
Abstract
During the analysis of suspensions of PI-stained nuclei from stem, adventitious roots, and leaves of in vitro germinated juvenile D. majalis plants, a range of one to three consecutive rounds of strict partial genome duplications could be discovered. Repeated cycles of under-replications were found again when axenically grown material was moved from solid nutrient media to potting soil, where it further maturated within and outside the laboratory. On average, 83–84% of the 2C DNA content participated in partial genome duplication processes while the remaining 16–17% were untouched. Different frequency distributions were detected for the affected nuclei, depending on the individual, the plant organ, and the ontogenetic stage. The above data were in line with the signs of endomitosis, as shown by acetocarmine-stained smear preparations from the lower epidermis of leaves and from adventitious roots sampled at some distance above the meristematic tip. Endomitosis is one known pathway of endoreplications, which leads to an increased DNA content in non-dividing somatic nuclei. Using a collection of young anthers in the microspore stage, where the regular haplophasic number of 1n = 2x = 40 chromosomes could be counted, foliar cells of the related vegetative plant body were considered to contain the whole set of 2n = 4x = 80 chromosomes that contributed to a mean mass of 14.71 pg DNA per nucleus at the 2C genome level. The diverse biological source materials and investigated ontogenetic stages as well as the different nuclear DNA amount calculations assessed in this study are of instructional value to avoid erroneous estimations of the present nuclear DNA amount per weight using flow cytometry. This is important with respect to the orchid genus Dactylorhiza, for example, when trying to achieve a full understanding of genome size variation in established polyploids and when searching for nascent polyploids in the future.
Similar content being viewed by others
Introduction
Although nucleus and cytoplasm do no more divide, somatic cells of many plant organisms preserve an opportunity to synthesize DNA when extending in volume and differentiating in biological characters (Joubès and Chevalier 2000). This phenomenon, termed endoreplication, leads to an increasing number of DNA molecules and makes nuclei therefore endopolyploid, as reviewed in Edgar et al. (2014). By contrast, the regular cell cycle, passing cells through somatic divisions, executes one round of DNA synthesis and then partitions replicated sister chromatids of every chromosome equally into daughter cells, so that no changes in ploidy occur. The regular cell cycle starts with the S phase (synthesis of DNA) and concludes with the M phase (mitosis). In the latter, nuclear division (karyokinesis) and cytoplasmic division (cytokinesis) arise. Intervening gap phases allow the cells to grow and represent checkpoints, namely G1 and G2, which separate the M phase from the S phase and the S phase from the M phase, respectively (Alberts et al. 2015). When plant cells escape from proper mitosis to run under the guidance of the endocycle, which is one out of two known pathways responsible for endoreplications (Breuer et al. 2014), DNA duplication happens without producing visible complexes between DNA and the proteins that make up the chromosome structure. Remarkable exceptions are gigantic polytene chromosomes in Phaseolus vulgaris L. that lump together thousands of sister chromatids, due to an extraordinarily high number of iterated endoreplications and the lack of sister-chromatid segregation (W Nagl in Von Denffer et al. 1978). In the last two decades, the occurrence of polytenic chromosomes could be detected even with low levels of polyploidy thanks to the application of fluorescence in situ hybridization (Bourdon et al. 2012). Interestingly, endocycle progression and mitosis share interrelated protein factors (Edgar et al. 2014). Initial stages of mitosis (e.g. higher-ordered structures of DNA packaging in the chromatin fibers) have been observed during the other known endoreplication pathway, called endomitosis, where mononucleate cells with low levels of endopolyploidy arise because of an arrested karyokinesis and absence of cytokinesis (see e.g. Kausch and Horner 1984).
If chromosome counts are not available to recognize endopolyploid cells, flow cytometry (FCM) overcomes this problem by preparing nuclear suspensions from a chosen tissue, as reviewed in Doležel et al. (2007). After staining liberated nuclei with a DNA-specific fluorochrome and its following excitation, light emissions are received one at a time when the nuclei pass in a fluid stream through the FCM equipment (see Lister 1990 for technical details). The amount of fluorescence light, emitted by each labelled nucleus at the observation point, is proportional to the total mass of DNA and, therefore, reflects the current position in the regular cell cycle. A histogram, displaying the frequency distribution of fluorescence intensities among all nuclei sampled from a tissue, hence provides two distinct peaks, which have been first described by Heller (1973): nuclei at the G1 phase, prior to DNA replication, form a peak that represents the 2C DNA content, where C refers to the DNA content of an unreplicated haplophasic chromosome complement (n) (see Greilhuber et al. 2005 for a terminology review); nuclei within the G2 phase as well as the M phase, after DNA replication, contribute to a joined 4C band at twice the intensity of the 2C peak. It is of note that diploid sexually reproducing eukaryotic organisms own each chromosome as a member of a pair; one chromosome is inherited from the mother and the other one is derived from the father, so that the 1C status only appears when the members of a chromosome pair segregate equally into the meiotic products during mating of germinal cells. Note furthermore that the presence of both the 2C and the 4C peak in non-dividing cells marks a first cycle of endoreplication. Consecutive rounds of endoreplications, involving not the whole 2C DNA content and providing more than two bands, represent repetitious cycles of under-replications. Within the plant kingdom, under-replications are exclusively connected to the orchid family (Trávniček et al. 2015; Hřibová et al. 2016). Because a small genomic part remains unaffected, fluorescence peaks above the 2C status can be detected at positions that disagree with complete genome replications, as will be explained later within the 'Material and methods' section. Certain orchid species, however, perform conventional whole-genome endoreplications as shown by the seminal work of Trávniček et al. (2015).
The Western European orchid Dactylorhiza majalis (Reichenbach) P.F. Hunt and Summerhayes (1965), naturally occurring in marshy meadows, is known to be an ancient allotetraploid species that contains 2n = 4x = 80 small circular chromosomes within each vegetative nucleus (4x refers to four somatic sets of chromosomes because the allotetraploid species unifies genetic information from two diploid parental species). Besides well-marked morphological characters and chromosome counts, the allotetraploid origin has been furthermore supported by genetic inventories with allozymes (Hedrén 1996) and diverse DNA markers (Devos et al. 2006). In a recent investigation, we detected extremely high rates of nonviable embryos at 12 D. majalis stands of the Upper Lusatia region in Saxony, Germany. The low seed fertility was attributed to an improper meiosis, which has been confirmed by aneuploid chromosome numbers in developing germinal cells (Schubert et al. 2020). These survey data motivated us to apply FCM-based nuclear DNA mass estimations to aid a fast identification of aneuploid offspring at the population level. As an alternative to plants collected from the wild, indoor cultivation of abundant individuals provides an attractive experimental system to study nucleic acid replication, including DNA content analyses, because D. majalis is endangered by habitat disturbance and therefore protected by law in Germany (e.g. Bräutigam and Otto 2012; Rode and Schadwinkel 2014). Samples are therefore not readily obtainable from native stands. Most importantly, seed embryos of orchid species are known to exist in the absence of a cotyledon and are moreover not surrounded by a nutritionally rich endosperm tissue. In natural environments, they therefore form upon germination a unique protocorm body to obtain nutrition from an associated living mycorrhizal fungus (Yeung 2017). Instead of establishing beneficial mycorrhizae, we tried to germinate immature embryos of D. majalis in axenic cultures that supply the necessary nutrients for further development, as outlined by Debergh (1994). Assessing the fostered plant material by FCM, the objectives of the study were to: (i) identify true peak positions with respect to the occurring endoreplication rounds, (ii) prove whether the whole nuclear genome or only a part of it becomes amplified, (iii) compare endoreplication patterns amongst different organs at diverse ontogenetic stages, (iv) to count the present number of chromosomes as well as to visually investigate the cellular process underlying endoreplication in D. majalis nuclei using cytological acetocarmine preparations, and (v) to estimate the nuclear DNA amount in relation to the present number of chromosomes.
Material and methods
Collecting and storing seeds
In the growing season of the year 2016, ripe capsules were collected from open-pollinated mother plants of D. majalis by sampling one natural population in Waltersdorf (Saxony, Germany) and two natural populations in Greiz (Thuringia, Germany). For each harvested mother plant, capsules and silica-gel particles were put in a bag of dry blotting paper and transferred to a refrigerator at 6 °C. Seeds of Pisum sativum L. cv. 'Ctirad', necessary to raise pea plants of known nuclear DNA mass, were received from J. Doležel (Olomouc, Czech Republic) and were stored as mentioned above.
Germinating seeds and cultivating plants
Beginning in December 2017, seeds were removed from the stored capsules of D. majalis and were subsequently transferred into a commercial sodium hypochlorite bleach (1% active chlorine by weight) to carry out surface disinfection for 6 min at room temperature. Seeds were then separately plated for each lot on solid sowing medium in lid-covered 190-ml plastic containers, which have been sterilized and delivered by the Greiner Bio-One GmbH (Frickenhausen, Germany). According to the Swedish orchid breeder S. Malmgren (personal communication), sowing medium was prepared by dissolving 10 g sucrose, 50 mg Ca3PO4, 50 mg KH2PO4 and 50 mg MgSO4 in tap-water, followed by the addition of 1 g of activated charcoal plus 5 g of agar and 50 ml of pineapple juice (fresh fruits were juiced by a food processor) to obtain a volume of 1 L. The pH was set to 6.0 with ammonia water. After autoclaving (121 °C for 20 min), sowing medium was allowed to cool to 55 °C and then 4.6 ml Vaminolact® (Fresenius, Graz, Austria) was added, which represents a ready-to-use mixture of amino acids. Sowing medium was finally dispensed in the above-mentioned containers (60 ml medium per vessel) where the agar hardened after closing lids. Because germs were cultured over a period of up to 20 months, the above sowing medium was newly made at roughly 7-months intervals, but using higher proportions of sucrose (20 g/l) and agar (15 g/l) to furnish protocorms and to convert them into seedlings with evident roots, stems and leaves. From July 2019 onwards, clumps of seedlings were transferred to solid orchid maintenance medium in capped steam-sterilized preserving jars (Weck, Wehr, Germany). From the factory-produced P668 orchid maintenance medium powder (PhytoTechnology Laboratories®, Lenexa, KS, USA), 27.31 g were mixed with 8.5 g of agar in distilled water to obtain 1-l batches. Afterwards, autoclaving (121 °C for 20 min) and dispensing in jars (200 ml medium per 580-ml vessel) were carried out. To keep the in vitro grown material for prolonged periods alive, orchid maintenance medium was supplemented with agar (8.5 g/l) in conjunction with B852 banana powder (30 g/l, PhytoTechnology Laboratories®). When, after a couple of weeks, the color of the used orchid maintenance media altered from gray to black, biological material was moved on freshly-made media with the same nutrient composition as mentioned above. All necessary transfers of disinfected seeds, living protocorms, seedlings, and plantlets were done under aseptic conditions inside a laminar air-flow cabinet. Single jar-grown orchid individuals were finally recovered from solid nutrient medium by washing their roots in running water. They were then planted for further maturation onto a special potting soil (Phytesia, Cry, Belgium). Pea plants were quickly grown from seeds in pots, using a standard garden soil provided by a local florist shop. Potting soils were watered once a week without adding a chemical fertilizer. The above-mentioned plant stock was cultured in the laboratory under the following conditions: temperature 21 °C; daylight; extra light was furnished by Lumilux® Cool White Lamps (Osram, Augsburg, Germany) for 9 to 13 h per day during winter and summer seasons, respectively. Orchid seedlings were kept cool (4 °C) by storing them in a dark refrigeration room from November 2018 until February 2019.
Preparing labelled nuclear suspensions and performing FCM
Three commercial kits were chosen from the Sysmex Partec GmbH (Görlitz, Germany) to estimate the amount of DNA in plant nuclei using FCM. The CyStain® PI OxProtect kit (protocol I) contains propidium iodide (PI). This DNA-intercalating fluorochrome emits light rays at the orange to red regions of the spectrum (λ > 590 nm) when becoming excited at 532 nm. The CyStain® UV OxProtect kit (protocol II) and the CyStain® UV Precise P kit (protocol III) employ 4',6-diamidino-2-phenylindole (DAPI) that binds to adenine as well as to thymine of the DNA, generating fluorescence at the 435- to 560-nm waveband in response to UV irradiation (λ = 365 nm). Both above-mentioned OxProtect kits were designed to liberate and label nuclei from oxidation-sensitive plant tissues in the cold, minimizing the interference of secondary metabolites. Starting genome size analyses, tissues were cut from the above actively growing D. majalis plants to dissect them into smaller pieces (1 cm2 or less) by means of scalpels. Moreover, source material was obtained from the fresh leaves of pea plants. After putting defined tissue pieces from both species together, they were collectively chopped with a razor blade to prepare filtered suspensions of free marked nuclei using the detailed protocols and necessary reagents included in the kits. Alternatively, entire seed embryos of D. majalis were ground in extraction buffer with a mortar and pestle. Since the utilized Sysmex Partec CyFlow® ploidy analyzer was equipped to stimulate PI as well as DAPI for fluorescence emission, light appeared in conjunction with the above DNA-labelled nuclei allowing their registration by photomultiplier tubes (PMT). Fluorescence peak identification and particle counts were accessed with the tools included in the CyView™ software version 1.6 (Sysmex Partec). To make the 2C as well as the 4C band of P. sativum and the corresponding peaks of D. majalis simultaneously visible on the screen, individual PMT gains were used according to the different plant materials and dying procedures. Recorded data files were finally imported to the FCS4 Express™ cytometry package (De Novo Software, Pasadena, CA, USA) for drawing the histograms presented below.
Interpreting data from FCM analyses and statistics
Fluorescence intensity (I) is expected to double when the entire nuclear genome becomes amplified during one round of endoreplication. Affected nuclei, offering a fluorescence intensity called Ik, correspond for that reason in the FCM-based histogram to a twofold increased channel position in comparison to nuclei, which own the starting DNA amount (their fluorescence intensity is called Ik-1). The ratio between the measured fluorescence intensities Ik/Ik-1 hence attains the value 2. This behavior, connected here to the reference species P. sativum, is indicated by a geometric progression of C values rising from 2 to 4C for both recorded fluorescence signals. If, however, a partial genome endoreplication occurred, then Ik/Ik-1 turns out to be lower than 2. When the 2C sample peak and the first endoreplication peak, for example, yielded I2/I1 = 1.88, this implies that 88% of the 2C DNA content participated in endoreplication. On the other hand, 12% of the 2C DNA stayed untouched. If the endoreplicated genomic parts stay constant across multiple consecutive rounds of endoreplication cycles, this is called strict partial endoreplication (Brown et al. 2017). According to the terminology given in Brown et al. (2017), the replicated part of the original 2C genome is P, whereas the unreplicated, fixed part is F. In every new strict partial endoreplication cycle, the part P of the original 2C genome, together with all pre-replicated copies of that part, undergo another round of endoreplication. Hence, for strict partial endoreplications, P and F are constant proportions of the 2C genome. With non-strict partial endoreplications, the duplicated proportions and parts of the original 2C genome may vary across consecutive endoreplication cycles (Brown et al. 2017). Having found ratios below 2 for the test species D. majalis by carrying out the above logic pair-wisely with up to four detected fluorescence bands, we called the peaks in sequence to the 2C band that were caused by iterated partial endoreplications 4E, 8E, and 16E as proposed by Brown et al. (2017). Due to the linear relationship between the present amount of DNA and the emitted fluorescence yield, it was moreover possible to determine DNA weights per nucleus for an orchid individual in question by comparing the PI-born fluorescence intensity of the 2C sample peak with that provided by an internal calibration standard. For this, pea nuclei of cv. 'Ctirad' were treated and measured in a single sample together with the orchid nuclei to be tested, as described above. To meet the requirements of a reliable data acquisition, every considered measurement employed mostly more than a total of 5000 analyzed particles, providing at least 1000 nuclei for each the sample and the reference peak used for the DNA mass calculations, as has been recommended by Loureiro et al. (2023). A notable exception, however, were the rare foliar nuclei with 2C status of the blooming plant no. B1, for which the DNA mass determination was ascertained by a rather small sum of 2000 liberated nuclei using four repeated nuclear preparation experiments and corresponding FCM runs. Pea nuclei of the cv. 'Ctirad' are known to contain 9.09 picogram (pg) DNA at the 2C level (Doležel et al. 1998), which is equivalent to 8.89 × 109 base pairs because the number of base pairs will precisely be the DNA mass in pg × 0.978 × 109 (Doležel et al. 2003). It is of note that we only used the PI-born fluorescence peak intensities for the determination of absolute DNA weights, whereas DAPI-born fluorescence data were not applied for such cases according to Doležel and Bartoš (2005). Statistics were assessed with each investigated orchid specimen by carrying aliquot sample parts through the same laboratory procedure to get mean values (ȳ) and standard deviations (sd) for the estimated nuclear DNA weights. The coefficient of variation (CV) was calculated by the formula \({\text{CV}}=[\text{sd}/\overline{y }] \times 100\text{\%}.\) This allows the comparison of data acquired from the same sampled specimen at different PMT gain settings (Lister 1990).
In addition, we calculated the obtained fluorescence intensity ratios Ik/Ik-1 in order to draw further statistical conclusions about the nature of the endoreplication process. Specifically, the mean values gathered from all relevant plant specimens were used to calculate an estimate for the P and F parts of the genome using linear regression as outlined in Brown et al. (2017) to corroborate the presence of strict partial endoreplications in D. majalis. All calculations were carried out using the software R version 4.2.2 (R Core Team 2022).
Preparing and imaging acetocarmine smears
Pieces of leaves and adventitious roots as well as young anthers were cut from the actively growing D. majalis plants. All subjects were immediately immersed into a generous quantity of killing fluid, consisting of absolute methyl alcohol and glacial acetic acid (ratio of 3:1 by volume), to process fixation for at least 1 h at −20 °C. Each fixed specimen was then soaked for 10 min at 95 °C in 200 µl acetocarmine reagent, where the cellular middle lamella was hydrolyzed and nuclear structures became stained. A stock solution was previously prepared by dissolving 1 g carmine (Thermo Fisher Scientific, Geel, Belgium) in 100 ml of boiling 45% (v/v) acetic acid, as has been thoroughly described by Sass (1958). Stained material was finally mounted on a slide in a fresh drop of acetocarmine, divided into the smallest possible linear strips with needle and scalpel, and squashed flat by gently pressing a coverslip against the glass slide. After removing excess fluid by a paper tissue, smear preparations were studied under considerable magnification with an Olympus-CX43 light microscope (Hamburg, Germany). Photomicrograph records were taken using an attached Olympus digital camera model LC30, which was operated with version 2.2 of the LC micro software package (Olympus Soft Imaging Solutions GmbH, Berlin, Germany).
Results
Seed germination and formation of plants
Having seed embryos plated on solid sowing medium, with changes of freshly prepared medium occurring at 7-months intervals and after moving the clumps of germinated seedlings to orchid maintenance medium, clusters of juvenile plantlets spread over the surface of the nutritional substratum two years later. The success of the above processing is shown for seed lot G02 (Fig. 1 a). Approximately 1 mg of seeds were sown per container, representing an average of 125 to 500 embryos, because the weight of a single D. majalis seed is known to range from 2 × 10–6 to 8 × 10–6 g in accordance with Schoenichen (1940). Thus, the roughly obtained 10 jar-grown plantlets suggest that 2 to 8% of the disinfected seeds were converted into plants. Over the following three years, when further cultivation was done on orchid maintenance medium with or without addition of banana powder, vigorous roots repeatedly sprouted at their heads to raise single annual stems anew (Fig. 1 b) that later exposed spirally arranged juvenile leaves (Fig. 1 c). Representative plantlets, which were recovered from solid maintenance medium 62 months after sowing, exhibited tall roots with two finger-like protuberances at the bottom, approximating the palmately lobed structure known from D. majalis in natural populations, and furthermore expressed thinner adventitious roots at the top (Fig. 1 d). The foregoing in vitro germination and cultivation systems on solid nutrient agar media were a source for abundant immature plantlets to examine different vegetative organs by FCM and acetocarmine smears, as outlined below. When agar-grown in vitro germinated individuals of D. majalis were transferred to potting soil, a few of them already reached a climax of differentiation in the laboratory sphere (Fig. 1 e, f). This made rare indoor subjects available for a collection of data from mature pot-bound plants, during flowering and afterwards, which will be presented below. Pot-bound biological materials that left the laboratory and then spent three additional years under natural climatic conditions at a private garden site were all in blossom in midsummer 2023 (Fig. 1 g).

Photographs, showing examples of plant regeneration from in vitro germinated seed embryos of D. majalis at different time points of cultivation (MAS – months after sowing). a. Nested juvenile green plantlets (seed lot G02), occupying maintenance agar medium, 24 MAS, to try out further cultivation; b. Rooted shoot, recovered from maintenance agar medium, 38 MAS; c. Single juvenile green plantlet, grown on maintenance agar medium, 37 MAS; d. Plantlets, removed from maintenance agar medium, 62 MAS, exhibiting finger-like protuberances at the bottom of the roots and expressing juvenile leaves as well as adventitious roots at the top in vertical and in horizontal positions, respectively; e. Mature pot-grown indoor plant in blossom, 37 MAS; f. Dormant pot-grown indoor plant, 44 MAS; g. Mature pot-grown outdoor plants in blossom, originating from in vitro germinated laboratory materials that were moved to a private garden site, where they became further cultivated for three years under natural climatic conditions
Flow cytometry
When PI-labelled nuclear preparations from in vitro germinated D. majalis were analyzed, a rooted stem with visible leaf primordia (Fig. 1 b) was first sampled. After removing this stem from the solid maintenance medium, with banana powder included, a piece shortly above the ground was chosen for FCM. Four fluorescence peaks were found at positions 2C, 4E, 8E, and 16E (Fig. 2 a). This pattern confirms three repeated rounds of partial genome duplications, where the original 2C DNA content became enlarged 1.88-fold (I2/I1 = 1.88) during the first cycle, the produced 4E DNA content was amplified 1.92-fold (I3/I2 = 1.92) during the second cycle, and the obtained 8E DNA content was extended 1.92-fold (I4/I3 = 1.92) during the third cycle. According to Table 1, about 30 to 36.36% of the counted nuclei were found in association with the 8E and the 4E band, respectively. Nuclei forming the 16E as well as the 2C band were detected with distinctly lower relative numbers of about 14 to 19.15%. The 2C band as well as the 4C band of co-prepared pea nuclei appeared in addition to the four peaks that were provided by D. majalis. This held true for all the other PI-born histograms presented below, where P. sativum served as an internal calibration standard; it allowed to determine the weights of nucleic acids in some orchid samples, as listed in Table 1 at the rightmost column, by comparing the fluorescence yield of the 2C band with that of the pea-leaf 2C standard band. Turning to nuclear preparations done with juvenile green leaves of in vitro germinated D. majalis, PI-born fluorescence signals at both 2C and 4E positions, and in one case even at 8E position were observed (Fig. 2 b–d). For this purpose, immature test individuals were employed that grew-up on orchid maintenance medium, as exemplarily depicted in Fig. 1 c. Foliar nuclei at 2C, 4E as well as at 8E peak positions were detected again with a blooming pot-bound plant (Fig. 2 e), while the last green leaf of a dormant pot-bound individual, after flowering, revealed only foliar nuclei at 2C and 4E peak positions (Fig. 2 f). Both in vitro germinated test plants described above were initially cultured on maintenance medium and were afterwards grown on potted soil (Fig. 1 e–f) until FCM was carried out. Their fully differentiated green leaves are characterized, in contrast to the preceding immature leaf samples, by an excessive brownish pigmentation that is well known from plants in natural habitats. Nuclei, which were investigated at separate stages of leaf differentiation with the above-mentioned test plants and the PI-fluorochrome, moreover indicated that the original 2C DNA content expanded in foliar cells 1.8-fold to 1.9-fold throughout a first round of partial genome duplication. A consecutive second round of partial genome duplication could be detected for two out of five individuals, increasing the yielded 4E DNA content 1.89-fold to 1.91-fold. These results are reflected by the ratios I2/I1 and I3/I2 given for plants symbolized by A3, B2, B1, D, and E1 in Table 1 (rows 3–7). It is furthermore noteworthy that during the whole ontogenesis the predominant fraction of foliar nuclei provided a 4E peak, whereas the frequency of foliar 2C cells was remarkably low at the time of flowering. This is demonstrated by the fact that 8.49 to 9.84% of the foliar nuclei signaled a 2C status when the test plant was in blossom. By contrast, 17.83 to 33.84% of the foliar nuclei represented a 2C status, when individuals were sampled prior to the flowering process or when a plant was approached after blooming. The very small fraction of foliar 2C cells, identified during the flowering process, was proven for two samples of plant individual B1 by labelling one batch with PI while the other one was labelled with DAPI (compare Fig. 2 e and Fig. 2 g as well as data presented in Table 1, row 6). When adventitious roots of in vitro germinated individuals, growing on maintenance medium, were hereafter sampled at some distance above the meristematic tip, the PI-born 2C band was analytically untraceable, as exemplarily shown for plant no. A3 (Fig. 2 h). Instead of providing a 2C peak, 30.22% of the labelled root nuclei revealed a signal at 4E position (see Table 1, row 4) that resulted from an initial round of partial genome duplication. The remaining detectable peak positions in the above-mentioned adventitious root sample indicated one to two additional rounds of partial genome duplications for 50.22% and 19.56% of the nuclei studied, where the produced 4E DNA content became amplified 1.85-fold (I3/I2 = 1.85) and the yielded 8E DNA content extended 1.97-fold (I4/I3 = 1.97), respectively. Putting the obtained fluorescence intensity ratios from all samples together into three subsets, according to the three detected partial genome endoreplication cycles, mean values were: I2/I1 = 1.86 (sd: 0.04, CV: 2.26%, n = 23); I3/I2 = 1.91 (sd: 0.01, CV: 0.64%, n = 10); I4/I3 = 1.93 (sd: 0.008, CV: 0.4%, n = 2). Due to the low CVs of the interpeak ratios, it seems plausible that the same part P of the original 2C genome is replicated in every new endoreplication cycle (Brown et al. 2017). The corresponding linear regression (Fig. 3) led to an estimated part P of 83.6% of the genome that took part in the endoreplication process, which corresponds to a part F of 16.4% that remained fixed. Finally, the regular cell cycle was recorded with diagrammatic clarity (Fig. 2 i) by examining field-collected seed embryos of D. majalis that were crushed and afterwards labelled with DAPI (see Table 1, row 8, recording high relative numbers of 2C cells in contrast to low relative numbers of 4C cells).

Flow cytometric pattern of nuclei that were liberated from different biological source materials and then became labelled by PI or alternatively by DAPI. For each instrumental run, the histogram plot provides the measured fluorescence intensities (x-axis, half-logarithmic scale) and the counted absolute numbers of nuclei (y-axis, peak area) that were found in association with the identified peaks. The peaks marked by 2C, 4E, 8E, and 16E represent different specimens of in vitro germinated D. majalis plants, carrying out iterated rounds of strict partial genome duplications at the stage of endomitosis (a–h). The peaks marked by 2C and 4C indicate sampled embryo seeds of D. majalis, performing a complete genome duplication under the guidance of the regular cell cycle (i). To test reliability of data and to allow nuclear DNA mass estimations, pea-leaf nuclei of known genome size and the orchid nuclei in question were co-prepared and labelled together in the same vessel. Pea nuclei yield separate peaks in positions [2C] and [4C], as marked by arrows, when they were pushed together with the orchid nuclei through the flow cytometer. The analyzed D. majalis specimens, with the applied instrumental PMT gains given in brackets, were: a. stem piece of shoot no. B4 (325); b. leaf piece of plantlet no. E1 (385); c. leaf peace of plantlet no. A3 (385); d. leaf piece of plantlet no. B2 (360); e. leaf piece of mature plant no. B1 (360); f. leaf piece of dormant plant no. D (385); g. leaf piece of mature plant no. B1 (379); h. adventitious root piece from plantlet no. A3 (325); i. open-pollinated seed embryos from the natural habitat (440). Information on the growing conditions and the ontogenetic stages of the sampled in vitro germinated D. majalis plants as well as the used protocols to liberate and to label nuclei from different source materials are summarized in Table 1

Regression line of the relative amount of DNA as compared to the 2C status, which was estimated using the FCM-based fluorescence intensity ratios, against the expected numbers of DNA copies relative to 2C assuming that the whole genome gets duplicated in every replication cycle
Cytological studies
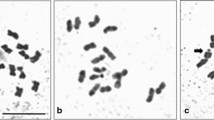
When acetocarmine smears of in vitro germinated plants were inspected by light microscopy, mononucleate leaf cells became visible. Epidermal cells, representing the lower surface of the leaf, were free from chloroplasts and exhibited large well-preserved spherical nuclei (Fig. 4 a). Stained chromocenters were observed inside the nuclei with clean color contrasts. Some of these spots were attached to the inner surface of the nuclear envelope in a chain-like structure, whereas the remainders spread over the surface of the nucleoplasm (Fig. 4 a). Data suggest the very beginning of a prophase when replicated chromatin starts to condense. Instead to continue with a regular mitosis, however, epidermal cells synchronously arrested at the stage of endomitosis. This developmental stage, indicated by the complete lack of metaphase, anaphase, telophase, and cytokinesis stages in smear preparations (Fig. 4 b), explains the consecutive rounds of partial DNA duplications previously measured by FCM (see above). It is of note that large stomates, which are sprinkled within the lower epidermis, contained masses of chloroplasts, so that endomitotic nuclei became rarely detectable in this cell type (Fig. 4 c). When hereafter adventitious roots were sampled from in vitro germinated plants at some distance above the meristematic tip, acetocarmine-stained abundant endomitotic nuclei appeared again (Fig. 4 d) corresponding to the above-described rounds of partial genome duplications that have been initially identified in this organ part. By contrast, preparations of acetocarmine-stained meristematic cells from the growing tip region of adventitious roots, which is confined to the terminal 1 to 2 mm, showed signs of a proper mitosis. Especially, distinct metaphase chromosomes (Fig. 4 e) and the pattern of cytokinesis (Fig. 4 f) were recorded, confirming complete cell divisions that occurred under guidance of the regular cell cycle. Unlike the above-mentioned meristematic root cells, specimens of young anthers that were collected and stained during the expression of the microspore stage illustrated the haplophasic number of 1n = 2x = 40 circular chromosomes, indicating meiosis (Fig. 4 g).

Light micrographs, recording examples of acetocarmine-stained smear preparations from different organs of in vitro germinated D. majalis plants. a–b. Abundant endomitotic nuclei, found in the lower epidermis region of a juvenile leaf (plant no. B2); c. The arrow indicates a rare example of an endomitotic nucleus, which has been detected inside a stomatal cell of the lower epidermis region of a juvenile leaf (plant no. B2); d. Abundant endomitotic nuclei of adventitious root tissues, sampled at some distance above the meristematic tip (plant no. A3); e. The arrow marks circular chromosomes in the metaphase stage of mitosis, when the growing tip region of an adventitious root was sampled (plant no. A3); f. Smear preparation, obtained from the growing tip region of an adventitious root, showing abundant cells at the stage of cytokinesis when division of the cytoplasm became finished as part of the regular cell cycle. The newly formed thin cell wall is exemplarily marked by an arrow (plant no. A3); g. Smear preparation, sampled from a collection of young anthers during the expression of the microspore stage. A single microspore that allowed to count the haplophasic number of 1n = 2x = 40 chromosomes is marked by an arrow (plant no. B1)
Discussion
Cultivation of plants in a mycorrhiza-free environment
We have established the methods in our laboratory sphere that have been previously assembled by Debergh (1994), which aimed at replacing the native symbiotic association between orchid seeds and mycorrhizal fungi with simple sucrose-containing media to assist protocorm production and in vitro cultivation of D. majalis plants. Since an auxin application, for instance, is known to increase the proportion of 4C and 8C nuclei in embryos during in vitro germination as compared to those in auxin-free medium (Lim and Loh 2003), no exogenous growth regulators were applied to prevent possible interferences with the objectives of the study. In this way, 2 to 8% of the disinfected seeds were respectively converted into plants, with no distinct differences observed amongst all tested seed lots.
Properties of the detected endoreplication process
When PI-stained nuclear suspensions from the stem, adventitious roots as well as leaves of the fostered indoor plants were analyzed by FCM, one to three successive rounds of partial genome duplications could be detected. Nuclei were identified in peak positions 2C, 4E, 8E, and 16E according to the fact that each considered ratio Ik/Ik-1 attained a value of less than 2. Pea-leaf nuclei were collectively liberated, stained, and measured with the orchid nuclei to be examined, but provided separate peaks at 2C and 4C positions (in these cases Ik/Ik-1 always turned out to be 2). Thus, it is unlikely that the above data suffered from interferential artefacts with secondary plant metabolites due to a possible inhibition of staining and/or probably overlapping fluorescence peaks. Moreover, the occurrence of iterated rounds of partial genome duplications was proven for one leaf of orchid individual no. B1 by two distinct fluorochromes. In this case, PI-labelled foliar nuclei showed up to two replication rounds, while DAPI-labelled foliar nuclei constantly signaled three replication rounds because of different target specificities and sensitivities for both tested fluorochromes. Since the sampled plants notably differed with respect to agar-grown and potted individuals, including diverse organs and age classes, we always employed an individual PMT gain (see legend of Fig. 2), for which the two sets of nuclei from the standard sample (pea) and the orchid sample in question produced separate 2C peaks (see Fig. 2) to enable reliable genome size estimations. Once selected, this gain was repeatedly used in the case of the analyzed specimen to ascertain the absolute DNA mass data presented in Table 1. The above-mentioned practice surprisingly demonstrated the absence of the 2C peak within the tested adventitious root sample (Fig. 2 h), indicating a newly discovered organ-specific phenomenon for D. majalis. Moreover, we detected abundant foliar 2C cells within the analyzed plants only as long as the flowers not appeared on the plant body (compare Fig. 2 b–d and e), reflecting a hitherto unknown ontogenetically controlled trait with respect to the leaf development. In conjunction with PMT gains 360 and 385 for certain samples, we can unfortunately not exclude the existence of potential 8E and 16E peaks that were out of the detection range. For that reason, both gain settings are probably not suited to record the whole range of naturally acting endoreplication processes in D. majalis by FCM technology. Based on calculations using the above-described ratios of fluorescence intensities (Ik/Ik-1), about 16–17% of the 2C genome (part F) do not take part in the endoreplication cycle within the investigated individuals due to an unknown mechanism, whereas the remaining 83–84% of the 2C genome (part P) content is endoreplicated. Because the consecutive genomic replication steps do not seem to be synchronized, a mosaic of somatic nuclei with highly different DNA amounts and diverse frequency distributions was detected. Yet, there is currently no nucleotide sequence information available with respect to both contrasting genomic parts. The above-mentioned DAPI-born peaks support the hypothesis that partial endoreplication is unable to selectively act on gene-coding regions (Hřibová et al. 2016). However, large-scale nucleotide sequencing analyses using orchids from the subtribe Pleurothallidinae (Chumová et al. 2021) and the model orchid Vanilla planifolia Andrews (Piet et al. 2022) have shown that uncoding repetitive DNA does not proliferate within the endoreplicated genome parts.
Iterated under-replications of genomic DNA, in which small target regions became discriminated by an unknown mechanism, are widespread amongst orchid taxa as has been initially investigated by Trávniček et al. (2015). In the case of many orchid species, repeated experiments have shown strongly conserved fluorescence intensity ratios for each round of DNA replication and additionally indicated stable relationships from cycle to cycle in agreement with a strict partial endoreplication process (Brown et al. 2017). When the mean fluorescence intensity ratios are converted to the DNA index R (relative amount of DNA compared to 2C) as proposed by Brown et al. (2017), and when this index R then scales linearly with increasing numbers of elapsed replication rounds, this suggests that a fixed part of the genome F does not take part in the endoreplication process. The slope of the corresponding regression line equals the proportion P of the genome, which becomes endoreplicated. However, there is no formal proof that the parts F and P are really constant during the endoreplication process. Yet, according to arguments given in Brown et al. (2017), the low variability of the fluorescence intensity ratios in our repeated experiments using D. majalis (see Table 1) can be interpreted in a way, such that F and P are likely to be constant genomic parts across all endoreplication cycles. In addition, our results were consistent across different sampled individuals, three vegetative plant organs, diverse ontogenetic stages, altered cultivation conditions and two geographic seed provenances. As a result, the endoreplication we found for D. majalis fits the model of consecutive rounds of strict partial genome endoreplications as defined by Brown et al. (2017) with a part P of 83–84% of the original 2C genome being endoreplicated. Most notably, the running cycle of repeated DNA under-replications started with axenically grown juvenile individuals of D. majalis during their cultivation on solid nutrient media. To the best of our knowledge, the current work is the first study which demonstrates that strict partial endoreplication occurs in orchid plants grown in a mycorrhiza-free environment. The running cycle of repeated under-replications was found again when in vitro germinated plantlets were moved to potting soil, where they further maturated under indoor conditions. Note that potted plants that were moved from the laboratory to the environment confirmed the above-mentioned phenomenon under outdoor conditions as well (data not shown).
Endomitosis causes the observed endoreplication process
Paying attention to cytological preparations, obtained from in vitro germinated D. majalis plants, we saw acetocarmine-stained chromocenters inside non-dividing nuclei with epidermal leaf cells and with adventitious root cells at some distance above the meristematic tip. Almost identical signs of endomitosis, which led to a moderately increased DNA content in response to arrested karyokinesis and missing cytokinesis, have been previously reported for mononucleated root cortex cells of the orchid species Vanilla planifolia forming intravacuolar crystals of calcium oxalate (Kausch and Horner 1984). Endomitosis has been rarely found in plant cells (D'Amato 1984) in contrast to various non-plant phyla, where this phenomenon frequently occurs during tissue-specific organ development (see e.g. van Rijnberk et al. 2022).
Possible consequences of the detected endoreplication process
Living cells, conducting consecutive rounds of genome endoreplications, are assumed to benefit from both large sizes and high protein synthesizing rates due to increased gene dosages. For example, a ploidy-dependent increase in gene transcription has been detected during tomato fruit growth (Bourdon et al. 2012). Endopolyploid cells are furthermore proposed to grow faster by reducing the elementary processes required for life (Edgar et al. 2014). Specifically, the complex cellular machinery orderly operating mitosis is switched off under such conditions. Indeed, it is important to emphasize that endoreplication exclusively concerns somatic tissues. The amplified DNA amounts are not inherited to the next generation, which we confirmed by successfully counting the expected haplophasic number of 40 chromosomes within a microspore of D. majalis as well as by demonstrating regular cell cycle nuclei in field-collected seed embryos, which only showed fluorescence intensities at normal 2C and 4C positions.
Genome size variation
When estimating the DNA content of vegetative D. majalis nuclei at the 2C level by FCM and using the PI-fluorochrome, a narrow span from 14.60 pg to 14.92 pg was obtained. Due to these marginal differences, it seems likely that all investigated plants belong to the same allotetraploid species. Based on the regular haplophasic number of 40 chromosomes, as recorded from the microspore stage of young anthers when the flowers appeared, foliar cells of plant no. B1 were considered to contain the whole vegetative set of 80 chromosomes, leading to a mean amount of 14.71 pg DNA per nucleus at the 2C genome level (this is equivalent to 1.4386 × 1010 base pairs). Wróblewska et al. (2019) and Šmarda et al. (2019) published mean values of 15.16 pg and 13.74 pg, respectively, while Eriksson et al. (2022) presented 2C values for eight accessions, ranging from 13.96 to 15 pg.
Although many species show a remarkable constancy in the genome size, there are some striking exceptions of intraspecific variation relating e.g. to chromosome polymorphisms (e.g. the occurrence of sex-specific chromosomes of special shape, present and absent B chromosomes), whole-scale duplication events, massive transposable element activity (Gregory 2004), and environmental impacts (Hrabovský et al. 2024). It is furthermore known that systematic inter-laboratory differences due to different FCM devices in genome size estimates exist, which might also lead to small, but significant deviations in DNA mass estimates (Doležel et al. 1998). Eriksson et al. (2022) proposed that the rapid evolutionary rate of a tandem-repeat, which probably derived from a repetitive miniature inverted-repeat transposable element, can result in considerable genome size differences among D. majalis populations. Moreover, the maternally inherited chloroplast DNA sequences, occurring in different accessions of D. majalis, share similarity with both current Dactylorhiza fuchsii (Druce) Soó and Dactylorhiza maculata (L.) Soó plastomes (Brandrud et al. 2020), or were not yet detected in any of the extant parental species (Nordström and Hedrén 2009). Such data provide strong evidence that allopolyploid D. majalis has developed from recurrent processes of hybridization, which possibly could also lead to different nuclear genome sizes. Altering the present number of chromosomes by irregular nondisjunction at meiosis makes a major contribution to the chromosomal abnormalities found in allopolyploid species due to possible recombinations between both sets of homoeologous chromosomes. The provided extraordinary genetic variation will become enriched, if it is more efficiently adapted to the occurring environment in comparison to the previously available genetic make-up (Comai 2005). Interestingly, an aneuploid D. majalis karyotype, confirming 78 chromosomes of uniform size, was recently recorded from a natural habitat (Jonsdorf, Saxony, Germany) close to the Waltersdorf population investigated in the present study, yet without a corresponding nuclear DNA mass estimation (Schubert et al. 2020). Based on the assumption that each single D. majalis chromosome among 80 equal-sized chromosomes contributes to a DNA mass of 0.1839 pg (14.71 pg divided by 80), we argue that the fluctuating genome size variation of the present study might indicate an aneuploid chromosome number of 81 with an increased nuclear DNA content of 14.92 pg (CV = 0.214%) for plant no. E1. It is of note that physicochemical methods have been generally accepted to provide sample data in repeated experiments with sufficiently good reliability when the obtained CV ranges between 2 and 5% (Rasch 1983), which would correspond to two to four missing or additional D. majalis chromosomes. Hence, chromosome counts via cytological preparations are indispensable for a correct identification of aneuploids and even to rule out the many other causes of intraspecific DNA variation mentioned above.
Factors influencing genome size estimation using FCM
To attain correct nuclear genome mass data for D. majalis by FCM technology, it is generally advisable to avoid sampling of leaves during the flowering period and to avoid adventitious root pieces for some distance above their meristematic tips because, under these circumstances, the 2C band drops from the analysis or falls behind the detection limit (Fig. 2 e and 2 h). If the 2C signal is missing and the 4E peak of the first endoreplication round is mistaken for the regular 2C position, this leads to an overestimation of the nuclear DNA mass (e.g., 28.31 pg vs 14.15 pg in case of the sampled adventitious tissue of plant no. A3). The phenomenon that foliar 2C cells are rare or sometimes missing when the plants exhibit their flowers has been initially described by Trávniček et al. (2015) for a couple of orchid species other than D. majalis. For D. majalis, we found abundant foliar 2C cells before the flowers appeared, so that almost every sampled leaf permitted suitable genome size estimation by FCM. Rare foliar 2C cells in blooming D. majalis individuals and repeated rounds of endoreplications have also been detected in a previous field study in natural populations of Saxony (unpublished data). However, FCM analyses of these cells were impaired by the high amount of secondary plant metabolites in the corresponding nuclear suspensions, attributable to the inappropriate nuclear isolation and staining procedures originally employed, which, in addition to the limited availability of plant material from natural populations due to environmental protection law, made the in-depth experiments necessary reported here. In contrast to Trávniček et al. (2015), we have neither tested young ovaries nor pollinaria by FCM-technology. Since flowers, comprising the above-mentioned reproductive organs, are available in the nature only for a small span of time, and because not every individual is blooming during the season, their use in the identification of aneuploids in natural populations is limited. Further possible sources of error in the context of FCM-based DNA mass estimation, e.g. the species to be analyzed and the species used as internal calibration standard show their 2C bands at identical channel positions or one of them did not release nuclei during the joined preparation of suspensions, have been described by Zonneveld (2019).
Implications for nature conservation
Survey data on sexual reproduction, including tests on seed fertility, will be urgently needed for Germany, where up to 30% of the world-wide remaining D. majalis populations have been assumed to exist under the challenging conditions of global warming (Ludwig et al. 2007). During the past, when numerous prezygotic and/or postzygotic barriers amongst species collapsed, vigorous hybrids appeared in disturbed habitats after irregular germinal cells with unreduced chromosome numbers mated and became cross-fertilized. The newly-born polyploid chromosome sets (3x, 4x, 5x, 6x, etc.), bringing genetic information from two or more species together in one nucleus, were able to process high levels of genetic variability via DNA recombination and allowed a fast adaptation on diverse environmental stresses by genotypic selection (a collection of such data has been assembled by Futuyma 1986; for a current review see Kopecký et al. 2022; for a recent work on D. majalis see Wolfe et al. 2023). FCM, as applied in the current work, can be used to detect hybrids among members of the genus Dactylorhiza. The necessary cytological confirmation of the hybrid status should be advisably done using germinal tissues with a haplophasic chromosome set, due to the high chromosome numbers expected in vegetative tissues. For those scientists, wishing to contribute to the above-mentioned fields of ecological genetics in the future, the techniques and data described here represent a valuable guidance with a focus on D. majalis and other species of the genus Dactylorhiza.
Data Availability
Not applicable.
References
Alberts B, Johnson A, Lewis J, Morgan D, Raff M, Roberts K, Walter P (2015) Molecular biology of the cell, 6th edn. Garland Science, Taylor & Francis Group, Abingdon, New York
Bourdon M, Pirrello J, Cheniclet C, Coriton O, Bourge M, Brown S, Moïse A, Peypelut M, Rouyère V, Renadin J-P, Chevalier C, Frangne N (2012) Evidence for karyoplasmic homeostasis during endoreplication and a ploidy-dependent increase in gene transcription during tomato fruit growth. Development 139:3817–3826
Brandrud MK, Baar J, Lorenzo MT, Athanasiadis A, Bateman RM, Chase MW, Hedrén M, Paun O (2020) Phylogenomic relationships of diploids and the origins of allotetraploids in Dactylorhiza (Orchidaceae). Syst Biol 69(1):91–109
Bräutigam S, Otto H-W (2012) Rote Liste der Farn- und Samenpflanzen in der Oberlausitz: aktualisierte Fassung. Berichte der Naturforschenden Gesellschaft der Oberlausitz 20:99–116
Breuer C, Braidwood L, Sugimoto K (2014) Endocycling in the path of plant development. Curr Opin Plant Biol 17:78–85
Brown SC, Bourge M, Maunoury N, Wong M, Bianchi MW, Lepers-Andrzejewski S, Besse P, Siljak-Yakovlev S, Dron M, Satiat-Jeunemaître B (2017) DNA remodeling by strict partial endoreplication in orchids, an original process in the plant kingdom. Genome Biol Evol 9:1051–1071
Chumová Z, Záveská E, Hloušková P, Ponert J, Schmidt PA, Čertner M, Mandáková T, Trávniček P (2021) Repeat proliferation and partial endoreplication jointly shape the patterns of genome size evolution in orchids. The Plant J 107:511–524
Comai L (2005) The advantages and disadvantages of being polyploid. Nature Rev Genet 6:836–846
D’Amato F (1984) Role of polyploidy in reproductive organs and tissues. In: Johri BM (ed) Embryology of angiosperms. Springer, New York, pp 519–566
Debergh P (1994) In vitro culture of ornamentals. In: Vasil IK, Thorpe TA (eds) Plant cell and tissue culture. Kluwer Academic Publishers, Dordrecht, pp 561–573
Devos N, Raspe O, Oh S-H, Tyteca D, Jacquermat A-L (2006) The evolution of Dactylorhiza (Orchidaceae) allotetraploid complex: insights from nrDNA sequences and cpDNA PCR-RFLP data. Mol Phylogen Evol 38:767–778
Doležel J, Bartoš J (2005) Plant DNA flow cytometry and estimation of nuclear genome size. Ann Bot 95(1):99–110
Doležel J, Greilhuber J, Lucretti S, Meister A, Lysák MA, Nardi L, Obermayer R (1998) Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann Bot 82(Suppl. A):17–26
Doležel J, Bartoš J, Voglmayr H, Greilhuber J (2003) Nuclear DNA content and genome size of trout and human. Cytometry 51A:127–128
Doležel J, Greilhuber J, Suda J (2007) Flow cytometry with plant cells. Wiley-VCH, Weinheim
Edgar BA, Zielke N, Gutierrez C (2014) Endocycles: a recurrent evolutionary innovation for post-mitotic cell growth. Nature Rev Mol Cell Biol 15:197–210
Eriksson MC, Mandáková T, McCann Temsch EM, Chase MW, Hedrén M, Weiss-Schneeweiss H, Paun O (2022) Repeat dynamics across timescales: a perspective from sibling allotetraploid marsh orchids (Dactylorhiza majalis s.l.). Mol Biol Evol 39(8):msac167
Futuyma DJ (1986) Evolutionary biology. Sinauer Associates, Sunderland
Gregory TR (2004) Macroevolution, hierarchy theory, and the C-value enigma. Paleobiol 30:179–202
Greilhuber J, Doležel J, Lysák MA, Bennett MD (2005) The origin, evolution and proposed stabilization of the terms “genome size” and “C-value” to describe nuclear DNA contents. Ann Bot 95(1):255–260
Hedrén M (1996) Genetic differentiation, polyploidization and hybridization in northern European Dactylorhiza (Orchidaceae): evidence from allozyme markers. Plant Syst Evol 201:31–55
Heller FO (1973) DNS-Bestimmung an Keimwurzeln von Vicia faba L. mit Hilfe der Impulscytophotometrie. Berichte der deutschen Botanischen Gesellschaft 86:437–441
Hrabovský M, Kubalová S, Mičieta K, Ščevková J (2024) Environmental impacts on intraspecific variation in Ambrosia artemisiifolia genome size in Slovakia, Central Europe. Environ Sci Pollut Res 31:33960–33974
Hřibová E, Holušová K, Trávniček P, Petrovská B, Ponert J, Šimková H, Kubátová B, Jersáková J, Čurn V, Suda J, Doležel J, Vrána J (2016) The enigma of progressively partial endoreplication: new insights provided by flow cytometry and next-generation sequencing. Genome Biol Evol 8:1996–2005
Hunt PF, Summerhayes VS (1965) Dactylorhiza Nevski, the correct generic name of the Dactylorchids. Watsonia 6:128–133
Joubès J, Chevalier C (2000) Endoreplication in higher plants. Plant Mol Biol 43:735–745
Kausch AP, Horner HT (1984) Increased nuclear DNA content in raphide crystal idioblasts during development in Vanilla planifolia L. (Orchidaceae). Eur J Cell Biol 33:7–12
Kopecký D, Martín A, Smýkal P (2022) Interspecific hybridization and plant breeding: From historical retrospective through work of Mendel to current crops. Czech J Gen Plant Breed 58:113–126
Lim WL, Loh CS (2003) Endopolyploidy in Vanda Miss Joaquim (Orchidaceae). New Phytol 159:279–287
Lister A (1990) Flow cytometry for selection of plant cells in vitro. In: Dix PJ (ed) Plant cell line selection. VCH, Weinheim, pp 39–85
Loureiro J, Čertner M, Lučanová M, Sliwinska E, Kolář F, Doležel J, Garcia S, Castro S, Galbraith DW (2023) The use of flow cytometry for estimating genome sizes and DNA ploidy levels in plants. Methods Mol Biol 2672:25–64
Ludwig G, May R, Otto C (2007) Verantwortlichkeit Deutschlands für die weitere Erhaltung der Farn- und Blütenpflanzen - vorläufige Liste. BfN-Skripten 220:1–102
Nordström S, Hedrén M (2009) Genetic diversity and differentiation of allopolyploid Dactylorhiza (Orchidaceae) with particular focus on the Dactylorhiza majalis ssp. traunsteineri/lapponica complex. Biol J Linnean Soc 97:52–67
Piet Q, Droc G, Marande W, Sarah G, Bocs S, Klopp C, Bourge M, Siljak-Yakovlev S, Bouchez O, Lopez-Roques C, Lepers-Andrzejewski S, Bourgois L, Zucca J, Dron M, Besse P, Grisoni M, Jourda C, Charron C (2022) A chromosome-level, haplotype-phased Vanilla planifolia genome highlights the challenge of partial endoreplication for accurate whole-genome assembly. Plant Communications 3(5):100330
R Core Team (2022) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Rasch D (1983) Biometrie. Deutscher Landwirtschaftsverlag, Berlin
Rode P, Schadwinkel H-J (2014) Dactylorhiza majalis (Rchb.) P.F. Hunt and Summerh. Breitblättriges Knabenkraut, Mai-Kuckucksblume. In: Heinrich W, Voelckel H, Dietrich H, Feldmann R, Geithner A, Kögler V, Rode P, Westhus W (eds) Thüringens Orchideen. Arbeitskreis Heimischer Orchideen Thüringen e.V, Uhlstädt-Kirchhasel, pp 395–405
Sass JE (1958) Botanical Microtechnique. Iowa State University Press, Ames
Schoenichen W (1940) Biologie der geschützten Pflanzen Deutschlands. Eine Einführung in die lebenskundliche Betrachtung heimischer Gewächse. Verlag Gustav Fischer, Jena
Schubert R, Brugger M, Kühnel S, Hohlfeld H, Heidger CM (2020) Analyses of sexual reproductive traits in Dactylorhiza majalis: a case study from East Germany. Biologia 75:507–521
Šmarda P, Knápek O, Březinová A, Horová L, Grulich V, Danihelka J, Veselý P, Šmerda J, Rotreklová O, Bureš P (2019) Genomes sizes and genomic guanine+cytosine (GC) contents of the Czech vascular flora with new estimates for 1700 species. Preslia 91:117–142
Trávniček P, Ponert J, Urfus T, Jersáková J, Vrána J, Hřibová E, Doležel J, Suda J (2015) Challenges of flow-cytometric estimation of nuclear genome size in orchids, a plant group with both whole-genome and progressively partial endoreplication. Cytometry A 87A:958–966
Van Rijnberk LM, Barrull-Mascaró R, van der Palen RL, Schild ES, Korswagen HC, Galli M (2022) Endomitosis controls tissue-specific gene expression during development. PLOS Biol 20:e3001597
Von Denffer D, Ehrendorfer F, Mägdefrau K, Ziegler H (eds) (1978) Lehrbuch der Botanik. Gustav Fischer Verlag, Jena, p 46
Wolfe TM, Balao F, Trucchi E, Bachmann G, Gu W, Baar J, Hedrén M, Weckwerth W, Leitch AR, Paun O (2023) Recurrent allopolyploidizations diversify ecophysiological traits in marsh orchids (Dactylorhiza majalis s.l.). Mol Ecol 32:4777–4790
Wróblewska A, Szcepaniak L, Bajguz A, Jedrzejczyk I, Tałałaj L, Ostrowiecka B, Brzosko E, Jermakowicz E, Mirski P (2019) Deceptive strategy in Dactylorhiza orchids: multidirectional evolution of floral chemistry. Ann Bot 123:1005–1016
Yeung EC (2017) A perspective on orchid seed and protocorm development. Bot Studies 58:33–46
Zonneveld BJM (2019) The DNA weights per nucleus (genome size) of more than 2350 species of the flora of The Netherlands, of which 1370 are new to science, including the pattern of their DNA peaks. Forum Geobot 8:24–78
Acknowledgements
This project was financially supported by a grant donated by the Ministry of Science and Art in Saxony.
Funding
Open Access funding enabled and organized by Projekt DEAL. Sächsisches Staatsministerium für Wissenschaft und Kunst
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schubert, R., Hohlfeld, H. & Brugger, M. Flow cytometry and cytological studies reveal consecutive rounds of strict partial genome duplications and endomitosis in cellular nuclei from vegetative organs of seeded Dactylorhiza majalis (subfamily Orchidoideae). Biologia 80, 823–837 (2025). https://doi.org/10.1007/s11756-025-01879-7
Received:
Accepted:
Published:
Version of record:
Issue date:
DOI: https://doi.org/10.1007/s11756-025-01879-7