
Abstract
To identify if and how the age of in vitro potato plantlets at planting affects the later tuber formation, an extensive pot experiment was performed under controlled conditions. In vitro-derived plantlets of different ages (14, 28, 42 and 56 days old at planting) and cultivars varying in maturity type (Gloria, very early; Bintje, mid-early; Elkana, late) were grown after planting for 10 weeks at photo/thermoperiods of 16 h (standard conditions) or 12 h (short-day conditions). Plants raised from older in vitro plantlets produced higher total dry weights, harvest index and tuber dry and fresh weights than plants from younger in vitro plantlets when grown under standard conditions after planting. By contrast, total and tuber dry weights were not affected by plantlet age when grown under short-day conditions. The number of tubers per plant increased with increase in in vitro plantlet age in the later cultivars and in the oldest age classes of the very early cultivar when plants were grown under standard conditions. Short-day conditions resulted in fewer, but larger tubers than standard conditions. Results support the ideas that (1) plants from older in vitro plantlets and earlier maturing cultivars are more advanced in the tuber formation process; however, this may benefit tuber yield only when plants are grown under conditions that are not strongly inducing tuberization, and (2) higher tuber numbers can be achieved by more below-ground nodes—as in older in vitro plantlets—and/or by later or less strong tuber induction—as in less inducing conditions (16-h vs. 12-h), later cultivars or younger in vitro plantlets.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Many seed potato production schemes use in vitro-derived potato plantlets as starting material for the production of several generations of seed tubers. The plantlets are planted into soil or other substrates, or used for aeroponic production, and are then grown in glasshouses for the production of minitubers or transplants that are later planted in the field or screenhouses (e.g. Struik and Wiersema 1999; Tadesse et al. 2001a, b; Huarte 2005; Pruski 2007; van der Veeken and Lommen 2009; Lommen 2015; Dimante et al. 2022).
The state of the potato plantlets at the moment of planting—i.e. at the end of the last in vitro phase—affects their performance after planting (Kozai et al. 1988; Hagman 1990; Tadesse et al. 2000; Lommen 2023). This state is influenced by the conditions during in vitro growth and the duration of the last in vitro phase determining the age of the plantlets at the moment of planting. Planting older in vitro plantlets was shown to result in higher leaf area growth and higher absolute growth rates early after planting (Lommen 2023). The higher growth rates were attributed in part to higher leaf areas of the upper leaves of older in vitro plantlets kept above-ground at planting. Below-ground, a higher age of the in vitro plantlets planted results in more main stem nodes being present in the soil at planting, and more plants already producing stolons, with a higher number of stolons per plant, soon after planting (Lommen 2023). Plants from older in vitro plantlets also more likely initiate tubers soon after planting, especially in very early cultivars. A higher age of the in vitro plantlets was both reported to increase the number of tubers produced (Milinkovic et al. 2012) and to lead to a lower number of tubers (Hassanpanah and Khodadadi 2009). Overall, there is still limited understanding and limited published data on effects of age on tuber yield and number and little published information on the precise conditions under which the tuber production was studied.
The present research focuses on the effects of the age of the in vitro plantlets used for planting on tuber yield and number. Earlier research has shown that in vitro-derived plantlets or transplants from these plantlets are very sensitive to external conditions (cf. Lommen and Struik 2006), especially to conditions that are known to enhance tuber induction, like short photoperiods. The conditions under which the in vitro-produced plantlets are raised after planting may therefore affect the degree to which physiological or genetic differences between in vitro plantlets at the moment of planting are expressed thereafter.
The aims of this study were (1) to assess if and how differences in age of the in vitro plantlets planted affect final tuber production in cultivars differing in earliness and (2) to assess if and how conditions after planting influence the expression of the differences due to plantlet age and cultivar.
Materials and Methods
Experimental Design
A pot experiment was carried out in six walk-in growth chambers in which in vitro-derived potato plantlets were grown for 10 weeks. The in vitro-derived plantlets had four ages (14, 28, 42 and 56 days) at the moment of planting and were from three cultivars differing in maturity type (Gloria, very early; Bintje, mid-early; Elkana, late). They were grown after planting under two conditions [a 16-h photo/thermoperiod (‘standard conditions’) and a 12-h photo/thermoperiod (‘short days’)]. The experimental design had in the first stratum the conditions in six walk-in growth chambers, three for each of the two conditions. Twenty-four blocks were laid out across the growth chambers with first cultivar randomised within a condition and then age randomised within a cultivar. Additionally, 24 in vitro plantlets per cultivar and age class were used to characterise the in vitro plantlets at the moment of planting.
In Vitro Cultivation
In vitro plants were multiplied by single-node cuttings. Plantlets were cultured in glass tubes of 2.0 cm ∅ × 20 cm h with 9 ml medium containing M&S mineral salts, sucrose 25.0 g l−1, agar 8.0 g l−1, alar-64 (daminozide) 13.3 mg l−1, glycine 2.0 mg l−1, myo-inositol 100.0 mg l−1, nicotinic acid 0.5 mg l−1, pyridoxin HCl 0.5 mg l−1 and thiamine HCl 0.1 mg l−1. Plants were grown at 23 °C and 16-h photoperiod with light supplied by Philips TL-84 fluorescent tubes at an intensity of 4 W m−2. The medium and conditions were the same during stock multiplication and the last in vitro phase when plantlets were produced for planting. One explant, consisting of a nodal cutting including stem part, bud and leaf, was cultured per tube. Tubes were closed with a polycarbonate cap and sealed with household plastic film. Production of in vitro plantlets started several months in advance and was scheduled in such a way that in the experiment, the in vitro plants of all different ages could be planted to the in vivo conditions on the same day, whereas all explants used for producing these plantlets were nodal cuttings derived from 5-week-old plantlets. The latter ensured that all plantlets of different age were derived from comparable source (ex)plants.
Cultivation Practices Ex Vitro
In vitro plantlets with roots were planted into black plastic pots of 18 × 18 × 18 cm filled with potting soil, one plantlet per pot, leaving four visible leaves per plant above the soil level for all cultivars and age classes and a variable number of nodes/leaves (increasing with increase in plantlet age) below-ground.
Plants were grown in walk-in growth chambers at day/night temperatures of 20/12 °C and RH 70%; radiation was supplied by SON-T and HPI-T lamps (Philips) in a 1:1 ratio at 100–170 W m−2 depending on the block. The photo/thermoperiod was 16 or 12 h according to treatment. Plants were fertilised 16, 30, 44 and 58 days after planting (DAP) with 100 ml of a concentrated nutrient solution [NH4NO3 6.4 g l−1, Ca(NO3)2 9.442 g l−1, KNO3 8.09 g l−1, KH2PO4 5.4428 g l−1, MgSO4.7H2O 9.853 g l−1, FeEDTA 0.04375 g l−1, MnSO4.1H2O 0.08 g l−1, H3BO3 0.12 g l−1, ZnSO4.7H2O 0.02 g l−1, Na2MoO4.2H2O 0.004 g l−1, CuSO4.5H2O 0.004 g l−1, pH 6.0] that was diluted 5 times before application.
It was ensured that from 34 DAP onwards the minimum distance between pots within a block was 10 cm and between blocks 15 cm, which corresponds to a minimum space of 28 × 43 cm per plant.
Plants were hilled 35 DAP by adding 2.5 cm (± 800 cm3) additional potting soil per pot.
Observations
Plants were analysed at planting and after 10 weeks ex vitro growth (at ‘harvest’). At planting, dry weights of shoot and root of the in vitro plants were determined and the length and leaf/node number of the stem. At harvest after 10 weeks growth, plants were separated in three fractions: the ‘tuber’ fraction (including tubers only), the ‘above-ground shoot’ (stems and leaves), and the ‘below-ground stems and stolons’; roots were not included in any fraction at this harvest moment. The sum of ‘above-ground shoot’ and ‘below-ground stems and stolons’ was regarded to be the ‘non-tuber’ fraction. The fresh tuber weight and total tuber number per plant were determined. Dry weights of the fractions were assessed after drying for at least 16 h at 70 °C. The total dry weight was the sum of the tuber and the non-tuber fractions. Harvest index was the dry weight of the tuber fraction divided by the total dry weight.
Statistical Analysis
Genstat 19.1 edition (VSN International Ltd. 2018) was used for analysis of variance. Differences between means within significant main effects or interactions were analysed by Fisher’s protected LSD test at α = 0.05.
Results
Initial Characteristics of the In Vitro Plantlets at Planting
At the moment of planting, older in vitro plantlets were more advanced in growth and development than younger in vitro plantlets. Stem length, node/leaf number on the main stem, and shoot dry weight increased across the full range of age classes (14–56 days) in all cultivars, whereas root dry weight seemed to level off between 42 and 56 days in cvs Gloria and Elkana but continued to increase significantly with increase in age in cv. Bintje (Table 1). Because in vitro-derived plantlets of all age classes were planted with exactly four visible leaves above the soil, the higher node/leaf numbers in the older plantlets also resulted in higher node/leaf numbers buried below-ground (1.7–2.5 nodes for 14-day-old plantlets up to 7.6–9.0 nodes for 56-day-old plantlets, depending on the cultivar).
Of the three cultivars, cv. Gloria grew fastest in vitro and the oldest plantlets of cv. Gloria had higher shoot and root weights at planting than those of cv. Elkana, with cv. Bintje taking an intermediate position (Table 1). Stems were longest in cv. Bintje and shortest in cv. Elkana. Also for number of stem nodes/leaves, there was an interaction between cultivar and plantlet age. In the youngest in vitro plantlets, cv. Gloria had the highest number of stem nodes/leaves, in the older in vitro plants cv. Bintje had the highest number; cv. Elkana was the cultivar with the lowest number of nodes/leaves across age classes.
Expression of Effects of Age of In Vitro Plantlet and Cultivar after 10 Weeks Growth Under Different Conditions
The growth conditions after planting had a strong effect on the overall performance of the plants and also determined if and how differences in age of in vitro plants and differences between cultivars were expressed, as shown by significant two-way interactions between age and conditions (age × cond) and cultivar and conditions (cv × cond) for all variates studied and by significant three-way interactions between age, conditions and cultivar (age × cv × cond) for number of tubers and weight per tuber (Table 2). Also, age × cultivar interactions were significant for some characteristics (Table 2).
The significant two- and three-way interactions will further guide the presentation in the results section on the effects of conditions on the expression of differences between age classes and between cultivars. The description of the effects of age and cultivar was based on three-way interactions when significant, or on the significant two-way interactions including conditions when the three-way interaction was not significant. To facilitate comparisons, individual means are presented in Figs. 1 and 2, but with the statistical information being based on the relevant interaction(s) (three- or two-way) between conditions and other factors (Table 2) as explained above. The full set of means is provided in the Appendix (Table 3).

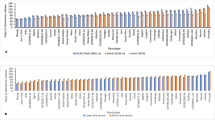
Effects of the age of the in vitro plant used for planting and cultivar on dry matter production and allocation in plants grown under standard conditions (left panels A, C, E, G) and short days (right panels B, D, F, H), 10 weeks after planting. A, B Total dry weight; C, D harvest index (HI); E, F tuber dry weight; G, H above-ground shoot dry weight. To facilitate comparisons, all individual means are presented. In bold, the relevant interactions for mean separation. Two-way interactions between age and conditions (age × cond) and cultivar and conditions (cond × cv) were significant for all variates. Different letters reflect significant differences between age class means (black letters) or cultivar means (coloured letters) within a condition. Fisher’s protected LSD test (α = 0.05) was used to separate the relevant means. ***Significant at P < 0.001, **significant at 0.001 ≤ P < 0.01, *significant at 0.01 ≤ P < 0.05, ns not significant, P ≥ 0.05. For full statistical information, see Table 2

Effects of the age of the in vitro plant used for planting and culivar on tuber-related characteristics in plants grown under standard conditions (left panels A, C, E, G) and short days (right panels B, D, F, H), 10 weeks after planting. A, B Tuber fresh weight; C, D tuber dry matter fraction; E, F tuber number; G, H fresh weight per tuber. All individual means are presented. In bold, the relevant interactions for mean separation. Two-way interaction between age and conditions (age × cond) and cultivar and conditions (cond × cv) were significant for the upper two variates and three-way interaction for the lower two. Different letters reflect significant differences between age class means (black letters) or cultivar means (coloured letters) within a condition for the upper two variates (A, B, C, D) and between age classes within a cultivar per condition for the lower two variates (E, F, G, H). In the lower diagrams (E, F, G, H), significant differences between cultivars are indicated per age class and condition by tick marks. Fisher’s protected LSD test (α = 0.05) was used to separate the relevant means. ***Significant at P < 0.001, **significant at 0.001 ≤ P < 0.01, *significant at 0.01 ≤ P < 0.05, ns not significant, P ≥ 0.05. For full statistical information, see Table 2
Effects of Age of In Vitro Plantlets
Dry Matter Production and Allocation
When in vitro plantlets of different age were grown after planting for 10 weeks under standard conditions, the total dry weight of the plants was higher when older in vitro plantlets, up to an age of at least 42 days, had been used for planting (Fig. 1A). Also harvest index (Fig. 1C) and tuber dry weight (Fig. 1E) were higher when older in vitro plantlets had been planted under standard conditions, whereas the dry weight of the above-ground shoot at harvest was lower in plants from older in vitro plantlets than in plants from the youngest in vitro plants under standard conditions (Fig. 1G).
By contrast, under short-day conditions, no significant effects of in vitro plantlet age were observed 10 weeks after planting for total dry weight, tuber dry weight and above-ground shoot dry weight (Fig. 1B, F, H). Harvest indexes were in general high under the short-day conditions and the increase in harvest index with increase in age class of the in vitro plantlets planted was small and hardly significant (Fig. 1D).
Across conditions, differences in harvest index between age classes were smaller in the very early cultivar Gloria than in the other cultivars, as indicated by significant age × cv interaction (Table 2; underlying averages are shown in the Appendix, Table 3).
Tuber characteristics
Under standard conditions, tuber fresh weight per plant produced at 10 weeks after planting increased slightly with increase in age of the in vitro plantlets planted (Fig. 2A), whereas under short-day conditions a small decrease in tuber fresh weight per plant with increase in in vitro plantlet age was observed (Fig. 2B). Tuber dry matter concentration was higher in tubers from older in vitro plantlets under standard conditions (Fig. 2C) and—to a smaller extent—also under short days (Fig. 2D). Under standard conditions, the number of tubers per plant increased with increase in age of the in vitro plantlet planted over the full range of ages (14–56 days) in the late cv. Elkana, up to 42 days in the mid-early cv. Bintje and from 28- to 56-day-old plantlets in the very early cv. Gloria, whereas in cv. Gloria, plants from the youngest (14-day-old) in vitro plantlets had a relative high number of tubers, comparable to that of the oldest (56-day-old) in vitro plantlets (Fig. 2E). Under short days, the number of tubers per plant was usually much lower than under standard conditions and varied much less with age of the planted in vitro plantlets; differences between age classes were not significant in cvs Gloria and Elkana, whereas in cv. Bintje a small increase in tuber number with age was observed in the three older age classes, while the youngest plants had an intermediate number of tubers, not differing significantly from any of the other age classes (Fig. 2F). The average weight per tuber seemed in general to be lower when older in vitro plantlets had been used as planting material, although the trend was not always statistically significant, especially not under standard conditions (Fig. 2G); there was one exception: the high number of tubers produced in the plants from the youngest (14-day-old) in vitro plants of cv. Gloria under standard conditions resulted in a lower average weight per tuber than in plants of this cultivar grown from 28- and 42-day-old in vitro plantlets under standard conditions.
Effects of Cultivar
Dry matter production and allocation
Under standard conditions, the late cultivar Elkana had lower total dry weights than the mid-early cultivar Bintje and the very early cultivar Gloria (Fig. 1A), whereas under short-day conditions, cv. Elkana had higher total dry weights than the other cultivars (Fig. 1B). Under standard conditions, the cultivars followed a logical cultivar ranking for harvest index, with the late cultivar Elkana having the lowest harvest index, the mid-early cultivar Bintje having an intermediate harvest index and the very early cultivar Gloria having the highest harvest index (Fig. 1C). Under short-day conditions, all harvest indices were high, but with cv. Bintje having on average a higher harvest index than the other two cultivars, which did not differ significantly from each other (Fig. 1D). Cultivar rankings for tuber dry weight were comparable to those for harvest index within a condition, but with tuber dry weights on average being lower under short-day conditions than under standard conditions. Cv. Elkana significantly had the highest above-ground shoot dry weight of the three cultivars under both conditions, but the differences between cultivars were pronounced and followed a logical cultivar ranking under standard conditions (Fig. 1G) and were much smaller and not always significant under short-day conditions (Fig. 1H).
Tuber Characteristics
Under standard conditions, cv. Gloria had the highest tuber fresh weights, cv. Bintje had intermediate levels and cv. Elkana had the lowest tuber fresh weights; the cultivar ranking was the same under short-day conditions, but the differences between cultivars were much smaller, and only significant for cv. Gloria versus the other cultivars. The cultivar ranking differed between conditions for tuber dry matter concentration, tuber number and weight per tuber. Cultivar Elkana had a higher tuber dry matter concentration than the other cultivars under short days but took an intermediate position under standard conditions. Cultivar Gloria produced—for most age classes—fewer but heavier tubers than the other cultivars under standard conditions, and more and smaller tubers than the other cultivars under short days.
Discussion
Results showed that differences in tuber production arising from differences in in vitro plantlet age were effectively expressed under standard conditions after planting, but largely suppressed or overruled under short-day conditions. Moreover, conditions after planting also strongly affected the expression of differences due to cultivar earliness.
Below, first the effects of in vitro plantlet age on dry matter production and allocation under standard conditions after planting will be discussed, followed by the changes when grown under short photo/thermoperiods. Thereafter, the processes governing the number of tubers produced will be discussed and finally the behaviour of the cultivars.
Dry Matter Production and Allocation Under Standard Conditions: Tuber Formation is Enhanced in Plants from Older In Vitro Plantlets and from Earlier Cultivars
Under standard conditions (16 h photo/thermoperiod of 20/12 °C) after planting, planting older in vitro-derived plantlets resulted 10 weeks later in higher total and tuber dry weights in all cultivars (Fig. 1A, E).
The higher total and tuber dry weights are in line with the small differences in weight of the in vitro plantlets at planting (cf. Table 1) and the larger total dry matter production in the first 2 weeks after planting noted earlier when planting older in vitro plantlets up to c. 42–56 days old (Lommen 2023). All this will have given the plants from older plantlets a head start in growth and radiation interception.
Additionally, the higher tuber dry weight will have resulted from a proportionally higher dry matter allocation to the tubers in plants from the older plantlets suggesting these plants were also more advanced in development. At 10 weeks after planting, under standard conditions, plants from the youngest in vitro plantlets still had the highest above-ground shoot dry weight (Fig. 1G) and the lowest harvest index (Fig. 1C). This likely was associated with delayed and/or reduced allocation of dry matter to the tubers. Time of tuber initiation will have been later in plants from the younger in vitro plantlets, in line with the stolon formation being delayed in plants from the youngest in vitro plantlets—as shown by lower fractions of plants having produced stolons early after planting, and the lack of tuber initiation in the first weeks after planting (Lommen 2023).
Additionally, total dry matter production may also have been higher in plants from the older in vitro plantlets because photosynthesis and dry matter production can be more efficient when tubers are produced and can act as strong carbohydrate sinks (cf. Nösberger and Humphries 1965; Moll 1986).
Dry Matter Production and Allocation Under Short Days: Differences Between Plantlet Ages and Cultivars are Suppressed
Under short-day conditions (12-h photo/thermoperiod of 20/12 °C), plants were exposed to a shorter photoperiod than under standard conditions (12 vs. 16 h) and to a slightly lower average temperature (16 vs. 17.33 °C) due to shorter day period. Because of the shorter day period, lower total amounts of radiation were available for the plants from 12 h irradiance per day than from 16 h. Under these short-day conditions, total dry matter yields were generally lower than under standard conditions, harvest indices generally higher and no significant effects of in vitro plantlet age on total and tuber dry weights were observed (Fig. 1B, F). The overall lower yields under short-day conditions will have been primarily lower due to the lower total amounts of radiation available for interception and photosynthesis per day. Under these short-day conditions, plants from all age classes and cultivars will have become strongly induced to form tubers relatively soon after planting because of the short photoperiod. In vitro-derived plants were shown to differ in degree of tuber induction when the conditions after planting differed more than c. 10 days (cf. Lommen and Struik 2006); moreover, early tuber initiation and allocation of dry matter to tubers is also in line with the generally high harvest indices (c. 0.81) found under short-day conditions and with the above-ground dry weights under short-day conditions being about half of those under standard conditions.
Under these strongly inducing conditions, even a slight delay in tuber initiation to a moment at which the canopy was larger, might have had a positive effect on yield. The age of the in vitro plantlets might not have had significant effects on total and tuber dry matter yield, because a possible slower initial growth after planting of plants derived from the youngest in vitro plantlets might have been compensated for by a slightly later tuber initiation—allowing dry matter to be invested in extra leaf growth. This is in line with the harvest index being very high for all plantlet ages under short-day conditions, but still increasing to a very small extent with an increase in plantlet age (Fig. 1D).
The positive effect of a later moment of tuberization or less strong allocation of dry matter to tubers under the short-day conditions is shown most clearly by the response of the different cultivars, with the latest cultivar Elkana still having more above-ground dry matter than the earlier cultivars (Fig. 1H) and higher total and tuber dry matter yields (Fig. 1B, F).
Tuber Number Seems to be Affected Along at Least Two Mechanisms: Number of Below-ground Nodes and Development at the Moment of Tuber Initiation
Significant three-way interaction for number of tubers per plant indicated that the effects of the in vitro plantlet age at planting on the number of tubers produced after 10 weeks depended on the environmental conditions, the cultivar and their interaction. The most prominent effect on tuber number was caused by the conditions, with short-day conditions resulting in few tubers per plant and little variation over cultivars and ages compared to standard conditions.
Concerning the effects of age two trends were visible. Under standard conditions, only plants from the youngest (14-day-old) in vitro plantlets in cv. Gloria had a higher number of tubers than those of intermediate age, whereas the number of tubers increased with increase in age over the full range of ages (14–56 days) in the late cv. Elkana, up to 42 days in the mid-early cv. Bintje and from 28 to 56 days old age in the very early cv. Gloria (Fig. 2E). Under short-day conditions, the number of tubers varied little with plantlet age and usually not significantly; yet also here a kind of downward trend was visible in plants from younger plantlets followed by an upward trend in plants from older plantlets. Consequently, at least two different—and partly counteracting—mechanisms will have determined the observed differences in number of tubers between ages, cultivars and conditions.
First, there seemed to be a negative effect of age on tuber number, which was expressed under conditions that were strongly inducing, and from which plants from the youngest plantlets could ‘escape’ to a certain degree. This ‘escape’ was visible in the youngest plants of the earliest cultivar Gloria under standard conditions and was weakly visible (though not significant) in all cultivars under short-day conditions. The effect may be caused by a mere delay of the moment of tuber initiation resulting in a slightly larger canopy at tuber initiation and/or a more extensive development of stolons and potential sites for tuber production. In in vitro-derived plants, the number of tubers initiated is associated with the size of the canopy at the moment of initiation (van der Veeken and Lommen 2009). Moreover, shallow stem buds may be delayed in stolon formation because of the light conditions (cf. Kumar and Wareing 1972) or being too close to active leaves for rapid production of (sessile) tubers (cf. Khan and Ewing 1983). In the present experiment, where plants were grown at c. 8.3 plants per m2, the number of tubers (Fig. 2E, F) by far exceeded the number of nodes planted below soil in all age classes, but especially in the youngest age class which on average had only 2 shallow nodes below soil at the moment of planting (cf. Table 1). This means that in all treatments more than one tuber (on top of one stolon) per node was produced and that more extensive formation of possible sites of tuber production was necessary to accommodate tuber initiation. Tuber formation being delayed in plants from the youngest (14 days old) in vitro plantlets [as also indicated by later stolon formation (Lommen 2023) and in line with a lower harvest index (Fig. 1C)] may have resulted in more tubers in plants raised from the 14-day-old plantlets in cv. Gloria than in plants raised from 28- or 42-day-old plantlets. This view is also confirmed by a much higher dry weight of the below-ground stem and stolon parts in plants from these young plantlets (Appendix, Table 3).
Second, there was—especially in the older age classes—a positive effect of higher age resulting in more tubers; this might be related to more below-ground nodes, more stolons from these nodes and more potential tuber positions. In crops from conventional seed tubers, tuber numbers are well known to be associated with stolon numbers within a cultivar (e.g. Haverkort et al. 1991; Struik and Wiersema 1999). In the present experiment, older in vitro-derived plantlets had more stem nodes (Table 1) and more stem nodes buried below soil. In addition, previous research has shown that at 2 weeks after planting comparable plantlets in standard conditions, the plants from older plantlets had more stolons per plant and more potential tuber positions on the stolons (Lommen 2023). This will give the plants from older in vitro plantlets a head start in developing below-ground sites for potential tuber production, but the plants will only be able to exploit this potential under conditions that are not too strongly inducing.
A positive effect of in vitro plantlet age on tuber number was also found by Milinkovic et al. (2012), who compared tuber numbers in plants from 4-, 10- and 12-weeks-old in vitro plantlets after growing them 14 weeks under greenhouse conditions (14-h thermoperiod of 25/15 °C, natural day light; planted 20 October in Toolangi, Victoria, Australia). They found in both cultivars studied fewer tubers in plants from 4- than 10-week-old plantlets, although at a much higher planting density than in the present experiment; this higher planting density may in part may explain their lower tuber numbers (2.50–4.94) per plant compared with the present experimentation (Fig. 2E, F). By contrast, Hassanpanah and Khodadadi (2009) harvested 2 months after planting more tubers when planting 20- and 30-day-old in vitro-derived plantlets than when planting 40- and 50-day-old plantlets. The exact greenhouse conditions or time of year at the latter location (Ardabil, Iran) were not published.
Finally, it cannot be excluded that also a larger/longer green canopy (LAD, leaf area duration) may have increased the number of tubers. It may keep tubers from being resorbed or allow continuation of tuber initiation during a longer timeframe. By contrast to plants from seed tubers, in vitro-derived plants can initiate tubers during a long period (Lommen 2015), longer than what is common in seed tuber grown crops.
All this means that a higher number of tubers could be expected when the moment of tuber initiation is delayed and/or dry matter allocation to tubers is less strong, the canopy is more developed at the moment of tuber initiation and enough sites for tuber production are available. This will also explain why the number of tubers (Fig. 2E, F was much lower under short days than under standard conditions (tuber initiation will have occurred earlier under the strongly inducing short-day conditions).
Effects on the weight per tuber (Fig. 2G, H) were the resultant of the effects on fresh tuber yield and on number of tubers per plant.
Susceptibility of In Vitro Plantlets to External Conditions Affects Cultivar Rankings
Because of the idea that the absence of a mother tuber makes the in vitro-derived plants more depending on and susceptible to external conditions for regulation of development (Lommen 1999), especially with regard to tuber induction, the plants in the present experiment were grown under two photo/thermoperiods and three cultivars differing in earliness were included in the experiment (in order to study a possible effect of age).
Effects of cultivars on dry matter production and dry matter allocation under standard conditions were fully as expected based on their maturity type, with the latest cultivar, Elkana, having the lowest harvest index after 10 weeks growth, still the highest above-ground dry weights and the lowest tuber and total dry weights. Differences between cultivars in dry matter production and allocation were reduced under inducing conditions.
Under both conditions, the cultivar behaviour/response in tuber number and weight per tuber of the in vitro-derived plants from the three cultivars differed considerably from what is commonly found for these cultivars in crops from seed tubers. Of the cultivars used, cv. Elkana is known for producing the lowest number but the largest tubers under common cultivation conditions and cv. Bintje is known for producing the highest number of tubers. In the present investigation, tuber numbers never were higher for Bintje than for Elkana (Fig. 5E, F). This confirms earlier findings that there is little correlation between plants from in vitro culture and from seed tubers, and that the cultivar ranking depends on the type of propagule used for planting (Schenk 1989; Lommen 1999).
In general, the cultivar performance highly depended on the type of conditions—as shown by significant interactions between cultivars and conditions for all variates (Table 2, Figs. 1 and 2). These interactions did not just result from a different magnitude of the differences between cultivars under the two conditions (Fig. 1C, D, G, H; Fig. 2A, B) but also from a different ranking of cultivars for important variates like total and tuber dry matter production and tuber number and size under different conditions (Fig. 1A, B, E, F; Fig. 2C–H).
Conclusions
The results support the idea that plants grown from older in vitro plantlets and earlier maturing cultivars are more advanced in the tuber formation process; however, this may benefit tuber yield only when plants are grown after planting under conditions that are not strongly inducing tuberization or induce a relative high allocation of dry matter to tubers; under inducing conditions early initiation and bulking of tubers will be stimulated in all age classes and cultivars, and using older in vitro plantlets or early cultivars may not affect or even compromise tuber yield due to insufficient allocation of dry matter to canopy production. Under these conditions, using young in vitro plantlets may alleviate the negative effect.
Higher numbers of tubers per plant can be achieved (a) by more below-ground nodes—as in older in vitro plantlets—and/or (b) by later or less strong tuber induction or a longer tuber initiation period—as in less inducing conditions (16-h vs. 12-h photo/thermoperiod), later cultivars or younger in vitro plantlets. A higher in vitro plantlet age thus may affect the number of tubers produced positively through increasing below-ground node number but negatively through advancing tuber initiation and bulking.
In general, potential differences due to in vitro plantlet age and cultivar were only effectively expressed under standard conditions and strongly suppressed under short-day conditions.
Data Availability
Data is available upon request.
References
Dimante I, Gaile Z, Cerina S, Proskina L (2022) High microplant densities in greenhouses – Are they always efficient in small potato minituber production systems? Am J Potato Res 99:243–257. https://doi.org/10.1007/s12230-022-09876-1
Hagman J (1990) Micropropagation of potatoes. Comparison of different methods. Crop Production Science 9:31–80
Hassanpanah D, Khodadadi M (2009) Study the plantlet age effect and planting beds on Agria potato mini-tuber production under in vivo condition. J Biol Sci 9(3):243–248
Haverkort AJ, van de Waart M, Bodlaender KBA (1991) Interrelationships of the number of initial sprouts, stems, stolons and tubers per potato plant. Potato Res 33:89–96
Huarte M (2005) Seed potato systems in Latin America. In: Haverkort AJ, Struik PC (eds) Potato in progress: science meets practice. Wageningen, Wageningen Academic Publishers, pp 86–92
Khan BA, Ewing EE (1983) Factors controlling the basipetal pattern of tuberization in induced potato (Solanum tuberosum L.) cuttings. Ann Bot 52:861–871
Kozai T, Koyama Y, Watanabe I (1988) Multiplication of potato plantlets in vitro with sugar free medium under high photosynthetic photon flux. Acta Hort 230:121–127
Kumar D, Wareing PF (1972) Factors controlling stolon development in the potato plant. New Phytol 71:639–648
Lommen WJM (1999) Causes for low tuber yields of transplants from in vitro potato plantlets of early cultivars after field planting. J Agric Sci 133:275–284
Lommen WJM (2015) How age of transplants from in vitro derived potato plantlets affects crop growth and seed tuber yield after field transplanting. Potato Res 58:343–360
Lommen WJM (2023) Effects of age of in vitro derived potato plantlets on early above- and below-ground development after planting of different cultivars. Potato Res. https://doi.org/10.1007/s11540-023-09621-z
Lommen WJM, Struik PC (2006) Tuber induction and initiation during production and early field growth of transplants from in vitro-derived potato plants. Ann Appl Biol 149:281–290
Moll A (1986) Effect of tuber removal on the rate of 14CO2 fixation in potato leaf discs. Photosynthetica 20:14–19
Milinkovic M, Horstra CB, Rodoni BC, Nicolas ME (2012) Effects of age and pretreatment of tissue-cultured potato plants on subsequent minituber production. Potato Res 55:15–25
Nösberger J, Humphies EC (1965) The influence of removing tubers on dry-matter production and net assimilation rate of potato plants. Ann Bot 29:579–588
Pruski K (2007) The canon of potato science: 22. In vitro multiplication through nodal cuttings. Potato Res 50:293–296
Schenk G (1989) Sortenechtheidsprüfung von In-vitro-Herkünften der Kartoffel. Archiv Für Züchtungsforschung, Berlin 19:317–326
Struik PC, Wiersema SG (1999) Seed potato technology. Wageningen, Wageningen Pers
Tadesse M, Lommen WJM, Struik PC (2000) Effects of in vitro treatments on leaf area growth of potato transplants during transplant production. Plant Cell, Tissue Organ Cult 61:59–67
Tadesse M, Lommen WJM, Struik PC (2001a) Effect of nitrogen pre-treatment of transplants from in vitro produced potato plantlets on transplant growth and yield in the field. Neth J Agric Sci 49:67–79
Tadesse M, Lommen WJM, Struik PC (2001b) Effect of temperature pre-treatment of transplants from in vitro produced potato plantlets on transplant growth and yield in the field. Potato Res 44:173–185
van der Veeken AJH, Lommen WJM (2009) How planting density affects number and yield of potato minitubers in a commercial glasshouse production system. Potato Res 52:105–119
VSN International Ltd. (2018). GenStat for Windows, edition 19.1. Hemel Hempstead, VSN International Ltd.
Acknowledgements
I gratefully acknowledge the role of Ms PM van de Putte, who carried out the experiment as part of her MSc study. I also thank the phytotron team of Wageningen University involved in this work for taking care of the plants, and Wageningen University for providing the six walk-in growth chambers simultaneously for the described experiment.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The author declares no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix
Appendix
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lommen, W.J.M. Environmental Conditions After Planting Affect the Expression of Differences in Tuber Formation of In Vitro-Derived Potato Plantlets from Different Ages and Cultivars. Potato Res. 67, 117–137 (2024). https://doi.org/10.1007/s11540-023-09622-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11540-023-09622-y