
Abstract
Cotton cultivation is conquered by transgenic Bt upland cotton hybrids in India. Bt gene does not provide resistance against sucking insect pests. Due to the inherent vulnerability of extant Bt cotton hybrids to sap-sucking insect pests including leafhopper, upland cotton cultivation is seriously threatened by surging populations of these pests. Consistent and extensive screening of upland cotton germplasm over the years has revealed absence of adequate resistance against leafhopper. Here, we report introgression of leafhopper tolerance from a diploid A-genome cotton species, Gossypium arboreum into G. hirsutum. The dominance of leafhopper tolerance was observed over its susceptibility. Genetic analysis revealed that tolerance to leafhopper was inherited in a simple Mendelian fashion and was controlled by two genes, either singly or in combination. Using bulked segregant analysis, two simple-sequence repeat markers, namely NAU 922 and BNL 1705, located on chromosomes A5 and A11 respectively, were tagged with leafhopper tolerance. To the best of our knowledge, this is the first report of molecular tagging of leafhopper tolerance introgressed from G. arboreum into G. hirsutum. A significant negative association was observed between leaf trichome density and leafhopper nymph population.
Similar content being viewed by others


Introduction
The word 'cotton' is used to represent four cultivated cotton species namely Gossypium hirsutum L., G. barbadense L., G. arboreum L. and G. herbaceum L. It is the chief source of natural fibre for the textile industry. G. hirsutum, also known as upland cotton, is a predominantly cultivated cotton species and occupies more than 98% of the world’s cotton area. India has the maximum area (nearly 13 million hectares) under cotton cultivation which accounts for about 41% of the global cotton area (Kranthi 2021). Although, India is the leader in cotton production (6.307 million tonnes) and contributes about 25% towards global cotton production, it is not listed among the top 20 countries with the highest cotton productivity. India registered cotton productivity of 487 kg lint ha−1 during 2020 compared to the world average of 761 kg lint ha−1 (Kranthi 2021). Damage to the cotton crop by the insect pests at all the growth stages is one of the major causes of low cotton productivity in the country. Although India has distinction of growing all the four cultivated species of cotton, currently, the species profile is dominated by upland cotton due to a large scale adoption of transgenic Bt cotton. The hirsutum Bt hybrids that are grown in India are highly vulnerable to sap-sucking insects (Kranthi and Stone 2020). It has led to a shift in the pest profile status from bollworms to sap feeders (leafhopper, whitefly, cotton aphid, mealybug, mirid bug etc.) due to the suspension on the use of pyrethroids and other conventional insecticides in Bt cotton. For instance, the whitefly epidemic in 2015 destroyed cotton on nearly 1.5 million ha in north Indian cotton growing states of Punjab, Haryana and Rajasthan. Due to this pandemic, cotton productivity in Punjab reduced from a five-year (2010–2014) average of 573 kg lint ha−1 to 197 kg lint ha−1 (Kumar et al. 2020). Leafhopper, Amrasca biguttula (Ishida) (Cicadellidae: Hemiptera) is among the most economically important sucking pests of cotton after the whitefly (Ghelani et al. 2014). It has the potential of causing a 25–45% loss in seed cotton yield and may have a negative impact on fibre quality (Kalyan et al. 2017).
Leafhopper incidence is observed on cotton during the vegetative phase. The adults are small, about 3-mm long and greenish-yellow in colour during summer, and acquire a reddish tinge in the winter. During sap sucking, both nymphs and adults inject saliva into the tissues resulting in toxaemia. Tender leaves infested by leafhopper become yellow along the margins and show downward curling of leaves. Sap sucking by nymphs as well as adults results in 'hopper burn' characterized by yellowing and bronzing of leaves leading to desiccation in plants. The punctures made by leafhoppers cause shortened internodes, thus making the plant less vigorous and leads to reduced yield (Thirasack 2001). Since the use of earlier proven pesticides, namely organophosphates and pyrethroids for the management of leafhoppers became ineffective, the chemical control of jassids shifted towards the use of neonicotinoids (Ahmad et al. 1999). Seed treatment with neonicotinoids protects the cotton plants from leafhopper damage for 40–60 days. However, resistance against neonicotinoid group of insecticides has been widely reported in cotton jassid (Kshirsagar et al. 2012; Sagar and Balikai 2014; Halappa and Patil 2016; Mahalakshmi and Prasad 2020). Besides the development of resistance to insecticides, chemical control measure causes environmental pollution, adverse effects on nontarget insects, the resurgence of secondary pests and enhances the cost of cultivation. Host–plant resistance is the most attractive and viable approach for insect pest management because it is durable and eco-friendly, and is one of the key components of integrated pest management (IPM) strategies. It underscores the need to incorporate genetic resistance in upland cotton to leafhopper. Low genetic variation has been reported among the upland cotton cultivars and these are considered prone to pathogen/insect epidemics (Brubaker and Wendel 1994; Bowman et al. 1996). Consistent and extensive screening of upland cotton germplasm over the years at various agricultural universities/institutes in India has revealed the absence of adequate resistance against leafhopper. Whereas, strong resistance to leafhopper has been reported in a related A-genome diploid (2n = 26) cotton species, G. arboreum L. (Sidhu and Dhawan 1980; Ansingkar et al. 2004; Mehetre et al. 2004; Nibouche et al. 2008; Kulkarni et al. 2009). Several morphological characteristics of the plants are known to impart tolerance or susceptibility to insect pests. One such appendage is the trichome––the epidermal outgrowths present on most of the aerial parts of the cotton plant. Several experimental studies have demonstrated that leaf trichomes reduce the leafhopper population in cotton (Parnell et al. 1949; Batra and Gupta 1970; Khan and Agarwal 1984; Butler et al. 1991). Limited literature showing the association of molecular markers with sucking insect pest tolerance in cotton is available (Sankeshwar et al. 2018; Ahmed et al. 2020; Abdelraheem et al. 2021).
Therefore, the objectives of this study were to (i) introgress leafhopper tolerance from G. arboreum to G. hirsutum, (ii) to determine the genetic control of leafhopper tolerance, (iii) to identify microsatellite markers linked to leafhopper tolerance, and (iv) to test the association of leaf trichome density with leafhopper infestation.
Material and methods
Plant material
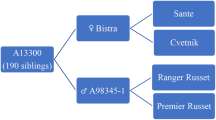
The interspecific cotton hybrid used to generate the segregating populations was developed by hybridizing G. hirsutum accession LH 2107 as the female parent and G. arboreum cv. LD 491 as the pollen parent. LD 491 is a commercial variety of desi cotton tolerant to leafhopper (figure 1a), whereas, LH 2107 (figure 1b) is a leafhopper susceptible line of upland cotton. These parental lines have been developed at the Punjab Agricultural University, Ludhiana, India. The interspecific F1 hybrid (G. hirsutum acc. LH 2107/G. arboreum cv. LD 491) was backcrossed as pollen parent with LH 2107 to develop BC1F1 (G. hirsutum acc. LH 2107/G. arboreum cv. LD 491/G. hirsutum acc. LH 2107) population (designated as population I). Similarly, this interspecific hybrid was also crossed as a male parent with an upland cotton variety F 2164 to develop another population. For genetic analysis, this population (designated as population II) was also treated as backcross generation as F 2164 (figure 1c) like LH 2107 is vulnerable to leafhopper.

Leafhopper tolerant G. arboreum cv. LD 491 (a); G. hirsutum acc. LH 2107 (b) and G. hirsutum cv. F 2164 (c) showing symptoms of leafhopper damage.
Phenotyping for leafhopper response
Jassid injury grade (JIG) was recorded as per the guidelines of ICAR-All India Coordinated Research Project on Cotton (2020-21) (www.cicr.org.in/aicrp-2021.htm): the entire foliage free of curling and yellowing (JIG I); curling of a few leaves in the lower portion of plant + marginal yellowing (JIG II); curling of leaves almost all over the plant (JIG III); and extreme curling, bronzing and drying of leaves (JIG IV). A total of 293 and 190 plants were assessed for leafhopper response in populations I and II, respectively. The plants exhibiting JIG I and II were considered leafhopper tolerant, whereas plants showing JIG III and IV were categorized as susceptible. No insecticide treatment was given to the cotton seeds before sowing. Similarly, no chemical control was undertaken for leafhopper control throughout the crop season. The number of leafhopper tolerant and susceptible plants was counted in backcross generations and the χ2 test was used to determine the number of genes governing the leafhopper tolerance. Data on nymph population were recorded on three leaves per plant, one each in the top, middle and lower canopy. The nymphal population was recorded thrice until 90 days after sowing (DAS) and data recorded during peak infestation were used for analysis.
Molecular analysis
Total genomic DNA was extracted from tender leaves following cetyl trimethyl ammonium bromide (CTAB) method (Saghai-Maroof et al. 1984). The quality and quantity of DNA was assessed on a nano-drop spectrophotometer and 0.8% agarose gel. Genomewide cotton-specific simple-sequence repeat (SSR) / microsatellite markers were used for molecular analysis. Amplification of specific DNA fragments was carried out in vitro through polymerase chain reaction (PCR) in the 96-well Applied Biosystems thermocycler. PCR reaction mixture of 12 μL consisted of 3 μL (60 ng) template DNA, 5 μL master mix (2\(\times\) premix), 2 μL autoclaved water and 1 μL of each forward (0.5 μM) and reverse primer (0.5 μM). Thermal profile for PCR amplification was as follows: initial denaturation at 94°C for 5 min followed by 30 cycles of denaturation at 94°C for 1 min; annealing at 52–58°C (depending on the primer pair) for 1 min and extension at 72°C; final extension at 72°C for 7 min. 2.5% agarose gel prepared in 0.5 \(\times\) TBE buffer was used for the resolution of amplified PCR products.
Bulked segregant analysis (BSA) proposed by Michelmore et al. (1991) was used to identify SSR markers associated with leafhopper tolerance. For this, an equal quantity of DNA from 10 BC1F1 (G. hirsutum acc. LH 2107/G. arboreum cv. LD 491/G. hirsutum acc. LH 2107) plants having JIG I was pooled to constitute the tolerant bulk. Similarly, susceptible bulk was created by pooling DNA from 10 plants with JIG IV. These bulks were genotyped with SSR markers polymorphic between the parental lines.
Measurement of trichome density
Data on leaf trichome density were recorded on parental lines and individual plants of the backcross population following Wright et al. (1999). Briefly, a young leaf trichome count was made on the underside of fully expanded but glossy leaves using a 6-mm ring (28.27 mm2). Similarly, fully expanded (but not glossy) leaves were used for trichome count on mature leaves. A total of three counts (one each on the left and right side of the midrib above the convergence of two large veins and one on the midrib) were made on each leaf using a stereomicroscope. An average of three counts was computed and used for statistical analysis. The Shapiro–Wilk test was conducted to assess the normal distribution of trichome density. Pearson’s correlation coefficient was used for establishing an association between various parameters.
Results and discussion
Introgression for leafhopper tolerance and its inheritance
G. arboreum, one of the two diploid cultivated Asiatic cotton species, belongs to secondary gene pool of cotton. It possesses resistance to several biotic stresses such as leafhopper, whitefly, thrip, cotton leaf curl disease etc. G. arboreum is cross-incompatible with upland cotton, and hybridization between these two species is relatively difficult. In the present study, several upland cotton lines, including LH 2107 were used as female parents and many desi cotton lines including LD 491 as pollen parents for the development of primary cross. A total of 12,392 flowers of upland cotton genotypes were pollinated to obtain four true interspecific (G. hirsutum × G. arboreum) hybrids. Nature of gene action and genetic control of a trait are important considerations for designing strategies to incorporate a trait successfully. The F1 hybrid derived from G. hirsutum acc. LH 2107/G. arboreum cv. LD 491 was observed to be tolerant to leafhopper indicating the dominant nature of leafhopper tolerance. This finding is consistent with the results of Sikka and Singh (1953); Annapan (1960); Pushpam and Raveendran (2005) in cotton and Sharma and Gill (1984) in okra where dominant expression of leafhopper tolerance has been reported.
In the present investigation, two populations, namely G. hirsutum acc. LH 2107/G. arboreum cv. LD 491/G. hirsutum acc. LH 2107 (population I) and G. hirsutum acc. LH 2107/ G. arboreum cv. LD 491/G. hirsutum cv. F 2164 (population II) were used to study the inheritance of leafhopper tolerance. Of the 293 plants in population I, 218 plants were found to be tolerant to leafhopper whereas, 75 plants were observed to be susceptible. Similarly in population II, 142 leafhopper tolerant and 48 susceptible plants were registered. According to the two gene model, the number of leafhopper tolerant and susceptible plants should segregate in a Mendelian ratio of 3:1 in BC1F1 generation. Chi-square analysis revealed that the observed and expected numbers did not differ significantly for digenic control (table 1). Thus, it is apparent that two dominant genes, either singly or in combination governed tolerance to leafhoppers in both populations. Similar duplicate gene action has been observed in F2 intervarietal upland cotton population segregating for leafhopper resistance in 15:1 ratio (Pushpam and Raveendran 2005). Further, using other populations they also showed leafhopper resistance to be under the monogenic (3 resistant: 1 susceptible) and digenic (13 resistant : 3 susceptible; 9 resistant : 7 susceptible) control in upland cotton. Two gene control of leafhopper resistance has also been reported by Sikka and Singh (1953) in cotton. Simple monogenic dominant genetic control for leafhopper resistance has been demonstrated by Painter (1958) and Sikka and Singh (1953). On the other hand, Roy et al. (2017) observed inhibitory gene action (13 susceptible: 3 resistant) for resistance to leafhopper in an F2 population derived from G. hirsutum × G. barbadense cross. The foregoing discussion reveals that tolerance to leafhopper in cotton is controlled by major genes but the gene action varies with different genetic backgrounds.
Identification of molecular markers linked to leafhopper tolerance
DNA-based markers have a variety of applications in crop improvement programmes including gene tagging. Gene tagging is the pre-requisite for marker-assisted selection (MAS) in which selection of a plant in the segregating/backcross generation is based on its genotype and not on its phenotype. It enhances the efficiency of a breeding programme by advancing breeding material across generations in a shorter time. For this purpose, a set of 304 cotton- specific SSR markers belonging to all the 13 A-genome chromosomes was used to discriminate the parental lines, namely G. hirsutum acc. LH 2107 and G. arboreum cv. LD 491. SSR markers have several merits as these are PCR based, co-dominant, highly reproducible, distributed throughout the genome, and follow Mendelian inheritance. Of the 304 SSR markers employed to detect polymorphism between the parental lines, 99 (32.6%) were observed to be polymorphic.
BSA was conducted to identify the SSR markers associated with leafhopper tolerance. Accordingly, DNA of 10 BC1F1 plants showing Jassid Injury Grade I was pooled to constitute 'tolerant' bulk in population I. Similarly, 'susceptible' bulk was created by pooling DNA of an equal number of BC1F1 plants exhibiting JIG IV symptoms. The 99 polymorphic SSR markers were genotyped on the leafhopper 'tolerant' and 'susceptible' bulks so as to identify the uncommon markers between the two bulks. Two SSR markers, namely NAU 922 (chromosome A5) and BNL 1705 (chromosome A11) were associated with leafhopper tolerance (figure 2). The marker genotype of the tolerant bulk resembled the leafhopper tolerant parent LD 491. Likewise, the genotype of the susceptible bulk was identical to that of the leafhopper susceptible parent, i.e. LH 2107 for both the markers (BNL 1705 and NAU 922). Interestingly, a correspondence between the genetic control and molecular analysis was observed. Both SSR markers are unlinked since both belong to two different chromosomal locations.

Gel picture showing association of SSR markers (a) NAU 0922 and (b) BNL 1705 with leafhopper tolerance; P1, G. arboreum cv. LD 491; P2, G. hirsutum acc. LH 2107; B1, Leafhopper tolerant bulk; B2, Leafhopper susceptible bulk; M, 50-bp DNA ladder.
Not many reports showing the association of molecular markers with sucking insect-pests including leafhopper tolerance are available in cotton. Recently, a QTL linked with jassid injury resistance mapped on chromosome A12 has been identified in a RIL population derived from G. hirsutum × G. barbadense cross (Sankseshwar et al. 2018). The genes governing leafhopper tolerance identified in the present study are located on chromosomes A5 and A11 and are different from Sankseshwar et al. (2018) in terms of origin and chromosomal location. The two genes identified in our study have been introgressed from a related diploid G. arboreum.
Association of leaf trichome density with leafhopper tolerance
Various studies have explored the contribution of trichomes (epidermal outgrowths on aerial plant parts) towards tolerance/susceptibility to insect pests in cotton. Trichomes alter host preference of the insects by affecting their feeding or oviposition behaviour. Higher leaf trichome density has been shown to influence the feeding and reproduction of jassid (Murugesan and Kavitha 2010; Kanher et al. 2016). On the other hand, a positive association of leaf trichome density with the whitefly population has been reported in cotton (Nawab et al. 2011; Grover et al. 2016; Suthar et al. 2021). Similarly, a significant positive correlation between the hair density with number of eggs laid down and the larval population of spotted bollworm (Earias vitella) in Asiatic cotton has also been documented (Keshav et al. 2013).
In the present investigation, leaf trichome density was assessed on one young and one mature leaf of each of 200 BC1F1 individuals in population I and five plants of each parental line. The mean and range of leaf trichome density recorded on parental lines and the backcross population are listed in table 2. Trichome density on the young and mature leaves was found to be 194 and 82.8, respectively, in LD 491. LH 2107 registered trichome density of 91 and 49 on young and mature leaves, respectively. Higher trichome density was present on young leaves in parents as well as in the backcross population compared to that of mature leaves. Similar findings on trichome density have been reported by Wright et al. (1999); Nawab et al. (2011); Turley and Vaughn (2012); Grover et al. (2016); Suthar et al. (2021) in cotton. However, Desai et al. (2008) had recorded a slightly less trichome number on young leaves in the F2 population derived from an interspecific cross of G. arboreum × G. herbaceum. The P-value of 0.0095 obtained in the Shapiro–Wilk test indicated that the distribution of leaf trichomes deviated significantly from the normal distribution in the BC1F1 population. These results are consistent with the findings of Grover et al. (2016) and Desai et al. (2008) in desi cotton. A significant positive association (r = 0.81*) of trichome density on young and mature leaves was observed in the present study. Similar to this observation, a prior study by Grover et al. (2016) in desi cotton also correlates young and mature leaf trichome density positively (r = 0.83*). A higher number of nymphs was recorded on American cotton parent LH 2107 (6.9) in comparison to desi cotton parent LD 491 (2.6). Nymph count ranged from 2.1 to 8.1 with a mean value of 5.12 in the backcross population I. A significant negative correlation (r = −0.39*) between nymph count and overall leaf trichome density was observed in population I. Similarly, a significant negative association of nymph count with young (r = −0.36*) and mature leaf trichome density (r = −0.37*) was recorded in the backcross population in the present investigation. Thus, it is evident that higher trichome density on the cotton leaves is unfavourable to leafhopper incidence. Similar reports of the negative association of trichome density with the jassid population are available in cotton (Ashfaq et al. 2010; Murugesan and Kavitha 2010; Khalil et al. 2017; Khan et al. 2017).
Presently, cotton cultivation in India is dominated by transgenic Bt upland cotton hybrids. It is important to note that the Bt gene does not provide resistance against sucking insect pests. Due to the inherent vulnerability of extant Bt cotton hybrids to sap-sucking insect pests, upland cotton cultivation is seriously threatened by the surging populations of these pests. Farmers have to pay out more on pesticides now than before the introduction of Bt cotton (Kranthi and Stone 2020). We successfully transferred leafhopper tolerance from a related diploid cotton G. arboreum into G. hirsutum, the most widely cultivated cotton species worldwide. Transfer of tolerance to sucking pests from desi cotton to tetraploid cotton has also been reported earlier (Kulkarni 2002; Ansingkar et al. 2004). However, we unambiguously demonstrate here that leafhopper tolerance introgressed from desi cotton to upland cotton is inherited in a simple Mendelian fashion and is controlled by two genes, either singly or in combination. To the best of our knowledge, it is the first report of molecular tagging of leafhopper tolerance introgressed from G. arboreum into G. hirsutum. The results reported here set the stage for mapping and the use of these genes in breeding new upland cotton cultivars with leafhopper tolerance for sustained cotton production and productivity.
References
Abdelraheem A., Kuraparthy V., Hinze L., Stelly D., Wedegaertner T. and Zhang J. 2021 Genome-wide association study for tolerance to drought and salt tolerance and resistance to thrips at the seedling growth stage in US Upland cotton. Ind. Crop. Prod. 169, 113645.
Ahmad M., Arif M. I. and Ahmad Z. 1999 Detection of resistance to pyrethroids in field populations of cotton jassid (Homoptera: Cicadellice). J. Econ. Entomol. 92, 1246–1250.
Ahmed H., Nazir M. F., Pan Z., Gong W., Iqbal M. S., He S. et al. 2020 Genotyping by sequencing revealed QTL hotspots for trichome-based plant defense in Gossypium hirsutum. Genes 11, 368.
Annapan R. S. 1960 Breeding for jassid resistance - new findings. Indian Cott. Grow. Rev. 14, 501–508.
Ansingkar A. S., Kadke P. P., Borikar S. T. and Bhosle S. S. 2004 Altering G. hirsutum cotton at cellular level to impart multiple sucking pest resistance through interspecific hybridization. In Proceedings of International symposium on “Strategies for Sustainable Cotton Production- A Global Vision (ed. B. M. Khadi, I. S. Katageri, S. S. Patil, H. M. Vamadevaiah, B. R. Patil and S. M. Manjula), pp. 101–103. University of Agricultural Sciences, Dharwad, India.
Ashfaq M., Ane M. N., Zia K., Nasreen A. and Hasan M. 2010 The correlation of abiotic factors and physicomorphic characteristics of (Bacillus thuringiensis) Bt transgenic cotton with whitefly, Bemisia tabaci (Homoptera: Aleyrodidae) and jassid, Amrasca devastans (Homoptera: Jassidae) populations. Afr. J. Agri. Res. 5, 3102–3107.
Batra G. R. and Gupta D. S. 1970 Screening of varieties of cotton for resistance to Jassid. Cot. Grow. Rev. 47, 285–291.
Bowman D. T., May O. L. and Calhoun D. S. 1996 Genetic base of Upland cotton cultivars released between 1970 and 1990. Crop Sci. 36, 577–581.
Brubaker C. L. and Wendel J. F. 1994 Reevaluating the origin of domesticated cotton (Gossypium hirsutum; Malvaceae) using nuclear restriction fragment length polymorphisms (RFLPs). Am. J. Bot. 81, 1309–1326.
Butler J. G. D., Wilson F. D. and Fishler G. 1991 Cotton leaf trichomes and populations of Empoasca lybica and Bemisia tabaci. Crop Prot. 10, 461–464.
Desai A., Chee P. W., May O. L. and Paterson A. H. 2008 Correspondence of trichome mutations in diploid and tetraploid cottons. J. Hered. 99, 182–186.
Ghelani M. K., Kabaria B. B. and Chhodavadia S. K. 2014 Field efficacy of various insecticides against major sucking pests of Bt cotton. J. Biopestic. 7, 27–32.
Grover G., Kaur B., Pathak D. and Kumar V. 2016 Genetic variation for leaf trichome density and its association with sucking insect-pests incidence in Asiatic cotton. Indian J. Genet. 76, 365–368.
Harappa and Patil 2016 Detoxifying enzyme studies on cotton leafhopper, Amrasca biguttula biguttula (Ishida), resistance to neonicotinoid insecticides in field populations in Karnataka, India. J. Plant Prot. Res. 56, 346–352.
Kalyan R. K., Saini D. P., Meena B. M., Pareek A., Naruka P., Verma S. et al. 2017 Evaluation of new molecules against jassids and white flies of Bt cotton. J. Entomol. Zool. Stud. 5, 236–240.
Kanher F. M., Syed T. S., Abro G. H., Jahangir T. M. and Tunio S. A. 2016 Some physio-morphological leaf characters of gamma irradiated cotton lines to resistance against Jassid (Amrasca devastans Dist.). J. Entomol. Zool. Stud. 4, 80–85.
Keshav A., Shera P. S. and Singh J. 2013 Morphological basis of resistance to spotted bollworm, Earias vittella (Fabricius) in Asiatic cotton. Phytoparasitica 41, 235–240.
Khalil H., Raza A. B. M., Afzal M., Aqueel M. A., Khalil M. S. and Mansoor M. M. 2017 Effects of plant morphology on the incidence of sucking insect pests complex in few genotypes of cotton. J. Saudi Soc. Agric. Sci. 16, 344–349.
Khan Z. R. and Agarwal R. A. 1984 Ovipositional preference of jassid, Amrasca biguttula biguttula Ishida on cotton. J. Entomol. Res. 8, 78–80.
Khan M. M., Ali A., Nadeem I., Khan M. Y. and Muhammad F. A. R. 2017 Morphological and biochemical Bt cotton plant trait induced resistance against cotton jassid Amrasca biguttula biguttula (Ishida) and cotton whitefly Bemisia tabaci (Genn.). J. Innov. Bio. Res. 1, 36–45.
Kranthi K. R. 2021 ICAC cotton data book 2021, International Cotton Advisory Committee, Washington.
Kranthi K. R. and Stone G. D. 2020 Long-term impacts of Bt cotton in India. Nat. Plants 6, 188–196.
Kshirsagar S. D., Satpute N. S. and Moharil M. P. 2012 Monitoring of insecticide resistance in cotton leafhoppers, Amrasca biguttula biguttula (Ishida). Ann. Plant Prot. Sci. 20, 283–286.
Kulkarni V. N. 2002 Hirsutization of G. arboreum cotton and genetic emendation of G. hirsutum for sucking pest resistance. Ph.D. thesis, University of Agricultural Sciences Dharwad, India.
Kulkarni V. N., Khadi B. M., Maralappanavar M. S., Deshapande L. A. and Narayanan S. S. 2009 The worldwide gene pools of Gossypium arboreum L. and G. herbaceum L., and their improvement. In Genetics and genomics of cotton (ed. A. H. Paterson), pp. 69–97. Springer, New York.
Kumar V., Kular J. S., Kumar R., Sidhu S. S. and Chhuneja P. K. 2020 Integrated whitefly [Bemisia tabaci (Gennadius)] management in Bt-cotton in North India: An agroecosystem-wide community-based approach. Curr. Sci. 119, 618–624.
Mahalakshmi M. S. and Prasad N. V. V. S. D. 2020 Insecticide resistance in field population of cotton leaf hopper, Amrasca devastans (Dist.) in Guntur, Andhra Pradesh, India. Int. J. Curr. Microbiol. App. Sci. 9, 3006–3011.
Mehetre S. S., Gomes M., Susan E., Aher A. R. and Shinde G. C. 2004 RAPD and cytomorphological analyses of F1, F2 and amphidiploid (A1) generations of Gossypium arboreum × Gossypium capitis-viridis. Cytologia 69, 367–379.
Michelmore R. W., Paran I. and Kesseli R. V. 1991 Identification of markers linked to disease resistance genes by bulk segregant analysis: a rapid method to detect markers in specific genomic regions using segregating populations. Proc. Natl. Acad. Sci. USA 88, 9828–9832.
Murugesan N. and Kavitha A. 2010 Host plant resistance in cotton accessions to the leafhopper Amrasca devastans (Distant). J. Biopest. 3, 526–533.
Nawab N. N., Khan I. A., Khan A. A. and Amjad M. 2011 Characterization and inheritance of cotton leaf pubescence. Pak. J. Bot. 43, 649–658.
Nibouche S., Brevault T., Klassou C., Dessauw D. and Hau B. 2008 Assessment of the resistance of cotton germplasm (Gossypium spp.) to aphids (Homoptera, Aphididae) and leafhoppers (Homoptera: Cicadellidae, Typhlocybinae): methodology and genetic variability. Plant Breed. 127, 376–382.
Painter R. H. 1958 Resistance of plants to insects. Annu. Rev. Entomol. 3, 267–290.
Parnell F. R., King H. E. and Ruston D. F. 1949 Jassid resistance and hairiness of the cotton plant. Bull. Entomol. Res. 39, 539–575.
Pushpam R. and Raveendran T. S. 2005 Artificial screening and inheritance studies on resistance to jassid (Amrasca devastans) in Gossypium hirsutum L. Indian J. Genet. 65, 199–201.
Roy U., Patil R. S., Katageri I. S., Patil S. B. and Udikeri S. S. 2017 Genetic studies on resistance to jassid (Amrasca biguttula biguttula) in segregating F2 population of interspecific (Gossypium hirsutum L. × Gossypium barbadense L.) cotton. Int. J. Curr. Microbiol. App. Sci. 6, 2683–2686.
Sagar D. and Balikai R. A. 2014 Insecticide resistance in cotton leafhopper, Amrasca biguttula biguttula (Ishida) – a review. Biochem. Cell. Arch. 14, 283–294.
Saghai-Maroof M. A., Soliman K. M., Jorgensen R. A. and Allard R. W. 1984 Ribosomal DNA spacer- length polymorphism in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc. Natl. Acad. Sci. USA 81, 8014–8018.
Sankeshwar M., Jadhav M., Adiger S., Patil R. S. and Katageri I. 2018 Mapping of QTLs for traits related to leaf pubescence, jassid resistance and yield in cotton (Gossypium spp.). Indian J. Genet. 78, 252–260.
Sharma B. R. and Gill B. S. 1984 Genetics of resistance to cotton jassid, Amrasca biguttula biguttula (Ishida) in okra. Euphytica 33, 215–220.
Sidhu A. S. and Dhawan A. K. 1980 Incidence of some insect pests on different varieties of cotton. J. Res. Punjab Agric. Univ. 17, 152–156.
Sikka S. M. and Singh A. 1953 Inheritance of jassid (Empoasca devastans Dist) resistance in some upland American cottons. Indian Cott. Grow. Rev. 7, 113–117.
Suthar T., Gupta N., Pathak D., Sharma S. and Rathore P. 2021 Morpho-anatomical characterization of interspecific derivatives of Gossypium hirsutum L. × G. armourianum Kearney cross for whitefly tolerance. Phytoparasitica, https://doi.org/10.1007/s12600-021-00963-3.
Thirasack S. 2001 Yield losses assessment due to pests on cotton in Lao PDR. Agric. Nat. Resour. 35, 271–283.
Turley R. B. and Vaughn K. C. 2012 Molecular biology and physiology differential expression of trichomes on the leaves of Upland cotton (Gossypium hirsutum L.). J. Cotton Sci. 16, 53–71.
Wright R. J., Thaxton P. M., El-Zik K. M. and Paterson A. H. 1999 Molecular mapping of genes affecting pubescence of cotton. J. Hered. 90, 215–219.
Acknowledgements
The interspecific hybrids and backcross populations used in the present study were developed under the Programme Support on ‘Enhancing Durability of Resistance to Biotic Stresses in Selected Cereal and Fiber Crops through Biotechnological Approaches (BT/01/CE1B/121/01)’ funded by the Department of Biotechnology, Government of India. Ministry of Science and Technology (Grant No. 102/IFD/SAN/1307/2014-15).
Author information
Authors and Affiliations
Corresponding author
Additional information
Corresponding editor: Shrish Tiwari
Rights and permissions
About this article

Cite this article
Jindal, S., Pathak, D., Pandher, S. et al. Inheritance and molecular tagging of genes introgressed from Gossypium arboreum to G. hirsutum for leafhopper tolerance. J Genet 101, 42 (2022). https://doi.org/10.1007/s12041-022-01379-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12041-022-01379-6