
Abstract
Under global warming, drought will reduce productivity of Pinus halepensis s.l. (subspecies halepensis and brutia) and cause a retreat from its rear edge distribution (latitudinal/elevational) in the Mediterranean. To test whether topography can influence this scenario, we studied for approximately 40 years the growth of six natural pine stands in water-limited habitats on the islands of Zakinthos and Samos (eastern Mediterranean Greece), and determined the critical moisture sources that drove pine growth. Dominant pines were selected with no permanent water sources under contrasting moisture conditions created by topography (“wet”-gulley/valley vs. “dry”-upslope habitats). The responses of P. halepensis s.l. to drought under a moderate and a worst case scenario were tested, projected under global warming (approx. − 25% and 40% in annual precipitation compared to 1961–1990 average). Our results show that “wet” habitat pines had higher productivity under normal to wet climate. However, the more precipitation declined, “wet” habitat tree growth was reduced at a significantly faster rate, but also showed a faster recovery, once rainfall returned. Thus, Pinus halepensis s.l. populations in gullies/valleys, may be more drought resilient and less likely to retreat towards higher elevation/latitudes under global warming, compared to pines on dry upslope sites. Under moderate drought, both ecosystems relied on deeper moisture pools supplied by rainfall of the previous 3–6 years (including the year of growth). However, valley/gully habitat pines on significantly deeper soils (and probably on deeper heavily weathered bedrock), appeared to utilize surface moisture from winter/spring rainfall more efficiently for survival and recovery. Thus, deep soils may provide the key “buffer” for pine survival in such ecosystems that could act as potential refugia for P. halepensis s.l. under climate change.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Under global warming scenarios, climate models project intense warming and reduced precipitation for mid-latitude regions of the planet, with the rate of warming for hot extremes to double that of the average rate of global warming (IPCC 2023). For the Mediterranean region, average temperatures are predicted to increase by 20%, precipitation to decline between 4% and 22% and droughts to be 5–10 times more frequent (Cherif et al. 2020). At global warming levels of 2.5–3.5 °C, climate risks are expected to become very high for species of the Mediterranean drylands, leaving migration from their current distribution range as a possible option (Guiot and Cramer 2016; Balzan et al. 2020).
The expected increase in stress conditions will impact forests and affect their growth and survival, mainly in arid areas (Vicente-Serrano et al. 2010). Thus, plant species are expected to shift poleward and upwards in response to warming and drought intensification, leading to geographic shifts towards more climatically suitable areas (Pounds et al. 1999; Christmas et al. 2016), fundamentally altering forest composition, structure, and biogeography (Allen et al. 2010).
There is evidence to suggest that forests are responding to recent changes in climate. For instance: (1) changes in plant distribution across temperate and Mediterranean mountain forests in western Europe (Lenoir et al. 2008); (2) upward migration of forest species in the Italian Apennines (Palombo et al. 2013), Spain (Peñuelas and Boada 2003; Peñuelas et al. 2007; Améztegui et al. 2010, 2016) and in southeastern France (Bodin et al. 2013); (3) significant growth reductions for eastern Mediterranean pine forests growing close to sea level caused by recent reductions in precipitation unprecedented in more than 100 years (Sarris el al. 2011; Dorman et al. 2015b). These last phenomena have been accompanied by increases of stand mortality across different species (Körner et al. 2005; Sarris et al. 2007, 2011; Allen et al. 2010; Dorman et al. 2015c), especially for species growing at their lowest latitudinal/elevational range (Sarris el al. 2011; Dorman et al. 2015c). Investigating these responses is critical for Mediterranean ecosystems, as they may experience the greatest proportional change in biodiversity under global warming (Sala et al. 2000).
However, it is not only climate that determines plant species productivity and distribution. Upward migration of lower zone species can be influenced by topography and geomorphology (Chen et al. 2007; Kazakis et al. 2007). Topography creates contrasting microclimates, especially between northern and southern slopes in drylands, which results in distinct differences in plant cover patterns in the landscape, effecting tree establishment, growth rates and survival (Principe et al. 2019; Balzan et al. 2020). Besides climatic variables, topography affects soil water movement and therefore soil moisture availability in complex terrains (Stephenson 1990; Chen et al. 2007). Therefore, to better understand and predict the response of ecosystems or their components to climatic change, it is very important that the interaction of climate with topography is investigated, as has been shown, e.g., for modeling net primary production (Xu et al. 2008; Govind et al. 2009; Huang et al. 2010), for determining species richness (Denslow 1995; Daws et al. 2002) and for accessing stand mortality (Allen and Breshears 1998; Guarin and Taylor 2005).
To investigate these effects, this study focused on Pinus halepensis s.l., the dominant conifer at low elevations in the Mediterranean (Ne’eman and Trabaud 2000). Its subspecies halepensis is located at the western part of the basin, while the subspecies brutia is found at its eastern edge (Christensen 1997). Both taxa combined occupy an area of some 7.5 million ha (Fady et al. 2003; Fig. 1). Other researchers consider them as separate species, i.e., Pinus halepensis Mill. (Aleppo pine) and Pinus brutia Ten. (Turkish or Calabrian pine) (Mauri et al. 2016). However, both have been found to be equally sensitive to changes in moisture availability or resilience to drought (Sarris et al. 2011; Veuillen et al. 2023a).

Under global warming, the degree of Pinus halepensis sl. retreat is unclear. Some models project a slight northward expansion with contraction from the extreme southeast and southwest of its current range, whereas others suggest that this contraction will be minute (Thuiller 2003). At is southwestern limits in Spain, approximately 20% of current stands are predicted to become unsuitable for P. halepensis by 2050–2080 according to different models under climatic change (Keenan et al. 2011). However, other studies document its resilience capacity and suggest that it will thrive in a drier future climate (Gazol et al. 2022). Although several studies have investigated the role of climate on the distribution and growth of P. halepensis, few have considered the effect of local site conditions nor their interaction with climate (Vennetier et al. 2018).
In addressing topographic effects on the productivity of P. halepensis s.l., one hypothesis is that in valleys or gullies, (land depressions), deep soils are formed supported by runoff from surrounding slopes, creating “wetter” habitats compared to neighboring hill slopes as suggested by Vennetier et al. (2018). It would be expected that forest productivity in such habitats would benefit from the additional supply of moisture and nutrients created by runoff even when soil and bedrock type do not permit permanent surface water accumulation (e.g., calcareous habitats; Sarris et al. 2007). As a result, better growth rates and denser stands may be expected compared to “drier” neighboring slopes. However, this may increase within stand competition, (if climate becomes drier), which in turn may increase water stress (Innes 1992; Dolph et al. 1995). Thus, these trees cannot be excluded from vulnerability to drought triggered dieback (Allen and Breshears 1998; Guarin and Taylor 2005).
Fast-growing tree species, before the onset of drought, may exhibit higher mortality than slower-growing ones, possibly due to physiological adaptations prior to drought, with micro-site or genetic conditions favorable for rapid growth early in life predisposing trees to vulnerability to drought later in life (Levanič et al. 2011). Higher resource expenses for readjusting to cope with drought, e.g., for reallocation or/and expansion of root systems, compared to trees already adapted to drier conditions, could also play a role. Thus, since both high and low growth levels are associated with mortality (Jenkins and Pallardy 1995; Sarris et al. 2007; Haavika et al. 2008; Wyckoff and Bowers 2010), it is still not clear which trees of the two systems (“wet” vs. “dry” or “fast” vs. “slow” growing or “less” vs. “more” drought adapted) will become more vulnerable under global warming.
This paper explores the impact that climatic change may have on P. halepensis s.l. productivity and future distribution in the Mediterranean by considering the effects of climate and topography. By determining to what degree pine responses differ between the two systems of potentially diverse moisture and growth conditions (“wet” vs. “dry”), it may be possible to identify in which of the two P. halepensis s.l. forest stands will become more vulnerable under climate change. Such information will improve our understanding of the process behind the current and future distribution of tree species in drylands, permit more accurate projections for distribution models and improve management practices to address the impacts of climatic change. The two tree responses that we investigated were radial growth and critical moisture utilized to sustain annual variability. A novel tree ring analysis was applied focusing on natural populations from the Greek islands of Zakinthos and Samos.
Specifically, the following questions were addressed:
-
1)
Is soil depth and soil moisture holding capacity or stand density more likely to differ in topographic systems of upslope vs. gully/valley sites without permanent water sources under Mediterranean climate conditions? We hypothesized that gullies/valleys will hold denser tree stands with deeper and wetter soils.
-
2)
In which of the above two ecosystems will pines grow better under moderate or under extreme drought conditions, and how is water utilization that sustains annual growth affected? Our hypothesis is that the more drought intensifies, the more growth is expected to decline in the driest habitat, while the higher the dependence on deep ground moisture supplies for annual growth.
-
3)
Which of the above drivers is more important for the sustainable management of such stands (e.g., using selective thinning) and for identifying potential refugia to foster climate change adaptation for P. halepensis s.l. at its xeric limits? We consider that both retention of soil moisture and stand density would play a role and that forest management would need to conserve the most favorable conditions in each habitat to provide the highest resistance to the retreat of pine populations towards colder/wetter regions.
Materials and methods
Site parameters and tree selection
We investigated the combined long-term effect of topography and climate on P. halepensis s.l. populations on two islands of the eastern Mediterranean, Zakinthos for Pinus halepensis subsp. halepensis, and Samos for Pinus halepensis subsp. brutia, both islands of southern Greece (Fig. 1). Both receive winter rainfall > 600 mm (data for 1986–2000). In addition, they experience a six-month dry season. Forests grow on a calcareous substrate, rendzina type soils that drain rapidly. The latter provide conditions where tree growth is water- limited, placing low elevation pine populations close to P. halepensis s.l. drought tolerance margins. All habitats selected belong to the thermo-Mediterranean vegetation belt (Oleo-Ceratonion), the driest of vegetation belts in the Mediterranean. To obtain even stronger responses to drought, a sharp contrast in time and space was used, taking advantage of a quasi-manipulation experiment “by nature”: (1) for replicating significant differences in moisture availability, we used a short distance moisture contrast between valley/gully and upper slope habitats; (2) to simulate conditions for moderate and for severe drought expected towards the mid or the end of the twenty-first century (− 25% and − 40% in annual precipitation, respectively, compared to 1961–1990 average; IPCC 2007; Giorgi and Lionello 2008), we took advantage of extreme drought conditions the eastern Mediterranean experienced towards the end of the 20th Century (Sarris et al. 2011; Hoerling et al. 2012).
We analyzed low elevation growth (200–300 m a.s.l.) Pinus halepensis s.l. forests (natural stands; no afforestation/reforestation) from south facing “dry” upper slope habitats (no permanent water for the last three decades of the twentieth century). Dominant, undamaged trees without signs of past harvesting in their neighborhood were sampled and core samples taken at breast height, including a wide range of stem diameters (Table 1). We excluded trees with bent trunks or with asymmetric crowns (avoiding reaction wood). From each tree, two or more core samples were taken from opposite sides with a 5 mm increment corer (Mora, Sweden). The core reaching closest to the pith was kept. These data were compared with “wet” habitat ring data sampled in the same manner (e.g., trees in valleys, gullies, dry river beds at 60–70 m a.s.l.). The “wet” habitat trees received additional moisture from hillside precipitation run-off (Figs. 2 and 3). A total of six pine populations were evaluated, three “wet” and three “dry” habitats, two pairs for Samos and one for Zakinthos.

The four habitats on Samos Island. a “Wet” P. brutia site (depression/gully; 65 m a.s.l.; foreground) and its “dry” counterpart in Limnionas area (upper slope; 210 m a.s.l.; b “Wet” habitat (valley; 60 m a.s.l) and its “dry” counterpart (upper slope; 290 m a.s.l) in Vourliotes area (photographs by D. Sarris)

The two habitats on Zakinthos Island. a “Wet” P. halepensis site (valley; 70 m a.s.l.); b its “dry” counterpart (upper slope; 225 m a.s.l)
In each habitat, 500 m2 wide plots were delimited to record vegetation parameters such as: (1) number of trees and (2) their circumference at breast height for trunks above 10 cm circumference (to be transformed into diameter at breast height (DBH) and basal area (BA) per tree, as a proxy of total forest biomass and structural complexity (McElhinny et al. 2005), (3) height and (4) ground cover for pines as well as for plant species of the understory taller than 30 cm. For each island, three plots were selected per habitat type, except for the upper slope habitat on Zakinthos where two plots were established, for a total of 11 plots (Table 2).
For evaluating soil conditions in each plot, two noncontiguous assessment zones for profile and depth were carefully selected as representative of within plot conditions (e.g., in proximity to dominant pines, avoiding exposed bedrock, etc.). Within each zone, measurements were made by digging a 0.5- to 1-m wide trench down to the bedrock or to course rock debris (22 trench profiles). Immediately after excavation, a Delta-T soil moisture sensor (SM150-UM-1) was used to record volumetric soil moisture content (%) at the side of each trench, 10 cm from the surface. A minimum of two measurements were made per trench before mid-day in May on Zakinthos and early July on Samos.
Dendrochronological analysis
The age of cores with pith was determined by counting the number of annual rings from bark to pith. The number of missing rings in cores that failed to hit the pith was estimated by assuming homocentric circles of rings. Annual increments to the nearest 0.01 mm, from bark to pith, were measured by a computer-linked mechanical platform under a stereomicroscope and the software package, Time Series Analysis and Presentation, TSAP, (Frank Rinn, Heidelberg, Germany). The first 15 years of growth at coring height were not used, to avoid early growth anomalies. After cross-dating our tree-ring series (Sarris et al. 2007, 2011), we selected those that provided the highest synchronicity (GL > 65%, CDI > 95; P < 0.01; Rinn 2003) to obtain trees with the strongest possible common climatic signal.
To account for the impact of “age trend”, i.e., the tendency of tree rings to become narrower as trees age, the tree ring width series were converted into basal-area increment series in order to be able to assess tree growth variability (BAI; Biondi and Qaedan 2008; Linares and Tíscar 2011; Dorman et al. 2015a). A raw BAI chronology in cm2 was built for each site as the average of individual-tree BAI series per year. In contrast to ‘classical’ detrending, this method is among those still capable of removing the effects of “age trend”, while retaining, at most, the valuable low-frequency growth variability needed to assess growth trends and potential long-term climatic effects on tree growth (see Mazza and Sarris 2021).
Moreover, to compare growth and its possible climatic drivers between gully/valley trees and trees growing upslope, we selected trees of similar age within mature stands. Trees require time to establish and develop canopies and root systems. By selecting mature trees of similar age, we limited the effect of differences related to the tree’s life stage, e.g., avoiding the comparison between young trees with insufficient time for their root systems to mature compared to older trees with fully established roots.
Therefore, 25 trees for Samos out of 68 sampled (10 trees for “wet” and 15 for “dry” habitats) satisfied the above criteria related to homogeneity of growth patterns (i.e. cross-dating) and age. For Zakinthos, 18 trees out of 43 sampled (8 for “wet” and 10 for “dry” habitats) were used. The selected trees were 53 (“wet”) to 58 (“dry”) years-old on average for the two habitats of Samos and 75 years-old (“wet”) 88 (“dry”) for Zakinthos (in the year 2000).
BAI chronologies were converted into 5-year and 10-year running means to obtain general growth patterns. Standard errors were calculated and plotted to test whether differences in growth between the two habitats remained significant throughout the changes in drought conditions within the 1963–2000 period of investigation. For 1963–2000, BAI chronologies included growth values from all trees selected for analysis.
Research has identified trees and shrubs growing on slopes as being the most sensitive to drought based on past dieback phenomena within the regions of analysis (Körner et al. 2005; Sarris et al. 2007). Thus, to determine periods of the greatest growth decline and recovery, BAI chronologies from the upslope habitat were used (Figs. S1 and S2; Table 3).
Dendroclimatological analysis
We examined whether the selected trees showed differential changes in ground water utilization per habitat related to the variability of precipitation over 1963–2000. A longer integration period of precipitation needed for growth would suggest that trees, without access to the water table, have been utilizing moisture stored in deeper ground, accumulated from rainfall events provided even multiple years prior to growth (see Sarris et al. 2007, 2013; Sarris and Mazza 2021).
To access such climatic influences, differences in growth for each tree related to habitat were first removed, i.e., each annual ring series was standardized for habitat effects. This was achieved by dividing each annual BAI by the mean BAI of all rings from the specific tree (mean for all years of growth; Mazza and Sarris 2021). Thus, the original data was transformed into indices which retained the year to year variation. The indexed tree-ring series were then averaged, creating standardized ring width chronologies unbiased from habitat effects, but still potentially influenced by climatic signals since they retained both high and low frequency variability.
As a tool in investigating ground water utilization under variability in climate dryness, a running correlation analysis was preformed using moving “windows” between growth and periods of precipitation known to influence growth (Sarris et al. 2007; Mazza et al. 2018; Mazza and Sarris 2021). A 10-year correlation “window” was selected (Sarris et al. 2007; Mazza and Sarris 2021). The significance of correlations between growth and precipitation variables ranged from P < 0.05, to P < 0.001; considered as satisfactory for identifying the best climatic drivers for growth per analysis “window”. This method provided a detailed tracking of moisture utilization zones using tree rings. The precipitation periods correlated with tree growth, included potentially effective rainfall periods promoting growth in Mediterranean climate conditions (Sarris et al. 2007; Dorman et al. 2015b; Mazza and Sarris 2021). The lower range of these integration periods was monthly precipitation from 1 to 12 months within the year of ring formation. The upper range was multiple years of precipitation, from 2 to 6 years prior to and including the year of ring formation. A total of 21 integration periods of precipitation were tested (Figs. 6 and 7).
The deviation in rainfall from 1961 to 1990 has been used in various climate model studies as a measure for projecting precipitation in the Mediterranean region during the twenty-first century (IPCC 2007; Giorgi and Lionello 2008; Somot et al. 2008). Annual precipitation for Zakinthos was 943 mm and 782 mm for Samos over 1961–1990. We calculated the deviation of recent precipitation conditions (5- and 10-year running means) from the 1961–1990 annual precipitation average as a measure of climatic dryness. This provided an assessment of the degree to which current conditions simulated the projected precipitation regime for the region towards the end of the twenty-first century.
Precipitation data, annual and monthly, were provided by the Hellenic National Meteorological Service from the nearest weather stations to our sites (< 30 km). The characteristics of these two stations and available data periods are given in Table 1. To verify the records provided by a particular weather station, data were compared with those of the nearest other weather station. In both cases, annual precipitation trends were found to be similar (Sarris et al. 2007, 2011).
Other factors such as temperature were considered in explaining annual ring variability. Based upon the literature, the tree growth at the elevations and latitudes investigated is controlled by temperature seasonally, i.e., no growth in midwinter (see Liphschitz et al. 1984; Sarris et al. 2013), and thus, rain falling during this period contributes to deep water storage but has no immediate effect on growth (Sarris et al. 2013). Furthermore, there had not been any significant increase in temperatures during winter or during the rest of the year until 2000 that could have resulted in any significant effect on growth, as has generally been the case for the eastern Mediterranean over the last decades of the twentieth century (Mitchell and Hulme 2000; Klein Tank et al. 2002).
Statistical analysis
The IBM SPSS program v. 25.0. for statistical analysis tested our data for normal distribution with the Kolmogorov–Smirnov test and for homogenous variance throughout the series with the Levene’s test. For data normally distributed, one-way ANOVA was performed to compare means. Pearson’s correlation coefficient (r) was used as the standard for correlating normally distributed data. Its coefficient of determination (r2) determined the degree to which ring variability is explained by precipitation. Differences in slope (b) between regression lines were tested by applying an analysis of covariance (ANCOVA).
Results
Stand and soil parameters across the study habitats
The analysis of stands within the 11 selected plots revealed that “wet” habitats had generally a higher number of trees on Zakinthos (approx. 50% more; Table 2). For Samos, “wet” habitat tree numbers exceeded the “dry” habitat by approximately 10% when including olive and carob populations > 10 cm circumference (> 3 cm DBH) in mixed pine stands. In all cases, maximum pine height was about 30% greater in “wet” habitats as was the undergrowth tree and shrub vegetation. “Wet” habitat pines covered some 75% more ground on Zakinthos and 40% more in Samos compared to “dry” habitats. When considering total basal area (BA), “wet” habitats produced approx. 20% more biomass per ha, whereas on Samos, the BA was the same when mixed pine, olive and carob species were included. There were no statistically significant differences between the two habitats for average basal area per tree, with or without including the mixed pine stands on Samos.
On the other hand, the analysis of soil parameters showed distinct differences between the two systems on both islands. Soils were found ̴ 4 times deeper in gully/valley habitats (P < 0.01), whereas soil moisture for gully/valley habitats was ̴ 105% higher on Zakinthos at the beginning of the dry season (6.5% vs. 3.2%), and some 85% higher on Samos in mid-summer (3.9% vs. 2.1%) at the 10-cm depth (P < 0.01; Table 2; Fig. S3).
Growth trends under drought and rainfall recovery
“Wet” habitat trees on both Samos and Zakinthos showed significantly improved growth (approximately 4.5 and 2 times higher basal area increment; 32.4 cm2 cf. 7.2 cm2 for Samos, 34.2 cm2 cf. 16.1 cm2 for Zakinthos) than their “dry” habitat counterparts over the period of investigation (1968–1998 in 5-year running means; 1963–2000 in actual calendar years; Figs. 4 and 5; Figs. S4 and S5 for annual growth for 1963–2000). Thinner solid and broken lines mark standard error (n = 10 or 15; Figs. 4a, b; n = 8 or 10; Figs. 5a, b). Boxes ds and rs (Figs. 4a, c and Figs. 5a, c) cover the period of greatest growth decline and recovery for “dry” habitat trees (Figs. S1 and S2).

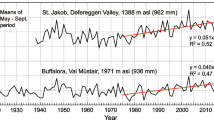
Samos Island BAI chronologies for a, b “wet” valley-gully and “dry” upper slope trees and c, d precipitation for January-December in a, c 5- and b, d 10- year running means

Zakinthos Island BAI chronologies for a, b “wet” valley-gully and “dry” upper slope trees and c, d precipitation for January–December in a, c 5- and b, d 10- year running means
Nevertheless, when precipitation declined, the differences in radial increment between the two habitats were considerably reduced (Figs. 4 and 5), as also supported by the significant correlation between precipitation and the differences in BAI of the two habitats. For Samos, this correlation was significant at P = 0.004 (r = 0.52 for 1969–1998; n = 29) and for Zakinthos at P < 0.0001 (r = 0.62 for 1965–1998; n = 34; correlations not shown). The greatest decline in precipitation was synchronous for both islands and occurred in the early 1990s. This period was selected for an in-depth growth analysis based on “dry” habitat tree performance (Table 3, Figs. S1 and S2). In all cases, differences in slopes between two regression lines of the same period (one line from “wet” the other from “dry” habitat trees) were statistically significant. As a measure of climatic dryness, the minimum or maximum precipitation (5-year mean) within each investigated period is given (min for Zakinthos and for Samos at –42% and –26% below the 1961–1990 annual precipitation mean).
“Wet” habitat tree growth appeared to decline at significantly faster rates on both islands. For Samos, it was 4.5 times faster for “wet” habitat trees (–2.3 cm2 a−1 vs. –0.5 cm2 a−1 for “dry” habitat trees; P < 0.001; 1986–1992), whereas for Zakinthos, it was twice as fast (− 1.5 cm2 a−1 vs. − 0.7 cm2 a−1 for “dry” habitat trees; P < 0.001; 1982–1991). However, when precipitation started to recover in the mid-1990s, “wet” habitat trees responded more rapidly. For Samos, this growth recovery was 3.2 times faster for “wet” habitat trees (1.8 cm2 a−1 vs. 0.6 cm2 a−1 for “dry” habitat trees; P < 0.001; 1992–1997). For Zakinthos, “wet” habitat tree recovery was 2.4 times faster (1 cm2 a−1 vs. 0.4 cm2 a−1 for “dry” habitat trees; P = 0.025; 1991–1995; Table 3).
Growth periods of “dry” or “wet” habitat trees that were best correlated with monthly to annual rainfall are highlighted in light blue shading under normal-wet climate, while light green shading is applied when climate was dry. Growth periods are highlighted in light red shading when multiple years of rainfall were the best in explaining growth under dry climate. Precipitation is color shaded for periods when both “dry” and “wet” growth-climate correlations produce the same pattern (Figs. 4 and 5). For actual correlation patterns see Fig. 6 and 7.

Significance levels from correlating “dry” (top) and “wet” (bottom) habitats from Samos Island with key periods of precipitation known to influence annual growth; correlations using 10-year moving “windows” (5th year of 10-year “window” shown). * for P < 0.05, **P < 0.01, ***P < 0.001. Asterisk in red and bold for monthly to full year rainfall signals (see text for color interpretation)

Significance levels from correlating “dry” (top) and “wet” (bottom) habitats from Zakinthos Island with key periods of precipitation known to influence annual growth; correlations using 10-year moving “windows” (5th year of 10-year “window” shown). * for P < 0.05, **P < 0.01, ***P < 0.001. Asterisk in red and bold for monthly to full year rainfall signals (see text for color interpretation)
Correlations between growth and precipitation
For each “window” of Figs. 6 and 7, the difference (%) between annual precipitation for 10-year means and its 1961–1990 mean is given as a measure of climate dryness. The integration periods of precipitation are ranked from the lowest to the highest supplied amounts as a proxy of surface cf. deeper ground water utilization; dry climate years in bold; peak drought years underlined; peak wet years in italics. Monthly to full year rainfall signals in red and bold. Thick boxes mark the main range for precipitation integration periods influencing growth as climatic dryness levels fluctuate. Color shaded P when at least “dry” or “wet” habitat trees are correlated with monthly to annual rainfall under normal-wet climate (light blue), or under dry climate (light green), and when best correlated with multiple years of rainfall under dry climate (light red). Colored precipitation when both “dry” and “wet” growth-climate correlations produce the same pattern.
Growth of “wet” habitat trees on Samos was significantly correlated with seasonal to annual precipitation, known to influence tree growth, from 1967 to 1984 (central year of 10-year running analysis “windows” reported; i.e., the 5th year) when precipitation ranged from–6% to + 7% compared to the 1961–1990 mean (light blue and green shading in Figs. 4 and 6). Thus, seasonal to annual rainfall signals recorded in tree-rings can be related to both moderate dry and moderate wet climatic conditions for “wet” habitat trees. However, when rainfall ranged between −14% and −18% below the 1961–1990 mean, only long-term rainfall supplies from the previous 2–6 years including the year of growth (4 or 6 years at P < 0.001) were significantly correlated with BAI (from 1986 until 1995; light red shading in Figs. 4 and 6). “Dry” habitat trees of Samos produced a similar pattern to their “wet” habitat counterparts during the period of drought intensification from 1987 to 1995 (Fig. 6). It was then that long periods of rainfall, mostly 4 to 6 years, could explain growth at P < 0.001 (light red shading; Figs. 4 and 6). On the other hand, during periods where precipitation recovered and reached its peak, − 4% to + 7% compared to the 1961–1990 mean; 1972–1982), growth of “dry” habitat trees was best correlated with somewhat shorter integration periods of precipitation, centered around 2-year rainfall supplies (P < 0.001). “Wet” habitat tree growth appeared to have been driven by even shorter rainfall amounts and related to December–April or October–April precipitation (P < 0.001), under this period of higher moisture availability (light blue shading Figs. 4 and 6).
For Zakinthos, both “wet” and “dry” habitat trees were significantly correlated with seasonal to annual precipitation under moderate wet (+ 5% to + 14%; 1967–1971; light blue shading), but also under extreme dry conditions (− 19% to − 27%; compared to the 1961–1990 mean; 1988–1995; light green shading; Figs. 5 and 7). This indicates that seasonal to annual rainfall signals recorded in tree rings can be related not only to moderately wet climates but can also appear under extreme dry climatic conditions for both “wet” and “dry” habitat trees. However, such signals from “wet” habitat trees appeared stronger. For instance, BAI climate correlations for “wet” habitat trees produced significant correlations at P < 0.001, e.g., for winter or winter-spring precipitation when the climax of drought occurred (− 26% and − 27% below the 1961–1990 mean; early 1990s). “Dry” habitat trees of the same time period showed significant correlations at P < 0.01, but only for February precipitation (Fig. 7).
In almost all cases under moderately dry climate conditions within 1974–1986 (− 3% to − 12% below the 1961–1990 mean; light red shading; Fig. 7), BAI from both habitats was correlated best with multiple year precipitation from 2 to 6 years. However, it was “dry” habitat trees that showed stronger BAI-climate correlations. These drivers were related to long-term rainfall supplies for almost the entire 1974–1986 period and even reached significance levels of P < 0.001 (e.g., for 3 and 4-year precipitation). “Wet” habitat trees produced similar correlation signals but at P < 0.01. On the other hand, they did not appear to be solely driven by multiple year rainfall, under this time period of normal to moderate dry climate. Seasonal rainfall (e.g., May) also appears to have played a role, for example in 1979 (light green shading; − 7% rainfall) or in 1983 (when rainfall returned to average; light blue shading; central year of 10-year analysis “window”). Such signals suggest that “wet” habitat trees are more sensitive to precipitation variability.
To summarize, average to above average precipitation (light blue shading), coincided with growth correlated significantly with monthly to seasonal precipitation signals exclusively. The signals were found in both “wet” habitat trees and for Zakinthos, mostly in “dry” habitat trees. All such cases were characterized by high growth rates, particularly when rainfall reached peak levels (Figs. 4, 5, 6, and 7).
Below average rainfall periods were connected with two opposite signals, long and short-term. Long-term signals, produced from significant correlations between growth and rainfall levels from 2 to 6 previous years, including the year of growth, appeared on both islands and for both habitat types. The significant influence of such rainfall levels on pine growth from both “wet” habitats, as well as from the “dry” habitat on Zakinthos, coincided with declining growth or with growth at the lowest. This occurred under declining precipitation or during rainfall minima (light red shading; Figs. 4, 5, 6, and 7). On the other hand, short-term signals of seasonal to annual precipitation were found to drive growth in both habitat types. Again they coincided with declining precipitation or at rainfall minima levels, followed by declining or very low growth rates. These signals also matched with growth recovery phases after decadal rainfall had reached a minimum (light green shading).
Discussion
Soil depth and moisture as the key growth drivers influenced by dryland topography
Our original hypothesizes that gullies/valleys will have deeper soils that hold significantly more moisture at their upper soil horizon compared to upper slope sites is supported by our results. Accumulation for millennia of loose soil and rock debris from weathered bedrock, together with plant litter material from higher ground, has created deep formations in the gullies/valleys investigated (Fig. 8) as a result of surface water runoff and other geophysical processes (Daws et al. 2002; Lanza et al. 2010). Whereas trees of both systems appear equally equipped to utilize deeper moisture pools (Figs. 6 and 7), the soil and weathered bedrock conditions of valley-gully systems provides pines with more moisture from winter/spring rainfall; critical for their survival and recovery during and after drought, particularly when deep moisture pools dry out. Such habitats have higher tree growth rates than upper slopes (Figs. 4 and 5) and the potential to act as refugia for P. halepensis s.l. and likely other drought adapted species under climate change.

Example of soil and tree growth conditions similar to the habitats from Samos Island. “dry”/upslope trees with shallow top soil and larger runoff potential (a). “wet”/ valley-gully trees with potential to accumulate larger runoff and debris, creating deeper top soils over deep and heavily weather bedrock with higher moisture holding capacity (b)
Topography directly contributes to water balance by controlling water flows and indirectly drives important soil variables (depth, colluvium, texture, past land uses; Vennetier et al. 2018). The 150–200 m elevational difference between the two habitat types, together with their deeper soil formations at lower elevations, are most likely the factors that explain the significantly higher moisture retained in the upper soil horizon of gully/valley habitats as a result of infiltrating rainfall runoff. Moreover, our gully/valley habitats most likely have a deeper matrix of weathered bedrock (Fig. 8). As reviewed by Dawson et al. (2020), the greater quantities of water stored in the fractures and matrix of weathered bedrock, characterized as rock moisture within the critical zone for sustaining life, can become a valuable water source that regulates plant sensitivity to drought and plant community distribution, particularly in seasonally dry climates (Klos et al. 2018; Rempe and Dietrich 2018; Szutu and Papuga 2019). All the above confirms our characterization of “wet” vs. “dry” for the systems investigated. Note that our soil moisture assessment was conducted within the dry season when moisture levels are expected to be low. Thus, the actual soil moisture differences are likely to be greater within the wet season of a year with normal rainfall. Moreover, the calcareous substrate (rendzina type soils) that characterized all habitats investigated, drains quickly. Combined with high summer temperatures, such conditions do not permit rainfall to permanently accumulate, even within “wetter” soils, causing, most likely, a depletion of surface soil moisture pools in all habitats and particularly during dry years towards the end of the dry season.
We expected that the above topographic drivers would create more favorable conditions for vegetation within “wet” habitats. The indicators evaluated included both the dominant pine vegetation and the understory. In Zakinthos, which received on average 20% more precipitation than Samos over 1961–1990, there was evidence that “wet” habitats may contain denser pine stands (50% more trees and 20% higher BA), with greater ground cover produced from both pines and the understory. Higher fracture densities in limestone may explain increased woody plant cover (Liu et al. 2019). Parameters for Samos vegetation, however, appear to be more similar between the two habitat types; approximately 10% more trees were measured in “wet” habitats, but without resulting in higher BA, when also considering the understory vegetation in mixed pine stands with olive and carob (Table 2). Hence, our original hypothesis that gullies/valleys contain denser stands, although it seems likely, requires further investigation. One additional aspect to consider would be biomass from all woody vegetation with stem circumferences < 10 cm, not considered in this study.
“Wet” habitat pines on deeper soils experience a faster growth decline and recovery
Pines on both islands have been adapted to average rainfalls of > 600 mm, falling mainly in winter, followed by a six-month dry season, typical of the Mediterranean climate. Dominant “wet” habitat pines on both Samos and Zakinthos showed significantly improved growth rates (4.5 times and 2 times higher increment), respectively, compared to their “dry” habitat counterparts, considering the entire period of investigation and particularly during normal or wet years (Figs. 4 and 5). Higher growth rates in “wet” habitats are a product of higher accumulation of soil moisture due by topography (Table 2; Fig. S3) in these water limited systems (Sarris et al. 2011; Dorman et al. 2015a).
It was hypothesized that the more drought intensifies, the more pine growth would decline for the driest habitat. Nevertheless, when precipitation drastically declined by 20–40% below the 1961–1990 average in the early 1990s, the difference in radial increment between the two habitats of contrasting moisture availability was considerably reduced. “Wet” habitat pine growth declined 1.5–2 times faster than its “dry” counterpart. On the other hand, “wet” habitat trees responded more rapidly once precipitation started to recover (Table 3). These findings create at first impression a paradox. “Wet” habitat pines could be more sensitive to rainfall decline and together more resilient than “dry” habitat pines. Veuillen et al. (2023b), focusing on P. halepensis across its distribution range, concluded that its resilience was improved by favorable conditions following drought. Similarly, favorable post-drought conditions have been found to produce a positive effect on ecosystem recovery (Jiao et al. 2021). Therefore, why have our two systems responded differently in terms of growth recovery to favorable post drought conditions?
Stand densities could play a role (Gea-Izquierdo et al. 2009; Sohn et al. 2016). The gully/valley habitats could contain denser stands as a result of their topographically improved moisture status compared to upslope sites. This higher stand density may in turn result in a negative impact under declining moisture conditions. It could create greater pressure from competition in gully/valley stands compared to upslope sites, explaining the former’s faster rate of growth decline. When moisture conditions improve, gully/valley habitats again receive more moisture and thus trees recover faster. However, for this to be valid, we would at least expect to see different responses between the two islands in relation to differences in stand density. This does not definitely appear to be the case. Zakinthos, despite being more likely to contain denser stands in valley/gully habitats compared to Samos (34.2 vs. 32.2 m2/ha BA), and despite a steeper rainfall decline (− 45% for Zakinthos vs. − 18% for Samos; from 982 mm in 1984 to 544 mm in 1991 vs. from 706 mm in 1986 to 577 mm in 1991; Figs. 4 and 5), its pines do not appear to experience a faster decline in growth compared to Samos (− 1.5 vs. − 2.3 in regression slopes; Table 3).
Under severe soil water depletion, growth can decline dramatically in trees with significantly higher growth levels than ones that survived mortality related to a drainage event followed by natural drought. This was presumed to be the result of fast growing trees being hydraulically under built for dry conditions (Levanič et al. 2011). A lower root biomass or a lower ratio of root area to leaf area for fast growing trees can, in turn, predispose trees to severe hydraulic constraints and subsequent mortality under drought (Levanič et al. 2011). Similarly, Zhang et al. (2021) suggested that favorable past climates could stimulate vegetation growth to surpass ecosystem carrying capacity, leaving an ecosystem vulnerable to climate stresses, a phenomenon known as structural overshoot. In our case, dominant “wet” habitat trees were by approximately 5 m taller than their “dry” habitat counterparts (Table 2), suggesting better developed crowns and foliage (above ground biomass). Such features could place them in a more stressful situation under drought intensification. Thus, it cannot be ruled out that our “wet” habitat trees experienced higher evapotranspiration, particularly if they retain less root biomass or lower ratios of root area to leaf area compared to trees on slopes.
Nevertheless, our results do not support the less root biomass hypothesis. By correlating tree-ring width with precipitation that determine groundwater availability, it is possible to indirectly determine whether pines are deep rooted and the origin of the moisture resources their roots have access to (Sarris and Mazza 2021). Signals of long integration periods of precipitation recorded in annual rings from trees not connected to the water table are associated with deeper moisture utilization by pines and correspond to deep root systems (Sarris et al. 2007, 2013; the latter for carbon and oxygen isotope confirmation). The climate signals recorded in annual rings suggest that trees from both habitat types had the ability to utilize deep moisture resources accumulated below ground from precipitation of past years. The particular moisture resources may have been the result of four to six years of rainfall on Samos, coinciding with the climax of drought from the early to mid-1990s (P < 0.001; light red shade; Fig. 6), and by three to four years of rainfall on Zakinthos in periods of moderate to severe rainfall decline, which occurred in the late 1970s or mid to late 1980s (P < 0.001; light red shade; Fig. 7). These findings also contradict our original hypothesis that, under drought, the driest habitat would exhibit a higher dependence on deep ground moisture resources for annual growth. Pines in valleys and depressions appear as deep rooted as pines growing upslope, making it highly unlikely that less root biomass can alone explain the faster growth decline and recovery of “wet” habitat trees under precipitation decrease or increase.
Hence, as drought intensifies and as long as deeper moisture pools are available, both “wet” and “dry” habitat trees appear to be equally equipped to exploit these resources. Note, that our sites are considered as well supplied in winter rainfall in terms of P. halepensis s.l. distribution across the Mediterranean (precipitation > 600 mm, 1986–2000 data; Mauri et al. 2016), permitting sufficient rainfall infiltration in deeper ground layers during rainy periods. In such cases, roots of some, but not all plants, follow water depletion down the soil/bedrock layers (Bledsoe et al. 2014). Pines in Mediterranean climates have been known to track water. As the seasonal climate becomes drier and the overlying substrate is depleted of moisture, progressively deeper water sources, including weathered bedrock, can be exploited to a depth of several meters, enabling trees to escape desiccation (Rose et al. 2003; Sarris et al. 2013). Plants commonly access such rock moisture pools from bedrock, which provides an important water source during drought, as reflected by tree water status and dieback phenomena (McCormick et al. 2021; Nardini et al. 2021).
However, this does not seem to be the case under normal or wet climates, where surface moisture supplies provided to trees within their growing season appear to drive growth. Moisture utilization signals related to surface monthly to seasonal rainfall appeared more frequently for Samos “wet” habitat trees, while a similar tendency appeared for Zakinthos pines under periods of average or increased precipitation (up to 7% above the 1961–1990 average for Samos, up to 14% for Zakinthos; light blue shading; Figs. 6 and 7). These signals were best related to rainfall supplies of March up to September–August for Samos trees and April or May up to September–August for trees from Zakinthos. “Dry” habitat trees during the same normal to rainy years, clearly took up water supplied by longer periods of rainfall provided by two to three-years on Samos (Fig. 6). In Zakinthos, either signals were less strongly related to seasonal moisture supplies (P < 0.01 for April–May rainfall vs. P < 0.001 for April rainfall signals produced by “wet” habitat trees in the early 1970s), or no signals were captured during the same rainy time periods (early 1980s; Fig. 7).
All the evidence suggests that the dominant P. halepensis s.l. trees growing in the valleys/gullies have a wider range of options in terms of utilizing moisture sources. Under normal or wet climate, they can rely on upper soil moisture pools supplied by monthly or seasonal rainfall within the growing season. Such resources permit high growth rates. However, these pools can also be rapidly depleted under drought intensification, particularly under high evapotranspiration, which possibly explains the rapid growth decline for “wet” habitat trees. Under a dry climate, these trees can still utilize deeper moisture pools, most likely “rock moisture” stored in weathered bedrock (Rempe and Dietrich 2018) supplied by past rainfall events, as is the case for their counterparts growing on upslope sites. However, low supplies of water and nutrients likely reduces growth to the level of “dry” habitat trees. Such shifts in water utilization from different soil horizons is typical of plants with dimorphic roots in Mediterranean-type ecosystems (Dawson and Pate 1996).
Under rainfall recovery, a key moisture source again becomes available, supplied by monthly and seasonal rainfall infiltration, particularly if deeper “rock moisture” pools run dry. During such conditions, trees on deeper soils have a key advantage. Deep soils, together with the usually underlying co-occurring thicker and heavily weathered bedrock formations, provide increased moisture holding capacities that can store more of these surface moisture inputs, amplified by runoff accumulating from higher ground. In contrast, “dry” habitats with shallow soils on top of lightly weathered bedrock lack such storage capacity (Fig. 8). As a result, surface moisture can become more easily lost to evaporation. Water content of soil on a slope is greatest at the base and becomes progressively less towards the top (Daws et al. 2002). Our “dry” habitat trees were all selected at upper to mid-slope, i.e., at the driest part of a slope. Such soil captured less moisture from any winter or spring precipitation, still available during severely dry years or during rainfall recovery, compared to soil (and it’s just underlying geological formations) on valleys/gullies. This became evident in “wet” habitats of both Samos and Zakinthos, boosting growth recovery. As exemplified by “wet” habitat trees of Zakinthos, pines showed 2.5 times faster growth recovery than their “dry” habitat counterparts (Table 3; Fig. 5) by relying on February rainfall (decade centered in 1994; light green shading; Fig. 7). “Dry” habitat trees on Zakinthos, at the same time, produced water utilization signals of seasonal to annual (December–February to January–December). This suggests that they depended on longer rainfall supply periods for growth because their shallower soils did not permit an as efficient exploitation of monthly rainfall as valley/gully trees. The outcome was poor growth recovery rates.
There have been increasing examples over the past decade across the Mediterranean region of similar findings, with pines shifting to surface moisture pools during drought intensification. Dorman et al. (2015b), for P. halepensis in central Israel, found that in habitats classified between mesic to xeric, pines depended on deeper moisture resources in rainy years. Although after severe drought, deeper moisture pools most likely became depleted, and these pines shifted their water uptake to surface moisture. Mazza et al. (2018) reported that P. nigra subsp. laricio in Calabria, within the species mesic to xeric distribution range, that under a dry climate, approximately one-third of the trees significantly reduced their capacity to utilize deeper ground moisture and depended mostly on surface rainfall provided by summer rainfall for growth (see also Mazza and Sarris (2021) for P. pinea). Therefore, we are possibly confronted with a situation that deserves further attention, since it appears widespread across different pine species. However, the preceding findings have been mostly linked with poor growth rates from poor habitats, resembling responses produced by our “dry” habitat trees. In this study, we were able to detect for the first time that a shift towards surface moisture utilization can also be linked with growth recovery phases when more favorable habitats are included.
Role of soil depth in delaying the retreat of P. halepensis s.l. under climate change at its drought tolerance limits
Soil depth, by influencing rooting depth, together with the degree of exposure to the sun’s radiation, has been considered to be among the prime factors that determines pine desiccation or survival (Dorman et al. 2015c). Deep soils, for instance, may influence tree architecture, prohibiting deep rooting into the bedrock, which in turn, can lead to elevated vulnerability in times of drought. This has been suggested for failures in pine afforestation in arid climates (average rainfall < 300 mm) and on south-facing slopes with sparse pines, particularly when the entire soil profile dries out, while water stored in rock layers is beyond the reach of roots (Fensham and Fairfax 2007; Dorman et al. 2015c).
Our findings, however, suggest that valley/gully trees take better advantage of surface moisture inputs from late winter and spring rainfall compared to trees on slopes. Such inputs are known to drive early wood formation (Sarris et al. 2013). Taking better advantage of late winter–early spring rainfalls, did not permit growth levels of dominant pines from “wet” habitats to fall below those of “dry” habitat trees (Figs. 4 and 5). Trees on upper slope sites apparently lack such “buffering effects” of deep soils, and heavily weathered bedrock formations, most likely, that can store additional “rock moisture” (Fig. 8; Rempe and Dietrich 2018; McCormick et al. 2021; Luo et al. 2023). Thus, in conditions where chronic drought has depleted deeper ground moisture pools, but still some late winter- early spring rainfall occurs, habitats with greater moisture holding capacities may have an advantage for tree survival under severe drought. Indeed, drought- driven incidents of desiccation have been documented for vegetation on slopes in Samos, irrespective of life form, i.e., for macchia-type sclerophyllous woodland as well as low elevation pine forests (Körner et al. 2005; Sarris et al. 2007).
A limitation of our study was the scarcity of natural P. halepensis and P. brutia populations at gully/valley sites in the islands investigated, out of which more “dry”- “wet” habitat systems could be studied to provide higher tree numbers at the mature life-stage. Human disturbances, and particularly wildfires, which spread rapidly in the habitat produced by these fire-adapted pines (Moreno et al. 2021) could make such efforts more difficult in the near future. Using more sites, tree numbers and species, will be needed though for future research efforts investigating the impact of topography in shaping the future distribution of drought- adapted species under climate change. Towards this direction, apart from Vennetier et al. (2018), there is more recent evidence in support of our results. Juniperus thurifera growing in valley bottoms of Spain experience conditions that buffer them against intensified aridification, compared to junipers growing on steep slopes (Camarero et al. 2023).
Therefore, it is likely that “wet” habitats may considerably delay the retreat of P. halepensis s.l. within its natural distribution range compared to what some models project under global warming (Thuiller 2003; Keenan et al. 2011) if the effects of topography are considered. It is possible that this is also the case for other species adapted to semi-arid conditions. Such effects should be considered in modeling the future biogeography of P. halepensis s.l. or of similar species growing in semiarid conditions. They can also be used for detecting potential refugia (Christmas et al. 2016) at species rear-end distribution limits. More favorable habitats for growth should provide the highest resistance to the future retreat of pine populations towards colder/wetter regions. This is as long as the taxa inside such favorable sites can be sustained within their drought tolerance limits and not outcompeted by better drought adapted species (Rodriguez-Ramirez et al. 2017). The precipitation decline in our ecosystems falls within the mid and upper range of projected reductions for the Mediterranean region towards the end of the twenty-first century (based on climate model studies using 1961–1990 as a reference period for climate projections; IPCC 2007; Giorgi and Lionello 2008). Thus, our study provided a unique test for simulating the future responses of P. halepensis s.l. to climate change and for evaluating the potential of “wet” habitats as refugia.
In dense stands, increased water stress because of drought may increase the vulnerability and susceptibility of trees to mortality (Allen and Breshears 1998; Guarin and Taylor 2005; Gea-Izquierdo et al. 2009). Thinning stands, and thus reducing density, is one management practice that could improve resistance for individual trees to drought stress (Laurent et al. 2003; Moreno and Cubera 2008; Sohn et al. 2016), as has also been suggested for P. halepensis (Helluy et al. 2020; Manrique-Alba et al. 2020). Increasing rooting depth, where deep moisture pools might exist, and removing individual pines that have low productivity and shallow root systems can be a management option for increasing forest resilience to drought (Sarris and Mazza 2021). For Pinus pinaster, it has been shown that stands of lower density utilize deeper moisture resources provided by rainfall of past years compared to denser stands (Mazza et al. 2014). After thinning, trees may develop more extensive root systems over time, hence increasing their ability to extract soil water during and after drought periods, compared to trees in unthinned stands (Laurent et al. 2003; Raz-Yaseef et al. 2010).
However, our findings suggest that deep soils, and most likely the heavily weathered bedrock formations underneath, are key for increasing survival for natural P. halepensis s.l populations under a drier climate. They are in agreement with Vennetier et al. (2018), which state the importance of deep, rich soils and a favorable local topography for good Aleppo pine productivity, even on the driest, hottest sites in southern France. Such findings question whether selective thinning of stands would be necessary for productivity improvement and chances of survival under climatic change, if reducing density would also accelerate soil erosion, particularly under increasing storm intensities and torrential rain events. This would, in turn, lead to decreasing surface moisture storage capacity of soils. Such thinning practices for climatic change adaptation have also been questioned for holm oak stands (Quercus Ilex L.) in the Iberian Peninsula (Gea-Izquierdo et al. 2009) and for P. halepensis populations in Israel (Dorman et al. 2015b, c). Thus, the chances of increasing erosion and heat stress on soil due to thinning (Dorman et al. 2015b, c) should also be considered, as well as the role of forests and their soils as C sources and sinks (Govind et al. 2009; Waring et al. 2020). Hence, before any thinning is carried out, a dendroecological study, as provided here, would be important for understanding the particular ecosystem in terms of rooting structure and function, and for evaluating the role of soil depth within the system.
Conclusion
The results of this study indicate that topography and climate will be key drivers for the future distribution of P. halepensis s.l. in the Mediterranean region. These factors determine water movement, the accumulation of debris and nutrients, soil depth and bedrock weathering, in turn regulating moisture holding capacity within the Critical Zone for sustaining life. If, under global warming, moderate drought conditions persist in the Mediterranean basin (–25% below the mean annual rainfall of 1961–1990), water limited tree species because of summer drought growing on slopes and in valleys-land depressions, are likely to respond similarly as the trees in this study in terms of ground water utilization. This for sites well-supplied by winter rainfall but with no permanent water. However, if severe drought conditions persist (–40% below the mean annual rainfall of 1961–1990), pine forests of valleys/gullies with deeper soils are more likely to still occupy the habitats of their current biogeographical distribution, although productivity will most likely be reduced close to levels of trees currently growing on much drier upslope sites.
In the dilemma of whether to reduce stand density or to protect soil from erosion to enhance low elevation pine resilience to drought, our findings stress the importance of the latter. Soil depth is a key feature for the long- term buffering of such affects. Therefore, thinning should be applied with great caution and after evaluating tree root structure, its functioning, together with soil depth and bedrock weathering to account for the key resources needed by trees to overcome drought stress.
References
Allen CD, Breshears DD (1998) Drought-induced shift of a forest-woodland ecotone: rapid landscape response to climate variation. Proc Natl Acad Sci USA 95(25):14839–14842. https://doi.org/10.1073/pnas.95.25.14839
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim JH, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259(4):660–684. https://doi.org/10.1016/j.foreco.2009.09.001
Améztegui A, Brotons L, Coll L (2010) Land-use changes as major drivers of mountain pine (Pinus uncinata Ram.) expansion in the Pyrenees. Glob Ecol Biogeogr 19(5):632–641. https://doi.org/10.1111/j.1466-8238.2010.00550.x
Améztegui A, Coll L, Brotons L, Ninot JM (2016) Land-use legacies rather than climate change are driving the recent upward shift of the mountain tree line in the Pyrenees. Glob Ecol Biogeogr 25(3):263–273. https://doi.org/10.1111/geb.12407
Balzan MV, Hassoun AER, Aroua N, Baldy V, Bou Dagher M, Branquinho C, Dutay J-C, El Bour M, Médail F, Mojtahid M, Morán-Ordóñez A, Roggero PP, Rossi Heras S, Schatz B, Vogiatzakis IN, Zaimes GN, Ziveri P (2020) Ecosystems. In: Cramer W, Guiot J, Marini K (eds) Climate and environmental change in the mediterranean basin–Current situation and risks for the future first mediterranean assessment report. Union for the Mediterranean, Plan Bleu, UNEP/MAP, Marseille, pp 151. 10.5281/zenodo.4768833
Biondi F, Qeadan F (2008) A theory-driven approach to tree-ring standardization: defining the biological trend from expected basal area increment. Tree Ring Res 64(2):81–96. https://doi.org/10.3959/2008-6.1
Bledsoe CS, Allen MF, Southworth D (2014) Beyond mutualism: complex mycorrhizal interactions. In: Lüttge U, Beyschlag W, Cushman J (eds) Progress in botany. Springer, Heidelberg, pp 311–334. https://doi.org/10.1007/978-3-642-38797-5_10
Bodin J, Badeau V, Bruno E, Cluzeau C, Moisselin JM, Walther GR, Dupouey JL (2013) Shifts of forest species along an elevational gradient in Southeast France: climate change or stand maturation? J Veg Sci 24(2):269–283. https://doi.org/10.1111/j.1654-1103.2012.01456.x
Camarero JJ, Gazol A, Valeriano C, Pizarro M, González de Andrés E (2023) Topoclimatic modulation of growth and production of intra-annual density fluctuations in Juniperus thurifera. Dendrochronologia 82:126145. https://doi.org/10.1016/j.dendro.2023.126145
Chen XF, Chen JM, An SQ, Ju WM (2007) Effects of topography on simulated net primary productivity at landscape scale. J Environ Manage 85(3):585–596. https://doi.org/10.1016/j.jenvman.2006.04.026
Cherif S, Doblas-Miranda E, Lionello P, Borrego C, Giorgi F, Iglesias A, Jebari S, Mahmoudi E, Moriondo M, Pringault O, Rilov G, Somot S, Tsikliras A, Vila M, Zittis G (2020) Drivers of change. In: Cramer W, Guiot J, Marini K (eds) Climate and environmental change in the mediterranean basin—current situation and risks for the future. First Mediterranean Assessment Report. Union for the Mediterranean, Plan Bleu, UNEP/MAP, Marseille, France, p 128. https://doi.org/10.5281/zenodo.4768833
Christensen KI (1997) Gymnospermae. In: Strid A, Tan K (eds) Flora hellenica 1, Königstein, pp 1–17
Christmas MJ, Breed MF, Lowe AJ (2016) Constraints to and conservation implications for climate change adaptation in plants. Conserv Genet 17(2):305–320. https://doi.org/10.1007/s10592-015-0782-5
Daws MI, Mullins CE, Burslem DFRP, Paton SR, Dalling JW (2002) Topographic position affects the water regime in a semideciduous tropical forest in Panamá. Plant Soil 238(1):79–89. https://doi.org/10.1023/A:1014289930621
Dawson TE, Pate JS (1996) Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: a stable isotope investigation. Oecologia 107(1):13–20. https://doi.org/10.1007/BF00582230
Dawson TE, Hahm WJ, Crutchfield-Peters K (2020) Digging deeper: what the critical zone perspective adds to the study of plant ecophysiology. New Phytol 226(3):666–671. https://doi.org/10.1111/nph.16410
Denslow JS (1995) Disturbance and diversity in tropical rain forests: the density effect. Ecol Appl 5(4):962–968. https://doi.org/10.2307/2269347
Dolph KL, Mori SR, Oliver WW (1995) Long-term response of old-growth stands to varying levels of partial cutting in the eastside pine type. West J Appl for 10(3):101–108. https://doi.org/10.1093/wjaf/10.3.101
Dorman M, Perevolotsky A, Sarris D, Svoray T (2015a) Amount vs. temporal pattern: on the importance of intra-annual climatic conditions on tree growth in a dry environment. J Arid Environ 118:65–68. https://doi.org/10.1016/j.jaridenv.2015.03.002
Dorman M, Perevolotsky A, Sarris D, Svoray T (2015b) The effect of rainfall and competition intensity on forest response to drought: lessons learned from a dry extreme. Oecologia 177(4):1025–1038. https://doi.org/10.1007/s00442-015-3229-2
Dorman M, Svoray T, Perevolotsky A, Moshe Y, Sarris D (2015c) What determines tree mortality in dry environments? A multi-perspective approach. Ecol Appl 25(4):1054–1071. https://doi.org/10.1890/14-0698.1
EUFORGEN (2009a) Distribution map of Alepo pine (Pinus halepensis). https://www.euforgen.org/species/pinus-halepensis/. Accessed 20 Aug 2021
EUFORGEN (2009b) Distribution map of Brutia pine (Pinus brutia). https://www.euforgen.org/species/pinus-brutia/. Accessed 20 Aug 2021
Fady B, Semerci H, Vendramin GG (2003) EUFORGEN Technical Guidelines for genetic conservation and use for Aleppo pine (Pinus halepensis) and Brutia pine (Pinus brutia). International Plant Genetic Resources Institute, Rome, p 6
Fensham RJ, Fairfax RJ (2007) Drought-related tree death of savanna eucalypts: species susceptibility, soil conditions and root architecture. J Veg Sci 18(1):71–80. https://doi.org/10.1658/1100-9233(2007)18[71:dtdose]2.0.co;2
Gazol A, Oliva J, Valeriano C, Colangelo M, Camarero JJ (2022) Mixed pine forests in a hotter and drier world: the great resilience to drought of Aleppo pine benefits it over other coexisting pine species. Front for Glob Change 5:899425. https://doi.org/10.3389/ffgc.2022.899425
Gea-Izquierdo G, Martin-Benito D, Cherubini P, Cañellas I (2009) Climate-growth variability in Quercus ilex L. West Iberian open woodlands of different stand density. Ann for Sci 66:802. https://doi.org/10.1051/forest/2009080
Giorgi F, Lionello P (2008) Climate change projections for the Mediterranean Region. Glob Planet Change 63(2–3):90–104. https://doi.org/10.1016/j.gloplacha.2007.09.005
Govind A, Chen JM, Ju WM (2009) Spatially explicit simulation of hydrologically controlled carbon and nitrogen cycles and associated feedback mechanisms in a boreal ecosystem. J Geophys Res 14:G02006. https://doi.org/10.1029/2008JG000728
Guarín A, Taylor AH (2005) Drought triggered tree mortality in mixed conifer forests in Yosemite National Park, California, USA. For Ecol Manag 218(1–3):229–244. https://doi.org/10.1016/j.foreco.2005.07.014
Guiot J, Cramer W (2016) Climate change: The 2015 Paris agreement thresholds and Mediterranean basin ecosystems. Science 354(6311):465–468. https://doi.org/10.1126/science.aah5015
Haavik LJ, Stephen FM, Fierke MK, Salisbury VB, Leavitt SW, Billings SA (2008) Dendrochronological parameters of northern red oak (Quercus rubra L. (Fagaceae)) Infested With Red Oak Borer (Enaphalodes rufulus (Haldeman) (Coleoptera: Cerambycidae)). For Ecol Manag 255(5–6):1501–1509. https://doi.org/10.1016/j.foreco.2007.11.005
Helluy M, Prévosto B, Cailleret M, Fernandez C, Balandier P (2020) Competition and water stress indices as predictors of Pinus halepensis Mill. radial growth under drought. For Ecol Manag 460:117877. https://doi.org/10.1016/j.foreco.2020.117877
Hoerling M, Eischeid J, Perlwitz J, Quan XW, Zhang T, Pegion P (2012) On the increased frequency of Mediterranean drought. J Clim 25(6):2146–2161. https://doi.org/10.1175/jcli-d-11-00296.1
Huang W, Zhang LP, Furumi S, Muramatsu K, Daigo M, Li PX (2010) Topographic effects on estimating net primary productivity of green coniferous forest in complex terrain using Landsat data: a case study of Yoshino Mountain, Japan. Int J Remote Sens 31(11):2941–2957. https://doi.org/10.1080/01431160903140829
Innes JL (1992) Forest decline. Prog Phys Geogr 16:1–64. https://doi.org/10.1177/030913339201600101
IPCC (2007) Climate change 2007: The physical science basis contribution of working group I. In: Solomon S, Qin D, Manning M, Chen Z et al (eds) Fourth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge
IPCC (2023) Weather and climate extreme events in a changing climate. In: Climate change 2021—the physical science basis: working group I contribution to the sixth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge. pp 1513–1766. https://doi.org/10.1017/9781009157896.013
Jenkins MA, Pallardy SG (1995) The influence of drought on red oak group species growth and mortality in the Missouri Ozarks. Can J for Res 25(7):1119–1127. https://doi.org/10.1139/x95-124
Jiao T, Williams CA, De Kauwe MG, Schwalm CR, Medlyn BE (2021) Patterns of post-drought recovery are strongly influenced by drought duration, frequency, post-drought wetness, and bioclimatic setting. Glob Chang Biol 27(19):4630–4643. https://doi.org/10.1111/gcb.15788
Kazakis G, Ghosn D, Vogiatzakis IN, Papanastasis VP (2007) Vascular plant diversity and climate change in the alpine zone of the Lefka Ori, Crete. Biodivers Conserv 16(6):1603–1615. https://doi.org/10.1007/s10531-006-9021-1
Keenan T, Maria Serra J, Lloret F, Ninyerola M, Sabate S (2011) Predicting the future of forests in the Mediterranean under climate change, with niche- and process-based models: CO2 matters! Glob Change Biol 17(1):565–579. https://doi.org/10.1111/j.1365-2486.2010.02254.x
Klein Tank AMG, Wijngaard JB, Können GP, Böhm R, Demarée G, Gocheva A, Mileta M, Pashiardis S, Hejkrlik L, Kern-Hansen C, Heino R, Bessemoulin P, Müller-Westermeier G, Tzanakou M, Szalai S, Pálsdóttir T, Fitzgerald D, Rubin S, Capaldo M, Maugeri M, Leitass A, Bukantis A, Aberfeld R, van Engelen AFV, Forland E, Mietus M, Coelho F, Mares C, Razuvaev V, Nieplova E, Cegnar T, Antonio López J, Dahlström B, Moberg A, Kirchhofer W, Ceylan A, Pachaliuk O, Alexander LV, Petrovic P (2002) Daily dataset of 20th-century surface air temperature and precipitation series for the European Climate Assessment. Int J Climatol 22(12):1441–1453. https://doi.org/10.1002/joc.773
Klos PZ, Goulden ML, Riebe CS, Tague CL, O’Geen AT, Flinchum BA, Safeeq M, Conklin MH, Hart SC, Berhe AA, Hartsough PC, Holbrook WS, Bales RC (2018) Subsurface plant-accessible water in mountain ecosystems with a Mediterranean climate. Wires Water 5(3):e1277. https://doi.org/10.1002/wat2.1277
Körner C, Sarris D, Christodoulakis D (2005) Long-term increase in climatic dryness in the East-Mediterranean as evidenced for the island of Samos. Reg Environ Change 5(1):27–36. https://doi.org/10.1007/s10113-004-0091-x
Lanza NL, Meyer GA, Okubo CH, Newsom HE, Wiens RC (2010) Evidence for debris flow gully formation initiated by shallow subsurface water on Mars. Icarus 205(1):103–112. https://doi.org/10.1016/j.icarus.2009.04.014
Laurent M, Antoine N, Joël G (2003) Effects of different thinning intensities on drought response in Norway spruce (Picea abies (L.) Karst). For Ecol Manag 183(1–3):47–60. https://doi.org/10.1016/S0378-1127(03)00098-7
Lenoir J, Gégout JC, Marquet PA, de Ruffray P, Brisse H (2008) A significant upward shift in plant species optimum elevation during the 20th century. Science 320(5884):1768–1771. https://doi.org/10.1126/science.1156831
Levanič T, Căter M, McDowell NG (2011) Associations between growth, wood anatomy, carbon isotope discrimination and mortality in a Quercus robur forest. Tree Physiol 31(3):298–308. https://doi.org/10.1093/treephys/tpq111
Linares JC, Tíscar PA (2011) Buffered climate change effects in a Mediterranean pine species: range limit implications from a tree-ring study. Oecologia 167(3):847–859. https://doi.org/10.1007/s00442-011-2012-2
Liphschitz N, Lev-Yadun S, Rosen E, Waisel Y (1984) The annual rhythm of activity of the lateral meristems (cambium and phellogen) in Pinus halepensis Mill. and Pinus pinea L. IAWA J 5(4):263–274. https://doi.org/10.1163/22941932-90000413
Liu HY, Jiang ZH, Dai JX, Wu XC, Peng J, Wang HY, Meersmans J, Green SM, Quine TA (2019) Rock crevices determine woody and herbaceous plant cover in the Karst critical zone. Sci China Earth Sci 62(11):1756–1763. https://doi.org/10.1007/s11430-018-9328-3
Luo ZB, Fan J, Shao MA, Yang Q, Gan M (2023) Weathered bedrock converts hydrological processes in loess hilly-gully critical zone. J Hydrol 625:130112. https://doi.org/10.1016/j.jhydrol.2023.130112
Manrique-Alba À, Beguería S, Molina AJ, González-Sanchis M, Tomàs-Burguera M, del Campo AD, Colangelo M, Camarero JJ (2020) Long-term thinning effects on tree growth, drought response and water use efficiency at two Aleppo pine plantations in Spain. Sci Total Environ 728:138536. https://doi.org/10.1016/j.scitotenv.2020.138536
Mauri A, Di Leo M, de Rigo D, Caudullo G (2016) Pinus halepensis and Pinus brutia in Europe: distribution, habitat, usage and threats In: San-Miguel-Ayanz J, de Rigo D, Caudullo G, Houston Durrant T, Mauri A (eds) European Atlas of Forest Tree Species. Publications Office of the European Union, Luxembourg, p e0166b8+
Mazza G, Sarris D (2021) Identifying the full spectrum of climatic signals controlling a tree species’ growth and adaptation to climate change. Ecol Indic 130:108109. https://doi.org/10.1016/j.ecolind.2021.108109
Mazza G, Cutini A, Manetti MC (2014) Influence of tree density on climate-growth relationships in a Pinus pinaster Ait. forest in the northern mountains of Sardinia (Italy). Iforest 8(4):456–463. https://doi.org/10.3832/ifor1190-007
Mazza G, Sarris D, Chiavetta U, Ferrara RM, Rana G (2018) An intra-stand approach to identify intra-annual growth responses to climate in Pinus nigra subsp. laricio Poiret trees from southern Italy. For Ecol Manag 425:9–20. https://doi.org/10.1016/j.foreco.2018.05.029
McCormick EL, Dralle DN, Hahm WJ, Tune AK, Schmidt LM, Chadwick KD, Rempe DM (2021) Widespread woody plant use of water stored in bedrock. Nature 597(7875):225–229. https://doi.org/10.1038/s41586-021-03761-3
McElhinny C, Gibbons P, Brack C, Bauhus J (2005) Forest and woodland stand structural complexity: Its definition and measurement. For Ecol Manag 218(1–3):1–24. https://doi.org/10.1016/j.foreco.2005.08.034
Mitchell TD, Hulme M (2000) A country-by-country analysis of past and future warming rates Tyndall Centre Internal Report, No1, November UEA, Norwich
Moreno G, Cubera E (2008) Impact of stand density on water status and leaf gas exchange in Quercus ilex. For Ecol Manag 254(1):74–84. https://doi.org/10.1016/j.foreco.2007.07.029
Moreno JM, Morales-Molino C, Torres I, Arianoutsou M (2021) Fire in Mediterranean pine forests: past, present and future. In: Pines and their mixed forest ecosystems in the Mediterranean Basin. Springer International Publishing, pp 421–456. https://doi.org/10.1007/978-3-030-63625-8_21
Nardini A, Petruzzellis F, Marusig D, Tomasella M, Natale S, Altobelli A, Calligaris C, Floriddia G, Cucchi F, Forte E, Zini LC (2021) Water ‘on the rocks’: a summer drink for thirsty trees? New Phytol 229(1):199–212. https://doi.org/10.1111/nph.16859
Palombo C, Chirici G, Marchetti M, Tognetti R (2013) Is land abandonment affecting forest dynamics at high elevation in Mediterranean Mountains more than climate change? Plant biosystem. Int J Deal Aspects Plant Biol 147(1):1–11. https://doi.org/10.1080/11263504.2013.772081
Peñuelas J, Boada M (2003) A global change-induced biome shift in the Montseny Mountains (NE Spain). Glob Change Biol 9(2):131–140. https://doi.org/10.1046/j.1365-2486.2003.00566.x
Peñuelas J, Ogaya R, Boada M, Jump AS (2007) Migration, invasion and decline: changes in recruitment and forest structure in a warming-linked shift of European beech forest in Catalonia (NE Spain). Ecography 30(6):829–837. https://doi.org/10.1111/j.2007.0906-7590.05247.x
Pounds JA, Fogden MPL, Campbell JH (1999) Biological response to climate change on a tropical mountain. Nature 398:611–615. https://doi.org/10.1038/19297
Príncipe A, Matos P, Sarris D, Gaiola G, do Rosário L, Correia O, Branquinho C (2019) In Mediterranean drylands microclimate affects more tree seedlings than adult trees. Ecol Indic 106:105476. https://doi.org/10.1016/j.ecolind.2019.105476
Raz-Yaseef N, Rotenberg E, Yakir D (2010) Effects of spatial variations in soil evaporation caused by tree shading on water flux partitioning in a semi-arid pine forest. Agric for Meteor 150(3):454–462. https://doi.org/10.1016/j.agrformet.2010.01.010
Rempe DM, Dietrich WE (2018) Direct observations of rock moisture, a hidden component of the hydrologic cycle. Proc Natl Acad Sci USA 115(11):2664–2669. https://doi.org/10.1073/pnas.1800141115
Rinn F (2003) TSAP-Win professional, time series analysis and presentation for dendrochronology and related applications, Version 030 for Microsoft Windows, Quick Reference. Rinntech, Heidelberg
Rodriguez-Ramirez N, Santonja M, Baldy V, Ballini C, Montès N (2017) Shrub species richness decreases negative impacts of drought in a Mediterranean ecosystem. J Veg Sci 28(5):985–996. https://doi.org/10.1111/jvs.12558
Rose KL, Graham RC, Parker DR (2003) Water source utilization by Pinus jeffreyi and Arctostaphylos patula on thin soils over bedrock. Oecologia 134(1):46–54. https://doi.org/10.1007/s00442-002-1084-4
Sala OE, Chapin FS 3rd, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287(5459):1770–1774. https://doi.org/10.1126/science.287.5459.1770
Sarris D, Christodoulakis D, Körner C (2007) Recent decline in precipitation and tree growth in the eastern Mediterranean. Glob Change Biol 13(6):1187–1200. https://doi.org/10.1111/j.1365-2486.2007.01348.x
Sarris D, Christodoulakis D, Körner C (2011) Impact of recent climatic change on growth of low elevation eastern Mediterranean forest trees. Clim Change 106(2):203–223. https://doi.org/10.1007/s10584-010-9901-y
Sarris D, Siegwolf R, Körner C (2013) Inter- and intra-annual stable carbon and oxygen isotope signals in response to drought in Mediterranean pines. Agric for Meteor 168:59–68. https://doi.org/10.1016/j.agrformet.2012.08.007
Sarris D, Mazza G (2021) Mediterranean pine root systems under drought. In: Ne’eman G, Osem Y (eds) Pines and their mixed forest ecosystems in the Mediterranean Basin. Managing Forest Ecosystems vol 38. Springer International Publishing, pp 129–140. https://doi.org/10.1007/978-3-030-63625-8_8
Sohn JA, Hartig F, Kohler M, Huss J, Bauhus J (2016) Heavy and frequent thinning promotes drought adaptation in Pinus sylvestris forests. Ecol Appl 26(7):2190–2205. https://doi.org/10.1002/eap.1373
Somot S, Sevault F, Déqué M, Crépon M (2008) 21st century climate change scenario for the Mediterranean using a coupled atmosphere–ocean regional climate model. Glob Planet Change 63(2–3):112–126. https://doi.org/10.1016/j.gloplacha.2007.10.003
Stephenson NL (1990) Climatic control of vegetation distribution: the role of the water balance. Am Nat 135(5):649–670. https://doi.org/10.1086/285067
Szutu DJ, Papuga SA (2019) Year-round transpiration dynamics linked with deep soil moisture in a warm desert shrubland. Water Resour Res 55(7):5679–5695. https://doi.org/10.1029/2018wr023990
Thuiller W (2003) BIOMOD–optimizing predictions of species distributions and projecting potential future shifts under global change. Glob Change Biol 9(10):1353–1362. https://doi.org/10.1046/j.1365-2486.2003.00666.x
Vennetier M, Ripert C, Rathgeber C (2018) Autecology and growth of Aleppo pine (Pinus halepensis Mill.): a comprehensive study in France. For Ecol Manag 413:32–47. https://doi.org/10.1016/j.foreco.2018.01.028
Veuillen L, Prévosto B, Alfaro-Sánchez R, Badeau V, Battipaglia G, Beguería S, Bravo F, Boivin T, Camarero JJ, Čufar K, Davi H, De Luis M, Del Campo A, Del Rio M, Di Filippo A, Dorman M, Durand-Gillmann M, Ferrio JP, Gea-Izquierdo G, González-Sanchis M, Granda E, Guibal F, Gutierrez E, Helluy M, El Khorchani A, Klein T, Levillain J, Linares JC, Manrique-Alba A, Martinez Vilalta J, Molina AJ, Moreno-Gutiérrez C, Nicault A, Olivar J, Papadopoulos A, Perevolotsky A, Rathgeber C, Ribas M, Ripullone F, Ruano I, Saintonge FX, Sánchez-Salguero R, Sarris D, Serra-Maluquer X, Svoray T, Tallieu C, Valor T, Vennetier M, Voltas J, Cailleret M (2023a) Pre- and post-drought conditions drive resilience of Pinus halepensis across its distribution range. Agric for Meteor 339:109577. https://doi.org/10.1016/j.agrformet.2023.109577
Veuillen L, Prévosto B, Zeoli L, Pichot C, Cailleret M (2023b) Pinus halepensis and P. brutia provenances present similar resilience to drought despite contrasting survival, growth, cold tolerance and stem quality: Insights from a 45 year-old common garden experiment. For Ecol Manag 544:121146. https://doi.org/10.1016/j.foreco.2023.121146
Vicente-Serrano SM, Lasanta T, Gracia C (2010) Aridification determines changes in forest growth in Pinus halepensis forests under semiarid Mediterranean climate conditions. Agric for Meteor 150(4):614–628. https://doi.org/10.1016/j.agrformet.2010.02.002
Waring B, Neumann M, Prentice IC, Adams M, Smith P, Siegert M (2020) Forests and decarbonization–roles of natural and planted forests. Front for Glob Change 3:58. https://doi.org/10.3389/ffgc.2020.00058
Wyckoff PH, Bowers R (2010) Response of the prairie–forest border to climate change: impacts of increasing drought may be mitigated by increasing CO2. J Ecol 98(1):197–208. https://doi.org/10.1111/j.1365-2745.2009.01602.x
Xu SY, Peddle DR, Coburn CA, Kienzle S (2008) Sensitivity of a carbon and productivity model to climatic, water, terrain, and biophysical parameters in a Rocky Mountain watershed. Can J Remote Sens 34(3):245–258. https://doi.org/10.5589/m08-029
Zhang Y, Keenan TF, Zhou S (2021) Exacerbated drought impacts on global ecosystems due to structural overshoot. Nat Ecol Evol 5(11):1490–1498. https://doi.org/10.1038/s41559-021-01551-8
Acknowledgements
We thank F. Schweingruber (Birmensdorf) and F. Meyer (Basel) for dendrochronological advice, N. Tsantas and P. Dimopoulos (Patras) for statistical and logistic support. A part of the data shown derives from the diploma work of S. Poulou (Patras). Our immense gratitude to Ch. Koerner (Basel) for inspiring this work, for his constructive comments and for hosting D.S. The cooperation of the Hellenic National Meteorological Service is gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding:This work was supported by the Research Committee of the University of Patras (“Karatheodori” research program code: Β.090), the COST Action ES1308–Climate Change Manipulation Experiments in Terrestrial Ecosystems–Networking and Outreach (ClimMani) and Exchange Grant (reference number 2850).
The online version is available at http://www.springerlink.com.
Corresponding editor: Tao Xu.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sarris, D., Christodoulakis, D. Topographic and climatic effects on Pinus halepensis s.l. growth at its drought tolerance margins under climatic change. J. For. Res. 35, 102 (2024). https://doi.org/10.1007/s11676-024-01755-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11676-024-01755-1