of the maar lake.
To reduce possible sources of error that may occur when determining and counting the palynomorphs, the raw data set has been simplified. Reduction of potential errors in identification has been achieved by grouping together some species of pollen and spores, such as, e.g. trilete spores, which only occur occasionally with single grains ("other trilete spores"), or morphologically similar species of the genus Plicatopollis. The simplification of the data set also reduces the effect of morphological variation, which may lead to a taxonomical over-splitting in pollen and spores compared to the true diversity of parent plants (e.g. Lenz et al. 2021, Wilde et al. 2021). In addition, only those taxa that have been recorded in at least one sample with a frequency of more than 1% were considered for statistical analysis. In doing so the Groß-Zimmern data set has finally been reduced to 44 palynological “variables” (see Online Resource 3).
Statistical analyses are based on standardised raw data values using the Wisconsin double standardisation (Bray and Curtis 1957; Cottam et al. 1978; Gauch and Scruggs 1979; Oksanen 2007). Hereby, the raw data values of each taxon are first divided by its maximum value (species maximum standardisation) and afterwards the abundance of each taxon is represented by its proportion in the sample (sample total standardization; Mander et al. 2010). This excludes the effect of sample size and equalises the influence of rare and abundant taxa on the results of the multivariate analyses (Van Tongeren 1995; Jardine and Harrington 2008; Mander et al. 2010).
A robust zonation of the pollen diagram using software PAST 4.05 (Hammer et al. 2001) was achieved by applying a constrained cluster analysis with the unweighted pair group average method (UPGMA) and the Bray-Curtis dissimilarity as the distance measure. Furthermore, to objectively illustrate differences and similarities between the palynomorph assemblages of the individual pollen zones at Groß-Zimmern, non-metric multidimensional scaling (NMDS; Shepard 1962a, b; Kruskal 1964) has been used. NMDS uses ranked distance values to project the data into a two- or three-dimensional coordinate system (ter Braak 1995; Hammer and Harper 2006). It is the most robust unconstrained ordination method in ecology (Minchin 1987) and has been successfully applied to palynological data in many previous studies (e.g. Jardine and Harrington 2008; Mander et al. 2010; Broothaerts et al. 2014, Lenz and Wilde 2018; Moshayedi et al. 2020). The method avoids the assumption of a linear (e.g. principal component analysis) or unimodal (e.g. correspondence analysis) response model between the individual palynomorph “variables” and underlying environmental gradients. Furthermore, as NMDS is a non-parametric method, normality of the data is not required.
For a joint NMDS of quantitative palynomorph data from Groß Zimmern and Messel, data of 598 samples from Messel (Lenz and Wilde 2018) have been used following Moshayedi et al. (2020). Thereby, the original Messel data set of Lenz and Wilde (2018) has been reduced by excluding outliers in the data set that do not represent the typical palynomorph assemblages, which are characteristic for different evolutionary stages of the vegetation at Messel. According to Moshayedi et al. (2020), the different vegetation phases, which were described for Lake Messel (Lenz et al. 2007a; Lenz et al. 2011; Lenz and Wilde 2018) have been simplified and reduced to five phases. These include the Early Initial Lake Phase (EILP) and the Late Initial Lake Phase (LILP) of the Lower Messel Formation as well as the three phases MMFbase, MMFcentre and MMFtop representing the Middle Messel Formation.
Diatom analysis
The sample from a depth of 27.90 m commonly showed diatom valves in the palynological slide and was therefore chosen for additional diatom analysis. For sample preparation the sediment was not chemically treated but sieved through stacked 125 μm and 63 μm meshes. Permanent diatom slides were subsequently prepared using Naphrax® as mountant.
Light microscope (LM) analysis was performed at ×1000 magnification using a Leica DM 5000 B LM equipped with a ProgRes® CT5 camera with Differential Interference Contrast under oil immersion. The material for SEM was mounted on pieces of photographic film as strew slides, sputtered with gold and observed in a JEOL FM6490 LV.
To assess species abundance, 400 valves were counted. Subsequently, slides were carefully checked to document additional taxa. Valve identification is mainly based on standard references (Krammer and Lange-Bertalot 1986, 1988, 1991a, 1991b; Houk et al. 2017; Lange-Bertalot et al. 2017) and relevant taxonomic publications (e.g. Crawford et al. 2003; Siver et al. 2010). Because of preservation effects and identification mainly to genus level, only qualitative ecological interpretations were made. These are based on the autecology of A. italica and known habitat requirements of the identified genera.
Results
Palynology
The occurrence of the most abundant pollen, spores, and algae in the lacustrine record at Groß-Zimmern is presented in the pollen diagram of Fig. 5. Based on a constrained cluster analysis (Fig. 6) four palynological zones (PZ) can be distinguished (PZ 2 to PZ 5), whereby PZ 2 is further divided into subzones 2a and 2b. The sample at 33.90 m (PZ 1) was excluded from cluster analysis and NMDS as only 32 palynomorphs have been identified in total by scanning several slides. Therefore, the sample has been distinguished from the other samples as a separate PZ.

Pollen diagram showing the most important taxa in the lacustrine succession of Lake Groß-Zimmern. The diagram shows the five palynological zones (PZ 1 to PZ 5) as derived from constrained cluster analysis (see Fig. 6). A algae; P pollen; S spore; F fungal spore

Results of a constrained cluster analysis (UPGMA) of 25 samples from Lake Groß-Zimmern between 32.90 m and 4.90 m. Samples are labelled with their core depth in meters. The sample at 33.90 m that is assigned to palynological zone 1 (PZ 1) has been excluded from cluster analysis, since the low number of palynomorphs in the sample is not of statistical significance
Palynozone 1
PZ 1 is represented by the lowermost sample in the lacustrine record at 33.90 m which is characterised by a low number of poorly preserved palynomorphs without statistical significance. Pollen of Pinaceae (Pityosporites labdacus, Fig. 7g) are dominant. In addition, there are fern spores (Punctatosporites palaeogenicus), pollen of Cupressaceae s.l. (Inaperturopollenites spp., Fig. 7h) and of some angiosperms, such as Triatriopollenites excelsus (Myricaceae, Fig. 7n, o), the Tricolporopollenites cingulum group (Fagaceae, Fig. 8b-d) and Emmapollis pseudoemmaensis (Chloranthaceae, Fig. 7j). With 25%, the highest amount of poorly preserved and unidentifiable palynomorphs ("Varia") among the studied samples has been noted here.

Pollen and spores from the lacustrine record at Lake Groß-Zimmern. The scale bar represents 10μm. Slide numbers and related depths are given in brackets, a Leiotriletes microadriennis (Schizaeaceae; GZ-113, 28.28 m); b Intrapunctisporis sp. (Pteridaceae; GZ-225, 4,90 m); c Polypodiaceoisporites gracillimus (Polypodiaceae; GZ-133, 15.90 m); d Laevigatosporites haardtii (Polypodiaceae; GZ-86, 23.90 m); e Verrucatosporites favus (Polypodiaceae; GZ-143, 14.90 m); f Reticuloidosporites dentatus (Polypodiaceae; GZ-76, 24.90 m); g Pityosporites labdacus (Pinaceae; GZ-204, 7.90 m); h Inaperturopollenites concedipites (Cupressaceae; GZ-153, 13.90 m); i Milfordia minima (Restionaceae; GZ-133, 15.90 m); j Emmapollis pseudoemmaensis (Chloranthaceae; GZ-14, 31.90 m); k Monocolpopollenites tranquillus (Palmae; GZ-54, 27.90 m); l, m Punctilongisulcites microechinatus (Hydrocharitaceae; GZ-4, 32.90 m, GZ-184, 9,90 m); n Triatriopollenites excelsus (Myricaceae; GZ-76, 24.90 m); o Triatriopollenites excelsus minor (Myricaceae; GZ-76, 24.90 m); p Triatriopollenites rurensis (Myricaceae; GZ-96, 22.90 m); q Plicatopollis plicatus (Juglandaceae; GZ-34, 29.95 m); r Plicatopollis lunatus (Juglandaceae; GZ-86, 23.90 m); s Plicatopollis hungaricus (Juglandaceae; GZ-54, 27.90 m); t Momipites punctatus (Juglandaceae; GZ-4, 32.90 m); u Pterocaryapollenites stellatus (Juglandaceae; GZ-184, 9,90 m); v Subtriporopollenites magnoporatus (Juglandaceae; GZ-54, 27.90 m); w Celtipollenites intrastructurus (Ulmaceae; GZ-173, 11.90 m); x Subtriporopollenites constans (Juglandaceae; GZ-54, 27.90 m); y Polyporopollenites eoulmoides (Ulmaceae; GZ-76, 24.90 m); z Intratriporopollenites pseudoinstructus (Tiliaceae; GZ-24, 30.95 m)

Pollen and spores from the lacustrine record at Lake Groß-Zimmern. The scale bar represents 10μm. Slide numbers and related depths are given in brackets. a Bombacacidites reticulatus (Bombacaceae; GZ-34, 29.95 m); b Tricolporopollenites cingulum fusus (Fagaceae; GZ-204, 7.90 m); c Tricolporopollenites cingulum pusillus (Fagaceae; GZ-54, 27.90 m); d Tricolporopollenites cingulum oviformis (Fagaceae; GZ-86, 23.90 m); e Tricolporopollenites megaexactus (Cyrillaceae; GZ-34, 29.95 m); f Tricolporopollenites satzveyensis (Cornaceae; GZ-354, 29.9 m); g Tricolporopollenites edmundii (Cornaceae; GZ-14, 31.90 m); h Tricolporopollenites microreticulatus (Oleaceae; GZ-54, 27.90 m); i Tricolporopollenites marcodurensis (Vitaceae; GZ-204, 7.90 m); j, k Tricolpopollenites parmularius (Eucommiaceae; GZ-96, 22.90 m, GZ-215, 5.90 m); l Reticulataepollis intergranulatus (Nyctaginaceae; GZ-86, 23.90 m); m Triatriopollenites crassostriatus (Solanaceae; GZ-14, 31.90 m); n Nyssapollenites kruschii (Nyssaceae; GZ-14, 31.90 m); o Tetracolporo-pollenites manifestus contractus (Sapotaceae; GZ-173, 11.90 m); p Tetracolporopollenites sapotoides (Sapotaceae; GZ-113, 20.28 m); q Tricolporopollenites eofagoides (unknown botanical affinity; GZ-76, 24.90 m); r Tetracolporopollenites sp. (unknown botanical affinity; GZ-54, 27.90 m)
Palynozone 2 and its subdivision
Seven samples between 32.90 m and 27.00 m represent PZ 2, whereby the uppermost sample shows a slightly different composition of the palynomorph assemblage and can be separated as subzone 2b from the other samples forming subzone 2a (Fig. 6). The compositional difference is also shown by NMDS, since the sample from 27.00 m depth (=PZ 2b) is plotted in the ordination space at the negative end of NMDS axis 1, clearly separate from the samples of PZ 2a (Fig. 9a). This is due to the occurrence of the coccal green alga Botryococcus (Fig. 9b), which already appears regularly in PZ 2a, but reaches the maximum for the entire record with 8.6% in PZ 2b.

Non-metric multidimensional scaling (NMDS) of palynological data of 25 samples from the lacustrine succession of Lake Groß-Zimmern using the Bray-Curtis dissimilarity and the Wisconsin double standardised raw data values. a Scatter plot of the first two axes showing the arrangement of samples. The different symbols represent samples from the palynological zones PZ 2a/b – 5 (see Fig. 6). The arrow illustrates the temporal succession of palynozones. b Scatter plot of the first two axes showing the arrangement of taxa. The dots represent the different samples (see a)
The dominant elements in PZ 2 are the fagaceous pollen Tricolporopollenites cingulum/Tricolpopollenites liblarensis, which reach their maximum value here with 19.6% at a depth of 31.90 m. However, their abundance is steadily decreasing by more than 12% towards the top of PZ 2. In contrast, Plicatopollis spp. (Juglandaceae, Fig. 7q-s) increase from less than 7% at the base of PZ 2a to 18.6% at the top. In spite of a slight decrease there is still a dominance of Plicatopollis spp. compared to fagaceous pollen in PZ 2b
Other elements from the surrounding forest in the vicinity of the maar lake, which often occur in PZ 2, are pollen of the Triporopollenites robustus/rhenanus group (Myricaceae, up to 6%), Triatriopollenites rurensis/bituitus (Myricaceae, up to 9.4%; Fig. 7p) and Momipites punctatus/quietus (Juglandaceae, subfamily Engelhardioideae, up to 8.1%, Fig. 7t). They show their maximum distribution for the complete record in zone PZ 2a.
Further common elements in PZ 2 are Inaperturopollenites spp. (Cupressaceae s.l., up to 10.4% in PZ 2a), which, however, become primarily more abundant in the subsequent PZ 3, as already heralded by a value of 15.4% in PZ 2b. In contrast, pollen of the Nyssaceae (Nyssapollenites spp., Fig. 8n), which are often associated with Cupressaceae s.l. in Eocene swamp forests of the region (Moshayedi et al. 2020, 2021), is almost missing in PZ 2a and reach the maximum value for the entire record with only 3.6% in zone PZ 2b.
Other elements such as Emmapollis pseudoemmaensis (Chloranthaceae) or Monocolpopollenites tranquillus (Palmae, Fig. 7k) occur regularly in lower numbers but more commonly than in the following PZs. In contrast, Milfordia minima (Restionaceae, Fig. 7i) also occurs consistently but does not reach the values of some samples in the subsequent PZs.
Palynozone 3
Seven samples between 25.90 m and 19.30 m can be assigned to PZ 3 (Fig. 6). In the NMDS ordination space, these samples are plotted in the lower left corner separate from the subsequent samples of PZ 4 and 5 (Fig. 9a) and there is only a slight overlap with the older PZ 2a (Fig. 9a).
There is a striking dominance of juglandaceous pollen of Plicatopollis spp. in PZ 3, which already show an increase at the top of PZ 2, but now rise to values between 22.9% and 29.4%, by far the highest for the entire succession. They clearly dominate compared to the fagaceous pollen Tricolporo-pollenites cingulum/Tricolpopollenites liblarensis which are also common but only reach a maximum of 10.9%.
Inaperturopollenites spp. (Cupressaceae s.l.) also occur regularly in PZ3 with values around 11% which is a slight increase compared to PZ 2a. In contrast, other taxa frequent in PZ 2 show a clear and abrupt decline in abundance. This concerns pollen of Myricaceae (Triatriopollenites rurensis/bituitus), which rise to 5.1% in a single sample of PZ 3, but are significantly less common otherwise. The Triporo-pollenites robustus/rhenanus group (Myricaceae) as well as Momipites spp. (Juglandaceae) occur only with low percentages or as single grains.
The high values for Nyssapollenites kruschii and Botryo-coccus as recorded in PZ 2b do not continue in PZ 3. Most of the remaining taxa are represented with frequency values comparable to those in PZ 2a.
Palynozone 4
Nine samples between 18.40 m and 7.90 m are included in PZ 4 (Fig. 6). In the NMDS ordination space, these samples plot on the positive side of NMDS axis 1, clearly separate from both, the older samples of PZs 2 and 3 and the two subsequent samples of PZ 5 (Fig. 9).
In PZ 4, a change in the dominance of juglandaceous (Plicatopollis spp.) and fagaceous (T. cingulum/T. liblarensis) pollen can be observed again. In samples from the middle of PZ 4, T. cingulum/T. liblarensis reach their maximum distribution for the complete record with up to 24.7%. Here they are clearly more abundant than Plicatopollis spp, except for the base and top where Plicatopllis spp. are slightly more frequent (20.6%) than fagaceous pollen.
Pollen of the Cupressaceae s.l. (Inaperturopollenites spp., up to 12.5%) are also common in PZ 4, but less abundant than in the older PZ 3. The proportions of myricaceous pollen such as Triatriopollenites rurensis/bituitus and the Triporopollenites robustus/rhenanus group also decrease significantly compared to PZs 2 and 3. Botryococcus, which occurred regularly to frequently in the older PZs is even missing in some samples of PZ 4.
In contrast, some taxa show higher values for the first time in the studied record. This applies, for example, to Tetracolporopollenites spp. (Sapotaceae, up to 4.4%; Fig. 8o, p, r), Tricolporopollenites pseudocingulum (Styraca-ceae?, Fagaceae?, up to 5.1%), Tricolporopollenites megaexactus (Cyrillaceae, up to 3.8%, Fig. 8e) and Tricolporopollenites microreticulatus (Oleaceae, up to 9.5%, Fig. 8h).
Palynozone 5
The uppermost two samples (5.90 m and 4.90 m) can be separated as PZ 5 (Fig. 6). In the NMDS ordination space, PZ5 is situated between PZ 3 and PZ 4 on the negative side of NMDS axis 2 thus indicating similarities to the pollen assemblages of PZ 3 and PZ 4 (Fig. 7).
Compared to PZ 4, the differences are related to few taxa. Plicatopollis spp. (Juglandaceae, up to 21.2%) and T. cingulum/T. liblarensis (Fagaceae, up to 17.6%) remain dominant and inaperturate pollen of the Cupressaceae s.l. (up to 9.1%) are still common. Pollen of Eucommiaceae (Tricolpopollenites parmularius, Fig. 8j, k) show a significant increase reaching their maximum abundance for the whole succession with 4.6% in the uppermost sample. In contrast, some taxa which were common in PZ 4, such as Tetracolporopollenites spp. or Tricolporopollenites pseudo-cingulum decrease in abundance to values comparable to those for PZ 3.
The diatom assemblage
In total, seven diatom genera were observed (Fig. 10) with the centric species Aulacoseira italica being overall dominant (>99% abundance). Eunotia, Stauroneis, Nupela, and Ulnaria occur very rarely, while only a single specimen of both, Pinnularia and ?Achnantidium was found.

Diatoms from the lacustrine record at Groß-Zimmern (sample at 27.90 m depth). a, b Scanning Electron Microscope images, scale bar = 5 μm, c-k Light Microscope images, scale bar = 10 μm. a - e Aulacoseira italica; f Ulnaria sp.; g Eunotia sp.; h ?Achnanthidium sp.; i Stauroneis sp.; j Nupela sp.; k Pinnularia sp.
However, a preservation and/or sieving related abundance bias cannot be excluded. The heavily silicified A. italica valves were often intact, while valves of other taxa were generally broken and therefore problematic to assign to species level. Moreover, Ulnaria species, for example, often exceed a length of 125 μm and were possibly for the most part excluded from the analysed material by sieving, except for smaller fragments. Observed quantities and diversity therefore may not be fully representative of the original assemblage. Nonetheless, A. italica likely was the most abundant species.
Discussion
Age of the succession
For a stratigraphic analysis the palynomorph assemblage of Lake Groß-Zimmern has been compared to stratigraphically relevant Paleogene microfloras in northwestern Central Germany (Pflug 1986) and Eastern Germany (Krutzsch 1966, 1970a). Detailed stratigraphic information for palynomorphs occurring in the nearby record of Lake Messel has been provided by Thiele-Pfeiffer (1988).
In general, the pollen assemblage from Lake Groß-Zimmern includes taxa typical for the Paleogene. Most of them are known in Central Europe between the upper Paleocene and the Oligocene (Fig. 11). Only few taxa have their first occurrence in the Eocene such as the different species of sapotaceous pollen (Tetracolporopollenites spp.) or the pollen of Pterocarya (Pterocaryapolenites stellatus, Fig. 7u). Other taxa such as Intratriporopollenites pseudoinstructus (Tiliaceae, Fig. 7z) or Bombacacidites reticulatus (Fig. 8a) disappear after the middle Eocene or decrease significantly in abundance (Fig. 11).

All of the pollen and spore taxa from Groß-Zimmern are also known from the nearby lacustrine record of Messel (see Thiele-Pfeiffer 1988) which is of latest early Eocene to early middle Eocene age (Lenz et al. 2015). Due to an almost identical composition of the pollen assemblage it can be assumed that the lacustrine sequence at Groß-Zimmern is of a similar age. Accordingly, the palynomorph assemblage of the samples from Lake Groß-Zimmern fits best to zones Hu4 to Ho2 of Pflug (1986) and 14 to 15 of Krutzsch (1966), which correspond to a middle Eocene (Lutetian/Bartonian) age (Fig. 11). Index species typical for the lower Eocene (Krutzsch 1970a) such as Pistillipollenites mcgregorii, which occurs regularly in the deepest part of the Lower Messel Formation at Messel (Lenz and Wilde 2018) and in the lacustrine sequence of the neighbouring Lake Prinz von Hessen (Moshayedi et al. 2018), are missing at Groß-Zimmern. Few taxa that have been found at Groß-Zimmern are even restricted to the middle Eocene, such as Punctilongisulcites microechinatus (Fig. 7l, m), which has been described for Central Europe so far only from Lake Messel (Thiele-Pfeiffer 1988) and the Helmstedt Lignite Mining District (Lenz 2005), or Bombacacidites cf. reticulatus, which has been found in Messel (Thiele-Pfeiffer 1988) and in the Geiseltal area (Krutzsch 1970b). The quantitative composition and diversity of the palynomorph assemblage at Groß-Zimmern is also in accordance with a typical middle Eocene association. Especially the high diversity and mass-occurrence of juglandaceous pollen of the genus Plicatopollis is characteristic for Zone 15 of Krutzsch (1970a) (Fig. 11).
Among the diatom assemblage almost all genera are known since the early or middle Eocene and therefore in accordance with palynological age determination. The genus Aulacoseira was even found in Late Cretaceous deposits (Benson et al. 2012 and references therein). On the other hand, the earliest record of Ulnaria (as Synedra) was hitherto documented from the late Eocene, although Siver et al. (2018) report a poorly preserved morphotype from Upper Cretaceous deposits in Canada which’s form could fit a Fragilaria or small Ulnaria/Synedra taxon. Nonetheless, Lake Groß-Zimmern now probably represents the earliest confirmed record for this genus.
The disturbance of the lacustrine succession
The most striking differences compared to the lacustrine records of the other maar lakes which were studied on the Sprendlinger Horst (Messel and Offenthal) are the strongly disturbed succession and the lack of well-layered clastic but oilshale-free sediments at the base of the lacustrine succession at Groß-Zimmern. According to the standard succession of lithological units in maar lakes (Pirrung 1998), sediments of a holomictic initial lake phase, which are well developed at Messel and Offenthal (Lenz et al. 2007a; Moshayedi et al. 2020), should also be expected for Lake Groß-Zimmern. However, at Groß-Zimmern oilshale abruptly follows the breccia of MLZ C without any transition. While in the maar lakes of Messel and Offenthal a continuous and almost undisturbed sequence of lacustrine sediments is developed (Felder et al. 2001; Felder and Harms 2004), a strongly disturbed succession mainly of oilshale belonging to MLZ D1 and 2 is characteristic for Lake Groß-Zimmern.
In the area of the Sprendlinger Horst a high tectonic activity can be assumed during the early and middle Eocene, which predates the formation of the URG but is accompanied by pre-rift volcanism (Feciakova 2007). Considerable influence of tectonic activity has been demonstrated in particular for the lower Eocene Lake Prinz von Hessen, located approx. 5 km west of Lake Groß-Zimmern (Fig. 1; Moshayedi et al. 2021). The lacustrine sequence of this small pull-apart basin is characterised by frequent and abrupt lithofacies changes, which point to periods of rapid uplift and subsidence during the evolution of the basin (Moshayedi et al. 2018). The frequent occurrence of mass flows, turbidites and redeposition in the sedimentary succession of Lake Prinz von Hessen has been assigned to tectonic events including earthquake tremor (Moshayedi et al. 2021). This scenario is supported by the fact that a chain of Eocene basins (Prinz von Hessen, Messel, Sellborn-Schneise and Erlenwiese; Fig. 1) is arranged along a concealed SW - NE striking major fault zone, the so-called Messel-Störungszone (Jakoby et al. 2000; Schulz et al. 2003) or Messel Fault Zone (MFZ), which was reactivated due to the initiation of the URG (Mezger et al. 2013). Lake Groß-Zimmern is slightly outside the MFZ, but tectonic activity may have also influenced areas in the vicinity.
Furthermore, processes within the underlying maar diatreme certainly played a role in the disturbance of the lacustrine sequence. Due to later diagenesis, the filling of a maar diatreme is subject to a long period of compaction and may thus be affected by syn- and postsedimentary subsidence causing deformation of the succession (Suhr et al. 2006). Due to subsidence and compaction of fine-grained sediment in the centre of maar lakes their filling often shows a bowl-shaped deformation (Suhr et al. 2006). However, the marginal facies in the respective basins frequently forms so-called “Randmulden” or marginal synclines (Suhr et al. 2006). They are probably related to both later compaction of the marginal facies and asymmetric subsidence of the funnel-shaped diatreme filling. Since space is restricted in a funnel-shaped structure the sedimentary filling may become more and more folded with increasing subsidence, especially at the margin of a diatreme (Suhr et al. 2006). A comparison with the similar-sized maar structure at Offenthal now suggests that the core at Offenthal was drilled more in the centre of the basin with largely undisturbed sediments whereas the core at Groß-Zimmern may have penetrated a more marginal area of the basin, where disturbance and redeposition of sediments due to compaction is expressed.
Correlation of lithological and palynological changes
When comparing compositional changes of the palynomorph assemblages from Lake Groß-Zimmern with lithological changes as observed in the core, a correlation can be seen on Fig. 5 already. The degree of disturbance and redeposition may thus have had an influence on the composition of the pollen assemblages as preserved in the samples. The sample representing PZ 1 that is characterised by an extremely low number of badly preserved palynomorphs, originates from the top of the grain-supported breccia of MLZ C, which is free of oilshale and composed of debris flows from the crater rim and crater wall (Felder et al. 2001). The change to MLZ D with the abrupt onset of oilshale is accompanied by the onset of a diverse and well-preserved microflora. While oilshale dominates in PZ 2a, it is missing in the turbidity and debris flow deposit of PZ 2b between 27.35 m and 26.30 m (Fig. 3). Redeposition here led to additional transport of palynomorphs from the edge of the lake into the basin which may be the reason for the slight change in composition of the palyno-morph assemblages from PZ 2b to PZ 2a. The lacustrine succession comprised by PZ 3 is also characterised by redeposition leading to turbidity and debris flows including bodies of oilshale partly showing a mor or less vertical lamination (Fig. 4d). The redeposited oilshale is sometimes even finely laminated (Fig. 4c) indicating longer periods with oligo- to meromictic conditions in the lake. Redeposited oilshale can also be observed in PZ 4 (Fig. 4e), but disturbance is no longer as strong as in the lower part of the lake deposits. Finally, at the top of the sequence a change in the palynomorph assemblage is documented by PZ 5 which shows almost undisturbed and finely laminated oil shale.
Reconstruction of the palaeoenvironment
Since the lacustrine filling in the maar lake of Groß-Zimmern is intensely disturbed any interpretation of palynological assemblages in terms of palaeoenvironmental trends and cyclicity must be treated with caution. Nevertheless, the micro-floristic assemblages can be compared with the plant communities as recorded at other sites on the Sprendlinger Horst.
The recolonisation of the crater walls and the devastated areas in the vicinity of the Eocene maar structures on the Sprendlinger Horst can be assigned to different vegetational phases, which are characterised by continuous gradual changes in the composition of the vegetation communities (Moshayedi et al. 2020). In the initial phase, immediately after a maar crater had formed, the surroundings were devoid of vegetation (Moshayedi et al. 2020). The development of a herbaceous and shrubby pioneering vegetation is characteristic for the subsequent recolonisation phase (Lenz et al. 2007a). The pioneering flora invaded the disturbed areas around the maar lake in successional stages, but the zonal forest vegetation reestablished around the lake not before the following recovery phase (Lenz et al. 2007a; Moshayedi et al. 2020). Due to the increasing vegetation cover of the crater wall and the related stabilisation of the slopes a change from holomictic to meromictic conditions and the onset of oilshale deposition in the lake is characteristic for the recovery phase (Lenz et al. 2007a). A robust climax vegetation follows during the climax phase, which is characterised by pollen assemblages without significant changes in systematic composition (Lenz et al. 2011, 2015). Slight quantitative changes can be assigned to minor changes in the climate (Lenz and Wilde 2018).
At Messel, all phases between the initial recolonisation and the establishment of a final climax vegetation can be distinguished (Lenz et al. 2015) by specific pollen assemblages. Therefore, the lacustrine succession at Messel may serve as a reference section for the succession of the vegetation on the Sprendlinger Horst following devastation by phreatomagmatic eruptions and a joint NMDS of samples from Lake Messel and Lake Groß-Zimmern was carried out for comparison (Fig. 12).

Non-metric multidimensional scaling (NMDS) of palynological data of 25 samples from the lacustrine succession of the Groß-Zimmern maar lake and 598 samples from the lacustrine succession of the Messel maar lake using the Bray-Curtis dissimilarity and the Wisconsin double standardised raw data values. a Scatter plot of the first two axes showing the arrangement of samples. The different symbols represent samples from the different palynozones at Groß-Zimmern and phases at Messel (see Lenz and Wilde 2018); b Scatter plot of the first two axes showing the arrangement of taxa. The dots represent the different samples (see a). c Scatter plot of the first two axes showing the different vegetational phases during the recolonisation of the volcanically disturbed sites on the Sprendlinger Horst (see Moshayedi et al. 2020). The coloured dots indicate the median NMDS scores for the record of Lake Groß-Zimmern and for the different vegetation phases at Messel (see Lenz and Wilde 2018)
In the NMDS ordination space, the samples from Messel are generally ordered from right to left along NMDS axis 1 from the oldest to the youngest samples (Fig. 12a). This indicates that the vegetation at Messel is characterised by a gradual and continuous change during the time of deposition of the lacustrine filling. On the right side, the samples from the Early Initial Lake Phase (EILP, Lenz et al. 2007a) are plotted, representing the recolonisation phase of the devastated area by a pioneering vegetation mainly characterised by a diverse assemblage of ferns while the intermittent deposition of oilshale started. During the Late Initial Lake Phase (LILP) the stability of the crater walls increased and the deposition of oilshale became more and more dominant while fern spores disappear. Therefore, the samples of the LILP are found at the centre of the NMDS ordination space clearly separate from the EILP (Fig. 12b). The change to almost continuous meromictic conditions during the LILP can be related to the reoccupation of the crater area by the forest vegetation during the recovery phase (Lenz et al. 2007a). On the left side of the NMDS ordination space the samples of the Middle Messel Formation (MMF) appear, which represent the climax vegetation (Fig. 12a). Slight compositional changes within the palynomorph assemblages of the MMF especially between the samples from the base (MMFbase), the centre (MMFcentre) and the top (MMFtop) of the succession are caused by a gradual loss of diversity within the thermophilic climax vegetation (Lenz et al. 2011; Lenz and Wilde 2018).
Pollen species characteristic for the MMF, such as Plicatopollis spp. (Juglandaceae), Tricolpopollenites liblarensis and Tricolporopollenites cingulum (both fagaceous), are also among the dominant taxa of the palynomorph assemblages of Lake Groß-Zimmern. However, the high values described for the MMF at Messel (Lenz et al. 2011) are not achieved. Therefore, a complete reoccupation of the crater area by the climax forest vegetation is obviously not covered by the pollen record here. Accordingly, the samples from Lake Groß-Zimmern are plotted at the positive end of the NMDS axis 1 together with the samples from the EILP of Lake Messel and clearly separate from the samples of the subsequent LILP (Fig. 12a). Thus, almost all samples fall into the range of the recolonisation phase at Lake Messel and Lake Offenthal (Fig. 12c) and only few samples are plotted in the ordination space in the transition to the recovery phase. This may indicate that just the beginning of the gradual reestablishment of a forest vegetation in the crater area is preserved in the lacustrine filling of Lake Groß-Zimmern.
At Messel (Lenz et al. 2007a) and Offenthal (Moshayedi et al. 2020) the recolonisation phase (=EILP) is documented predominantly in the holomictic oilshale-free lake phase. However, oilshale was formed in both lakes during the terminal recolonisation phase. Since the recolonisation phase is recorded entirely in oilshale-dominated deposits at Groß-Zimmern, this indicates that only a late phase is preserved in the succession (see below). Therefore, a hiatus has to be inferred between the breccia (PZ 1) and the oil shale (PZ 2).
Thus, the lacustrine succession of the maar lake at Groß-Zimmern only includes the initial phase, the late recoloni-sation phase and the early transition to the recovery phase of the vegetation following the devastation of the area by a phreatomagmatic eruption. This almost corresponds to the record as preserved in the lacustrine succession of Lake Offenthal (Moshayedi et al. 2020) where the recovery phase is more pronounced. However, since both lakes provide a similar thickness of the lacustrine record, which is lower than in Messel by a factor of eight, a significantly shorter time interval is documented at Groß-Zimmern and Offenthal.
Initial phase at Lake Groß-Zimmern
The initial recolonisation phase is represented at Groß-Zimmern by PZ 1, the lowest sample from the base of the lake sediments with few badly preserved pollen and spores. Only a number of pinaceous pollen (Pityosporites labdacus) and rare pollen grains of Myricaceae (Triatriopollenites excelsus), Fagaceae (Tricolporopollenites cingulum), Cupressaceae s.l. (Inaperturopollenites spp.) as well as some fern spores (Punctatosporites palaeogenicus) have been noted. The sample has been taken from a mass-flow breccia (Fig. 3) which was assigned by Felder et al. (2001) to MLZ C just at the beginning of the lacustrine filling when the crater area and the vicinity of the lake were still free of vegetation. Therefore, only pollen from the surrounding vegetation was transported into the crater lake by wind. This applies in particular to pinaceous pollen, which can be transported over long distances and are rare or missing in the rest of the sequence. Wind-dispersed pollen such as those of Pinaceae and Cupressaceae s.l. can even be regarded as typical for the base of lacustrine successions in crater lakes (Rampino and Koeberl 2006) and has also been observed at Offenthal in a similar position (Moshayedi et al. 2020). Rare evidence for pollen of Myricaceae and Fagaceae probably represents the (para)tropical forest as surviving in some distance, and monolete spores may indicate that some ferns initially had colonised the devastated area as it is typical for pteridophytes (Thomas and Cleal 2021).
Recolonisation phase of Lake Groß-Zimmern
The early recolonisation phase at Messel and Offenthal (Lenz et al. 2007a; Moshayedi et al. 2020) is characterised by the deposition of finely layered clastic but still oilshale-free sediments in the lower part of the lacustrine succession (Pirrung 1998). At Groß-Zimmern, however, such sediments are completely missing. Instead, oilshale immediately follows the initial breccia. This suggests that clastic sediments representing a purely holomictic lake phase have been removed later. The lithologic hiatus is confirmed by the palynomorph assemblages showing an abrupt change between the almost barren sample of PZ 1 and the diverse microflora in the samples at the base of PZ 2. In contrast, there is a gradual increase of palynomorphs in the clastic sediments of the holomictic phase of Lake Offenthal documenting the continuous progression of pioneering elements into the crater area at the transition from the initial to the recolonisation phase (Moshayedi et al. 2020).
The early recolonisation phase at Messel and Offenthal is characterised by the frequent occurrence of various fern spores (Lenz et al. 2007a, Moshayedi et al. 2020). At Groß-Zimmern various fern spores such as those of the Leiotriletes/Toroisporis group (Schizaeaceae) and Laeviagtosporites spp. (Polypodiaceae) occur regularly in the samples of PZs 2 to 5 (Fig. 5). However, they do not appear in frequencies that are reached at Messel and Offenthal during the early recoloni-sation phase (Lenz et al. 2007a; Moshayedi et al. 2020).
At the end of the recolonisation phase, ferns were followed by Restionaceae (Milfordia spp.). At Messel and Offenthal pollen of Restionaceae were mainly restricted to the later stage of the recolonisation phase and missing from the succeeding recovery phase (Lenz and Wilde 2018; Moshayedi et al. 2020). Restionaceae invaded the herbaceous communities instead of grasses as observed today on Krakatao or on the crater walls of volcanoes on Papua New Guinea (Taylor 1957). Since grasses were rare during the Eocene the Restionaceae with their grass-like habit and ecology probably occupied their role (Lenz et al. 2007a; Moshayedi et al. 2020). In Lake Groß-Zimmern, pollen of Restionaceae regularly occur with higher values in PZs 2 to 4 and are only missing in the two samples of PZ 5 (Fig. 5). Together with the regular but not abundant occurrence of fern spores, this supports that the recolonisation phase has not been fully preserved at Groß-Zimmern, but only its terminal stage when Restionaceae had replaced ferns in the pioneering vegetation.
Comparable to Offenthal (Moshayedi et al. 2020), high values for Cupressaceae s.l. (Inaperturopollenites spp., Fig. 5) indicate that swamp communities already existed around the lake during the recolonisation phase. This is supported by pollen of Nyssaceae (Nyssapollenites spp.) which regularly appears in the record of Groß-Zimmern (Fig. 5). Together with Nyssaceae, Cupressaceae often formed Nyssa-Taxodium swamps during the Eocene in Central Europe which were similar to those on the warm-temperate coastal plains of eastern North America today (Daubenmire 1978; Box 2015). Again comparable to Lake Offenthal (Moshayedi et al. 2020), floating and submersed aquatic plants existed in the littoral zone in front of such swamp communities as shown by regularly occurring Punctilongisulcites microechinatus (most probably Hydrocharitaceae) in the samples of PZs 2 to 5.
In addition to the accumulation of centric diatoms in the layer at 27.90 m depth, the frequent occurrence of the coccal green alga Botryococcus during PZs 2 and 3 (Fig. 5) shows that the lake experienced algal blooms. Interestingly, Botryococcus is limited to the holomictic lake phase in Lake Offenthal, during which no oilshale was deposited (Moshayedi et al. 2020). In contrast, mass occurrences of Botryococcus have been described from the oilshale of Lake Messel, in particular from the top of the MMF (Lenz et al. 2007b). While in Lake Offenthal Botryococcus is obviously associated to the formation of a stratified lake and the transition to meromictic conditions (Moshayedi et al. 2020), mass occurrences in the upper part of the MMF at Lake Messel have been attributed to increasingly drier conditions at the end of the lake development (Lenz et al. 2007b). However, in both lakes alkalinity probably may have triggered the growth of Botryococcus (e.g. Wetzel 1983; Wilde et al. 1993).
It is known that Botryococcus blooms may result in wind-driven strandline accumulation (De Deckker 1988). Such accumulations may have been transported into the Eocene maar lakes of Eckfeld (Nickel 1996) and Messel (Lenz et al. 2007b) by mass-flows or turbidites which could also explain the peak abundance of Botryococcus in PZ 2b at Groß-Zimmern.
Overall, it is obvious that the quantitative shifts among the pioneering elements and early colonisers in the palynomorph assemblages across the various PZs in the record of Lake Groß-Zimmern are not serious and related only to a few taxa. This is in contrast to the fact that at Messel and Offenthal the pioneering flora invaded the disturbed areas of the craters and their surroundings in successional stages which are charac-terised by significant changes of the palynomorph assemblages (Lenz et al. 2007a; Moshayedi et al. 2020). This is further evidence that only the late recolonisation phase is preserved at Groß-Zimmern.
Palaeoenvironmental changes in the hinterland of Lake Groß-Zimmern
While the azonal vegetation immediately around Lake Groß-Zimmern was hardly subject to change during deposition, wind transported pollen from the surrounding regional vegetation show significant fluctuations in frequency (Fig. 5). This allows for the distinction of 4 phases, which are equivalent to PZs 2 to 5 and characterised by the dominance of different plant families (Fig. 13).

Non-metric multidimensional scaling (NMDS) of palynological data of 25 samples from the lacustrine succession at Groß-Zimmern using the Bray-Curtis dissimilarity and the Wisconsin double standardised raw data values (see Fig. 9) showing different vegetational phases in the zonal climax vegetation in the hinterland of the maar lake
Phase A:
Pollen of Juglandaceae (Plicatopollis spp.) and, in particular, of Fagaceae (T. cingulum) belong to the dominant elements in this phase which is documented by PZ 2. This association is typical for inland sites of Western and Central Europe during the Eocene (Mai 1981, 1995; Schaarschmidt 1988; Wilde 1989; Collinson et al. 2012b). However, in the ordination space of the NMDS of Lake Groß-Zimmern various pollen of Myricaceae (Triatriopollenites rurensis/bituitus, Triatriopollenites excelsus, Triporopollenites rhenanus/robustus) plot together with samples from PZ 2 (Fig. 13) which indicates that members of the family were another common element of the vegetation. During the recolonisation phase of Lake Messel, Myricaceae belonged to a fast-growing pioneering community on the tuff rim (Lenz et al. 2007a). In particular, the genus Comptonia was regarded as a pioneer element already by Mai (1995).
Ulmaceae are well known at Messel from pollen, leaves and fruits (Thiele-Pfeiffer 1988; Wilde 1989; Collinson et al. 2012a). They may have been early colonisers of the volcanic substrate on the tuff rim (Lenz et al. 2007a), probably with the extinct genus Cedrelospermum as characteristic representative (Wilde and Manchester 2003). Pollen of Ulmaceae (Poly-poropollenites spp.) is also regularly represented in the pollen assemblages of Lake Groß-Zimmern, indicating their important role in the pioneer vegetation during the recolonisation phase.
The high values for T. cingulum and Plicatopollis spp. show that wind-pollinated Fagaceae and Juglandaceae dominated within the arboreal climax vegetation still in some distance around Lake Groß-Zimmern. However, high proportions of pollen of Myricaceae and Ulmaceae show that these fast-growing plants may have formed a transitional community on the volcanic substrates in the area between the lake itself and the climax forest in the background during reoccupation of the devastated area.
Phase B:
The next phase of the vegetation in the hinterland of Lake Groß-Zimmern is documented by PZ 3 with pollen of Myricaceae clearly decreasing in frequency. This may indicate that the thermophilic climax forest gradually approached the vicinity of the lake. Accordingly, pollen of Fagaceae (T. cingulum) and Juglandaceae (Plicatopollis spp.) now dominate the pollen assemblage.
Subordinate changes in the composition of the mid-Eocene climax vegetation, especially with regard to Fagaceae and Juglandaceae can be assigned to Milankovitch cyclicity (Lenz et al. 2011, 2015). During times of higher mean annual temperature (MAT) and lower seasonality with more balanced rainfall Fagaceae dominated within the climax vegetation. Therefore, the dominance of the juglandaceous pollen in PZ 3 of Lake Groß-Zimmern may indicate a period of slightly decreasing MAT but also drier conditions (see discussion in Lenz et al. 2015, Lenz and Wilde 2018).
Furthermore, the palynomorphs in the upper part of PZ 3 are significantly darker than in other parts of the lacustrine record at Groß-Zimmern. The higher thermal alteration (Fig. 14) is accompanied by a qualitative increase of black particles (charcoal). Both may be an effect of wildfires in the vicinity of the lake and support the assumption of an even drier period for the upper part of PZ 3.

a Colour chart indicating different thermal alteration of palynomorphs in different depths of the lacustrine record of the maar lake at Groß-Zimmern; b Box-and-whisker-plot for the three parts of the lacustrine record marked with different colours in a; c Results of a Kruskall Wallis test and Mann-Whitney pairwise Bonferroni corrected p-values indicating that the lacustrine part between 22.90 m and 19.30 m depth includes significant darker palynomorphs than other parts of the record; d Examples of palynofacies pictures indicating that the part between 22.90 m and 19.30 m depth is characterised mainly by opaque and semi-opaque phytoclasts, whereas other parts of the record show mainly translucent phytoclasts
Phase C:
In this phase, which is equivalent to PZ 4, the thermophilic forest was at least temporarily dominated by Fagaceae (T. cingulum). Lenz and Wilde (2018) described a similar association of fagaceous pollen with thermophilous and wetland taxa as well as aquatic/hydrophilous elements for the maar lake of Messel. Correspondingly, T. cingulum is plotted together with pollen of thermophilous taxa such as Sapotaceae (Tetracolporopollenites spp), Cornaceae (Tricolporopollenites satzveyensis, Mastixia), Vitacae (Tricolporopollenites marcodurensis, Cissus) and aquatic plants such as Hydrocharitaceae (Punctilongisulcites microechinatus) in the ordination space of the NMDS from Groß-Zimmern (Fig. 13). This confirms a general climatic change towards increasing temperatures and higher and balanced precipitation in PZ 4.
Despite strong disturbance of the sedimentary succession in the core from Groß-Zimmern, the pollen diagram (Fig. 5) shows gradual transitions between the Juglandaceae and Fagaceae dominated forests. This demonstrates that disturbance did not affect a proper succession of the pollen record in the lacustrine deposits.
Phase D:
In the last phase, which consists of the two samples of PZ 5, the gradual transition to the recovery phase at Lake Groß-Zimmern may have been captured. Typical elements of the recolonisation phase such as the Restionaceae (Milfordia spp.) disappear as in the beginning of the recovery phase at Messel and Offenthal (Lenz and Wilde 2018; Moshayedi 2020).
In addition, other elements from the thermophilic forest appear for the first time with higher values in the two samples of PZ 5, such as, e.g. Eucommiaceae (Tricolpopollenites parmularius). Furthermore, both, juglandaceous (Plicatopollis spp.) and fagaceous pollen (T. cingulum) now appear with high values (Fig. 5). In addition to the disappearance of the pioneer elements, this strongly indicates that the zonal forest had reached the crater area. The common occurrence of pollen from Myricaeae (T. rurensis/bituitus) and Ulmacae (Polyporopollenites spp.) could indicate that the crater wall itself was still colonised by these fast-growing plants.
In the NMDS, the samples from PZ 5 are plotted next to those from PZ 3, but separate from PZ 4 (Fig. 13). However, the low number of samples does not allow for an interpretation in terms of climate here.
Lake characteristics
Diatom-rich sediments occur regularly in the lacustrine succession of Lake Groß-Zimmern (Felder 2002) but only the sample from 27.90 m depth (PZ 2), which lithologically approached a diatomite, was specifically analysed for diatoms. All occurring genera still live in freshwater habitats (Lange-Bertalot et al. 2017). The respective assemblage is dominated by Aulacoseira italica which probably formed a massive bloom. However, because of taxonomic confusion the autecology of the species is not well known. Based on its actual occurrence together with mainly benthic genera (e.g. Gomphonema, Epithemia, Cymbella Eunotia, Pinnularia), some of which have also been observed in the sample from Groß-Zimmern, Crawford et al. (2003) suggest a different habitat compared to open water planktonic species such as A. granulata, with A. italica representing a littoral element in more or less eutrophic lakes, ponds, ditches or streams. A. italica can form long filamentous colonies and has been observed as a bloom forming species in the phytoplankton of lakes during spring (Reuter 1979), although it is not exclusive to planktonic habitats but likely rather tychoplanktonic (Potapova et al. 2010). Heavily silicified Aulacoseira species require turbulence to remain suspended in the photic zone and have therefore been associated with water column mixing and wind-induced turbulence (Lund 1954; Li et al. 2010; Rühland et al. 2015). In Brazilian floodplain lakes, A. italica occurrs mainly in oligotrophic environments, zones of low pollution or less turbid waters and shows the highest abundance in the most acidic lake (Nardelli et al. 2021). Several Aulacoseira species have the capacity to remain buried as resting spores in the bottom sediment for longer periods thus surviving unfavourable conditions in the water column and re-emerge in the epilimnion with turbulence, for example, after heavy rainfall or with strong winds (Edlund et al. 1996; Potapova et al. 2008; Nardello et al. 2021). Resting spores of A. italica are especially common in populations from shallow polymictic lakes and in fossil sediments (Potapova et al. 2010). Although nowadays a relatively rare cosmopolitan species, A. italica was repeatedly found in high abundance in other fossil assemblages (Krammer and Lange-Bertalot 1991; Kiss et al. 2012; Houk et al. 2017).
Many species of the genus Eunotia are also strongly linked to oligo- or dystrophic, low conductivity and acid freshwater habitats and are indicators of a very good ecological quality (Krammer and Lange-Bertalot 1991; Lange-Bertalot et al. 2017). Diatoms of the genus Nupela have been reported from almost all continents from waters with neutral pH and low conductivity (Spaulding and Edlund 2008). The genus is widespread but also often associated with acidic waters (Siver et al. 2010). In a study from the Colombian lowland, Nupela species were found in aquatic ecosystems with low conductivity and a pH ranging from acidic to relatively basic (Sala et al. 2013). Most species of Pinnularia live in oligo- and dystrophic, often acidic, low conductivity freshwater (Krammer and Lange-Bertalot 1986; Lange-Bertalot et al. 2017). Stauroneis species are found mostly in benthic habitats of wetlands and small freshwater lakes and ponds, although some taxa also occur in streams on moss and on moist soils (Bahls 2012), and Ulnaria is generally common in lakes and rivers (Spaulding 2013).
Therefore, the dominance of Aulacoseira italica along with the co-occurring mainly benthic taxa indicates a rather low conductivity, shallow to moderately deep freshwater lake or wetland with a pH probably ranging from somewhat acidic to circumneutral. The co-occurrence of the diatom assemblage with the coccal green alga Botryococcus confirms this interpretation. Although Botryococcus favours more alkaline water (Wetzel 1983; Wilde et al. 1993), it especially prefers oligothrophic conditions (Jankovská and Komárek 2000). Since these are also preferred by several of the present diatom taxa, a limited nutrient supply to the lake at least during PZ 2 can be assumed.
The diatom assemblage points to a shallow to moderately deep water body. This shows that the sediments from the core at Groß-Zimmern including such a diatom assemblage and Botryococcus were either transported from the littoral zone into their present position or that the core was actually drilled in the area of a marginal syncline (“Randmulde”) rather than in the centre of the basin.
Conclusions
The palynological investigation of the various lacustrine fillings of Eocene maar lakes on the Sprendlinger Horst, such as those at Messel, Offenthal and Groß-Zimmern show that the recolonisation of areas devastated by phreatomagmatic eruptions including the reestablishment of the climax vegetation followed similar processes. Despite the intense disturbance of the sedimentary succession in the core from Groß-Zimmern, an interpretation of the palynological succession is possible when compared to the more complete record from Lake Messel.
The strong disturbance of the lacustrine deposits in the core from Groß-Zimmern is unique for the studied Eocene lake deposits on the Sprendlinger Horst. However, it can be speculated that an interplay of tectonic activity and syn- and postsedimentary subsidence in the basin resulted in frequent mass movements from the lake shore and the crater wall including deformation and redeposition of oilshale. However, since similar processes can also be assumed for the maar lakes of Messel and Offenthal, it is obvious that the core from Groß-Zimmern has been drilled in a more marginal position within the crater (“Randmulde”) and not in its central part where only a saucer-shaped deformation of the lake sediments has to be expected (Suhr et al. 2006). This is confirmed by the assemblage of diatoms and coccal green algae, which point to a shallow to moderately deep lake rather than to the deep centre of the lake basin. Additionally, the assemblage indicates freshwater conditions with lower nutrient levels and a rather neutral ph for the lake.
A comparison of the record from the core at Groß-Zimmern with the succession of palynomorph assemblages from Lake Messel (Lenz and Wilde 2018) shows that the initial phase of recolonisation is directly followed by the late recolonisation phase here. The early recolonisation phase with a typical dominance of fast growing pioneering herbaceous plants such as ferns or Restionaceae is missing in the record at Groß-Zimmern. It is therefore likely that the respective deposits were removed by erosion or reworked causing a hiatus between PZs 1 and 2.
The initial stage of the succeeding recovery phase, which is characterised by the reoccupation of the crater area by a thermophilic forest vegetation, may have been recorded at Groß-Zimmern in PZ 5 at the top of the lacustrine succession. However, the majority of the recovery phase, and in particular the following climax phase, is not preserved here. This is comparable to the known record from Lake Offenthal, where the final climax phase is also missing.
Thanks to previous high-resolution palynological studies of the sedimentary records from Lake Messel (e.g. Lenz and Wilde 2018) and Lake Offenthal (Moshayedi et al. 2020) it is well-known which plants were part of the azonal flora and which belonged to the regional zonal vegetation in the area of the Sprendlinger Horst in middle Eocene times. A comparable distinction between a (par)autochthonous and an allochthonous component in the pollen record of the vegetation can also be inferred for Lake Groß-Zimmern. This allows to assume slight variations of climate conditions by the record of changes within the zonal forest vegetation, especially by the changing dominance of pollen from anemophilous Juglanda-ceae and Fagaceae. Therefore, a period with slightly increased temperatures as well as higher and more balanced precipitation prevailed during PZ 4 (dominance of fagaceous pollen), whereas PZ 3 (dominance of juglandaceous pollen) was characterised by slightly drier conditions.
Despite a strong disturbance of the sedimentary record, our palynological study of the lacustrine filling of Lake Groß-Zimmern surprisingly shows that the regular succession of the vegetation is still preserved. Therefore, the lacustrine record of Lake Groß-Zimmern is another example that the fillings of maar lakes are perfect archives for the reconstruction of the palaeoenvironment in deep-time records by using pollen and spores as proxies.
Data availability
All data generated or analysed during this study are included in this published article and its supplementary information files.
References
Backhaus, E., & Rahnama-Rad, J. (1991). Die Rutschgefährdung der Messel-Formation (Fundstätte Messel; Mittel-Eozän). Einflüsse der Tektonik, der Hydrogeologie und der Materialeigenschaften der Gesteine. Courier Forschungs-Institut Senckenberg, 139, 1–69.
Benson, M. E., Kociolek, J. P., Spaulding, S. A., & Smith, D. M. (2012). Pre-Neogene non-marine diatom biochronology with new data from the late Eocene Florissant Formation of Colorado, USA: Strati-graphy, 9(2), 131–152.
Bray, J. R., & Curtis, J. T. (1957). An ordination of the upland forest communities of southern Wisconsin. Ecological Monographs, 27, 325–349.
Broothaerts, N., Verstraeten, G., Kasse, C., Bohncke, S., Notebaert, B., & Vandenberghe, J. (2014). Reconstruction and semi-quantification of human impact in the Dijle catchment, central Belgium: a palynological and statistical approach. Quaternary Science Reviews, 102, 96–110. https://doi.org/10.1016/j.quascirev.2014.08.006
Box, E. O. (2015). Warm-Temperate Deciduous Forests of Eastern North America. In E. Box, K. Fujiwara (Eds.), Warm-Temperate Deciduous Forests around the Northern Hemisphere. Geobotany Studies (Basics, Methods and Case Studies) (pp. 225–255). Cham: Springer. https://doi.org/10.1007/978-3-319-01261-2_13
Collinson, M. E., Manchester, S. R., & Wilde, V. (2012a). Fossil fruits and seeds of the Middle Eocene Messel biota, Germany. Ab-handlungen der Senckenberg Gesellschaft für Naturforschung, 570, 1-251.
Collinson, M. E., Smith, S. Y., Manchester, S. R., Wilde, V., Howard, L. E., Robson, B. E., Ford, D. S., Marone, F., Fife, J. L., & Stampanoni, M. (2012b). The value of X-ray approaches in the study of the Messel fruit and seed flora. In T. Lehmann, & S. F. K. Schaal (Eds.), Messel and the terrestrial Eocene - Proceedings of the 22nd Senckenberg Conference. Palaeobiodiversity and Palaeoenvironments, 92(4), 403-416. https://doi.org/10.1007/s12549-012-0091-7.
Cottam, G., Goff, F. G., & Whittaker, R. H. (1978). Wisconsin Comparative Ordination. In R. H. Whittaker (Ed.), Ordination of Plant Communities. Handbook of Vegetation Science, 5-2 (pp. 185–213). Dordrecht: Springer. https://doi.org/10.1007/978-94-009-7989-5_7.
Crawford, R. M., Likhoshway, Y. V., & Jahn, R. (2003). Morphology and Identity of Aulacoseira italica and typification of Aulacoseira (Bacillariophyta). Diatom Research 18(1), 1–19. https://doi.org/10.1080/0269249X.2003.9705569.
Daubenmire, R. F. (1978). Plant Geography with special reference to North America. New York: Academic Press.
Deckker, P. De (1988). Large Australian lakes during the last 20 million years: sites for petroleum source rock or metal ore deposition, or both? In A. J. Fleet, K., Kelts & M. R. Talbot (Eds.), Lacustrine Petroleum Source Rocks. Geological Society, London, Special Publication, 40, 45–58. https://doi.org/10.1144/GSL.SP.1988.040.01.05.
Derer, C. E., Schumacher, M. E., & Schäfer, A. (2005). The northern Upper Rhine Graben: basin geometry and early syn-rift tectono-sedimentary evolution. International Journal of Earth Sciences, 94(4), 640–656. https://doi.org/10.1007/s00531-005-0515-y.
Dezes, P., Schmid, S. M., & Ziegler, P. A. (2004). Evolution of the European Cenozoic Rift System: interaction of the Alpine and Pyrenean orogens with their foreland lithosphere. Tectonophysics, 389(1-2), 1–33. https://doi.org/10.1016/j.tecto.2004.06.011.
Edlund, M. B., Stoermer, E. F., & Taylor, C. M. (1996). Aulacoseira skvortzowii sp. nov. (Bacillariophyta), a poorly understood diatom from Lake Baikal, Russia. Journal of Phycology, 32, 165–175. https://doi.org/10.1111/j.0022-3646.1996.00165.x.
Ellin, S., & McLean, D. (1994). The use of microwave heating in hydrofluoric acid digestions for palynological preparations. Palynology, 18, 23–31. https://doi.org/10.1080/01916122.1994.9989436.
Feciakova, Z, Mertz, D. F., & Renne, P. R. (2007). Geodynamic setting of the Tertiary Hocheifel volcanism (Germany), Part I: 40Ar/39Ar geochronology. In R. R. Ritter & U. R. Christensen (Eds.), Mantle Plumes, a multidisciplinary approach (pp. 185–206). Berlin, Heidelberg: Springer.
Felder, M. (2002). Paläolimnologische Untersuchungen zu Siderit- und Aragonitbildung in schwarzpelitdominierten paläogenen Seen Mitteleuropas. Dissertation, University of Jena.
Felder, M., & Harms, F. J. (2004). Lithologie und genetische Interpretation der vulkano-sedimentären Ablagerungen aus der Grube Messel anhand der Forschungsbohrung Messel 2001 und weiterer Bohrungen. Courier Forschungsinstitut Senckenberg, 252, 151–203.
Felder, M., Harms, F. J., & Liebig, V. (2001). Lithologische Beschreibung der Forschungsbohrungen Groβ-Zimmern, Prinz von Hessen und Offenthal sowie zweier Lagerstättenbohrungen bei Eppertshausen (Sprendlinger Horst, Eozän, Messel-Formation, Süd-Hessen). Geologisches Jahrbuch Hessen, 128, 29–82.
Gauch, H. G., & Scruggs, W. M. (1979). Variants of polar ordination. Vegetation, 40, 147–153. https://doi.org/10.1007/BF00228478.
Gruber, G., & Micklich, N. (2007). Messel - Treasures of the Eocene. Hessisches Landesmuseum Darmstadt.
Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontologia Electronica 4(1), https://palaeo-electronica.org/2001_1/past/issue1_01.htm.
Harms, F. J., Aderhold, G., Hoffmann, I., Nix, T., & Rosenberg, F. (1999). Erläuterungen zur Grube Messel bei Darmstadt, Südhessen. Schriftenreihe der Deutschen Geologischen Gesell-schaft, 8, 181–222.
Houk, V., Klee, R.., & Tanaka, H. (2017). Atlas of freshwater centric diatoms with a brief key and descriptions. Second emended edition of Part I and II. Melosiraceae, Liparogyraceae, Paraliaceae and Aulacoseiraceae. Fottea (Supplement), 17, 1–616.
Jacoby, W., Wallner, H., & Smilde, P. (2000). Tektonik und Vulkanismus entlang der Messeler-Störungszone auf dem Sprendlinger Horst: geophysikalische Ergebnisse. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 151(4), 493–510. https://doi.org/10.1127/zdgg/151/2001/493
Jankovská, V., & Komárek, J. (2000). Indicative value of Pediastrum and other coccal green algae in palaeoecology. Folia Geobotanica, 35, 59–82. https://doi.org/10.1007/BF02803087
Janssen, C. R., & Birks, H. J. B. (1994). Recurrent groups of pollen types in time. Review of Palaeobotany and Palynology, 82, 165–173. https://doi.org/10.1016/0034-6667(94)90028-0
Jardine, P. E., & Harrington, G. J. (2008). The Red Hills Mine palynoflora: A diverse swamp assemblage from the Late Paleocene of Mississippi, USA. Palynology, 32(1), 183–204. https://doi.org/10.2113/gspalynol.32.1.183
Juggins, S. (2007). C2 Software for ecological and palaeoecological data analysis and visualization. User guide Version 1.5. Newcastle upon Tyne: Newcastle University.
Kiss, K. T., Klee, R., Ector, L., & Ács, É. (2012). Centric diatoms of large rivers and tributaries in Hungary: Morphology and biogeographic distribution. Acta Botanica Croatica 1, 1-53. https://doi.org/10.2478/v10184-011-0067-0.
Krammer, K., & Lange-Bertalot, H. (1986). Bacillariophyceae. 1. Teil: Naviculaceae. In H. Ettl, J. Gerloff, H. Heynig, & D. Mollenhauer (Eds.), Süßwasserflora von Mitteleuropa, vol. 2/1, Jena: Gustav Fischer Verlag.
Krammer, K., & Lange-Bertalot, H. (1988). Bacillariophyceae. 2. Teil: Bacillariaceae. Epithemiaceae, Surirellaceae. In H. Ettl, J. Gerloff, H. Heynig, & D. Mollenhauer (Eds.), Süßwasserflora von Mitteleuropa, vol. 2/2. Stuttgart: Gustav Fischer Verlag.
Krammer, K., & Lange-Bertalot, H. (1991a). Bacillariophyceae. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. In H. Ettl, J. Gerloff, H. Heynig, & D. Mollenhauer (Eds.), Süßwasserflora von Mitteleuropa, vol 2/3, Stuttgart: Gustav Fischer Verlag.
Krammer, K., & Lange-Bertalot, H. (1991b). Bacillariophyceae. 4. Teil: Achnanthaceae. Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. In H. Ettl, J. Gerloff, H. Heynig, & D. Mollenhauer (Eds.), Süßwasserflora von Mitteleuropa, vol. 2/4 Stuttgart: Gustav Fischer Verlag.
Kruskal, J. B. (1964). Nonmetric multidimensional scaling: a numerical method. Psychometrika, 29, 115–129. https://doi.org/10.1007/BF02289694.
Krutzsch, W. (1966). Die sporenstratigraphische Gliederung des älteren Tertiär im nördlichen Mitteleuropa (Paläozän-Mitteloligozän), methodische Grundlagen und gegenwärtiger Stand der Untersuchungen. Abhandlungen des Zentralen Geologischen Instituts, 8, 112–149.
Krutzsch, W. (1970a). Die stratigraphisch verwertbaren Sporen- und Pollenformen des mitteleuropäischen Alttertiärs. – Jahrbuch für Geologie., 3, 309 - 379.
Krutzsch, W. (1970b). Einige neue Pollenformen aus den Familien der Tiliaceen, Bombacaceen, und Sterculiaceen aus dem mitteldeutschen Alttertiär. Jahrbuch für Geologie., 3, 275 - 307.
Lange-Bertalot, H., Hofmann, G., Werum, M., & Cantonati, M. (2017). Freshwater Benthic Diatoms of Central Europe. Over 800 common species used in ecological assessment. English edition with updated taxonomy and added species. Schmitten-Oberreifenberg: Koeltz Botanical Books.
Lenz, O. K. (2005). Palynologie und Paläoökologie eines Küstenmoores aus dem Mittleren Eozän Mitteleuropas Die Wulfersdorfer Flözgruppe aus dem Tagebau Helmstedt, Niedersachsen. Palaeontographica B, 271, 1–157.
Lenz, O. K., & Wilde, V. (2018). Changes in Eocene plant diversity and composition of vegetation: the lacustrine archive of Messel (Germany). Paleobiology, 44(4), 709735. https://doi.org/10.1017/pab.2018.25.
Lenz, O. K., Wilde, V., & Riegel, W. (2007a). Recolonization of a Middle Eocene volcanic site: quantitative palynology of the initial phase of the maar lake of Messel (Germany). Review of Palaeobotany and Palynology, 145(3), 217–242. https://doi.org/10.1016/j.revpalbo.2006.11.001.
Lenz, O. K., Wilde, V., Riegel, W., & Heinrichs, T. (2007b). Distribution and ecologic significance of a new freshwater dinoflagellate cyst (Messelodinium thielepfeifferae gen. et sp. nov) from the Middle Eocene Lake Messel (Germany). Palynology, 31, 119–134. https://doi.org/10.1080/01916122.2007.9989639.
Lenz, O. K., Wilde, V., & Riegel, W. (2011). Short-term fluctuations in vegetation and phytoplankton during the Middle Eocene greenhouse climate: a 640-kyr record from the Messel oil shale (Germany). International Journal of Earth Sciences, 100, 1851–1874. https://doi.org/10.1007/s00531-010-0609-z.
Lenz, O. K., Wilde, V., Mertz, D. F., & Riegel, W. (2015). New palynology-based astronomical and revised 40Ar/39Ar ages for the Eocene maar lake of Messel (Germany). International Journal of Earth Science, 104(3), 873–889. https://doi.org/10.1007/s00531-014-1126-2.
Lenz, O. K., Wilde, V., & Riegel, W. (2017). ENSO- and solar-driven sub-Milankovitch cyclicity in the Paleogene greenhouse world; high resolution pollen records from Eocene Lake Messel, Germany. Journal of the Geological Society 174, 110–128. https://doi.org/10.1144/jgs2016-046.
Lenz, O. K., Diekmeier, S., & Wilde, V. (2021). Nypa pollen diversity in middle Eocene mangroves at the edge of the southern Proto-North Sea (Helmstedt, northern Germany) – a statistical approach, Grana, 60(2), 97–118, https://doi.org/10.1080/00173134.2020.1808697.
Li, Y. M., Ferguson, D., Wang, Y. F., & Li, C. S. (2010). Paleoenvironmental inferences from diatom assemblages of the middle Miocene Shanwang Formation, Shandong, China. Journal of Paleolimnology, 43, 799–814. https://doi.org/10.1007/s10933-009-9369-9.
Lorenz, V. (2000). Formation of maar-diatreme-volcanoes. International Maar Conference. Terra Nostra, 2000/6, 284–291.
Lund, J. W. G. (1954). The seasonal cycle of the plankton diatom Melosira italica (Ehr.) Kütz. subsp. subarctica O. Müll. Journal of Ecology, 42, 151–179. https://doi.org/10.2307/2256984
Mai, D. H. (1981). Entwicklung und klimatische Differenzierung der Laubwaldflora Mitteleuropas im Tertiär. Flora, 171(6), 525–582. https://doi.org/10.1016/S0367-2530(17)31304-X.
Mai, D. H. (1995). Tertiäre Vegetationsgeschichte Europas—Methoden und Ergebnisse. Jena: Gustav Fischer Verlag.
Mander, L., Kürschner, W. M., & McElwain, J. C. (2010). An explanation for conflicting records of Triassic–Jurassic plant diversity. Proceedings of the National Academy of Sciences of the United States of America, 107, 15351–15356. https://doi.org/10.1073/pnas.1004207107-
Mezger, J. E., Felder, M., & Harms, F. J. (2013). Crystalline rocks in the maar deposits of Messel: key to understand the geometries of the Messel Fault Zone and diatreme and the post-eruptional development of the basin fill. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 164(4), 639–662. https://doi.org/10.1127/1860-1804/2013/0034.
Minchin, P. R. (1987). An evaluation of the relative robustness of techniques for ecological ordination. Vegetation, 69, 89–107. https://doi.org/10.1007/BF00038690.
Moshayedi, M., Lenz, O. K., Wilde, V., & Hinderer, M. (2018). Controls on sedimentation and vegetation in an Eocene pull-apart basin (Prinz von Hessen, Germany): evidence from palynology. Journal of the Geological Society, 175(5), 757–773. https://doi.org/10.1144/jgs2017-128.
Moshayedi, M., Lenz, O. K., Wilde, V., & Hinderer, M. (2020). The recolonisation of volcanically disturbed Eocene habitats of Central Europe: the maar lakes of Messel and Offenthal (SW Germany) compared. Palaeobiodiversity and Palaeoenvironments, 100(4), 951–973. https://doi.org/10.1007/s12549-020-00425-4.
Moshayedi, M., Lenz, O. K., Wilde, V., & Hinderer, M. (2021). Lake-level fluctuations and allochthonous lignite deposition in the Eocene pull-apart basin “Prinz von Hessen” (Hesse, Germany) - A palynological study. In M. R. Rosen, D. Finkelstein, L. Park Boush & S. Pla-Pueyo (Eds.), Limnogeology: Progress, challenges and apportunities: A tribute to Beth Gierlowski-Kordesch, (pp. 69–106). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-030-66576-0_3.
Nickel, B. (1996). Die mitteleozäne Mikroflora von Eckfeld bei Manderscheid/Eifel. Mainzer Naturwissenschaftliches Archiv. Beiheft, 18, 1–121.
Pflug, H. (1986). Palynostratigraphy of the Eocene/Oligocene in the area of Helmstedt, in North Hessia and in the adjacent Southern Range. In A. Tobien (Ed.), Nordwestdeutschland im Tertiär. Beiträge zur regionalen Geologie der Erde, 38, 567–583.
Oksanen, J. (2007). Standardization methods for community ecology. Documentation and user guide for package Vegan, 1.8-6.
Pirrung, M. (1998). Zur Entstehung isolierter alttertiärer Seesedimente in zentraleuropäischen Vulkanfeldern. Mainzer Naturwissenschaftliches Archiv Beiheft, 20, 1117.
Pirrung, M., Fischer, C., Büchel, G., Gaupp, R., Lutz, H., & Neuffer, F. O. (2003). Lithofacies succession of maar crater deposits in the Eifel area (Germany). Terra Nova, 15(2), 125132. https://doi.org/10.1046/j.1365-3121.2003.00473.x.
Potapova, M. G., Bixby, R. J., Charles, D. F., Edlund, M. B., Enache, M. E., Furey, P., Hamilton, P. B., Lowe, R. L., Manoylov, K. M., Ognjanova-Rumenova, N., Ponader, K. C., Ren, L., Siver, P. A., Spaulding, S. A., & Zalack, J. (2008). Representatives of the genus Aulacoseira Thwaites in NAWQA samples. Eighteenth NAWQA Workshop on Harmonization of Algal Taxonomy, April 27–29, 2007. Report 08–07 (pp.1-56). Philadelphia: Patrick Center for Environmental Research, The Academy of Natural Sciences.
Potapova, M., English, J., & Edlund, M. (2010). Aulacoseira italica. In Diatoms of the United States. Retrieved from https://diatoms.org/species/aulacoseira_italica, 03.07.2021.
Rampino, M. R., & Koeberl, C. (2006). Comparison of Bosumtwi Impact Crater (Ghana) and Crater Lake Volcanic Caldera (Oregon, USA): Implications for Biotic Recovery after Catastrophic Events. In C. Cockell, I. Gilmour, & C. Koeberl (Eds.), Biological Processes Associated with Impact Events. Impact Studies (pp. 100–120). Berlin, Heidelberg: Springer. https://doi.org/10.1007/3-540-25736-5_5.
Reinhold, C., Schwarz, M., Bruss, D., Heesbeen, B., Perner, M., & Suana, M. (2016). The northern Upper Rhine Graben - Re-dawn of a mature petroleum province? Swiss Bulletin für Angewandte Geologie, 21(2), 35–56. https://doi.org/10.5169/seals-658196.
Reuter, J. E. (1979). Seasonal distribution of phytoplankton biomass in a nearshore area of the central basin of Lake Erie, 1975-1976. The Ohio Journal of Science, 79(5), 218–226.
Rühland, K. M., Paterson, A. M., & Smol, J. P. (2015). Lake diatom responses to warming: reviewing the evidence. - Journal of Paleolimnology. 54, 1–35. https://doi.org/10.1007/s10933-015-9837-3.
Sabel, M., Bechtel, A., Püttmann, W., & Hoernes, S. (2005). Palaeoenvironment of the Eocene Eckfeld Maar lake (Germany): implication from geochemical analysis of the oil shale sequence. Organic Geochemistry, 36(6), 873891. https://doi.org/10.1016/j.orggeochem.2005.01.001.
Schaarschmidt, F. (1988). Der Wald, fossile Pflanzen als Zeugen eines warmen Klimas, In S. Schaal, & W. Ziegler (Eds.), Messel - ein Schaufenster in die Geschichte der Erde und des Lebens (pp. 27-52). Frankfurt am Main: Waldemar Kramer.
Schulz, R., Harms, F. J., & Felder, M. (2003). Die Forschungsbohrung Messel 2001: Ein Beitrag zur Entschlüsselung der Genese einer Ölschieferlagerstätte. Zeitschrift für Angewandte Geologie, 48(4), 9-17.
Schumacher, M. E. (2002). Upper Rhine Graben: Role of preexisting structures during rift evolution. Tectonics, 21(1), 6-1–6-17. https://doi.org/10.1029/2001TC900022.
Shepard, R. N. (1962a). Analysis of proximities: Multidimensional scaling with an unknown distance function. I. Psychometrica, 27, 125–140. https://doi.org/10.1007/BF02289630.
Shepard, R. N. (1962b). Analysis of proximities: Multidimensional scaling with an unknown distance function. II. Psychometrica, 27, 219–246. https://doi.org/10.1007/BF02289621.
Siver, P. A., Wolfe, A. P., & Edlund, M. B. (2010). Taxonomic description and evolutionary implications of Middle Eocene pinnate diatoms representing the extend genera Oxyneis, Actinella and Nupela (Bacillariophyceae). Plant Ecology and Evolution, 143, 340–351. https://doi.org/10.5091/plecevo.2010.419.
Siver, P. A., Velez, M., Cliveti, M. & Binda, P. (2018). Early freshwater diatoms from the Upper Cretaceous Battle Formation in Western Canada. Palaios, 33(12), 525-534. https://doi.org/10.2110/palo.2018.045.
Smith, K. T., Schaal, S. F. K., & Habersetzer, J. (2018). Messel – An Ancient Greenhouse Ecosystem. Senckenberg-Buch 80. Stuttgart: E. Schweizerbart’sche Verlagsbuchhandlung.
Suhr, P., Goth, K., Lorenz, V., & Suhr, S. (2006). Long lasting subsidence and deformation in and above maardiatreme volcanoes – a never ending story. Zeitschrift der Deutschen Gesellschaft für Geowissenschaften, 157(3), 491–511. https://doi.org/10.1127/1860-1804/2006/0157-0491.
Taylor, B. W., 1957. Plant succession on recent volcanoes in Papua. Journal of Ecology, 45, 233–243. https://doi.org/10.2307/2257087.
ter Braak, C. J. F. (1995). Ordination. In R. H. G Jongman, C. J. F Ter Braak, & O. F. R. Tongeren (Eds.), Data Analysis in Community and Landscape Ecology (pp. 91–274). Cambridge: Cambridge University Press.
ter Braak, C. J. F., & Looman, C. W. N. (1995). Regression. In R. H. G. Jongman, C. J. F. Ter Braak, & O. F. R. Tongeren, (Eds.), Data Analysis in Community and Landscape Ecology (pp. 29–77). Cambridge: Cambridge University Press.
Thiele-Pfeiffer, H. (1988). Die Mikroflora aus dem mitteleozänen Ölschiefer von Messel bei Darmstadt. Palaeontographica B, 211, 1–86.
Thomas, B. A., & Cleal, C. J. (2021) Pteridophytes as primary colonisers after catastrophic events through geological time and in recent history. Palaeobiodiversity and Palaeoenvironment, 102(1), 59–71. https://doi.org/10.1007/s12549-021-00492-1.
Thomson, P. W., & Pflug, H. (1953). Pollen und Sporen des mitteleuropäischen Tertiärs. Gesamtübersicht über die stratigraphisch und paläontologisch wichtigen Formen. Palaeontographica B, 94, 1–138.
Van Tongeren, O. F. (1995). Data analysis or simulation model: a critical evaluation of some methods. Ecological modelling, 78(1-2), 51–60. https://doi.org/10.1016/0304-3800(94)00117-Z
Weber, J & Hofmann, U. (1982). Kernbohrungen in der eozänen Fossillagerstätte Grube Messel bei Darmstadt. Geologische Abhandlungen Hessen, 83, 1-58.
Wetzel, R. G. (1983). Limnology. Philadelphia: Saunders College Publications.
Wilde, V. (1989). Untersuchungen zur Systematik der Blattreste aus dem Mitteleozän der Grube Messel bei Darmstadt (Hessen, Bundesrepublik Deutschland). Courier Forschungsinstitut Senckenberg, 115, 1–123.
Wilde, V., & Manchester, S. R. (2003). Cedrelospermum-fruits (Ulmaceae) and related leaves from the Middle Eocene of Messel (Hesse, Germany). Courier Forschungsinstitut Senckenberg, 241, 147–153.
Wilde, V., Frankenhäuser, F., & Lutz, H. (1993). Algenreste aus den mitteleozänen Sedimenten des Eckfelder Maares bei Manderscheid in der Eifel. Mainzer naturwissenschaftliches Archiv, 31, 127-148.
Wilde, V., Frankenhäuser, H., & Lenz, O. K. (2021). A myricaceous male inflorescence with pollen in situ from the middle Eocene of Europe. Palaeobiodiversity and Palaeoenvironments 101(4). https://doi.org/10.1007/s12549-020-00479-4.
Wilson, M. A., Burt, R., & Lee, C. W. (2006). Improved elemental recoveries in soils with heating boric acid following microwave total digestion. Communications in Soil Science and Plant Analysis, 37(3-4), 513524. https://doi.org/10.1080/00103620500449377.
Ziegler, P. A. (1994). Cenozoic rift system of Western and Central-Europe-an overview. Geologie en Mijnbouw, 73(2-4), 99–127.
Acknowledgements
The help of Karin Schmidt, Palaeobotanical Section, Senckenberg Research Institute and Natural History Museum Frankfurt, in sampling and sample preparation is gratefully acknowledged. We thank the Department for Messel Research and Mammalogy, Senckenberg Research Institute and Natural History Museum Frankfurt for supplying the core photographs of FIS/HLUG from the core B/97-BK 8. The authors would like to thank Mary Ellen Benson for helpful advice and literature. Reviews by Carlos Jaramillo and Manuel Casas-Gallego helped to finally improve the manuscript.
Funding
Open Access funding enabled and organised by Projekt DEAL. Maryam Moshayedi received funding through the Deutscher Akademischer Austauschdienst (DAAD) by grant number 110207-54900347.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mutzl, J., Lenz, O.K., Wilde, V. et al. Vegetation dynamics in a disturbed lacustrine record: The Eocene maar lake of Groß-Zimmern (Hesse, SW Germany). Palaeobio Palaeoenv 103, 249–276 (2023). https://doi.org/10.1007/s12549-022-00543-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00543-1