
Abstract
Reconstructions of palimpsest formation and dynamics in Early Pleistocene African archaeological deposits have undergone significant advances thanks to taphonomic research. However, the spatial imprint of different agents implicated in most of these accumulations still needs to be addressed. We hypothesize that different site formation dynamics may yield diverse spatial distributions of archaeological remains, reflecting the intervention of different agents (i.e., hominins, felids, hyaenids) in palimpsests. This study aims to investigate the spatial patterns of archaeological remains in a selected sample of Early Pleistocene accumulations with the goal of understanding and characterizing their spatial dynamics. Building on previous taphonomic interpretations of twelve paradigmatic archaeological deposits from Olduvai Bed I (FLK Zinj 22 A, PTK 22 A, DS 22B, FLK N 1–2 to 5, FLK NN 3, DK 1–3) and Koobi Fora (FxJj50, FxJj20 East and FxJj20 Main), we explore the spatial patterns of remains statistically and use hierarchical clustering on principal components analysis (HCPC) to group the highest-density spots at these sites based on a number of spatial variables. The results of this approach show that despite sharing a similar inhomogeneous pattern, anthropogenic sites and assemblages where carnivores played the main role display fundamentally different spatial features. Both types of spatial distributions also show statistical differences from modern hunter-gatherer campsites. Additional taphonomic particularities and differing formation processes of the analyzed accumulations also appear reflected in the classifications. This promising approach reveals crucial distinctions in spatial imprints related to site formation and agents’ behavior, prompting further exploration of advanced spatial statistical techniques for characterizing archaeological intra-site patterns.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The study of site formation and dynamics at significant Early Pleistocene sites like Olduvai Gorge Bed I (Tanzania) and Koobi Fora (Kenya) is critical for elucidating early hominin behavior. These sites provide essential information on the activities of hominins and their interactions and coexistence with carnivores, and have consequently been the subject of intense interest and research (e.g. Leakey 1971; Isaac 1983, 1984; Isaac 1997; Bunn 1981, 1982, 1986, 1991; Potts 1982, 1988; Bunn and Kroll 1986; Pobiner et al. 2008; Plummer and Bishop 2016; Plummer et al. 2009; Oliver et al. 2019; Parkinson 2018; Blumenschine 1995). Zooarchaeological and taphonomic studies have played a crucial role in understanding the site processes that led to the formation of these assemblages (Bunn 1982, 1991; Potts 1982, 1988; Bunn and Kroll 1986; Domínguez-Rodrigo et al. 2007a; Parkinson 2018). However, the spatial imprint left by various agents on these archaeological deposits remains underexplored. This paper seeks to address this gap by examining the spatial patterns of archaeological remains to contribute to discerning the roles and inputs of hominins, felids, and hyaenids within these sites.
The first investigations of these assemblages underestimated the role of carnivores and were interpreted as evidence of hominins’ repeated visits to these locations for animal carcass processing. This interpretation was primarily based on the spatial proximity of high concentrations of animal carcass remains and stone tools (Leakey 1971; Isaac 1981, 1983, 1984; Bunn 1982, 1991; Potts 1982, 1988; Bunn and Kroll 1986). However, subsequent taphonomic reanalyses revealed that most of these sites were more intricate palimpsests, wherein carnivores, including felids and hyaenids, had significantly contributed to the accumulation and modification of the assemblages (Domínguez-Rodrigo et al. 2007a). Among Olduvai Bed I sites (Fig. 1B-C), only FLK Zinj displayed dominant hominin activity (Domínguez-Rodrigo and Barba 2007a, 2007b; Parkinson 2018), and remained particularly noteworthy for being a very well-preserved, vertically discrete, and extensively excavated deposit (Table 1).
The discovery in 2012 and 2014 of the PTK and DS sites (Domínguez-Rodrigo et al. 2017a; Domínguez-Rodrigo and Cobo-Sánchez 2017b) on the same paleosurface as FLK Zinj (~ 1.84 Ma) (Deino 2012), separated by only a few hundred meters, presented a new opportunity to investigate early hominin behavioral variability at Olduvai Bed I. Recent taphonomic studies of these two new sites suggest that hominins were the primary accumulating and modifying agents at these locations, having early and primary access to small and medium-sized carcasses before other carnivores (Cobo-Sánchez 2020; Domínguez-Rodrigo et al. 2021; Organista et al. 2023). Furthermore, the involvement of felids in the formation of DS has been deemed very unlikely, with only hyenas altering some of the carcasses after hominin intervention. Nearly all activity, including carcass procurement, bone defleshing, and bone breakage targeting marrow is attributed to hominins (Cobo-Sánchez et al. 2022).
Several sites at Koobi Fora (Fig. 1A), such as FxJj50, FxJj20 East, and FxJj20 Main (Okote Member, ~ 1.6-1.4 Ma) (Brown and Feibel 1986; Brown and McDougall 2011), in which accumulations of bones and stone tools concur spatially, were also interpreted as hominin-made accumulations (Bunn et al. 1980; Bunn 1981, 1986; Pobiner et al. 2008; Plummer and Bishop 2016; Plummer et al. 2009; Oliver et al. 2019). Intra-site spatial analysis appears to confirm the integrity and agency of some of these sites (Kroll 1997; Bellomo 1994). At FxJj50, carnivore and rodent modifications were identified, as well as a few hominin modifications probably related to percussion and defleshing (Bunn 1997). Posterior revisions of the FxJj50 assemblage confirmed the primary access by hominins in that site (Brantingham 1998; Domínguez-Rodrigo 2002), although the intervention of carnivores as secondary access was also recognized. In contrast, at FxJj20 East and Main the poor cortical preservation made it difficult to identify surface modifications: only a few cut marks were found in FxJj20 Main, and there were no cut marks or tooth marks in FxJj20 East (Harris et al. 1997; Bunn 1997). In all sites, the presence of stone tools supports the idea of hominin involvement in the assemblages. However, further taphonomic analyses would be necessary to determine the main agent responsible for the faunal accumulation and draw additional inferences.

Map with the location of all sites mentioned in the text. A) Koobi Fora Okote Member sites, B-C) Olduvai Gorge Bed I sites
Regarding the non-anthropogenic sites from Bed I, FLK North levels 1–5 (~ 1.785 Ma) (Hay and Kyser 2001) hominin and carnivore activities seem to have been mostly independent. Analysis of tooth marks, types and frequencies of notch types, and breakage plane angles indicate that carnivores (mainly felids) were responsible for the bulk of the bone accumulations at these levels. Although stone tools are present, they appear to be unrelated to marrow extraction. The low abundance of flakes or cutting tools with edges also suggests that hominins were not generally engaging in carcass processing at the site. All levels are dominated by two bovid species, Parmularius altidens and Antidorcas recki, which suggests a specialized carcass collector, such as leopard and/or Dinofelis (Domínguez-Rodrigo and Barba 2007c; Domínguez-Rodrigo et al. 2007b; Egeland 2007a). Recently excavated trenches have revealed that only the high-density accumulation of levels 1–2 was clear. Level 3 was barely distinguishable, and levels 4 and 5 were difficult to differentiate (Domínguez-Rodrigo et al. 2010).
DK’s taphonomic results (~ 1.85 Ma) (Walter et al. 1992) indicate a slightly more complex formation history in which accumulations were made by both felids and hyenids. Some bones may have deposited independently as background scatters. In addition, a few cases of hominin-carnivore interdependence have been documented, although hominins were probably only using the site for carcass-processing activities intermittently (Egeland 2007c).
Due to the lack of anthropic evidence, the faunal assemblage from FLK NN (~ 1.84 Ma) (Deino 2012) has been attributed to the activity of carnivores. Most of the accumulations found in its three levels were the result of carnivore transport of carcasses to this preferential location in the landscape. The presence of complete bones and the low tooth mark frequencies support felids as the main agent (Barba and Domínguez-Rodrigo 2007; Domínguez-Rodrigo and Barba 2007d; Egeland 2007b). The virtual absence of tools, given that most of the so-called manuports are in fact ecofacts in many Bed I sites (de la Torre and Mora 2005), indicates that the degree of involvement of hominins in these accumulations was much lower than that reported on FLK N 1–5 and DK.
What have spatial studies contributed so far to Early Pleistocene archaeology? In general terms, they have complemented other analyses, focusing on the density of materials horizontally (activity zones) and vertically (temporal resolution) through visual inspection of excavation plans and using a few statistical indexes. The importance of ethnography in the early days of intra-site analysis has been fundamental in modeling the link between behavior, social organization, and spatial distribution of remains (Yellen 1977; Kroll and Price 1991; Bartram 1993). Initially, intra-site analysis aimed to obtain a precise resolution, yet few Pleistocene sites offer the ideal conditions for identifying spatially and temporally defined areas of activity. Skepticism about the interpretative potential of these techniques led to the partial abandonment of these methods (Clark and Gingerich 2022), but in recent years, spatial analysis and GIS have regained importance. These techniques are now crucial for studying the functionality of sites (Bargalló et al. 2015; Peters and van Kolfschoten 2020; Sánchez-Romero et al. 2020; Zilio et al. 2021; Saladié et al. 2021) and for spatial modeling (Cobo-Sánchez 2020), although the latter is generally applied in landscape archaeology (Mehrer and Wescott 2006; Castielo 2022; Kempf and Günther 2023).
The development in the last two decades of spatial statistics and the creation of libraries such as “spatstat” (Baddeley 2008; Bivand et al. 2013; Diggle 2014; Baddeley et al. 2016) has boosted the application of new methodologies that incorporate spatial point process theory. Although these techniques are widespread in other disciplines, their arrival in archaeology has been relatively recent. Spatial statistical tools have now made it possible to enhance the reliability of interpretations that previously depended solely on the visual inspection of the researcher, lacking solid evidence of statistical significance. Among other applications, they have proved their potential in discerning the social organization of both hunter-gatherer groups and early Homo (Domínguez-Rodrigo and Cobo-Sánchez 2017a, b), predicting site material density in areas not yet excavated (Domínguez-Rodrigo et al. 2017a), studying post-depositional processes (Domínguez-Rodrigo et al. 2017b), and particularly in interpreting site formation, especially when combined with taphonomy (Panera et al. 2019; Cobo-Sánchez 2020; Méndez-Quintas et al. 2022; Moclán et al. 2023a, b).
Building on this body of research, the present study proceeds under the assumption that different site formation dynamics may yield diverse spatial distributions of archaeological remains. Anthropogenic and non-anthropogenic assemblages may exhibit fundamental differences in their spatial patterns, as they were formed by distinct taphonomic agents (i.e., hominins, felids, or hyenids). Each of these agents potentially exhibits unique features in terms of space utilization, transport, processing, and consumption of carcasses. Our understanding of the spatial imprint of early hominin-made accumulations largely stems from various spatial studies conducted on PTK, DS, and FLK Zinj, which seems to indicate that Early Pleistocene hominins displayed spatial behaviors that resulted in well-defined mono-cluster patterns (Domínguez-Rodrigo and Cobo-Sánchez 2017a, b; Cobo-Sánchez 2020; Díez-Martín et al. 2021). Additionally, this study hypothesizes that carnivores will likely not exhibit as dense or well-defined accumulation patterns, given that actualistic studies have shown that carnivore ravaging can result in the partial dispersion of remains (Binford 1981; Binford et al. 1988; Marean and Bertino 1994; Camarós et al. 2013; Arilla et al. 2020). The main differences could lie in the intensity of the archaeological patterns, the number of hot-spots, the clustering patterning, and the correlation of remains.
Hypothetically, such disparities would be spatially reflected in cases of minimal post-depositional disturbance and without alteration to the original spatial configuration. The spatial composition of independent and interdependent site occupations might also display unique characteristics. An interdependent accumulation would arise from the original spatial configuration being modified or dispersed by a second agent, so it could therefore still preserve some aspects of the original spatial pattern. On the contrary, events with no interaction may exhibit a combination of two distinct spatial patterns.
Here, we aim to address several objectives. We first seek to determine if these sites display different types of spatial point patterns in terms of intensity and correlation. Specifically, we expect those differences to be more pronounced among sites where a single predominant agent contributes significantly to the accumulation process (Table 1). Secondly, we test a new approach that allows us to make quantitative comparisons between sites. This approach is based on measuring a set of spatial features of the main high-density areas of each site. Finally, we investigate whether these patterns reflect distinct taphonomic agents or processes based on the results and interpretations from previous studies.
Materials and methods
To this aim, we conducted a spatial statistical analysis of the distribution of archaeological remains and used hierarchical clustering on principal components analysis (HCPC) to group the highest-density areas at a number of Early Pleistocene sites based on several spatial variables.
Sample of selected sites
We selected the sites based on specific spatial and taphonomic criteria, ensuring their well-studied taphonomic history, preserved general spatial configuration, minimal post-depositional impact by abiotic processes, and sufficiently large spatial windows for proper analysis. The chosen sites for analysis included several Olduvai Bed I sites (FLK N 1–2 to 5, FLK NN 3, DK 1–3, PTK, FLK Zinj 22 A, PTK 22 A, and DS 22B) and three Koobi Fora sites (FxJj50, FxJj20 Main, and FxJj20 East) (Table 2), which represent a variety of agents and palimpsests responsible for the accumulations commonly found in Early Pleistocene archaeological assemblages (see Table 1 column Interpretation).
The archaeological assemblages of the sample analyzed in this study present a varied degree of time averaging, which challenges the interpretation of spatial patterns. As noted by Díez-Martín et al. (2010) and Domínguez-Rodrigo et al. (2010), the presence of a refuse continuum across the vertical stratigraphy at FLK N suggests a prolonged accumulation of materials over time. This complicates the assessment of discard activity intensity, which may result from both short-term episodes or continuous activity by hominin groups. In contrast, taphonomic analyses have shown that DS, PTK, and FLK Zinj assemblages were formed within a short period of a few years (estimated at 2–3 years in DS), according to the documented subaerial weathering stages (Domínguez-Rodrigo et al. 2007a; Cobo-Sánchez 2020; Organista et al. 2023). FxJj50 vertical distribution has been subjected to different interpretations. While some authors argued that refittings proved a unique depositional event (Bunn et al. 1980; Isaac 1997), posterior revisions by Kroll (1997) led her to interpret both FxJj50 and FxJj20 E as multiple occupations on various old surfaces, sometimes already buried between hominin visits, but with little spatial disturbance. Understanding the implications of time averaging is crucial for interpreting spatial patterns in archaeological assemblages. However, there is a scarcity of spatial studies addressing its effects comprehensively. One can hypothesize that if hominins visited the same spot repeatedly, it would leave a stronger trace over time. For instance, sporadic visits might leave a weaker imprint (e.g. FLK N 3–5), while more frequent visits could leave a stronger one (FLK N 1–2). However, this idea has not been tested appropriately from a spatial perspective, and there are high chances that natural post-depositional processes after and/or in between depositions could remove some of the evidence. The particularities in the formation of each assemblage explain partly why most spatial studies focussing on disentangling time-palimpsests are site-focused.
Regardless of the time span represented by each site, we presume that repeated occupations (numerous or sparse) have created distinct patterns based on the dominant agent. This assertion finds support in spatial statistics and point process theory. According to it, the superimposition of two similar point patterns, such as inhomogeneous Poisson processes (characterized by spatial inhomogeneity and randomness), results in a new point pattern that still follows an inhomogeneous Poisson distribution (Baddeley et al. 2016). Thus, the original pattern is preserved. Although numerous reoccupations may increase the overall intensity, other spatial traits might be preserved. It is noteworthy that not all spatial statistical methods focus on intensity estimation; rather, our analyses include a diverse array of spatial characteristics (i.e., correlation, spacing).
All analyses, when possible, were performed in the overall (all remains unmarked without distinction), bones, and lithics patterns. Using broad categories such as bones and lithics might carry some drawbacks too, as they cannot fully capture specific aspects of the sites’ patterns (i.e., spatial localized formation processes and agencies). The limitations stem partly from working with historical data, where the spatial component of the attributes of the remains is not usually available. The plans provided by Leakey (1971) indicate some artifact types and anatomical parts, which can facilitate more detailed spatial analyses in the future. However, such specific information is missing in the Koobi Fora plans, which limits the inclusion of these categories in our analyses. Furthermore, the absence of spatially recorded information regarding the size of remains and taphonomic marks, presents additional challenges, since these details could provide crucial insights into site formation processes. This information is only available in the recently excavated sites DS and PTK and has been used for spatial analyses elsewhere (Cobo-Sánchez 2020; Díez-Martín et al. 2021).
Digitalization of excavation plans
Since most sites were excavated during the second half of the 20th century, we had to digitalize the published maps and calculate the relative coordinates of all the remains in order to work with the spatial properties of the excavation planimetries. The excavation plans were obtained from the original excavation reports (Leakey 1971; Isaac 1997) and digitalized using ArcGIS (10.4.1 version). They were projected onto a Universal Transverse Mercator (UTM) projection. Subsequently, the digital plans were scaled, thus units corresponded to meters in reality. Coordinates corresponding to the remains were then extracted. The resulting shapefiles were read in R (R Core Team 2023) with the libraries “rgdal” and “maptools” (Bivand et al. 2023a, b). The DS and PTK spatial data stem directly from recent excavations and were recorded using total stations, so no previous data preparation was required.
In this sense, contemporary excavations offer more reliance on their data due to meticulous recording, advanced mapping technologies, and interdisciplinary approaches. DS and PTK went through a thorough excavation process, employing laser theodolites to map materials over 2 cm in size and stereo-photography for 3D reconstructions of unearthed trenches. Sediment was meticulously sifted through 5 mm and 3 mm meshes after being removed, while item orientations were measured using compasses and inclinometers (Cobo-Sánchez 2020; Organista et al. 2023). Unfortunately, however, we lack detailed information on the fieldwork process in most of the sites excavated during the 60–70s. During the excavation of the primary accumulation in FLK Zinj, it is acknowledged that not all remains’ locations were documented (Leakey 1971). However, lithic pieces smaller than 5 mm were recovered from FLK Zinj assemblage, indicating a careful collection of the material. For example, in FxJj20 M lithic remains sized less than 3 cm were recovered (Bellomo 1994), but the minimum size of plotted materials is unknown. The research team recognized in the reports that some bias generated by the excavation methodology may have led to the exclusion of smaller elements (Harris and Isaac 1997: 160). Regardless of data collection, a partial loss of information must be assumed before proceeding to analyze and interpret patterns. Notice the discrepancies between the reported number of remains and the actual number of plotted bones and lithics in the planimetries in the older sites (Table 2). In some cases, the plotted sample represents less than 10% of the total reported remains (see Table 2, DK and Koobi Fora sites percentage of analyzed remains). It is likely that only identifiable remains were plotted and used for spatial analysis at Koobi Fora (Kroll 1997), and similarly at Olduvai Bed I Leakey’s plans (1971). We do not expect this to significantly affect results since the general described spatial pattern is still recognizable. Previous spatial studies (Kroll 1997) and observations noted on reports (Leakey 1971) pointed towards the existence of certain clusters that we were able to confirm statistically (e.g. the two main clusters Mary Leakey noticed in FLK N 1–2 also appear when digitalizing the maps). However, the underrepresentation of remains at some historically excavated sites can directly affect intensity estimation. Therefore, methods based on intensity were only used to obtain graphic displays of the distribution of the entire assemblage and the delimitation of high-density areas. In addition to this, the focus of this paper is to understand the relationships and patterns that emerge from the distribution of points or remains across space.
Spatial statistical exploratory analysis: intensity and correlation
The first set of spatial methods aimed to understand the general spatial configuration of archaeological sites in order to characterize them and examine potential differences across sites. We applied several spatial statistical non-parametric tests and summary functions to the palimpsests’ point patterns. The variation of intensity of remains across the spatial window of a site, known as spatial inhomogeneity, is of particular interest. It is crucial to determine whether the inhomogeneity and the clustering tendency observed visually in the plans are supported statistically. Confirming inhomogeneity is necessary to then apply the corresponding tests and not to mix variability in intensity with clustering. This is essential since results indicating randomness in the distribution of remains may suggest the absence of agent-specific or activity-specific spatial patterns.
Consequently, we focused on two different objectives: estimating point pattern intensity (number of artifacts per square meter) to characterize inhomogeneity, and measuring point correlation (i.e., overall point interdependence, and types of association between bones and lithics). It is expected that potential differences might arise between anthropogenic and non-anthropogenic accumulations in terms of intensity and correlation. For the first objective, we tested the null hypothesis of Complete Spatial Randomness (CSR), created density maps for each site using kernel estimation of intensity, and examined the relative abundance of bones and lithics. The second objective was addressed by calculating nearest-neighbor distance functions and their mean, and examining point type associations using the nearest-neighbor correlation, the nearest-neighbor equality function, and the mark connection function. These are the most commonly used methods for statistical spatial exploration analysis. These techniques can be interpreted jointly and are complementary. The following section details each approach. Table 3 additionally summarizes the described methods. All spatial statistical analyses were performed in R using the “spatstat”, “sparr”, “sp”, and “maptools” packages (Baddeley et al. 2016; Davies et al. 2017; Pebesma and Bivand 2005; Bivand et al. 2013, 2023b).
Inhomogeneity and spatially varying intensity estimation
Since intensity variation appeared to be very marked throughout the sites’ excavated areas, the Hopkins-Skellam index was applied to assess inhomogeneity. This index results from comparing nearest-neighbor distances from aleatory samples of points with empty-space distances from random spatial locations (Hopkins and Skellam 1954). It was performed by applying the Cumulative Distribution Function method for edge correction and indicating “clustered” as the alternative hypothesis. Values greater than 1 indicate that points tend to avoid locations close to other points (regular pattern). If distances are smaller than expected, it is possible that the pattern is agglomerative and might be forming clusters. The Hopkins-Skellam index is less sensitive to inhomogeneity or edge effects. Given the marked intensity variation observed at the sites, this test provides a more reliable measure of clustering than other similar indexes such as Clark-Evans (1954), which assumes stationarity.
Kernel estimation of intensity was used to obtain density maps of the spatially varying intensity. We selected the smoothing bandwidth (σ) using the likelihood cross-validation method, which assumes an inhomogeneous Poisson process. The Diggle edge correction method was also applied to avoid possible biases caused by the window’s boundary (Baddeley et al. 2016; Diggle 1985). Three maps were created per site to display the intensity of bones, lithics, and the overall pattern. These maps were later used as the basis for the calculation of hot spots through scan tests.
For a better understanding of the point types’ (lithics and bones) relative spatial distribution, we used the spatially varying relative risk estimation (Kelsall and Diggle 1995). This nonparametric method estimates the probability of spatial distribution of each mark in a point pattern. The null hypothesis suggests that both bones and lithics have similar probabilities of appearing across the entire spatial area. Likelihood cross-validation was used again to select an appropriate bandwidth. Tolerance contours were drawn around the regions where the estimated probability of a given type was significantly different from the average proportion (Hazelton and Davies 2009; Davies et al. 2017). Significance was assessed using a Monte Carlo test. First, we relabel randomly the point pattern (i.e., giving the label bone or lithic randomly to all remains) and compute the type probability or relative risk. This is repeated n times, yielding n images of the probabilities of the mark types, and a Monte Carlo test is computed at each pixel of those images. We carried out 19 simulations, which produced values that are multiples of 1/20 = 0.05 (Davies et al. 2017; Hazelton and Davies 2009; for a similar archaeological application of this method see Cobo-Sánchez 2020 and Diez-Martín et al. 2021).
The K-function and other summary functions can help characterize the spatial correlation and dependence between remains (clustered, regular, random) by analyzing their distribution inside a radius r (Baddeley et al. 2000, 2016). However, they can be challenging to interpret or ambiguous because their values contain contributions for all interpoint distances less than or equal to r. It has been observed that using K or G inhomogeneous functions to assess correlation in patterns with low agglomerative tendency, such as those commonly found in Early Pleistocene assemblages, may not always yield successful results (Cobo-Sánchez 2020; Díez-Martín et al. 2021, 2023; Moclán et al. 2023b).
An alternative is the pair-correlation function, which is a standardized derivative K-function (Baddeley et al. 2016). The inhomogeneous pair correlation function g(r) compares the probability of observing a pair of points separated by a distance r with the probability expected for an inhomogeneous Poisson process (an inhomogeneous random pattern) when g(r) = 1. The g(r) values lower than 1 indicate regularity, while higher results are obtained for agglomerative patterns. We applied it to bones, lithics, and overall sites’ patterns using the divide-by-d estimator, which performs better for small r values. To test whether the deviation from the inhomogeneous Poisson line was statistically significant we created envelopes with 39 Monte Carlo simulations of inhomogeneous Poisson processes using the intensity function of the sites, giving the test a significance level of 2/40 = 0.05. This guarantees that the archaeological and the inhomogeneous Poisson curves are comparable and that the corresponding envelopes support a valid Monte Carlo test of the null hypothesis (Baddeley et al. 2016).
Overall interdependence and association types among remains
Once the intensity and inhomogeneity were estimated in all sites, we focused on methods that addressed correlation. Especially relevant to understanding palimpsest formation is the issue of whether bones and lithics from a particular deposit are functionally linked. The spatial co-occurrence in the same stratigraphic layer is not proof of association, which was in fact the assumption that led to the interpretation of many Early Pleistocene accumulations as hominin made. A statistical assessment of the type of spatial association between types of remains can be useful to understand whether there is a functional link between them. If bones and lithics are randomly mixed, the probability of a functional association would be low. In contrast, positive or negative associations are more likely to represent a functional association.
Since spatial variation in most palimpsests is extreme and hard to estimate accurately, it is a challenge to determine the type of correlation between bones and lithics using methods like the inhomogeneous cross-type K-function or the i to any Kdot function. Approaches based on nearest-neighbor distances should be more robust against spatial variation because they do not involve estimating the intensity of the point pattern. Methods like the nearest neighbor correlation, nearest neighbor equality function, and the mark connection function are useful to assess spatial clustering and more robust against spatial variation (Baddeley, pers. comm.). We thus explored interdependence between lithics and bones at each site additionally by means of these three approaches.
The nearest-neighbor correlation is a robust method that can be applied to stationary and non-stationary processes. This test analyzes the probability of the same types of points lying close to each other. Unnormalized correlation gives the probability that neighboring fossils are of the same type (lithic or bone). Normalized correlation compares the result with what is expected under random labeling of the point pattern (Baddeley et al. 2016). A value of 1 indicates consistency with random labeling, smaller values imply that neighbors are of different types, while larger values suggest that neighbors are of similar types.
An extension of the nearest-neighbor correlation is the nearest-neighbor equality function which counts the proportion of neighbors of a certain type against the order of the neighbor. The graph of the cumulative proportion shows the fraction amongst the kth nearest neighbors, which have a specified type. The envelopes to assess significance are effectively generated by keeping the positions of the points fixed and shuffling only the labels of the points (Cobo-Sánchez 2020; Diez-Martín et al. 2021).
Finally, the mark connection function calculates the ratio of the bivariate pair correlation (gij) and the unmarked pair correlation (g) and shows the resulting conditional probability that two points lying together are of the same or different types (Baddeley et al. 2016). The output is an array of graphics displaying the association between bones, lithics, bones-lithics, and lithics-bones. Results lying above and below the benchmark correspond to positive or negative correlation, respectively.
Cluster analysis on the spatial features of the highest intensity areas
Given that most non-parametric approaches discussed in the previous section do not yield quantifiable results in the form of single measures and that the estimates and p-values of some statistical tests were too similar to detect differences, we included a further approach that consists of comparing the sites based on quantifiable spatial measurements (Table 4). This is crucial as spatial comparisons among sites are not usual in the scientific literature due to a lack of approaches. While the methods presented above allow us to make comparisons, they are mostly subjective. Developing alternatives that address quantifiable spatial differences can provide robust support to our inferences regarding the previous spatial tests and analyses.
The considered spatial parameters include absolute and relative variables related to the major statistically significant high-density areas of each spatial pattern (e.g., perimeter, diameter, area, mean intensity, mean distance between the points inside the hot spots), but also variables related to the relation between the dense areas and the remaining spatial window, which ensure that features of the scattering and peripheral areas are considered as well. As high-density areas exist on all sites, their presence is not in itself a criterion for differentiating between palimpsest types and therefore they need to be looked at more closely. Ideally, other small accumulations or scatter areas should be analyzed separately. However, we believe that approach is useful only when information on the formation and agency of each of these areas is available. Given the lack of spatially localized taphonomic variables in almost all assemblages, only the largest areas were taken as reference, as we can think of them as a proxy for the focus of activity occurred. The exception is DS, whose in-depth spatial taphonomic study has resulted in a comprehensive interpretation of each of its accumulations (Cobo-Sánchez 2020; Díez-Martín et al. 2021), allowing them to be treated separately as three distinct areas similar in size (zones A, B, and C).
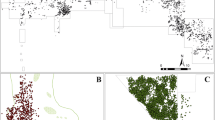
To calculate the parameters in each site, the statistically significant high-density areas were first isolated by performing a spatial scan test (Kulldorff 1997). The scan test detects statistically significant differences of intensity inside circles of fixed radius all around the pattern. It uses a likelihood ratio test statistic to assess whether intensities inside and outside a certain fixed radius are similar or not (Kulldorff 1997; Baddeley et al. 2016). Sigma values were estimated using the likelihood cross-validation method, sometimes multiplied by a value ranging from 0.5 to 2.5 to accurately estimate the radius in which a hot spot occurred (sigma values specified in Fig. 2 and SI Figs. I-II). We then computed the p-values for all locations. The largest elevated-intensity areas in each site were considered the focus of activity and the main object of analysis (Fig. 2). However, we have also taken into account smaller areas where secondary accumulations occurred (see variables 11, 16, and 21 in Table 4). The main activity areas were treated as new separate point patterns. Then, the parameters were estimated using functions from the “spatstat” library in R (Baddeley et al. 2016). Calculations were made for the overall point patterns, as well as separately for the bones and the lithics’ spatial patterns. In addition to all sites, four modern hunter-gatherer Kua (Bartram et al. 1991) and! Kung (Yellen 1977) campsites were included in the comparison of spatial patterns of bones, since stone tools are absent in these camps. Comparisons with the spatial distribution of debris at modern hunter-gatherer campsites have also demonstrated that contemporary human home bases lack these mono-cluster patterns. This suggests that early hominins utilized space in fundamentally different ways compared to modern humans today (Domínguez-Rodrigo et al. 2017Domínguez-Rodrigo and Cobo-Sánchez 2017b; Cobo-Sánchez 2020).

Maps of the overall spatial distribution of remains (bones and lithics) of the selected sites. The blue-shaded areas identify statistically significant high-density areas estimated with hot spot scan tests. All sigma values were multiplied by two except for FLK NN (2.5) and FxJj20 complex (2.5). The top two rows depict anthropogenic accumulations, whereas the bottom two rows primarily represent carnivore and palimpsests (FLK N 1–2, DK) accumulations. For the bones and lithics spatial distribution see Supplementary Information (SI) Fig. I-II. Bones distribution and statistically significant high-density areas of hunter-gatherer campsites can be found in SI Fig. III
With the aim of grouping similar high-density areas, we then performed hierarchical clustering on principal components (HCPC) analysis on the resulting dataset of spatial variables using the “FactoMineR” package in R (Lê et al. 2008; Husson et al. 2007). PCA is used as a preprocessing “denoising” method prior to performing the clustering analysis to better highlight the main features of the data set. Additionally, it reduces the dimensionality of the dataset, which has 28 variables with potential correlations. The analysis was carried out (a) for the overall spatial point patterns, (b) for the bones’ spatial patterns, (c) for the bones’ spatial patterns including the hunter-gatherer campsites, and (d) for the lithics’ spatial patterns.
Results
The varying intensity within assemblages
Spatial inhomogeneity is supported for all sites by the results of the Hopkins-Skellam index, suggesting that the hypothesis of CSR should be rejected. The obtained A values were statistically significant in all cases (p-value < 2.2e-16) and extremely low. This is compatible with agglomerative patterns in which points tend to lie closer to each other than would be expected for random patterns (Table 5). Yet, a clustering tendency does not necessarily entail clusters. Kernel density maps illustrate the varying intensity of the spatial distribution of remains (Fig. 3). Most sites seem to display a few main high-density areas surrounded by several smaller ones, however FLK Zinj, PTK, and DK overall patterns appear to be composed of one main accumulation. FLK NN, FLK N 5, and FxJj20 M show several small statistically significant high-density accumulations, based on the results of hot spot scan tests (Fig. 2). Hot spot intensity is higher in FLK Zinj, PTK, DS, and FLK N 1–2 than at the Koobi Fora sites. FLK N levels 3–5, DK, and FLK NN show significantly lower hot spot intensities. The limited contribution of hominins to sites such as FLK NN, and FLK N levels 3–5 is reflected in the low intensity of lithic remains, which also decreases the overall lower intensity. At these sites, bone remains therefore determine the overall spatial distribution (SI Fig. IV-V). However, it is noteworthy to mention that this observation varies notably for DK. When considering all remains at DK, including those not analyzed in this study, a significant increase in mean intensity is observed, rising from 0.47 to 5.12 for lithics and from 1.60 to 42.85 in bones (see Table 2).

Kernel intensity estimation for overall patterns. Bar scales = p/kds (estimated pieces per kernel density surface). The top two rows depict anthropogenic accumulations, whereas the bottom two rows primarily represent carnivore and palimpsests (FLK N 1–2, DK) accumulations
Relative risk maps suggest strong segregation between bones and lithics spatial distribution in all sites (Figs. 4a and 4b). However, in general, the probability of finding bones in the same areas as lithics is relatively higher than the other way around. This implies that in most sites lithics are more concentrated in specific locations, whereas bones are spread more widely across the window. Koobi Fora stands as an exception: the results show that lithics are more likely to be found across the entire spatial window, and bones as agglomerated events. It is worth mentioning FLK NN in this regard, where the concentrations of bones and lithics occur on opposite sides of the spatial window. This could indicate independent depositional events. Although the mean distance to nearest-neighbors does not vary substantially between sites, we noticed that FLK Zinj, DS, PTK, and FLK N 1–2 tend to display the lowest mean distances, also when considering different types of remains (Table 5).

Relative risk maps with Tolerance contours highlighting the areas where the probability of bones is significantly higher than the average proportion (p-value < 0.05). The top two rows depict anthropogenic accumulations, whereas the bottom two rows primarily represent carnivore and palimpsests (FLK N 1–2, DK) accumulations

Relative risk maps with tolerance contours highlighting the areas where the probability of lithics is significantly higher than the average proportion (p-value < 0.05). The top two rows depict anthropogenic accumulations, whereas the bottom two rows primarily represent carnivore and palimpsests (FLK N 1–2, DK) accumulations
variable at sites that have higher carnivore input, i.e., FLK N 3–5, FLK NN, and DK. This might suggest that bones and lithics display two very distinctive spatial patterns at these sites, as the relative risk maps suggest.
The inhomogeneous pair correlation functions of overall patterns in most sites follow similar tendencies: distances between remains are shorter than expected for a random pattern, but when the radius of analysis increases (around r = 1 m) the patterns turn more regular, and distances are greater than for an inhomogeneous distribution (Fig. 5).

Inhomogeneous pair correlation function of the overall patterns. The black line is the observed function, and the horizontal red dashed line is the theoretical estimated inhomogeneous Poisson pattern. Shaded areas represent envelopes generated with 39 Monte Carlo simulations of the inhomogeneous Poisson process with the intensity of each site. The top two rows depict anthropogenic accumulations, whereas the bottom two rows primarily represent carnivore and palimpsests (FLK N 1–2, DK) accumulations
The main distinction between patterns lies in the possibility of distinguishing inhomogeneous Poisson patterns from agglomerative patterns with potential clusters. We observe three general tendencies among sites. In sites like FLK Zinj, PTK, DS, FLK N 1–2, and 5, statistically significant results were found, suggesting that remains in high-density areas are positively associated and closely located, often within distances < 30 cm. This implies that remains may be spatially organized in distinct and well-defined possible clusters, with significant gaps between them. For FLK N 3, FLK NN, and Koobi Fora sites, the pair correlation function supports regularity, although not at all distances. The high amount of remains at short distances might be the result of an inhomogeneous Poisson pattern. Finally, for DK and FLK N level 4, the inhomogeneous distribution of remains cannot be ruled out, except for certain short intervals of distances.
The results for the lithic patterns (SI Fig. VII) in the carnivore sites indicate that the spatial event of lithics occurring is inhomogeneous and aleatory, so lithics do not present any positive or negative spatial correlation. Once more, FLKN levels 1–2 are the exception among sites with strong signals of carnivore activity, probably due to the high concentration of lithics in two large areas. However, unlike most anthropogenic accumulations, the function indicates a more regular to inhomogeneous tendency. As for bone distributions, results are similar to the overall patterns with the exception of FLK NN and the FxJj20 complex at Koobi Fora, which present a clustering tendency (SI Fig. VI).
The interdependence of bones and stone tools
Normalized nearest-neighbor correlation values of all sites were slightly greater than 1, apart from FxJj20 Main (Table 5). Notice that 1 is consistent with random labeling, and therefore independence between types. Unnormalized results suggest that the probability of remains being the same type (lithic or bone) is high. Once again, the high probability estimated for FLK NN (0.97) indicates that lithics and bones are strongly segregated.
The nearest neighbor equality function results (Fig. 6) align with this observation: in all sites bones and lithics appear to be spatially segregated, and graphics suggest a positive correlation between remains of the same type. Similar interpretations were previously drawn from relative risk maps and the Hopkins-Skellam test. However, results were non-significant in FLK N levels 3, 4, and 5, suggesting random labeling and no association between bones and lithics in these sites.
The mark connection function addresses bone-bone, lithic-lithic, and bone-lithic correlations (SI Fig. VIII-IX). Lithics and bones in all sites tend to exhibit certain types of correlation, which can be positive (clustering) or negative. However, in FxJj20 M and FLKN 1–2, 3, and 5 levels, it oscillates around the reference line of random labeling. This hints that these types might not be fully correlated in the entire assemblage. In most anthropic accumulations, bones typically show a negative correlation (with a brief positive correlation at close distances), while lithics exhibit a positive correlation. However, DS displays inverse correlation patterns, with bones showing a positive correlation and lithics displaying a positive-to-negative trend. Among carnivore and palimpsest accumulations, correlation patterns are similar between FLK N levels 1–2 and 3, as well as between FLK NN and DK (see details in SI Fig. VIII-IX). Mark connection function results align partly with the nearest neighbor equality function, but due to the lack of statistical significance reliance should be on the second method.

Nearest neighbor equality function. The black line displays the proportion of neighbors of a certain type. Values above the red dashed line indicate that the neighbors tend to be of the same type, while values below it describe patterns where points of different types are correlated or the same types inhibit. Shaded-area represent confidence envelopes for aleatory marks spatial distribution. The top two rows depict anthropogenic accumulations, whereas the bottom two rows primarily represent carnivore and palimpsests (FLK N 1–2, DK) accumulations
Classification of highest-intensity areas of overall, bones and stone tool patterns
The previous spatial statistical analysis provides insights into possible spatial differences among sites. Given the complexity of interpreting all those results, this second unsupervised method allows to jointly analyze several features and find hidden patterns regarding the statistical relationships between the sites. The results presented in the following paragraphs enable us to categorize the sites according to the spatial characteristics of their highest-density areas. Below we use the term cluster to refer to the groups identified in the HCPC analysis, leaving the term high-density area to define the largest hot-spots analyzed on each pattern. The spatial values used in the classification analyses can be found in SI Tables I-III. Additionally, the quantitative variables describing each cluster the most and the correlation between variables and dimensions are provided in SI Tables IV-XI.
Overall spatial patterns
The hierarchical clustering on principal components analysis (HCPC) on the archaeological assemblages explains around 50–60% of the variance of the dataset and classifies the high-density areas of the overall patterns into four different groups (Fig. 7a). The first group includes FLK N levels 3, 4, and 5, and FLK NN. In the dendrogram map, the height axis indicating distances between clusters suggests that differences are more marked between this group and the rest (SI Fig. Xa). All DS areas (A, B, and C), PTK, and FLK Zinj form another group, while on the other side of the factor map, the Koobi Fora sites are similar to FLK N 1–2. DK appears as an outlier event in the clustering diagram.
This distribution in the maps clearly separates anthropic from carnivore accumulations, with an intermediate group of sites formed by palimpsests and Koobi Fora sites. Dimension 1 explains the size and quantity of high-density areas. Anthropogenic accumulations typically exhibit larger main high-density areas with greater overlap between bones and lithics areas, particularly in PTK and DS areas C and B. This tendency is also observed among FLK N 1–2 and DK palimpsests. Interestingly, FLK N 1–2 and Koobi Fora sites share a similar ratio of bone intensity to lithic intensity, which is smaller than the overall sample mean. This implies that the intensity of both patterns is proportional. Carnivore accumulations seem to display opposite spatial features: distance between neighbors inside high-density areas and all remains to the centroid are larger. They also seem to count with a greater number of high-density areas, whose sizes are similar.
Dimension 2 emphasizes differences in the shape and intensity of main high-density areas. The intensity is higher and the distance of remains to the estimated boundary of the high-density area is greater among DS areas A and B, PTK and FLK Zinj. Perimeter to diameter ratio indicates that these sites have a more compact shape. Similar characteristics are shared with FLK N 1–2 palimpsest. These parameters decrease in Koobi Fora sites, area C, and carnivore sites. There, the large perimeter-to-diameter ratio points toward more irregular clusters, and distances between remains in the overall pattern are larger. This is significantly marked in FLK NN and DK patterns. The outlier status of DK is attributed to the large dimension of its cluster, which is an anomaly compared to the mean of the entire sample.
Bones spatial patterns
Results obtained from the analysis of the high-density areas of bones yielded slightly different results. While most anthropic accumulations remain in the same cluster, Area A of DS seems to be different from the rest of DS areas. Koobi Fora sites are grouped together with several carnivore accumulations (FLK N levels 3–5, and FLK NN). FLK N 1–2 and DK are situated apart from the rest of the groups, however DK shows closer proximity to anthropic accumulations (Fig. 7c).
Dimension 1 explains mostly differences in cluster size and shape (perimeter and diameter). Anthropic sites (DS, PTK, FLK Zinj) and palimpsests (FLK N 1–2, DK) exhibit larger clusters. Carnivore bone accumulations seem to present more significant high-density areas, with greater distances between remains within the cluster. Koobi Fora sites appear to fall between these two groups, sharing some spatial traits with carnivores, FLK Zinj, and FLK N 1–2. Minimal discrepancies are observed in dimension 2 across most sites. Sites demonstrating more irregular cluster shapes and smaller secondary high-density areas, such as FLK N 1–2, FLK N 3, DK, FLK Zinj, and FLK NN, exhibit lower values on dimension 2. In DS, area A stands out for having the greatest distances between neighbors across the entire window, as well as the largest secondary high-density areas. On the contrary, FLK N 1–2 exhibits the shortest distances between neighbors throughout the entire window, coupled with a dense bone pattern both within the significant area and across the entire window. Once more, DK distinguishes itself by having the largest high-density area.
As expected, when we introduce hunter-gatherer campsites these are classified into a separate group (Fig. 7d). The main differences in dimension 1 between these campsites and the archaeological accumulations lie in the greater extension of the entire pattern (window) and the isolation of high-density areas (distance centroid window). Additionally, bones are more closely spaced throughout the entire pattern, and intensity is notably higher within clusters compared to outside the window in most accumulations considered. This indicates that hunter-gatherers exhibit a set of spatial behaviors that encompass extensive areas and are structured in more clearly defined clusters. Dimension 2 reveals variations in the cluster areas of Kunahajina (34.505) and //Gakwe Dwa 2 (!Kung) (9.648), as well as //oabe1 (81.228) and Kani//am/odi (56.250), along with differences in the number of clusters within the windows. Kunahajina and //Gakwe Dwa 2 exhibit a greater number of significant areas. //Gakwe Dwa 2 shows the largest distances between bones within the cluster, whereas //oabe1 exhibits the shortest.
Lithics spatial patterns
Factor maps of overall and bone patterns follow similar tendencies, yet lithic high-density areas are classified in a different manner. For example, DS areas B, C, and PTK lie nearer to carnivore and palimpsest sites (DK and FLK N 3–5) (Fig. 7b). Despite this, variations in height on the dendrogram map suggest the existence of two different subgroups (SI Fig Xb). Two distinct clusters are formed by area A of DS and FLK NN on the one side, and FLK Zinj, FLK N levels 1–2, and the Koobi Fora sites on the other side (Fig. 7b). Dimension 1 highlights variations in the size of primary high-density areas and secondary areas. FLK NN and DS area A exhibit smaller high-density areas, with secondary areas of comparable size to the main one. Distances between remains inside and outside the main areas are greater. The largest lithic areas are found in PTK, DS areas C and B, DK, and FLK N 4–5. Unlike area A, areas B and C lack additional secondary clusters. Koobi Fora sites exhibit similar size parameters as PTK, while FLK N 1–2 and FLK Zinj fall in between both groups. Dimension 2 highlights discrepancies in the number of significant high-density lithic areas, pattern intensity, and nearest-neighbor distances of the pattern. FLK N 1–2 and Koobi Fora sites exhibit multiple significant areas, whereas FLK Zinj shares the high intensity of the lithics accumulation with this group. On the contrary, carnivore sites, DK palimpsest, PTK, and DS present between 4 − 1 significant areas, with larger distances between remains, particularly in DS. Overall intensity decreases, although higher estimates are observed in anthropogenic sites.

Factor maps of hierarchical clustering on principal components analysis (HCPC) performed on the main high-density area of each: (a) overall patterns, (b) lithic patterns, (c) bone patterns, (d) bone patterns with hunter-gatherer campsites. Dendrograms are available in SI X
Discussion
Spatial characterization of anthropogenic accumulations
The anthropogenic accumulations have yielded a variety of results, all pointing towards the confirmation of functional links between bones and lithics. FLK Zinj, DS, and PTK share numerous taphonomic similarities. These include dense accumulations of intensely fractured small and medium-sized bovid carcasses in reduced areas, similarly high cut-mark and percussion-mark frequencies distributed on most anatomical parts, a low to moderate degree of carnivore ravaging, etc. They all confirm the functional association of faunal and lithic remains in the three sites (Domínguez-Rodrigo and Cobo-Sánchez 2017b; Cobo-Sánchez 2020; Organista et al. 2023). The outcomes of different multivariate statistical and machine learning analyses are consistent in the classification of these taphonomic signatures within a hominin-to-carnivore scenario (Domínguez-Rodrigo et al. 2014; Cobo-Sánchez 2020; Organista et al. 2023). Nearly all activity, including carcass procurement, bone defleshing, and bone breakage targeting marrow is attributed to hominins. Bone ravaging by hyenas was limited (Cobo-Sánchez et al. 2022) and slightly more intensive in PTK (Organista et al. 2023).
The HCPC results of the overall patterns suggest that there are spatial similarities in the main high-intensity areas of these three sites (five different accumulations if we count DS areas separately). They appear to be distinct from the spatial variables measured in carnivores and palimpsest assemblages. Previous spatial analyses have already delved into the common spatial characteristics of FLK Zinj, PTK, and DS (Domínguez-Rodrigo and Cobo-Sánchez 2017b; Cobo-Sánchez 2020; Diez-Martín et al. 2021). The intensity of bone accumulations is higher than in any other site analyzed here, however, the density of the DS lithic sub-assemblages is exceeded by sites like FxJj20 complex or FLK N levels 1–2. Both bones and lithics in anthropogenic sites appear to be agglomerated at distances < 0.3 m to then follow a regular pattern at distances > 0.5 m (the inhomogeneous distribution of lithics in PTK could not be discarded, see SI Fig. VII). This supports the spatial distribution of high-density areas we see on these sites and suggests that the accumulations are not the result of inhomogeneous random phenomena (e.g., caused by post-depositional processes such as fluvial processes), but a behavioral structured pattern commonly found in these anthropic assemblages.
FLK Zinj and PTK are characterized by single high-density areas with scattered and not agglomerated remains surrounding the main accumulation. This pattern differs from that seen at DS, where three different accumulations similar in size can be observed and confirmed with the kernel intensity estimation and the hot spot scan tests (Figs. 1 and 2). They have been analyzed separately to find any taphonomic or spatial variations that may suggest different formation processes (Cobo-Sánchez 2020). However, they appear to be the result of the hominin agency and seem to be equivalent to those found in FLK Zinj and PTK. Several taphonomic signals have led to the interpretation of areas B and C as the main accumulation. They could have been separated later by carnivore activity (hyenas as tested in Cobo-Sánchez et al. 2022) that dispersed and created the gap that we now identify as two different high-density areas. On the other hand, area A may represent a hominin-made secondary refuse area due to the overrepresentation of appendicular remains of larger carcasses and the lack of axial bones (Cobo-Sánchez 2020; Diez-Martin et al. 2021). In fact, the HCPC results reveal distinctions among these areas, particularly when the patterns are evaluated by types of remains. Whereas areas B and C are consistently grouped together, area A stands apart from the remaining anthropic sites (Fig. 7c-d), and in the analyses of lithic patterns is coupled with FLK NN (Fig. 7b). However, this distinction does not persist when evaluating the entire assemblage, as all DS areas appear similar, along with PTK and FLK Zinj (Fig. 7a).
Another ambivalence is revealed in the lithics distribution of FLK Zinj, which bears closer resemblance to FLK N 1–2 and Koobi Fora sites in the clustering analysis (Fig. 7b). Many tests yielded similar results for FLK Zinj and PTK lithics patterns (e.g., small and defined areas of significant probability of lithics higher than the average proportion, Fig. 4b; high positive correlation among lithics, SI Fig. VIII). Nevertheless, the measured parameters in their main high-density areas seem to present certain differences related to the intensity and concentration of lithics inside them (SI Table III). The main lithics hot spot at FLK Zinj is smaller than in PTK and areas B and C of DS, yet the mean intensity is significantly higher (FLK Zinj: 30.6 lithics per square meter; PTK: 13.3; DS area A: 13.5; DS area B: 7.2; DS area C: 7.2), lithics lie closer (n-n standardized value, FLK Zinj: 1.46; PTK: 1.67; DS area A: 3.23; DS area B: 1.61; DS area C: 1.83) and the average intensity of the entire window pattern is higher (FLK Zinj: 5.20 lithics per square meter; PTK: 2.60; DS area A: 2.51; DS area B: 1.94; DS area C: 1.91). This is also noticeable at the Koobi Fora sites and FLK N 1–2, indicating that the lithics within these five assemblages tend to be more densely clustered and concentrated. The composition of the lithic assemblages recovered at FLK Zinj and FLK N 1–2 is dominated by production activities that might leave a more pronounced spatial imprint, but battering activities not related to flaking processes seem to be relevant in the latter (de la Torre 2004).
The degree of intervention of hominins in Koobi Fora sites cannot be adequately addressed due to the poor cortical preservation of bones, which hinders accurate estimations of carnivore and hominin agency in the accumulations. The slightly better preservation of the FxJj50 assemblage showed butchering activities on certain bones, percussion marks, and fracture patterns similar to those generated by anthropogenic action (Bunn 1997). Primary access by hominins to some of these carcasses was inferred, and the intervention of carnivores as secondary agents was also recognized (Isaac 1997; Brantingham 1998; Domínguez-Rodrigo 2002). This is not the case for the FxJj20 complex, in which only the high concentration of spatially overlapping bones and lithics has been used to infer hominin agency, in addition to its likeness to other Koobi Fora sites. Both FxJj20 Main and FxJj20 East assemblages share similar patterns. Identifiable bone elements were few, the most abundant were teeth, followed by appendicular elements. There is a noticeable absence of epiphyses, and in FxJj20 East researchers documented fewer ribs than in FxJj20 Main (Bunn 1997). The three sites were formed in similar contexts, on a floodplain next to the riverbank and buried in low-moderate energy conditions (Bunn et al. 1997). Despite this, it seems sure that these anomalous accumulations are not the result of fluvial processes (Harris and Isaac 1997; Kroll 1997), and only hominins and carnivores are left to explain their formation. Since for now the results of the clustering analyses appear to be consistent with the agency of sites, we may shed light on how similar these sites are to other anthropogenic accumulations in terms of their spatial characteristics.
All Koobi Fora sites seem to exhibit common spatial characteristics that differ at times from the results obtained for FLK Zinj, PTK, and DS. It is noticeable that faunal remains at Koobi Fora localities are more dispersed, like some FLK N levels and DK (Table 5). In the pair correlation functions of overall patterns inhomogeneity and randomness cannot be discarded, and only regularity is statistically confirmed at distances > 0.5 m. In the clustering analyses, all Koobi Fora sites are consistently grouped together, suggesting that similar agencies and processes are behind their formation. Since both FxJj50 and FxJj20 complex are always set apart from the rest of the anthropogenic accumulations in Olduvai Bed I (apart from lithic patterns, where similarities with FLK Zinj are detected), we can hypothesize that different mechanisms might be proceeding in these sites. There are several possible explanations for that distinction: (1) the degree of hominin-carnivore contribution and activities performed on sites; (2) post-depositional processes that might have affected the original spatial configuration of remains; (3) the span time of the palimpsest formation.
As commented above, the conditions of the bone assemblages do not allow further exploration of the first potential explanation. We know that carnivores played a secondary role in carcasses of FxJj50, and they were probably hyaenas. FLK N palimpsests represent a common amenity scenario in which felids and hominins participated independently in the formation of the accumulations, and hyaenas ravaged interdependently the assemblages. On the other hand, FLK Zinj, DS, and PTK represent clear scenarios of hominin agency with posterior intervention of hyaenas. The analysis of stone tools through traceology and refitting showed evidence of total and partial knapping episodes spatially identified (Schick 1987; Harris and Isaac 1997; Braun et al. 2008). Tools were used to process meat and soft plants (Harris and Isaac 1997; Keeley 1997). These diverse activities are found on PTK, DS, and FLK Zinj (meat processing) and FLK N assemblages (plant processing). From a spatial perspective, Koobi Fora sites appear to lie between those two scenarios. When the overall and lithic patterns are contemplated, these sites are closer to FLK N 1–2 scenario, a palimpsest with independent carnivore and hominin activities (Fig. 7a-b). However, bone patterns are closely related to those found on palimpsests with stronger carnivore signals (Fig. 7c-d), which might suggest a more complicated formation process.
Post-depositional processes unrelated to secondary carnivore intervention are expected to modify the original spatial distribution of remains. When quantified, these alterations usually lead to some randomness and less agglomeration that may or may not preserve certain original properties (Domínguez-Rodrigo et al. 2017b). The second instance does not seem probable for Koobi Fora assemblages since this randomness has not been detected in the spatial analysis except at close distances (pair correlation functions in Fig. 5 and SI Figs. VI-VII). Given that fluvial processes might have been involved in these sites, certain movements and in particular the reorientation of remains could have taken place to a limited extent (Kaufulu 1987; Bunn et al. 1997; Schick 1997). This could be behind some of the inhomogeneity observed at short distances. Nonetheless, it should be noticed that similar inhomogeneous tendencies are present in other anthropogenic lithic accumulations (see PTK SI Fig. VII), and the general spatial distribution seems to be preserved (Kroll 1997).
As for the third instance, it is known that the formation of the Koobi Fora sites was the result of several occupations on different old surfaces, sometimes already buried between hominin visits, which led to the formation of a thick deposit containing the vertical distribution. The posterior dispersion of materials was not significant since the vertical movement of refitted bones and lithics is minimal (Kroll 1997). Vertically indistinguishable accumulations could potentially result in horizontal palimpsests, where the spatial distribution differs slightly from other anthropogenic occupations occurring in the same area. The Koobi Fora sites exhibit a higher number of high-density areas compared to the Olduvai anthropogenic sites, particularly notable in FxJj 50 and FxJj20 Main (Fig. 2). This might be associated with the different “old ground occupations’’ reported by Kroll (1997). Since FLK N palimpsests are the result of different independent events of several agents (felids, hominins, hyenids), an alternative explanation for the similarities among these faunal accumulations might be in the time deposition of the assemblages. In several tests, Koobi Fora sites resembled other accumulations such as FLK N levels 1–4 and DK (pair correlation functions in Fig. 5; nearest-neighbor mean for bones in Table 5).
What can be confidently asserted is that Koobi Fora sites exhibit common spatial characteristics with both anthropogenic and carnivore accumulations. The unique spatial patterns observed in these assemblages could be attributed to a combination of factors, including the degree of hominin-carnivore interaction or the complex site formation, which involved multiple occupations over time. This would explain the proximity of these sites to FLK N 1–2, the carnivore palimpsest with the strongest signal of hominins. This site demonstrated that the spatial overlapping of lithics and fossil bones is no guarantee of functional correlation. Since we do not count with a confirmed taphonomic functional association in the FxJj20 complex, similar inferences drawn in Koobi Fora maxi-sites should be carefully reevaluated. It is important to note that spatial analysis should not be seen as a replacement for taphonomic interpretations, and one should not place complete reliance on spatial analysis alone. The optimal approach would involve a combination of both methodologies together with lithic frequencies and refits, which has the potential to offer a more comprehensive understanding of the processes shaping the Koobi Fora sites.
Palimpsests and carnivore accumulations
The site FLK N 1–2 presents a noteworthy example in this investigation of a palimpsest with spatial mixed signals of both carnivore and hominin activity. Its results were usually different from those of its analogous levels, even though the lack of functional connection between bones and lithics is well known. At FLK N 1–2, faunal remains attributed mainly to Parmularius and Antidorcas were probably accumulated by felids, specifically leopards or saber-tooth felids, while hyenas had occasional access to some of the carcasses, further modifying them (Domínguez-Rodrigo and Barba 2007c; Domínguez-Rodrigo et al. 2022). On the other hand, the hominin agency at this site has been mainly related to battering activities (de la Torre 2004). Given the lack of typical percussion-related modifications on the bone surfaces and breakage planes, however, this activity seems to have been unrelated to bone demarrowing. This suggests that the deposition of bones and lithics were functionally independent and that the site may have been generated through common-amenity processes involving multiple independent taphonomic agents (Domínguez-Rodrigo et al. 2007a; Díez-Martín et al. 2010; Bunn et al. 2010; Domínguez-Rodrigo et al. 2022).
While some graphics display a correlation of remains similar to those of anthropogenic accumulations, other tests suggest that this assemblage is similar to other carnivore palimpsest accumulations. Despite the lack of statistical significance of the mark connection function, in the bone-lithic correlation one can detect a slight increase of positive correlation at distances of 1–1.5 m (SI Fig. IX). Notice that the main high-density area detected in bones and lithics have an average radius of 1.5 m and are spatially overlapping. It could be inferred the existence of a spatial covariate (presumably the presence/absence of trees as a cover) that is interacting with the accumulation of remains. This would result in the formation of these two high-density areas whose correlation is slightly detected in the mark connection function. However, its statistical significance cannot be proved, as other functions such as pair correlation suggest.
Similar patterns of bones and lithics correlations can be traced in all FLK N levels, but certain spatial instability (fluctuations or discontinuities in the graphic functions) and randomness are frequently found in other levels too (Fig. 6) and stand out when lithic patterns are analyzed (SI Fig. VII). Hominin and carnivore activities at FLK N levels 3, 4, and 5 seem to have been independent. Analysis of tooth-marking types, frequencies of notch types, and breakage plane angles indicate that carnivores were responsible for the bulk of the bone accumulations at these levels (Domínguez Rodrigo et al. 2007b; Egeland 2007b). Although stone tools are present, they appear to be unrelated to marrow extraction. Less than five cut marks across the three levels on anatomical locations that are not compatible with scavenging also indicate that these traces of hominin butchery might correspond to an independent depositional event. Regarding the lithic assemblage on FLK N, the low abundance of flakes or cutting tools with edges documented (de la Torre 2004) also suggests that hominins were not generally engaging in carcass processing at the site. That together with the extremely low density of stone tools, many of which were, in fact, ecofacts and not manuports (de la Torre 2004; de la Torre and Mora 2005), indicates that the signal of hominin activity in these levels was sporadic and not significant for the formation of the main bones’ accumulations. Díez-Martín et al. (2010) reached similar conclusions regarding the nature of activities performed on FLK N sites. Battering activities seem to dominate the compendium of hominin activities, however, some knapping sequences were detected too. Core reduction is documented as fragmented, and it appears that a great number of flakes were transported off-site. The fragmentation of knapping operations suggests a short and non-intensive participation of hominins in the accumulations, leading to a “non-organized hominin impact on the site and its resources” (Díez-Martín et al. 2010: 384). This non-organized presence the authors refer to seems to be confirmed statistically in certain levels of FLK N (3–5), where some tests yielded inhomogeneity and randomness. Thus, the inhomogeneity would be the result of the accumulation of lithics’ patterns being too small to discern any clear spatial organization.
Later excavations around FLK N revealed that all levels identified by Leakey (1971) were hard to distinguish in the vertical distribution of remains. Only the high-density accumulation of levels 1–2 was clear. Level 3 was barely distinguishable, and levels 4 and 5 were impossible to differentiate (Domínguez-Rodrigo et al. 2010). The clustering classification we applied here together with their spatial properties seems to confirm the similarities among levels 3–5, and part of their inhomogeneity and randomness could be explained by the artificial distinction among them. Whether or not these three levels should be considered one, their taphonomic stories do barely change and reflect the time-average palimpsest nature of FLK N accumulations. The presence of a freshwater spring nearby and a dry peninsula projecting into a large wetland area could explain why carnivores -and to a lesser degree hominins- were repeatedly attracted to this location (Ashley et al. 2010; Barboni et al. 2010; Domínguez-Rodrigo et al. 2010; Bunn et al. 2010).
The HCPC results consistently reveal a separation between levels 1–2 and levels 3–5 across all analyzed patterns (Fig. 7). Levels 1–2 align with Koobi Fora and FLK Zinj, particularly in lithic assemblages, while levels 3–5 show a distinct pattern, sharing affinities with FLK NN (in overall and bone patterns), Koobi Fora (in bones), and DK (in lithics). While it is hard to address the similarities between FLK N and Koobi Fora, it is evident that FLK NN and DK share common taphonomic stories with these levels, and this is reflected spatially too.
The spatial characteristics shown by DK and FLK NN were similar in some instances to those found in FLK N. The formation of FLK NN is primarily attributed to carnivores, and the almost complete absence of tools suggests that the degree of hominin involvement in these accumulations was much less compared to FLK N 1–5 and DK (Barba and Domínguez-Rodrigo 2007; Domínguez-Rodrigo and Barba 2007d; Egeland 2007b). Unlike FLK NN and FLK N, DK shows evidence of a more complex formation process, where multiple factors contribute to the accumulation of materials. Carnivores, hominins, and natural processes all seem to play roles in shaping the palimpsest assemblage at DK (Egeland 2007c) and might explain the reason why this accumulation appears to be an outlier in most HCPC analyses.
These sites showed that the bone-lithic and bone correlation was clear at some distances, but the lithics correlation was irregular, with certain peaks of positive correlation (SI Fig IX). The segregation between both types of remains was marked, especially for FLK NN (see nearest-neighbor correlation values among other tests), and mean distances among neighbors were the highest in both sites. It is significant that inhomogeneity and randomness cannot be ruled out in the lithic distribution of these assemblages (SI Fig. VII). However, it should again be noted that the spatial sample analyzed in DK represents less than 10% of the total set for both industry and fauna. This could affect some results, especially those tests or analyses that rely on intensity estimation, and could also explain the outlier effect in the total and bone distributions.
DK is only grouped in the lithic classification together with other carnivore palimpsests and anthropogenic assemblages (FLK N 3–5, PTK, and areas B and C of DS). In the classification of lithics, the focus does not lie on the identification of carnivores or hominins, as these accumulations are unique to the latter. Instead, results may point out variations in carving and resource-processing behaviors among hominids. However, we face a limitation in terms of references from spatial studies on Early Pleistocene assemblages, hindering a comprehensive interpretation of these results. The main differences among lithic accumulations might be related to the temporal lapse of the formation of the site and the type of activities carried out on site. The number and density of lithics in most palimpsests and carnivore accumulations suggest that hominin activity in these referential places was sporadic and scattered. In the FLK N site, most activities appear to revolve around percussion, likely related to processing plant resources. Plant and wood processing has been reported in several Koobi Fora sites too (Keeley and Tooth 1981). At the same time, many stone tools originally classified as manuports are in fact ecofacts with no anthropogenic origin (de la Torre and Mora 2005). On the contrary, at sites like DS, it has been documented that both fleshing and bone percussion activities occur across all areas, with evidence of spatial segregation as well (Cobo-Sánchez 2020; Díaz Martín et al. 2021). The analysis of the DK lithic assemblage seems to indicate a predominance of knapping activities, with a predominance of local lava raw material (de la Torre 2004). Given the identification of some anthropic action in some bone assemblages, these carving activities could be linked to meat processing, such as those identified in DS. It appears that some minor fluvial processes might have removed lithic remains smaller than 20 mm. Despite this, the assemblage seems to preserve its integrity, and even some refittings have been documented (de la Torre 2004). Even knowing this, it is hard to explain the similarities found among DK and PTK, DS, and other main carnivore assemblages, or the closeness between DS area A and FLK NN.
Addressing the spatial imprint of agency and formation processes
After a thorough analysis of all the archaeological sites, it is pertinent to revisit the initial questions we posed. Certain dissonances detected in the spatial configuration of Early Pleistocene archaeological palimpsests may be related to the formation processes behind these accumulations. In our findings, it appears that we can indeed establish significant distinctions between anthropic and non-anthropic overall assemblages. Our hypothesis regarding carnivore accumulations being less dense and clustered compared to hominins’ seems to be confirmed. Furthermore, we have detected differences in the shape of the high-density areas (more irregular) and the number of accumulations (greater than in anthropogenic assemblages). We have observed that inhomogeneity and randomness can indicate the absence of significant spatial patterns when the agents responsible are barely involved in the formation process. This seems to be the case of lithics in carnivore assemblages. The correlation of remains cannot be used to address agency directly; however, cases of random labeling of remains at certain distances were only observed among carnivore and palimpsest accumulations.
Previous studies have shown that some aspects of the social behavior and organization of hominins can be traced in the distribution of the archaeological remains (Clark and Gingerich 2022; Domínguez-Rodrigo and Cobo Sánchez 2017a, 2017b; Kroll and Price 1991; Yellen 1977). Although all agents examined in this study (i.e., felids, hyenids, hominins) tend to accumulate carcasses (Cavallo and Blumenschine 1989; Fourvel 2012; Arriaza et al. 2016; Domínguez-Rodrigo and Cobo-Sánchez 2017b; Cobo-Sánchez 2020; Domínguez-Rodrigo et al. 2022), it is possible that some spatial differences between the assemblages may be attributable to variations in their socio-reproductive systems and food acquisition strategies. In this sense, we can assert that the social organization of the first hominins and its spatial imprint differ significantly for modern hunter-gatherer societies: the multi-cluster spatial distribution of bones found in modern hunter-gatherer camps, related to their organization in single households with multiple clusters (Yellen 1977; Bartram et al. 1991; Domínguez-Rodrigo and Cobo-Sánchez 2017a), differs in the clustering analysis from the single-cluster organization of the first archaeological sites (Domínguez-Rodrigo and Cobo-Sánchez 2017b), which resulted from the collective consumption of food in referential places. This study confirms the existence of such differences with the results of the clustering analysis of bones.
Another relevant question for disentangling hominin-carnivore interactions in Early Pleistocene sites is if we can distinguish spatially between independent and interdependent palimpsests. This task becomes challenging because the spatial occurrence of objects in the same location will inevitably lead to some form of correlation, whether positive or negative. For instance, one might expect cases of inhibition where events are independent. However, we have observed that inhibition can also occur among functionally related sets. Inhomogeneity cannot be equated solely to the independence of events. The absence of a driving agent of accumulation or the loss of the original distribution are other possible factors behind results like this.
It appears that interdependent deposits still preserve the original spatial configurations when an agent stands out. Such situations are found among DS, PTK, and FLK Zinj, where a very low carnivore ravaging took place and has not blurred the spatial imprint of hominins. Examples of independent palimpsests are those sites labeled as carnivore-made, formed by bone accumulations performed by carnivores (probably through interdependent events of primary consumption and posterior ravaging) and sporadic lithic knapping and battering episodes unrelated to bone assemblages (FLK N 3–5, FLK NN). However, due to the low contribution of hominins, these specific cases share similar spatial patterns, probably reflecting more carnivore than hominins spatial behavior. The contribution of hominins in FLK N 1–2 and DK independent palimpsests is significantly higher than in any other non-hominin sites. In these sites, mixed spatial characteristics are detected, which suggests that both carnivore and hominin spatial imprints are still present in the distribution of remains. In general, it seems like independent accumulations are clearly detected only when the degree of contribution of all agents is similar. However, we cannot evaluate the spatial properties of interdependent palimpsests as we do not have a strong representation of situations in which hominins and carnivores modify the same carcasses or where hominins act as scavengers of carnivores.
The spatial statistical analyses have revealed certain differences among sites, but quantifying the input of specific taphonomic agents remains a challenge. The focus on high-density areas helped us reach another level and yielded promising results, as several HCPC dendrograms (SI Fig. X) achieved accurate classifications of overall and bone accumulations in terms of agency. While its results provide some insight into the data, a significant portion of the variability remains unaccounted for. Therefore, the current approach has limitations in fully capturing the complexity and nuances present in the archaeological assemblages. And this can be especially true for lithics accumulation. Indeed, the discrepancy in the level of success between bones and overall patterns compared to lithics accumulation further highlights the complexity of interpreting these assemblages. Lithics might follow different spatial patterns or formation processes than bones in these assemblages. Further investigations and additional spatial methods might be necessary to address the unexplained variance and gain a more comprehensive understanding of the patterns and processes involved in the formation and functionality of these assemblages. Combining the HCPC results with other statistical techniques, taphonomic studies, and exploring new approaches to machine learning could potentially lead to a more robust and comprehensive interpretation of the archaeological data.
The main challenge encountered when dealing with old spatial data is the significant loss of information. In most planimetries, the entire assemblage is not plotted, and in some cases, only identifiable faunal remains and main lithic pieces were represented, with no spatial information of unidentifiable fauna or small debris. This reduces considerably the analyzed samples, as is the case of DK assemblage and faunal remains of FxJj20 complex sites, whose samples represent less than 10% of all recovered pieces (Table 2). Despite this having significant effects on the estimation of the overall intensity of the patterns, the general spatial arrangement could be assessed in all archaeological sites, and statistical detection of high-density areas has even converged with on-field observations noted in reports. This suggests that the distribution of remains still reflects the main spatial configuration of the accumulations and can serve as approximations.
Another problem any spatial study confronts when trying to interpret the behavioral patterns found in archaeological assemblages is the lack of referential frameworks. The diverse performances on carcasses among carnivores have been deeply studied (Blumenschine 1986, 1988; Domínguez-Rodrigo 2001; Pickering 2002; Arriaza et al. 2015, 2016, 2019), however, we lack equivalent models for how these carnivores accumulate and modify assemblages from a spatial perspective. Several experiments have recorded the spatial movement of bones produced by carnivore intervention in carcasses (Marean and Bertino 1994; Arriaza et al. 2019) and simulated hearths and camps (Camarós et al. 2013; Arilla et al. 2020), but none of these recorded data have been analyzed yet spatially or compared with anthropic accumulations. It is crucial that future spatial studies focus on conducting controlled experiments involving carnivores to recreate diverse scenarios of dependent and interdependent activities.
Conclusion
This study provides new evidence of significant spatial variations among Early Pleistocene assemblages related to the different agents that created and modified these accumulations. Importantly, FLK Zinj, DS, and PTK stand out from the other sites as anthropogenic assemblages due to their unique spatial traits. Sites with a higher amount of carnivore input, such as FLK N and FLK NN also share spatial similarities and appear clearly differentiated from anthropogenic sites, while the Koobi Fora assemblages lie in an intermediate position.
Approaching agency by means of spatial statistical methods proves to be useful. By concentrating on high-density areas where the main events occurred, we gain valuable insights and uncover spatial dissonances in archaeological assemblages. This approach provides information on the spatial imprint of different agents. Nevertheless, the interpretation of functionality and formation mechanisms of archaeological assemblages should not be based uniquely on spatial criteria. To gain a more comprehensive understanding, it is essential to complement spatial analyses with taphonomic studies, lithic frequencies and refits, and additional robust statistical methods. In addition to this, we advocate for future spatial taphonomic studies to approach not only main accumulations but other meaningful small accumulations or scatter areas identified in a site, as this can provide further insights into the formation processes of Early Pleistocene assemblages.
We wish to emphasize the importance of experimentation and the need for referential frameworks to enhance the robustness when interpreting the spatial signals identified in this study. By replicating different scenarios of hominin-carnivore interaction, such as hominin activities, carnivore accumulations and modifications, and other post-depositional processes, we can gain insights into the formation and alteration of accumulations that have shaped archaeological assemblages.
Data availability
Data is included as supplementary material.
Code Availability
Code is included as supplementary material.
References
Arilla M, Rosell J, Blasco R (2020) A neo-taphonomic approach to human campsites modified by carnivores. Sci Rep 10:6659. https://doi.org/10.1038/s41598-020-63431-8
Arriaza MC, Domínguez-Rodrigo M, Martínez-Maza C, Mabulla A, Baquedano E (2015) Differential predation by age and sex classes in blue wildebeest in Serengeti: study of a modern Carnivore den in Olduvai Gorge (Tanzania). PLoS ONE 10. https://doi.org/10.1371/journal.pone.0125944
Arriaza MC, Domínguez-Rodrigo M, Yravedra J, Baquedano E (2016) Lions as Bone accumulators? Paleontological and ecological implications of a modern bone assemblage from Olduvai Gorge. PLoS ONE 11. https://doi.org/10.1371/journal.pone.0153797
Arriaza MC, Domínguez-Rodrigo M (2016) When felids and hominins ruled at Olduvai Gorge: A machine learning analysis of the skeletal profiles of the non-anthropogenic Bed I sites. Quat Sci Rev 139: 43-52. https://doi.org/10.1016/j.quascirev.2016.03.005
Arriaza MC, Organista E, Yravedra J, Santonja M, Baquedano E, Domínguez-Rodrigo M (2019) Striped hyenas as bone modifiers in dual human-to-Carnivore experimental models. Archaeol Anthropol Sci 11:3187–3199. https://doi.org/10.1007/s12520-018-0747-y
Ashley GM, Barboni D, Domínguez-Rodrigo M, Bunn HT, Mabulla AZP, Díez-Martín F, Barba R, Baquedano E (2010) Paleoenvironmental and paleoecological reconstruction of a freshwater oasis in Savannah grassland at FLK North, Olduvai Gorge, Tanzania. Quat Res 74(3):333–343. https://doi.org/10.1016/j.yqres.2010.08.006
Baddeley A (2008) Analysing spatial point patterns in R. CSIRO, Australia
Baddeley AJ, Møller J, Waagepetersen R (2000) Non-and semi‐parametric estimation of interaction in inhomogeneous point patterns. Stat Neerl 54(3):329–350. https://doi.org/10.1111/1467-9574.00144
Baddeley A, Rubak E, Turner R (2016) Spatial point patterns: methodology and applications with R. CRC, New York
Barba R, Domínguez-Rodrigo M (2007) FLK North North 1: living floor or natural accumulation? In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 217–228
Barboni D, Ashley GM, Domínguez-Rodrigo M, Bunn HT, Mabulla AZP, Baquedano E (2010) Phytoliths infer locally dense and heterogeneous paleovegetation at FLK North and surrounding localities during upper Bed I time, Olduvai Gorge, Tanzania. Quat Res 74(3):344–354. https://doi.org/10.1016/j.yqres.2010.09.005
Bargalló A, Gabucio MJ, Rivals F (2015) Puzzling out a palimpsest: testing an interdisciplinary study in level O of Abric Romaní. Quat Int 417:51–65. https://doi.org/10.1016/j.quaint.2015.09.066
Bartram LE (1993) An etnoarchaeological analysis of Kua San (Botswana) bone food refuse. Ph.D. dissertation, University of Wisconsin
Bartram LE, Kroll EM, Bunn HT (1991) Variability in Camp structure and Bone Food Refuse Patterning at Kua San Hunter-Gatherer Camps. In: Kroll EM, Price TD (eds) The interpretation of Archaeological spatial patterning. Springer US, Boston, pp 77–148
Bellomo RV (1994) Methods of determining early hominid behavioural activities associated with the controlled use of fire at FxJj 20 Main. J Hum Evol 27:173–195. https://doi.org/10.1006/jhev.1994.1041
Binford LR (1981) Bones: ancient men and modern myths. Academic, Orlando
Binford LR, Bunn HT, Kroll EM (1988) Fact and fiction about the Zinjanthropus floor: data, arguments, and interpretations. Curr Anthropol 29(1):123–149. https://doi.org/10.1086/203618
Bivand RS, Pebesma E, Gómez-Rubio V (2013) Applied spatial data analysis with R. Springer, New York
Bivand R, Keitt T, Rowlingson B (2023a) Rgdal: Bindings for the ‘Geospatial’ Data Abstraction Library_. R package version 1.6-6, https://CRAN.R-project.org/package=rgdal
Bivand R, Lewin-Koh N, Pebesma E et al (2023b) Package ‘Maptools’: Tools for Reading and Handling Spatial Objects. R package version 1.1-8, https://CRAN.R-project.org/package=maptools
Blumenschine RJ (1986) Carcass consumption sequence and the archaeological distinction of scavenging and hunting. J Hum Evol 15(8):639–659. https://doi.org/10.1016/S0047-2484(86)80002-1
Blumenschine RJ (1988) An experimental model of the timing of Hominid and Carnivore influence on archaeological bone assemblages. J Archaeol Sci 15(5):483–502. https://doi.org/10.1016/0305-4403(88)90078-7
Blumenschine RJ (1995) Percussion marks, tooth marks, and experimental determinations of the timing of Hominid and Carnivore access to long bones at FLK Zinjanthropus, Olduvai Gorge. Tanzan J Hum Evol 29(1):21–51. https://doi.org/10.1006/jhev.1995.1046
Brantingham PJ (1998) Mobility, competition and plio-pleistocene hominid foraging groups. J Archaeol Method Theory 5(1):57–98. https://doi.org/10.1007/BF02428416
Brown FH, Feibel CS (1986) Revision of lithostratigraphic nomenclature in the Koobi Fora region, Kenya. J Geol Soc Lond 143(2):297–310. https://doi.org/10.1144/gsjgs.143.2.0297
Brown FH, McDougall I (2011) Geochronology of the Turkana depression of Northern Kenya and Southern Ethiopia. Evol Anthropol 20:217–227. https://doi.org/10.1002/evan.20318
Bunn HT (1981) Archaeological evidence for meat-eating by Plio-Pleistocene hominids from Koobi Fora and Olduvai Gorge. Nature 291:574–577. https://doi.org/10.1038/291574a0
Bunn HT (1982) Meat-eating and human evolution: studies on the diet and subsistence patterns of Plio-Pleistocene hominids in East Africa, Ph.D. Dissertation. University of California
Bunn HT (1986) Patterns of skeletal representation and hominid subsistence activities at Olduvai Gorge, Tanzania, and Koobi Fora. Kenya J Hum Evol 15:673–690. https://doi.org/10.1016/S0047-2484(86)80004-5
Bunn HT (1991) A taphonomic perspective on the Archaeology of Human origins. Annu Rev Anthropol 20:433–467. https://doi.org/10.1146/annurev.an.20.100191.002245
Bunn HT (1997) The bone assemblages from the excavated sites. In: Isaac GL, Isaac B (eds) Plio-Pleistocene archaeology. Koobi Fora Research Project. Volumen V. Cambridge University Press, Cambridge, pp 402–458
Bunn HT, Kroll EM (1986) Systematic butchery by Plio-Pleistocene hominids at Olduvai Gorge, Tanzania. Curr Anthropol 27:431–452
Bunn HT, Harris JWK, Isaac G, Kaufulu Z, Kroll E, Schick K, Toth N, Behrensmeyer AK (1980) FxJj50: an early pleistocene site in northern Kenya. World Archaeol 12:109–136. https://doi.org/10.1080/00438243.1980.9979787
Bunn HT, Mabulla AZP, Domínguez-Rodrigo M, Ashley GM, Barba R, Díez-Martín F, Remer K, Yravedra J, Baquedano E (2010) Was FLK North levels 1–2 a classic living floor of Oldowan hominins or a taphonomically complex palimpsest dominated by large Carnivore feeding behavior? Quat Res 74(3):355–362. https://doi.org/10.1016/j.yqres.2010.06.004
Camarós E, Cueto M, Teira LC, Tapia J, Cubas M, Blasco R, Rosell J, Rivals F (2013) Large carnivores as taphonomic agents of space modification: an experimental approach with archaeological implications. J Archaeol Sci 40:1361–1368. https://doi.org/10.1016/j.jas.2012.09.037
Castielo ME (2022) Computational and Machine Learning Tools for Archaeological Site modeling. Springer, Cham
Cavallo JA, Blumenschine RJ (1989) Tree-stored leopard kills: expanding the hominid scavenging niche. J Hum Evol 18(4):393–399. https://doi.org/10.1016/0047-2484(89)90038-9
Clark PJ, Evans FC (1954) Distance to Nearest Neighbor as a measure of spatial relationships in populations. Ecology 35(4):445–453. https://doi.org/10.2307/1931034
Clark AE, Gingerich AM (2022) Intrasite Spatial Analysis of Mobile and Semisedentary peoples. University of Utah, Salt Lake City
Cobo-Sánchez L (2020) Taphonomic and spatial study of the archaeological site DS from bed I (Olduvai Gorge). Ph.D. dissertation, Complutense University
Cobo-Sánchez L, Pizarro-Monzo M, Cifuentes-Alcobendas G, Jiménez García B, Abellán Beltrán N, Courtenay LA, Mabulla A, Baquedano E, Domínguez-Rodrigo M (2022) Computer vision supports primary access to meat by early Homo 1.84 million years ago. PeerJ 10:e14148. https://doi.org/10.7717/peerj.14148
R Core Team (2023) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Davies TM, Marshall JC, Hazelton ML (2017) Tutorial on kernel estimation of continuous spatial and spatiotemporal relative risk. Stat Med 37(7):1191–1221. https://doi.org/10.1002/sim.7577
de la Torre I (2004) Estrategias tecnológicas en el Pleistoceno inferior de África oriental (Olduvai y Peninj, norte de Tanzania). Ph.D. dissertation, Complutense University
de la Torre I, Mora R (2005) Unmodified lithic material at Olduvai Bed I: manuports or ecofacts? J Archaeol Sci 32:273–285. https://doi.org/10.1016/j.jas.2004.09.010
Deino AL (2012) 40Ar/39Ar dating of Bed I, Olduvai Gorge, Tanzania, and the chronology of early pleistocene climate change. J Hum Evol 63(2):251–273. https://doi.org/10.1016/j.jhevol.2012.05.004
Diez-Martin F, Sanchez Yustos P, Domínguez-Rodrigo M, Mabulla A, Bunn H, Ashley G, Barba R, Baquedano E (2010) New insights into hominin lithic activities at FLK North Bed I, Olduvai Gorge, Tanzania. Quat Res 74(3):376–387. https://doi.org/10.1016/j.yqres.2010.07.019
Diez-Martín F, Cobo-Sánchez L, Baddeley A, Uribelarrea D, Mabulla A, Baquedano E, Domínguez-Rodrigo M (2021) Tracing the spatial imprint of Oldowan technological behaviors: a view from DS (Bed I, Olduvai Gorge, Tanzania). PLoS ONE 16(7). https://doi.org/10.1371/journal.pone.0254603
Diez-Martín F, Cobo-Sánchez L, Fraile-Márquez C, de Francisco S, Duque-Martínez J, Sánchez-Yustos P, Baquedano E, Mabulla A, Domínguez-Rodrigo M (2023) Searching for intra-site spatial patterns in the African Early Acheulean: the lowermost archaeo-units at FLK West (Olduvai Gorge Tanzania). Archaeol Anthropol Sci 15(5) https://doi.org/10.1007/s12520-023-01773-w
Diggle PJ (1985) A Kernel Method for smoothing point process data. J R Stat Soc C: Appl Stat 34(2):138–147. https://doi.org/10.2307/2347366
Diggle PJ (2014) Statistical analysis of spatial and spatio-temporal point patterns. Taylor and Francis Group, Florida
Domínguez-Rodrigo M (2001) A study of Carnivore competition in riparian and open habitats of modern savannas and its implications for hominid behavioural modelling. J Hum Evol 40(2):77–98. https://doi.org/10.1006/jhev.2000.0441
Domínguez-Rodrigo M (2002) Hunting and scavenging by early humans: the state of the debate. J World Prehist 16(1):1–54. https://doi.org/10.1023/A:1014507129795
Domínguez-Rodrigo M, Barba R (2007a) New estimates of tooth-mark and percussion-mark frequencies at the FLK Zinjanthropus level: the Carnivore–Hominid–Carnivore hypothesis falsified (I). In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 39–74
Domínguez-Rodrigo M, Barba R (2007b) The behavioral meaning of cut marks at the FLK Zinj level: the Carnivore-Hominid-Carnivore hypothesis falsified (II). In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 75–100
Domínguez-Rodrigo M, Barba R (2007c) A palimpsest at FLK North 1–2: independent Carnivore- and hominid-made bone accumulations. In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 127–163
Domínguez-Rodrigo M, Barba R (2007d) Reanalysis of FLK North North 3: yet another case of a palimpsest? In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 239–252
Domínguez-Rodrigo M, Cobo-Sánchez L (2017a) The spatial patterning of the social organization of modern foraging Homo sapiens: a methodological approach for understanding social organization in prehistoric foragers. Palaeogeogr Palaeoclimatol Palaeoecol 488:113–125. https://doi.org/10.1016/j.palaeo.2017.06.008
Domínguez-Rodrigo M, Cobo-Sánchez L (2017b) A spatial analysis of stone tools and fossil bones at FLK Zinj 22 and PTK I (Bed I, Olduvai Gorge, Tanzania) and its bearing on the social organization of early humans. Palaeogeogr Palaeoclimatol Palaeoecol 488:21–34. https://doi.org/10.1016/j.palaeo.2017.04.010
Domínguez-Rodrigo M, Barba R, Egeland CP (2007a) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York
Domínguez-Rodrigo M, Barba R, Organista E (2007b) A taphonomic study of FLK North 3 and 4: a felid–hyaenid and hominid palimpsest. In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 165–189
Domínguez-Rodrigo M, Mabulla AZP, Bunn HT, Díez-Martín F, Baquedano E, Barboni D, Barba R, Domínguez-Solera S, Sánchez P, Ashley GM, Yravedra J (2010) Disentangling hominin and Carnivore activities near a spring at FLK North (Olduvai Gorge, Tanzania). Quat Res 74(3):363–375. https://doi.org/10.1016/j.yqres.2010.07.004
Domínguez-Rodrigo M, Bunn HT, Yravedra J (2014) A critical re-evaluation of bone surface modification models for inferring fossil hominin and Carnivore interactions through a multivariate approach: application to the FLK Zinj archaeofaunal assemblage (Olduvai Gorge, Tanzania). Quat Int 322–323:32–43. https://doi.org/10.1016/j.quaint.2013.09.042
Domínguez-Rodrigo M, Cobo-Sánchez L, Uribelarrea D, Arriaza MC, Yravedra J, Gidna A, Organista E, Sistiaga A, Martín-Perea D, Baquedano E, Aramendi J, Mabulla A (2017a) Spatial simulation and modeling of the early pleistocene site of DS (Bed I, Olduvai Gorge, Tanzania): a powerful tool for predicting potential archaeological information from unexcavated areas. Boreas 46:805–815. https://doi.org/10.1111/bor.12252
Domínguez-Rodrigo M, Cobo-Sánchez L, Yravedra J, Uribelarrea D, Arriaza MC, Organista E, Baquedano E (2017b) Fluvial spatial taphonomy: a new method for the study of post-depositional processes. Archaeol Anthropol Sci 10:1769–1789. https://doi.org/10.1007/s12520-017-0497-2
Domínguez-Rodrigo M, Baquedano E, Organista E, Cobo-Sánchez L, Mabulla A, Maskara V et al (2021) Early pleistocene faunivorous hominins were not kleptoparasitic, and this impacted the evolution of human anatomy and socio-ecology. Sci Rep 11(1):16135. https://doi.org/10.1038/s41598-021-94783-4
Domínguez-Rodrigo M, Organista E, Baquedano E, Cifuentes-Alcobendas G, Pizarro-Monzo M, Vegara-Riquelme M, Gidna A, Uribelarrea D, Martín-Perea D (2022) Neo-taphonomic analysis of the Misiam leopard lair from Olduvai Gorge (Tanzania): understanding leopard–hyena interactions in open settings. R Soc Open Sci 9. https://doi.org/10.1098/rsos.220252
Egeland CP (2007a) Zooarchaeology and taphonomy of FLK North 5. In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 191–200
Egeland CP (2007b) Zooarchaeology and taphonomy of FLK North North 2. In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 229–237
Egeland CP (2007c) Zooarchaeology and taphonomy of the DK Site. In: Domínguez-Rodrigo M, Barba R, Egeland CP (eds) Deconstructing Olduvai. A taphonomic study of the Bed I sites. Springer, New York, pp 253–268
Fourvel JB (2012) Hyénidés modernes et fossiles d’Europe et d’Afrique: taphonomie comparée de leurs assemblages osseux. Archéologie et Préhistoire. Ph.D. dissertation, University of Toulouse
Harris JWK, Isaac GL (1997) Sites in the upper KBS, Okote, and Chari members: reports. In: Isaac GL, Isaac B (eds) Plio-Pleistocene archaeology. Koobi Fora Research Project. Volume V. Cambridge University, Cambridge, pp 115–236
Hay RL, Kyser TK (2001) Chemical sedimentology and paleoenvironmental history of Lake Olduvai, a Pliocene lake in northern Tanzania. Geol Soc Am Bull 113(12):1505–1521. https://doi.org/10.1130/0016-7606(2001)113%3C1505:CSAPHO%3E2.0.CO;2
Hazelton ML, Davies TM (2009) Inference based on kernel estimates of the relative risk function in geographical epidemiology. Biom J 51:98–109. https://doi.org/10.1002/bimj.200810495
Hopkins B, Skellam JG (1954) A New Method for determining the type of distribution of Plant individuals. Ann Bot 18:213–227. https://doi.org/10.1093/oxfordjournals.aob.a083391
Husson F, Josse J, Lê S, Mazet J (2007) FactoMineR: Factor Analysis and Data Mining with R. R package version 1.04. http://CRAN.R-project.org/package=FactoMineR
Isaac GL (1983) Aspects of human evolution. Conference Paper. In: Isaac B (ed) The archaeology of human origins. Papers by Glynn Isaac, Cambridge University Press, Cambridge, pp. 96–119
Isaac GL (1997) Plio-Pleistocene archaeology. Koobi Fora Research Project. Volumen V. Cambridge University Press, Cambridge
Isaac GLl (1984) The archaeology of human origins: studies of Lower Pleistocene in East Africa, 1971–1981. Adv World Archaeol 3:1–87
Kaufulu ZM (1987) Formation and preservation of some earlier Stone Age sites at Koobi Fora, Northern Kenya. South Afr Archaeol Bull 42(145):23–33. https://doi.org/10.2307/3887770
Keeley L (1997) Microwear traces on a selected sample of stone artifacts from Koobi Fora. In: Isaac GL, Isaac B (eds) Koobi Fora Research Project. Plio-Pleistocene archaeology, vol 5. Cambridge University Press, Cambridge, pp 224–233
Keeley L, Toth N (1981) Microwear polishes on early stone tools from Koobi Fora, Kenya. Nature 293:464–465. https://doi.org/10.1038/293464a0
Kelsall JE, Diggle PJ (1995) Kernel Estimation of relative risk. Bernoulli 1(1/2):3–16. https://doi.org/10.2307/3318678
Kempf M, Günther G (2023) Point pattern and spatial analysis using archaeological and environmental data. A case study from the neolithic Carpathian Basin. J Archaeol Sci Rep 47:103747. https://doi.org/10.1016/j.jasrep.2022.103747
Kroll EM (1997) Lithic and faunal distributions at eight archaeological excavations. In: Isaac GL, Isaac B (eds) Koobi Fora Research Project. Plio-Pleistocene archaeology, vol 5. Cambridge University Press, Cambridge, pp 459–538
Kroll EM, Price TD (1991) The interpretation of Archaeological spatial patterning. Plenum, New York
Kulldorff M (1997) A spatial scan statistic. Commun Stat Theory Methods 26:1481–1496. https://doi.org/10.1080/03610929708831995
Lê S, Josse J, Husson F (2008) FactoMineR: an R package for multivariate analysis. J Stat Softw 25(1):1–18
Leakey MD (1971) Olduvai Gorge: volume 3, excavations in beds I and II, 1960–1963. Cambridge University Press, Cambridge
Marean CW, Bertino L (1994) Intrasite spatial analysis of bone: subtracting the Effect of secondary Carnivore consumers. Am Antiq 59(4):748–768. https://doi.org/10.2307/282346
Mehrer MW, Wescott KL (2006) GIS and Archaeological Site Location modeling. CRC Press Taylor & Francis Group, Boca Raton
Méndez-Quintas E, Santonja M, Pérez-González A, Díaz-Rodríguez M, Domínguez AS (2022) Exploring the formation processes on open-air palaeolithic sites: a late Middle Pleistocene Acheulean assemblage at Arbo site (Miño River basin, Spain). J Archaeol Sci Rep 43:103453. https://doi.org/10.1016/j.jasrep.2022.103453
Moclán A, Huguet R, Márquez B, Álvarez-Fernández A, Laplana C, Arsuaga JL, Pérez-González A, Baquedano E (2023a) Identifying activity areas in a neanderthal hunting camp (the Navalmaíllo Rock Shelter, Spain) via spatial analysis. Archaeol Anthropol Sci 15:44. https://doi.org/10.1007/s12520-023-01746-z
Moclán A, Cobo-Sánchez L, Domínguez-Rodrigo M, Méndez-Quintas E, Rubio-Jara S, Panera J, Pérez-González A, Santonja M (2023b) Spatial analysis of an early middle palaeolithic kill/butchering site: the case of the Cuesta De La Bajada (Teruel, Spain). Archaeol Anthropol Sci 15:91. https://doi.org/10.1007/s12520-023-01792-7
Oliver JS, Plummer TW, Hertel F, Bishop LC (2019) Bovid mortality patterns from Kanjera South, Homa Peninsula, Kenya and FLK-Zinj, Olduvai Gorge, Tanzania: evidence for habitat mediated variability in Oldowan hominin hunting and scavenging behavior. J Hum Evol 131:61–75. https://doi.org/10.1016/j.jhevol.2019.03.009
Organista E, Moclán A, Aramendi J, Cobo-Sánchez L, Egeland CP, Uribelarrea D, Martín-Perea D, Vegara-Riquelme M, Hernández-Vivanco L, Gidna A, Mabulla A, Baquedano E, Domínguez-Rodrigo M (2023) A taphonomic analysis of PTK (Bed I, Olduvai Gorge) and its bearing on the interpretation of the dietary and eco-spatial behaviors of early humans. Q Sci Rev 300:107913. https://doi.org/10.1016/j.quascirev.2022.107913
Panera J, Rubio-Jara S, Domínguez-Rodrigo M, Yravedra J, Méndez-Quintas E, Pérez-González A, Bello-Alonso P, Moclán A, Baquedano E, Santonja M (2019) Assessing functionality during the early Acheulean in level TKSF at Thiongo Korongo site (Olduvai Gorge, Tanzania). Quat Int 526:77–98. https://doi.org/10.1016/j.quaint.2019.09.013
Parkinson JA (2018) Revisiting the hunting-versus-scavenging debate at FLK Zinj: a GIS spatial analysis of bone surface modifications produced by hominins and carnivores in the FLK 22 assemblage, Olduvai Gorge, Tanzania. Palaeogeogr Palaeoclimatol Palaeoecol 511:29–51. https://doi.org/10.1016/j.palaeo.2018.06.044
Pebesma EJ, Bivand RS (2005) Classes and methods for spatial data in R. R News 5 (2). https://cran.r-pro-ject.org/doc/Rnews/
Peters C, van Kolfschoten T (2020) The site formation history of Schoningen 13II-4 (Germany): testing different models of site formation by means of spatial analysis, spatial statistics and orientation. J Archaeol Sci 114:105067. https://doi.org/10.1016/j.jas.2019.105067
Pickering TR (2002) Reconsideration of criteria for differentiating faunal assemblages accumulated by hyenas and hominids. Int J Osteoarchaeol 12(2):127–141. https://doi.org/10.1002/oa.594
Plummer T, Bishop L (2016) Oldowan hominin behavior and ecology at Kanjera South,Kenya. J Anthropol Sci 94:29–40. https://doi.org/10.4436/JASS.94033
Plummer TW, Bishop LC, Ditchfield PW, Ferraro JV, Kingston JD, Hertel F, Braun DR (2009) The environmental context of Oldowan Hominin activities at Kanjera South, Kenya. In: Hovers E, Braun DR (eds) Interdisciplinary approaches to the Oldowan. Springer Netherlands, Dordrecht, pp 149–160
Pobiner BL, Rogers MJ, Monahan CM, Harris JWK (2008) New evidence for hominin carcass processing strategies at 1.5 Ma, Koobi Fora, Kenya. J Hum Evol 55:103–130. https://doi.org/10.1016/j.jhevol.2008.02.001
Potts R (1982) Lower Pleistocene site formation and hominid activities at Olduvai Gorge, Tanzania. Ph.D. Dissertation. Harvard University
Potts R (1988) Early hominid activities at Olduvai: foundations of human behaviour. Aldine, New York
Saladié P, Rodríguez-Hidalgo A, Domínguez-Rodrigo M, Vallverdú J, Mosquera M, Ollé A, Huguet R, Cáceres I, Arsuaga JL, Bermúdez de Castro JM, Carbonell E (2021) Dragged, lagged, or undisturbed: reassessing the autochthony of the hominin-bearing assemblages at Gran Dolina (Atapuerca, Spain). Archaeol Anthropol Sci 13:65. https://doi.org/10.1007/s12520-021-01303-6
Sánchez-Romero L, Benito-Calvo A, Marín-Arroyo AB, Agudo-Pérez L, Karampaglidis T, Rios-Garaizar J (2020) New insights for understanding spatial patterning and formation processes of the neanderthal occupation in the Amalda I cave (Gipuzkoa, Spain). Sci Rep 10:8733. https://doi.org/10.1038/s41598-020-65364-8
Schick KD (1987) Modeling the formation of Early Stone Age artifact concentrations. J Hum Evol 16:789–807
Schick KD (1997) Experimental studies of site formation processes. In: Isaac GL, Isaac B (eds) Koobi Fora Research Project. Plio-Pleistocene archaeology, vol 5. Cambridge University Press, Cambridge, pp 244–256
Walter RC, Manega PC, Hay RL (1992) Tephrochronology of Bed I, Olduvai Gorge: an application of the laser-fusion 40Ar/39Ar dating to calibrating biological and climatic change. Quat Int 13/14:37–46. https://doi.org/10.1016/1040-6182(92)90008-P
Yellen JE (1977) Archaeological approaches to the present: models for reconstructing the past. Academic, New York
Zilio L, Hammond H, Karampaglidis T, Sánchez-Romero L, Blasco R, Rivals F, Rufà A, Picin A, Chacón MG, Demuro M, Arnold LJ, Rosell J (2021) Examining neanderthal and Carnivore occupations of Teixoneres Cave (Moià, Barcelona, Spain) using archaeostratigraphic and intra-site spatial analysis. Sci Rep 11:4339. https://doi.org/10.1038/s41598-021-83741-9
Acknowledgements
We thank the Tanzanian Commission for Science and Technology (COSTECH), the Department of Antiquities, the Ngorongoro Conservation Area Authority (NCAA), and the Ministry of Natural Resources and Tourism for permission to conduct research at Olduvai Gorge. We would like to thank the three anonymous reviewers for their insightful comments and constructive feedback, which greatly enhanced the quality of this manuscript.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This study was conducted with funding from the Spanish Ministry of Science and Innovation (grant: PID2020-115452GB-C21) and the Ministry of Culture (from the program of Archaeological Projects Abroad). This study was also supported by ICArEHB – The Interdisciplinary Center for Archaeology and Evolution of Human Behaviour, funded by the Portuguese Foundation for Science and Technology (FCT) under program UIDP/04211/2020. Amanda Merino-Pelaz was funded by the Ministry of Universities of Spain (FPU grant FPU21/03533). Lucía Cobo-Sánchez was funded by the European Union ERC-CoG-2021, FINISTERRA, 101045506. Views and opinions expressed are however those of the author(s) only and do not necessarily reflect those of the European Union or the European Research Council Executive Agency. Neither the European Union nor the granting authority can be held responsible for them. Elia Organista was funded by the Swedish Research Council (Vetenskapsrådet) (AR-HS 2018-01045-3).
Author information
Authors and Affiliations
Contributions
Conceptualization: A.M-P, L.C-S; Methodology: A.M-P, L.C-S; Data acquisition: A.M-P, L.C-S, E.O., E.B., M.D-R; Formal analysis and investigation. A.M-P, L.C-S; Writing – original draft preparation: A.M-P, L.C-S; Writing – review and editing: A.M-P, L.C-S, E.O., E.B., M.D-R; Funding acquisition: E.B., M.D-R; Resources: E.B., M.D-R; Supervision: E.B., M.D-R.
Corresponding author
Ethics declarations
Ethical approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Merino-Pelaz, A., Cobo-Sánchez, L., Organista, E. et al. Unraveling the spatial imprint of hominin and carnivore accumulations in Early Pleistocene African sites. Archaeol Anthropol Sci 16, 128 (2024). https://doi.org/10.1007/s12520-024-02020-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-024-02020-6