
Abstract
Introduction
Analysis of time-resolved postprandial metabolomics data can improve our understanding of the human metabolism by revealing similarities and differences in postprandial responses of individuals. Traditional data analysis methods often rely on data summaries or univariate approaches focusing on one metabolite at a time.
Objectives
Our goal is to provide a comprehensive picture in terms of the changes in the human metabolism in response to a meal challenge test, by revealing static and dynamic markers of phenotypes, i.e., subject stratifications, related clusters of metabolites, and their temporal profiles.
Methods
We analyze Nuclear Magnetic Resonance (NMR) spectroscopy measurements of plasma samples collected during a meal challenge test from 299 individuals from the COPSAC2000 cohort using a Nightingale NMR panel at the fasting and postprandial states (15, 30, 60, 90, 120, 150, 240 min). We investigate the postprandial dynamics of the metabolism as reflected in the dynamic behaviour of the measured metabolites. The data is arranged as a three-way array: subjects by metabolites by time. We analyze the fasting state data to reveal static patterns of subject group differences using principal component analysis (PCA), and fasting state-corrected postprandial data using the CANDECOMP/PARAFAC (CP) tensor factorization to reveal dynamic markers of group differences.
Results
Our analysis reveals dynamic markers consisting of certain metabolite groups and their temporal profiles showing differences among males according to their body mass index (BMI) in response to the meal challenge. We also show that certain lipoproteins relate to the group difference differently in the fasting vs. dynamic state. Furthermore, while similar dynamic patterns are observed in males and females, the BMI-related group difference is observed only in males in the dynamic state.
Conclusion
The CP model is an effective approach to analyze time-resolved postprandial metabolomics data, and provides a compact but a comprehensive summary of the postprandial data revealing replicable and interpretable dynamic markers crucial to advance our understanding of changes in the metabolism in response to a meal challenge.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
1 Introduction
Food ingestion triggers many parts of the human metabolism. While the fasting state may reveal certain metabolic differences among individuals, recently challenge tests have been used to study also the postprandial metabolism of individuals (Bermingham et al., 2023). Analyzing postprandial data may help us understand differences, stratify individuals in terms of their metabolic responses (Bermingham et al., 2023; Harte et al., 2012; Wopereis et al., 2017) and advance personalized nutrition (Berry et al., 2020; Zeevi et al., 2015). For example, postprandial hyperlipidemia, the abnormally increased levels of triglyceride-rich lipoproteins after food intake, is a risk factor for cardiovascular diseases (Botham & Wheeler-Jones, 2013; O’Keefe & Bell, 2007; Poppitt, 2005). Subjects with different body mass index (BMI) values have shown different postprandial responses in terms of amino acids (Bastarrachea et al., 2018; Bondia-Pons et al., 2014) and lipid metabolites (Rämö et al., 2017). Sex differences in insulin sensitivity and metabolome have been observed in the postprandial response (Kumar et al., 2020) (See Lépine et al. (2022) for a recent review on challenge tests and metabolomic responses.) To take it even further, personalized dietary recommendations based on integrating postprandial data with, e.g., gut microbiome characteristics, may lower postprandial glucose levels (Zeevi et al., 2015).
Mining a rich information source such as the postprandial metabolomics data requires advanced data analysis approaches to extract insights. However, there is currently a gap between the complexity of the data and the analysis methods used to analyze such data. The state-of-the-art methods to analyze challenge test data can be considered under supervised and unsupervised methods. In many cases, there is a study design underlying a challenge test with different groups, e.g., normal vs. hyper-triglycerides (Wojczynski et al., 2011), or healthy vs. diseased or pathological conditions (Kumar et al., 2020). The data in such studies is labelled, and commonly analyzed using supervised methods often using univariate methods in which metabolites are analyzed separately, e.g., t-tests (LaBarre et al., 2021; Wojczynski et al., 2011) or analysis of variance (ANOVA) models (Berry et al., 2020; Kumar et al., 2020; Rizi et al., 2019; Smilde et al., 2010; Wojczynski et al., 2011) studying group differences. More advanced methods consider explicitly the temporal aspect of the data and/or unbalanced designs, e.g., by using linear mixed models (Müllner et al., 2021), extensions of the analysis of variance-simultaneous component analysis (ASCA) (Erdos et al., 2023). A shortcut for analyzing the temporal behavior is to extract features from the dynamic profiles such as the area under the curve (AUC) (Lairon et al., 2007; Pellis et al., 2012; Rämö et al., 2017). Although these methods are powerful, they rely on labelled data, and may fail to reveal unknown subject stratifications, or they use derived features that do not reveal the full dynamic profile of the data. Univariate methods do not take into account the interplay between metabolites, and may miss the biomarkers by focusing on one metabolite at a time. If there is no a priori group information, e.g., cohort data, which is also the topic of this paper, then such data is unlabelled, and analyzed using unsupervised methods, commonly relying on multivariate methods such as principal component analysis (PCA) or clustering. PCA has been used by arranging the data as subjects-time points by metabolites (Zivkovic et al., 2009) or one time point at a time (Saito et al., 2020). Clustering has been used to cluster metabolites (e.g., averaged across subjects) to create groups of metabolites showing similar time courses (Pellis et al., 2012; Wopereis et al., 2017). These multivariate methods work by considering only summaries of the data or do not utilize the fact that challenge test data has a temporal structure.
Postprandial metabolomics data collected at multiple time points from multiple subjects is inherently three-way, and can be arranged as a subjects by metabolites by time points array (see Fig. 1). Multiway data analysis methods (also known as tensor factorizations) (Acar & Yener, 2008; Kolda & Bader, 2009; Smilde et al., 2004) are effective tools for extracting the underlying patterns from such multiway data (also referred to as higher-order tensors), and have been successfully used in data mining, e.g., (Acar et al., 2007; Becker et al., 2023; Williams et al., 2018; Yin et al., 2020).
With the goal of stratifying subjects with no prior labels and understanding differences among subjects based on their metabolic responses to a meal, in this paper, we analyze measurements of plasma samples collected during a meal challenge test from the COPSAC2000 cohort (Copenhagen Prospective Studies on Asthma in Childhood) (Bisgaard, 2004). We arrange the metabolomics measurements at several time points from a group of participants as a subjects by metabolites by time points tensor, and use the CANDECOMP/PARAFAC(CP) tensor factorization model (Carroll & Chang, 1970; Harshman, 1970) to reveal the underlying patterns in the data. The CP model summarizes the data by extracting the main patterns of variation in the subjects, metabolites and time modes simultaneously. Extracted patterns are unique under mild conditions (Kolda & Bader, 2009; Kruskal, 1977) and this facilitates interpretation. While CP-based tensor methods have been previously used in longitudinal data analysis, e.g., analysis of gut microbiome (Martino et al., 2021), urine metabolomics (Gardlo et al., 2016), and simulated dynamic metabolomics data (Li et al., 2022), its application to postprandial metabolomics data has been limited. We recently simulated postprandial metabolomics data using a human whole-body metabolic model (Li et al., 2024), and demonstrated that analysis of the T0-corrected data (i.e., the fasting state-corrected data, where the postprandial data is corrected by subtracting the fasting state data) using a CP model together with the analysis of the fasting state data provides a comprehensive picture of the underlying metabolic mechanisms.

An R-component CP model of a third-order tensor with modes: subjects, metabolites, and time points
The aim of this study is to investigate postprandial dynamics of human metabolism in comparison with the fasting state through the analysis of the time-resolved postprandial metabolomics data collected from the COPSAC2000 cohort. We analyze the T0-corrected data using a CP model and the fasting state data using PCA. We demonstrate that the CP model reveals biologically meaningful and replicable patterns (i.e., replicable across subsets of subjects) providing a compact but a comprehensive summary of the postprandial data.
2 Data description
The COPSAC2000 cohort contains 411 healthy subjects with mothers with a history of asthmaFootnote 1. 299 subjects, aged 18 years, attended the food challenge test using a standardized mixed meal (Stroeve et al., 2015) containing 60 g palm olein, 75 g glucose, and 20 g dairy protein in a total volume of 400 ml water-based drink. The size of the drink was made proportional to the daily recommended caloric intake (function of sex, age and height). Blood samples were collected from the participants at eight time points after an overnight fasting, i.e., at the fasting state, and 0.25, 0.5, 1, 1.5, 2, 2.5, and 4 h after the meal intake. The samples were put on ice, and within 4 h split into EDTA plasma before being stored at \(-80 ^{\circ }\)C.
Plasma samples were measured using NMR (nuclear magnetic resonance) spectroscopy through Nightingale Blood Biomarker Analysis, which provides 250 features.Footnote 2 These features consist of lipoproteins, apolipoproteins, aminoacids, fatty acids, glycolysis-related metabolites, ketone bodies and an inflammation marker. Lipoproteins consist of four main lipoprotein classes: HDL (high density lipoprotein), IDL (intermediate density lipoprotein), LDL (low density lipoprotein), and VLDL (very low density lipoprotein), and subclasses divided according to particle sizes: XXL (extremely large), XL (very large), L (large), M (medium), S (small) and XS (very small). Cholesterol (C), cholesterol ester (CE), free cholesterol (FC), triglyceride (TG), phospholipid (PL) levels of these classes and subclasses are among the list of metabolites. In addition, we included measurements of insulin and C-peptide. The total number of features we included in this study is 161, and these features are given in Table S1.1 in Supplementary Material S1.
Seven subjects are removed before the analysis since two subjects have a large amount of missing data, four subjects have extremely high levels of acetate, and one subject is detected as an outlier for the CP model (for outlier detection, see (Bro, 1997)). In total, there are 140 male and 152 female subjects included in the analysis.
The T0-corrected metabolomics data, i.e., the postprandial data corrected by subtracting the fasting state data, is arranged as a third-order tensor \({\varvec{\mathscr{X}}}\) of size \(I\times J \times K\), where I, J, K denotes the number of subjects (140 males or 152 females), metabolites (161), and time points (7 time points, i.e., fasting state data is subtracted from other time points and excluded in T0-corrected data), respectively. The fasting state data is arranged as a matrix of size \(I\times J\).
In addition to the metabolomics data, additional information such as different body composition, and insulin resistance measures were obtained from the subjects. Weight, height, and waist circumference were directly measured, and consequently BMI and waist/height ratio were calculated. Body composition was estimated by bioelectrical impedance using a Tanita scale (TANITA MC-780MA), to obtain body muscle mass and fat mass, and subsequently, body fat percentage, muscle to fat ratio, fat mass index (fat mass/height2 (kg/m2)), and fat free mass index ((muscle mass/2.2) × 2.20462/height2 (kg/m2)). Insulin resistance was assessed using HOMA-IR (Homeostatic Model Assessment for Insulin Resistance), by the formula: fasting insulin (microU/L) × fasting glucose (nmol/L) / 22.5. Descriptive statistics on these variables stratified by sex are provided in Table S1.2 in Supplementary Material S1.
3 Methods
3.1 CP model
The CP model (Carroll & Chang, 1970; Harshman, 1970; Hitchcock, 1927) approximates a higher-order tensor as the sum of a minimum number of rank-one tensors as in Fig. 1. An R-component CP model of a third-order tensor \({\varvec{\mathscr{X}}} \in {\mathbb{R}}^{I \times J \times K}\) with modes: subjects, metabolites, and time points represents the data as follows:
where \(\circ\) denotes the vector outer product; \({\varvec{\mathbf {\MakeLowercase {a}}}}_{r}\), \({\varvec{\mathbf {\MakeLowercase {b}}}}_{r}\), \({\varvec{\mathbf {\MakeLowercase {c}}}}_{r}\) corresponds to the rth column of factor matrices \({\varvec{\mathbf {\MakeUppercase {A}}}}\in {\mathbb {R}}^{I\times R}, {\varvec{\mathbf {\MakeUppercase {B}}}}\in {\mathbb {R}}^{J\times R}\), and \({\varvec{\mathbf {\MakeUppercase {C}}}}\in {\mathbb {R}}^{K\times R}\), respectively. Each component \(({\varvec{\mathbf {\MakeLowercase {a}}}}_{r},{\varvec{\mathbf {\MakeLowercase {b}}}}_{r},{\varvec{\mathbf {\MakeLowercase {c}}}}_{r})\) may reveal subject groups (in \({\varvec{\mathbf {\MakeLowercase {a}}}}_{r}\)), groups of metabolites (in \({\varvec{\mathbf {\MakeLowercase {b}}}}_{r}\)) related to those subject groups following a specific temporal profile (given by \({\varvec{\mathbf {\MakeLowercase {c}}}}_{r}\)). The CP model is unique up to permutation and scaling ambiguities under mild conditions (Kolda & Bader, 2009). Permutation ambiguity indicates that the order of rank-one components is arbitrary while the scaling ambiguity corresponds to arbitrarily scaling each vector in component r, i.e., (\({\varvec{\mathbf {\MakeLowercase {a}}}}_{r}\), \({\varvec{\mathbf {\MakeLowercase {b}}}}_{r}\), \({\varvec{\mathbf {\MakeLowercase {c}}}}_{r}\)), as long as the product of the norms of the vectors stays the same.
The uniqueness of the CP model without additional constraints on the extracted patterns is an advantage compared to other types of tensor factorizations such as the Tucker decomposition (Tucker, 1966). The uniqueness facilitates the interpretability of the CP components, i.e., when the model is unique, each component \(({\varvec{\mathbf {\MakeLowercase {a}}}}_{r},{\varvec{\mathbf {\MakeLowercase {b}}}}_{r},{\varvec{\mathbf {\MakeLowercase {c}}}}_{r})\) can be interpreted one at a time potentially revealing biomarkers for a specific stratification of subjects. The Tucker decomposition is a more flexible model than the CP model and it is not unique in terms of extracted patterns. Additional constraints on the factor matrices and core array in the Tucker decomposition can achieve uniqueness (Kolda & Bader, 2009). For instance, higher-order singular value decomposition (HOSVD) with orthogonality constraints on all factor matrices as well as all-orthogonal core can help with uniqueness. HOSVD has recently been used to analyze metabolomics measurements collected at multiple time points before and after an oral glucose challenge test from twenty subjects (Fujita et al., 2023). Additional constraints on the factor matrices such as orthogonality as in PCA or HOSVD imposed for uniqueness are not necessarily realistic assumptions, often limiting the interpretability of the methods, especially when the goal is to interpret one component at a time rather than to find subspaces.
The model fit shows how well the CP model describes the data and is defined as follows:
where \(\Vert \cdot \Vert\) denotes the Frobenius norm. If the model fully explains the data, the fit is \(100\%\); otherwise, the unexplained part remains in the residuals.
When reporting the results, we normalize each vector, \({\varvec{\mathbf {\MakeLowercase {A}}}}_{r}\), \({\varvec{\mathbf {\MakeLowercase {B}}}}_{r}\), \({\varvec{\mathbf {\MakeLowercase {C}}}}_{r}\), by its norm due to the scaling ambiguity.
3.1.1 Model selection
Choosing the right number of components, i.e., R, is crucial to extract the underlying patterns accurately; however, the selection of R in a CP model is a challenging task (Håstad, 1990; Kolda & Bader, 2009). Here, we choose R based on the replicability of the extracted components, where replicability is defined as the ability to extract similar patterns from random subsamples of the dataset (Adali et al., 2022), and is an extension of split-half analysis (Harshman & De Sarbo, 1984).
The replicability of an R-component CP model is assessed as follows:
- Step 1.:
-
Randomly split all subjects into ten parts (if there is a class information of potential interest, randomly split the subjects such that each part has the same class proportions as in the original data)
- Step 2.:
-
Leave out one part at a time, and form a subset using the remaining data, i.e., in total, ten subsets,
- Step 3.:
-
Fit an R-component CP model to each subset,
- Step 4.:
-
Calculate the similarity of factors between every pair of CP models (in the metabolites and time points modes), i.e., 45 similarity scores,
- Step 5.:
-
Repeat Step 1–4 ten times using different random splitting of subjects in Step 1.
The similarity of the factors in metabolites and time points modes from two CP models \(\llbracket {\varvec{\mathbf {\MakeUppercase {A}}}}, {\varvec{\mathbf {\MakeUppercase {B}}}}, {\varvec{\mathbf {\MakeUppercase {C}}}} \rrbracket\) and \(\llbracket \bar{{\varvec{\mathbf {\MakeUppercase {A}}}}}, \bar{{\varvec{\mathbf {\MakeUppercase {B}}}}}, \bar{{\varvec{\mathbf {\MakeUppercase {C}}}}} \rrbracket\) is measured using the factor match score (FMS) after finding the best matching permutation:
An R-component CP model is considered to be replicable if \(95\%\) of FMS values (of 450 FMS values computed at the end of Step 5) is higher than 0.9. We choose the highest number of components that produces a replicable CP model in order to explain the data as much as possible. There are various diagnostic approaches used to determine the number of CP components such as the core consistency diagnostic (Bro & Kiers, 2003), and the increase in model fit. These approaches are often used together since no single method can always give the right number of components. Here, we primarily rely on a replicability-based model selection approach since the replicability of the extracted patterns is particularly important when the goal is to reveal biomarkers.
3.2 Experimental setting
Before the analysis, the data is preprocessed by first centering across the subjects mode and then scaling within the metabolites mode (Bro & Smilde, 2003). When scaling, each slice in the metabolites mode is divided by the root mean squared value of non-missing entries.
All experiments are performed in MATLAB (2020b). For fitting the CP model, we use cp-wopt (Acar et al., 2011) from the Tensor Toolbox (version 3.1) (Bader & Kolda, 2022) using the nonlinear conjugate gradient algorithm from the Poblano Toolbox (Dunlavy et al., 2010). Multiple random initializations are used when fitting the model to avoid local minima, and the run returning the minimum function value is used for further analysis. See the Github repositoryFootnote 3 for details. For PCA, we use the MATLAB function svd after estimating the missing values using weighted optimization (Acar et al., 2011).
4 Results
4.1 The CP model of T0-corrected metabolomics data from males reveals a BMI-related group difference
We analyze the T0-corrected data from males using a 2-component CP model. The number of components is selected based on the replicability of extracted patterns. See Supplementary Material S2 for more details on model selection. The model fit is 44%.

2-component CP model of the T0-corrected metabolomics data from males. A Subjects mode (i.e., \({\varvec{\mathbf {\MakeLowercase {a}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {a}}}}_{2}\)), where subjects are colored according to BMI defined as lower BMI: BMI\(<25\) and higher BMI: BMI \(\ge 25\), B Metabolites mode (i.e., \({\varvec{\mathbf {\MakeLowercase {b}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {b}}}}_{2}\)), where metabolites are colored according to lipoprotein classes. The size of the marker for each metabolite is adjusted according to lipoprotein subclasses as indicated in the legend. We show the names of the metabolites with the highest coefficients for the component of interest, i.e., \({\varvec{\mathbf {\MakeLowercase {b}}}}_{2}\). The Rest group contains features other than the ones in the lipoprotein group, the Total group corresponds to total concentrations of certain metabolites, e.g., Total-C, Total-TG. See Tabel S1.1 in Supplementary Material S1 for details, C Time mode (i.e., \({\varvec{\mathbf {\MakeLowercase {c}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {c}}}}_{2}\))

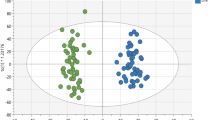
Scatter plots of the first and second component from the 2-component CP model of the T0-corrected data from males. A Subjects mode, (i.e., \({\varvec{\mathbf {\MakeLowercase {a}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {a}}}}_{2}\)) colored according to BMI defined as lower BMI: BMI\(<25\) and higher BMI: BMI \(\ge 25\), and B Metabolites mode, where metabolites are colored according to lipoprotein classes. The size of the marker for each metabolite is adjusted according to lipoprotein subclasses as indicated in the legend. The Rest group contains features other than the ones in the lipoprotein group, the Total group corresponds to total concentrations of certain metabolites, e.g., Total-C, Total-TG. See Table S1.1 in Supplementary Material S1 for details
Figure 2 shows the factors \(({\varvec{\mathbf {\MakeLowercase {a}}}}_{r},{\varvec{\mathbf {\MakeLowercase {b}}}}_{r},{\varvec{\mathbf {\MakeLowercase {c}}}}_{r})\) extracted by the 2-component CP model. To see whether the model reveals any underlying group structure among subjects, we consider additional information available about the participants after the analysis. In the subjects mode (i.e., \({\varvec{\mathbf {\MakeLowercase {a}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {a}}}}_{2}\)) in Fig. 2, subjects are colored according to BMI groups, i.e., lower vs. higher BMI groups. The lower BMI group contains subjects with BMI less than 25.0 (i.e., underweight and normal weight subjects) while the higher BMI group contains subjects with BMI greater than or equal to 25.0 (i.e., overweight and obese subjects). In the metabolites mode (i.e., \({\varvec{\mathbf {\MakeLowercase {b}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {b}}}}_{2}\)), metabolites are colored according to lipoprotein subclasses. Here, the Rest group contains features other than the ones in the lipoprotein group while the Total group consists of total concentrations of certain metabolites, e.g., Total-C, Total-TG. See Supplementary Material S1 (Table S1.1) for details. In the time mode (i.e., \({\varvec{\mathbf {\MakeLowercase {c}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {c}}}}_{2}\)), the model reveals the temporal patterns.
In this model, the second CP component, i.e., (\({\varvec{\mathbf {\MakeLowercase {A}}}}_{2}\), \({\varvec{\mathbf {\MakeLowercase {B}}}}_{2}\), \({\varvec{\mathbf {\MakeLowercase {C}}}}_{2}\)), is of particular interest since it reveals a statistically significant group difference (p-value \(=6 \times 10^{-4}\) using a two-sample t-test based on \({\varvec{\mathbf {\MakeLowercase {A}}}}_{2}\)) in terms of BMI. Supplementary Material S3—Fig. S3.4 gives a more detailed visualization of the second component. In the metabolites mode, i.e., \({\varvec{\mathbf {\MakeLowercase {B}}}}_{2}\) in Fig. 2B, we observe that the subject group difference is due to different behaviour of certain metabolites, i.e., metabolites with high coefficients in terms of absolute value. In particular, we observe that TG-related metabolites, VLDLs (XXL/XL/L), monounsaturated fatty acids (MUFA), saturated fatty acids (SFA), and total fatty acids (Total-FA) have high positive coefficients (indicating that changes in these metabolites positively relate to higher BMI) while several LDL-FCs, VLDLs (M/S), and unsaturation degree have high negative coefficients (indicating that changes in these metabolites negatively relate to higher BMI). Unsaturation degree is the level of unsaturation of fatty acids within a sample, high levels of which indicating that the fatty acid content is likely to be polyunsaturated rather than saturated for a specific sample. Time profiles of the raw data for these specific metabolites, i.e., the ones with high coefficients in terms of absolute value, are included in Supplementary Material S3. In the time mode, \({\varvec{\mathbf {\MakeLowercase {c}}}}_{2}\) increases gradually until 2.5 h and decreases afterwards showing the temporal profile of the metabolic response modelled by this component.
Note that while we discuss subject group differences in terms of BMI, this component also has significant correlation with several other variables. Figure 6 shows the correlation of the factor vector in the subjects mode for the second component, i.e., \({\varvec{\mathbf {\MakeLowercase {A}}}}_{2}\), with various variables of interest. High positive and negative correlations indicate that the second component is related to not only BMI but also a phenotype defined by these closely related variables.
The first component, i.e., (\({\varvec{\mathbf {\MakeLowercase {A}}}}_{1}\), \({\varvec{\mathbf {\MakeLowercase {B}}}}_{1}\), \({\varvec{\mathbf {\MakeLowercase {C}}}}_{1}\)), on the other hand, models an earlier response captured by \({\varvec{\mathbf {\MakeLowercase {c}}}}_{1}\) potentially modelling individual differences. Scatter plots of the first and second component in the subjects and metabolites modes showing subject groups and groups of lipoproteins based on their classes and subclasses are given in Fig. 3.
4.2 Analysis of the fasting state metabolomics data from males using PCA reveals a BMI-related group difference

Scatter plots from PCA of the fasting state data from males: A Subjects mode, where subjects are colored according to BMI defined as lower BMI: BMI\(<25\) and higher BMI: BMI \(\ge 25\), and B Metabolites mode, where metabolites are colored according to lipoprotein classes. The size of the marker for each metabolite is adjusted according to lipoprotein subclasses as indicated in the legend. We show the names of the metabolites with the highest coefficients for the first component, where we observe a statistically significant group difference in terms of BMI. The Rest group contains features other than the ones in the lipoprotein group, and the Total group corresponds to total concentrations of certain metabolites, e.g., Total-C, Total-TG. See Supplementary Material S1 (Table S1.1) for details
Figure 4 shows the scatter plots of principal components in the subjects and metabolites modes from the PCA of the fasting state data. In the subjects mode, a weak but statistically significant (p-value \(=1\times 10^{-3}\)) group difference is captured using the first principal component (PC1). In the metabolites mode, we observe that in PC1, IDLs, LDLs, VLDLs, Apolipoprotein B (ApoB), and Remnant cholesterol (Remnant-C) have high positive loadings showing positive relation with the concentrations of these metabolites and the higher BMI group.

2-component CP model of the T0-corrected metabolomics data from females. A Subjects mode (i.e., \({\varvec{\mathbf {\MakeLowercase {a}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {a}}}}_{2}\)), where subjects are colored according to BMI defined as lower BMI: BMI\(<25\) and higher BMI: BMI \(\ge 25\), B Metabolites mode (i.e., \({\varvec{\mathbf {\MakeLowercase {b}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {b}}}}_{2}\)), where metabolites are colored according to lipoprotein classes. The size of the marker for each metabolite is adjusted according to lipoprotein subclasses as indicated in the legend. The Rest group contains features other than the ones in the lipoprotein group, the Total group corresponds to total concentrations of certain metabolites, e.g., Total-C, Total-TG, and C Time mode (i.e., \({\varvec{\mathbf {\MakeLowercase {c}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {c}}}}_{2}\))
4.3 The CP model of T0-corrected metabolomics data from females does not reveal a BMI-related group difference
Factors captured by the analysis of the T0-corrected data from females using a 2-component CP model are given in Fig. 5 (see Supplementary Material S2 for the selection of the number of components). The fit of the 2-component model is \(47\%\).
In the subjects mode, we do not observe a group difference in either component in terms of the additional information available about the participants, see \({\varvec{\mathbf {\MakeLowercase {a}}}}_{1}\) and \({\varvec{\mathbf {\MakeLowercase {a}}}}_{2}\) in Fig. 5A, where subjects are colored according to lower and higher BMI groups. Note that correlations are also quite low for females in Fig. 6. The time mode shows two underlying profiles: early response in \({\varvec{\mathbf {\MakeLowercase {c}}}}_{1}\), and a later response in \({\varvec{\mathbf {\MakeLowercase {c}}}}_{2}\). Even though the second component is similar in the metabolites and time modes, i.e., \({\varvec{\mathbf {\MakeLowercase {B}}}}_{2}\) and \({\varvec{\mathbf {\MakeLowercase {C}}}}_{2}\) in Fig. 5, to the component captured by the CP model of the T0-corrected data from males, i.e., (\({\varvec{\mathbf {\MakeLowercase {A}}}}_{2}\), \({\varvec{\mathbf {\MakeLowercase {B}}}}_{2}\), \({\varvec{\mathbf {\MakeLowercase {C}}}}_{2}\)) in Fig. 2, no BMI-related group difference is observed in this component for females (see Fig. 8 for a clear comparison of the components from males vs. females).

Correlation between the second component (\({\varvec{\mathbf {\MakeLowercase {a}}}}_{2}\)) of the 2-component CP model of the T0-corrected data in the subjects mode with the additional meta data. Measurements from males and females are analyzed using a CP model separately as shown in Figs. 2 and 5. The second component vector in the subjects mode, i.e., (\({\varvec{\mathbf {\MakeLowercase {a}}}}_{2}\)), from each model is used to compute the correlations for males and females. Descriptions of these variables are as follows: HOMAIR homeostatic model assessment for Insulin Resistance; MuscleFatRatio muscle to fat ratio; FatPercent body fat percentage; MuscleMass amount of muscle in the body (kg); Weight (kg); BMI Body Mass Index; Waist Waist circumferance (cm); WaistHeightRatio waist measurement divided by height (cm); FatMass amount of body fat (kg); FatMassIndex; FFMI Fat Free Mass Index. The mean ± standard deviation for each variable and sex is shown at the bottom of the plot, and more details are provided in Table S1.2 in Supplementary Material S1
4.4 Analysis of the fasting state metabolomics data from females using PCA reveals a BMI-related group difference

Scatter plots from PCA of the fasting-state data from females: A Subjects mode, where subjects are colored according to BMI defined as lower BMI: BMI\(<25\) and higher BMI: BMI \(\ge 25\), and B Metabolites mode, where metabolites are colored according to lipoprotein classes. The size of the marker for each metabolite is adjusted according to lipoprotein subclasses as indicated in the legend. We show the names of the metabolites with the highest coefficients for the fourth component, where we observe a statistically significant group difference in terms of BMI. The Rest group contains features other than the ones in the lipoprotein group, the Total group corresponds to total concentrations of certain metabolites, e.g., Total-C, Total-TG
Scatter plots of PC1 and PC4 in the subjects and metabolites modes from PCA of the fasting state data from females are shown in Fig. 7. In the subjects mode, only PC4 reveals a statistically significant (p-value \(=5\times 10^{-6}\)) BMI-related group difference. Note that the fourth component only explains 5% of the variation in the data so the signal—although statistically significant—is weak in this data set. In the metabolites mode, we observe that in PC4, HDLs (S), C-peptide, glucose, glycoprotein acetyls (GlycA), total concentration of lipoprotein particles (Total-P), and valine (Val) have high positive loadings (positively relate to higher BMI) while the HDLs (XL), average diameter for HDL particles (HDL size), glycine (Gly), and XS-VLDL-PL have high negative loadings (negatively relate to higher BMI).
We summarize the metabolites observed to be related to BMI-related group difference in Table 1.
5 Discussion
5.1 Fasting vs. dynamic states
In males, groups of lipoproteins reveal BMI-related group difference, and the relations of those lipoproteins with BMI groups differ in the fasting vs. dynamic state. For the fasting state data, the positive relation of LDL-C (Bays et al., 2013; Lamon-Fava et al., 1996), VLDL-C (Lamon-Fava et al., 1996) and ApoB (Lamon-Fava et al., 1996) with higher BMI has been previously reported, and our findings are consistent with the literature. Likewise, we find similar patterns whereby large triglyceride rich VLDL lipoproteins are increased in individuals with higher BMI following a meal challenge containing fats assessed via conventional AUC postprandial metrics (Wang et al., 2017). However, we did not find complementary decreases in HDL, this may be in part due to gender stratification differences in our analyses. Likewise monozygotic twin studies point to that these observed differences in lipoprotein responses are driven by BMI differences, rather than underlying genetics (Rämö et al., 2017).
While Table 1 gives a summary of the most important metabolites, i.e. metabolites with high coefficients in terms of absolute value, we also observe in Fig. 4 that HDLs (XL/L) negatively relate to higher BMI in the fasting state (even though they are not among the most important metabolites; therefore, not reported in Table 1). This matches with previous results in terms of HDL-C (Bays et al., 2013; Lamon-Fava et al., 1996).
This study differs from previous challenge test studies as it is a comprehensive study in terms of the lipoproteins (classes and subclasses) considered in the analysis, and the comprehensive picture provided by the CP model from the postprandial data as well as its comparison with the fasting state analysis. We observe that dynamic and fasting state data provide different views of the same set of metabolites. These different views reveal different relations with the BMI groups as we observe in the case of VLDLs, i.e., while concentrations of all VLDLs positively relate to higher BMI in the fasting state (see Fig. 4), Fig. 2 shows that changes in VLDLs (XXL/XL/L) positively relate to higher BMI, and changes in VLDLs (M/S) negatively relate to higher BMI in the dynamic state.
These results not only improve our understanding of dynamic changes in the human metabolome but also reveal dynamic and static markers through the analysis of fasting and postprandial data.
5.2 Males vs. females (dynamic state)
Patterns of dynamic response to the challenge test, i.e., patterns in the metabolites and time modes in T0-corrected analyses, are similar in males vs. females. Figure 8 compares the components extracted using CP models from males vs. females in the metabolites and time modes. Figure 8A and C show the comparison of \({\varvec{\mathbf {\MakeLowercase {b}}}}_{1}\) from males vs. females, and \({\varvec{\mathbf {\MakeLowercase {c}}}}_{1}\) from males vs. females, respectively. The second components (i.e., \({\varvec{\mathbf {\MakeLowercase {b}}}}_{2}\) and \({\varvec{\mathbf {\MakeLowercase {c}}}}_{2}\)), which are the patterns of interest in terms of BMI-related group difference, are compared in Fig. 8B and D. We observe similar patterns in both modes indicating that patterns of dynamic response to the challenge test are similar in males and females.

Comparison of CP models of T0-corrected data from males and females. A and B Metabolites mode. For better comparison, the line \(y=x\) is plotted together with \(y=x-0.05\) and \(y=x+0.05\). C and D Time mode
We strengthen this argument by analyzing the T0-corrected data from all individuals (both males and females) using a CP model. When males and females are analyzed together, the components in the metabolites and time modes are the same for both sexes. In the subjects mode, BMI-related group difference is observed among males but still not among females (See Supplementary Material S4).
The fat distribution is usually quite different for males and females, with overweight males often having central obesity, i.e., adiposity (consisting of TGs) in and around the liver, where VLDL is produced. Young females have more subcutaneous fat than men and also deposit more fat non-centrally, e.g., on other places such as hips, thighs etc.; pre-menopausal women also export less TG from the liver, because they deposit it elsewhere, and the VLDL may therefore contain less TG and pack more efficiently, so the observed gender differences can be linked to the effect of such anthropometric differences on TG metabolism (Palmisano et al., 2018). The presence of gender differences can also be anticipated due to the inherent variances in sex hormones, psychosocial factors, and cardiovascular risk factor profiles (Loh et al., 2022). These postprandial gender differences may in part explain the cardiovascular risk reduction noted in pre-menopausal women (Zaman et al., 2012). Note that our cohort comprises individuals who are 18 years old from both sexes, and this uniformity allows us to infer that the observed sex-based differences might mirror the variations in cardiovascular risk profiles. While our study’s strength lies in the uniform age of the cohort, further research is needed to clarify these relationships and to better understand how our findings might extend to different age demographics and clinical domains.
5.3 Males vs. females (fasting state)
In the fasting state, the main patterns in the metabolites mode are similar in males vs. females while the relations of those patterns with BMI differ. Figures 4 and 7 show the scatter plots of PCs in the fasting state analysis of males and females, respectively. Similar patterns are observed for both males and females. More precisely, in PC1s (in Figs. 4 and 7), IDLs, LDLs, and VLDLs are the metabolites with high positive coefficients. While PC1 shows strong signals from lipoproteins in both males and females, it shows BMI-related group difference only in males. This could again be linked to anthropometric differences between genders. BMI in overweight young women is not strongly associated with abdominal obesity so we do not see PC1 as related to BMI. In men, abdominal fat is strongly associated with BMI, so BMI is a good proxy. This component is not only related to BMI but also to waist circumference, fat mass, and fat mass index which would all indicate that excess adiposity is probably more relevant to this component than BMI (which is also affected by muscle mass).
In females, PC4 in Fig. 7 reveals a statistically significant group difference in terms of BMI, and the metabolites with high positive coefficients are mainly the HDLs (S) and the ones with high negative coefficients are mainly the HDLs (XL). For males, we also observe in PC1 (even though the coefficients are small) that HDLs (S) positively relate to higher BMI while HDLs (XL) negatively relate to the higher BMI group capturing the relation revealed by PC4 in females. Thus, for both males (from PC1 in Fig. 4) and females (from PC4 in Fig. 7), we can say that HDLs (S) are positively related to higher BMI and HDLs (XL) are negatively related to higher BMI. In addition, in PC4 for females, we observe metabolites related to insulin sensitivity and inflammation, indicating that the obese and overweight women have higher glucose, C-peptide (marker of insulin synthesis), valine (a branched-chain amino acid known to increase in subjects being less insulin-sensitive (Zhao et al., 2016)), and GlycA (an inflammatory marker (Otvos et al., 2015))
These differences between genders may also be explained in the pre-menopausal cardioprotective lipid profiles of females (Kilim & Chandala, 2013). This gender disparity in postprandial atherogenic remnant lipoproteins may help explain why males are at greater cardiovascular risk in early life. Literature suggests gender differences in cardiovascular risk are abolished after menopause (Rich-Edwards et al., 1995), and this may point towards the sex hormones playing a moderating role on atherogenic lipoproteins.
6 Conclusion
In this study, we have demonstrated a comprehensive analysis of metabolomics measurements collected during a challenge test from the COPSAC2000 cohort. We have arranged time-resolved metabolomics measurements as a subjects by metabolites by time points tensor, and analyzed fasting state-corrected dynamic data using a CP model in comparison with the fasting state data. Our analysis demonstrates that the CP model reveals interpretable patterns showing how subject groups (i.e., lower vs. higher BMI groups) differ in terms of the dynamic response of certain metabolite groups. We also show that metabolites behave differently in fasting vs. dynamic states; therefore, analysis of fasting and postprandial data can reveal static as well as dynamic biomarkers. In addition, our results indicate that patterns of dynamic response to the challenge test are similar in males vs. females but differences are observed in terms of how those patterns relate to BMI groups. Extracted patterns have shown to be not only interpretable but also replicable.
While the CP model can reveal a comprehensive picture of time-resolved postprandial data and novel insights such as sex differences and metabolite groups behaving differently in fasting vs. dynamic states, there are still several limitations. The set of measured metabolites/features is dominated by lipoproteins; therefore, the main patterns of variation in the data mainly model the behavior of lipoproteins, which is the case for both CP and PCA. We do not observe glycolysis-related metabolites, amino acids or ketone bodies well-modelled due to the dominant set of lipoproteins. In order to get a better understanding of postprandial metabolic response in terms of glycolysis, ketogenesis, and the aminoacid metabolism, metabolites may be grouped into specific subsets and jointly analyzed. Also, in this study, we have mainly focused on one pattern,i.e., one CP component or PCA component, at a time (even if scatter plots are considered). However, multiple components together may explain the meta variables of interest, e.g., BMI groups, better. For both males and females, when multiple linear regression is used to predict BMI values using the factor matrix in the subjects mode, we do not observe any improvement using multiple components. However, in PCA, multiple components can be relevant. As we are not interested in a specific variable but rather in capturing components that may relate to a certain phenotype (characterized by multiple variables), we have focused on one component at a time and studied the correlation of the subject scores with the available meta variables of interest here. Depending on the goal, multiple components can be clustered or used in further prediction tasks. We should also mention that as an unsupervised method, the CP model focuses on the main patterns explaining the variation in the data. These main patterns may not necessarily be related to meta variables of interest. Such an unsupervised approach may miss the relation with meta variables if that relation does not significantly contribute to the main variation in the data. In those cases, supervised methods may perform better. However, the goal of using an unsupervised approach in this exploratory study is its potential to reveal unknown subject stratifications rather than predicting meta variables.
Another future research direction to consider is the joint analysis of postprandial metabolomics data with other omics data sets, in particular, gut microbiome data, through data fusion methods (Acar et al., 2015; Schenker et al., 2021). With recent technological advances, e.g., multi-omics microsampling (Shen et al., 2023), that may make such challenge test data more easily available, jointly analyzing dynamic metabolomics data with other omics data sets becomes even more crucial.
7 Supplementary information
Supplementary files contain the full list of metabolites, descriptive statistics of meta variables, selection of the number of components for the CP model, supplementary figures and comparison of CP models from males and females.
Data availability
NMR measurements of plasma samples are not publicly available but may be shared by COPSAC through a collaboration agreement. Data access requests should be directed to Morten A. Rasmussen (morten.arendt@dbac.dk). The GitHub repository https://github.com/eacarat/CP_Metabolomics_MealChallenge contains the scripts showing the preprocessing steps and analyzing the data using a CP model as well as the scripts for replicability.
Notes
While our study is nested within the COPSAC2000 mother-child cohort, which at its inception recruited mothers with a history of asthma, this current study is not intended, nor describes any connections with asthma.
The full list of metabolites is available at https://research.nightingalehealth.com/biomarkers.
References
Acar, E., & Yener, B. (2008). Unsupervised multiway data analysis: A literature survey. IEEE Transactions on Knowledge and Data Engineering, 21(1), 6–20.
Acar, E., Aykut-Bingol, C., Bingol, H., et al. (2007). Multiway analysis of epilepsy tensors. Bioinformatics, 23(13), i10–i18.
Acar, E., Dunlavy, D. M., Kolda, T. G., et al. (2011). Scalable tensor factorizations for incomplete data. Chemometrics and Intelligent Laboratory Systems, 106(1), 41–56.
Acar, E., Bro, R., & Smilde, A. K. (2015). Data fusion in metabolomics using coupled matrix and tensor factorizations. Proceedings of the IEEE, 103, 1602–1620.
Adali, T., Kantar, F., Akhonda, M. A. B. S., et al. (2022). Reproducibility in matrix and tensor decompositions: Focus on model match, interpretability, and uniqueness. IEEE Signal Processing Magazine, 39(4), 8–24.
Bader, B. W., & Kolda, T. G., et al. (2022) Tensor Toolbox for MATLAB, Version 3.1. www.tensortoolbox.org
Bastarrachea, R. A., Laviada-Molina, H. A., Nava-Gonzalez, E. J., et al. (2018). Deep multi-OMICs and multi-tissue characterization in a pre- and postprandial state in human volunteers: The GEMM family study research design. Genes, 9(11), 532.
Bays, H. E., Toth, P. P., Kris-Etherton, P. M., et al. (2013). Obesity, adiposity, and dyslipidemia: A consensus statement from the national lipid association. Journal of Clinical Lipidology, 7(4), 304–383.
Becker, F., Smilde, A. K., & Acar, E. (2023). Unsupervised EHR-based phenotyping via matrix and tensor decompositions. WIREs Data Mining and Knowledge Discovery, 13(4), e1494.
Bermingham, K. M., Mazidi, M., Franks, P. W., et al. (2023). Characterisation of fasting and postprandial nmr metabolites: Insights from the ZOE PREDICT 1 study. Nutrients, 15(11), 2638.
Berry, S. E., Valdes, A. M., Drew, D. A., et al. (2020). Human postprandial responses to food and potential for precision nutrition. Nature Medicine, 26(6), 964–973.
Bisgaard, H. (2004). The Copenhagen prospective study on asthma in childhood (COPSAC): Design, rationale, and baseline data from a longitudinal birth cohort study. Annals of Allergy, Asthma & Immunology, 93(4), 381–389.
Bondia-Pons, I., Maukonen, J., Mattila, I., et al. (2014). Metabolome and fecal microbiota in monozygotic twin pairs discordant for weight: A Big Mac challenge. The FASEB Journal, 28(9), 4169–4179.
Botham, K. M., & Wheeler-Jones, C. P. (2013). Postprandial lipoproteins and the molecular regulation of vascular homeostasis. Progress in Lipid Research, 52(4), 446–464.
Bro, R. (1997). PARAFAC. Tutorial and applications. Chemometrics and Intelligent Laboratory Systems, 38(2), 149–171.
Bro, R., & Kiers, H. A. L. (2003). A new efficient method for determining the number of components in PARAFAC models. Journal of Chemometrics, 17(5), 274–286.
Bro, R., & Smilde, A. K. (2003). Centering and scaling in component analysis. Journal of Chemometrics, 17(1), 16–33.
Carroll, J. D., & Chang, J. J. (1970). Analysis of individual differences in multidimensional scaling via an N-way generalization of “Eckart-Young" decomposition. Psychometrika, 35(3), 283–319.
Dunlavy, D. M., Kolda, T. G., & Acar, E. (2010). Poblano v1.0: A Matlab toolbox for gradient-based optimization. Tech. Rep. SAND2010-1422, Sandia National Laboratories, https://www.osti.gov/servlets/purl/989350
Erdos, B., Westerhuis, J. A., Adriaens, M. E., et al. (2023). Analysis of high-dimensional metabolomics data with complex temporal dynamics using RM-ASCA+. PLoS Computational Biology, 19(6), e1011221.
Fujita, S., Karasawa, Y., Hironaka, K., et al. (2023). Features extracted using tensor decomposition reflect the biological features of the temporal patterns of human blood multimodal metabolome. PLoS ONE, 18(2), e0281594.
Gardlo, A., Smilde, A. K., Hron, K., et al. (2016). Normalization techniques for PARAFAC modeling of urine metabolomic data. Metabolomics, 12(7), 1–13.
Harshman, R. A. (1970). Foundations of the PARAFAC procedure: Models and conditions for an “explanatory" multimodal factor analysis. UCLA Working Papers in Phonetics, 16, 1–84.
Harshman, R. A., & De Sarbo, W. S. (1984). An application of PARAFAC to a small sample problem, demonstrating preprocessing, orthogonality constraints, and split-half diagnostic techniques. Research Methods for Multimode Data Analysis (pp. 602–642). New York: Praeger.
Harte, A. L., Varma, M. C., Tripathi, G., et al. (2012). High fat intake leads to acute postprandial exposure to circulating endotoxin in type 2 diabetic subjects. Diabetes Care, 35(2), 375–382.
Håstad, J. (1990). Tensor rank is NP-complete. Journal of Algorithms, 11(4), 644–654.
Hitchcock, F. L. (1927). The expression of a tensor or a polyadic as a sum of products. Journal of Mathematics and Physics, 6(1), 164–189.
Kilim, S. R., & Chandala, S. R. (2013). A comparative study of lipid profile and oestradiol in pre- and post-menopausal women. Journal of Clinical & Diagnostic Research, 7(8), 1596–1598.
Kolda, T. G., & Bader, B. W. (2009). Tensor decompositions and applications. SIAM Review, 51(3), 455–500.
Kruskal, J. B. (1977). Three-way arrays: Rank and uniqueness of trilinear decompositions, with application to arithmetic complexity and statistics. Linear Algebra and its Applications, 18(2), 95–138.
Kumar, A. A., Satheesh, G., Vijayakumar, G., et al. (2020). Postprandial metabolism is impaired in overweight normoglycemic young adults without family history of diabetes. Scientific Reports, 10(1), 353.
LaBarre, J. L., Hirschfeld, E., Soni, T., et al. (2021). Comparing the fasting and random-fed metabolome response to an oral glucose tolerance test in children and adolescents: Implications of sex, obesity, and insulin resistance. Nutrients, 13(10), 3365.
Lairon, D., Lopez-Miranda, J., & Williams, C. (2007). Methodology for studying postprandial lipid metabolism. European Journal of Clinical Nutrition, 61(10), 1145–1161.
Lamon-Fava, S., Wilson, P. W., & Schaefer, E. J. (1996). Impact of body mass index on coronary heart disease risk factors in men and women. Arteriosclerosis, Thrombosis, and Vascular Biology, 16(12), 1509–1515.
Lépine, G., Tremblay-Franco, M., Bouder, S., et al. (2022). Investigating the postprandial metabolome after challenge tests to assess metabolic flexibility and dysregulations associated with cardiometabolic diseases. Nutrients, 14(3), 472.
Li, L., Hoefsloot, H., de Graaf, A. A., et al. (2022). Exploring dynamic metabolomics data with multiway data analysis: A simulation study. BMC Bioinformatics, 23(1), 31.
Li, L., Yan, S., Bakker, B. M., et al. (2024). Analyzing postprandial metabolomics data using multiway models: A simulation study. BMC Bioinformatics, 25, 94.
Loh, X., Sun, L., Allen, J. C., et al. (2022). Gender differences in fasting and postprandial metabolic traits predictive of subclinical atherosclerosis in an asymptomatic chinese population. Scientific Reports, 12(1), 16890.
Martino, C., Shenhav, L., Marotz, C. A., et al. (2021). Context-aware dimensionality reduction deconvolutes gut microbial community dynamics. Nature Biotechnology, 39(2), 165–168.
Müllner, E., Röhnisch, H. E., von Brömssen, C., et al. (2021). Metabolomics analysis reveals altered metabolites in lean compared with obese adolescents and additional metabolic shifts associated with hyperinsulinaemia and insulin resistance in obese adolescents: A cross-sectional study. Metabolomics, 17(1), 1–13.
O’Keefe, J. H., & Bell, D. S. (2007). Postprandial hyperglycemia/hyperlipidemia (postprandial dysmetabolism) is a cardiovascular risk factor. The American Journal of Cardiology, 100(5), 899–904.
Otvos, J. D., Shalaurova, I., Wolak-Dinsmore, J., et al. (2015). Glyca: A composite nuclear magnetic resonance biomarker of systemic inflammation. Clinical Chemistry, 61(5), 714–723.
Palmisano, B. T., Zhu, L., Eckel, R. H., et al. (2018). Sex differences in lipid and lipoprotein metabolism. Molecular Metabolism, 15, 45–55.
Pellis, L., van Erk, M. J., van Ommen, B., et al. (2012). Plasma metabolomics and proteomics profiling after a postprandial challenge reveal subtle diet effects on human metabolic status. Metabolomics, 8(2), 347–359.
Poppitt, S. D. (2005). Postprandial lipaemia, haemostasis, inflammatory response and other emerging risk factors for cardiovascular disease: The influence of fatty meals. Current Nutrition & Food Science, 1(1), 23–34.
Rich-Edwards, J. W., Manson, J. E., Hennekens, C. H., et al. (1995). The primary prevention of coronary heart disease in women. New England Journal of Medicine, 332(26), 1758–1766.
Rizi, E. P., Baig, S., Loh, T. P., et al. (2019). Two-hour postprandial lipoprotein particle concentration differs between lean and obese individuals. Frontiers in Physiology, 10, 856.
Rämö, J. T., Kaye, S. M., Jukarainen, S., et al. (2017). Liver fat and insulin sensitivity define metabolite profiles during a glucose tolerance test in young adult twins. The Journal of Clinical Endocrinology & Metabolism, 102(1), 220–231.
Saito, K., Hattori, K., Andou, T., et al. (2020). Characterization of postprandial effects on CSF metabolomics: A pilot study with parallel comparison to plasma. Metabolites, 10(5), 185.
Schenker, C., Cohen, J., & Acar, E. (2021). A flexible optimization framework for regularized matrix-tensor factorizations with linear couplings. IEEE Journal of Selected Topics in Signal Processing, 15(3), 506–521.
Shen, X., Kellogg, R., Panyard, D. J., et al. (2023). Multi-omics microsampling for the profiling of lifestyle-associated changes in health. Nature Biomedical Engineering, 8(1), 11–29.
Smilde, A. K., Geladi, P., & Bro, R. (2004). Multi-way analysis with applications in the chemical sciences. Wiley.
Smilde, A. K., Westerhuis, J. A., Hoefsloot, H. C. J., et al. (2010). Dynamic metabolomic data analysis: A tutorial review. Metabolomics, 6(1), 3–17.
Stroeve, J. H. M., van Wietmarschen, H., Kremer, B. H. A., et al. (2015). Phenotypic flexibility as a measure of health: The optimal nutritional stress response test. Genes & Nutrition, 10(3), 1–21.
Tucker, L. R. (1966). Some mathematical notes on three-mode factor analysis. Psychometrika, 31, 279–311.
Wang, F., Lu, H., Liu, F., et al. (2017). Consumption of a liquid high-fat meal increases triglycerides but decreases high-density lipoprotein cholesterol in abdominally obese subjects with high postprandial insulin resistance. Nutrition Research, 43, 82–88.
Williams, A. H., Kim, T. H., Wang, F., et al. (2018). Unsupervised discovery of demixed, low-dimensional neural dynamics across multiple timescales through tensor component analysis. Neuron, 98(6), 1099-1115.e8.
Wojczynski, M. K., Glasser, S. P., Oberman, A., et al. (2011). High-fat meal effect on LDL, HDL, and VLDL particle size and number in the genetics of lipid-lowering drugs and diet network (GOLDN): An interventional study. Lipids in Health and Disease, 10(1), 181.
Wopereis, S., Stroeve, J. H. M., Stafleu, A., et al. (2017). Multi-parameter comparison of a standardized mixed meal tolerance test in healthy and type 2 diabetic subjects: The PhenFlex challenge. Genes & Nutrition, 12(21), 1–14.
Yin, K., Afshar, A., Ho, J. C., et al. (2020). LogPar: Logistic PARAFAC2 factorization for temporal binary data with missing values. In: KDD’20: Proceedings of the 26th ACM SIGKDD International Conference on Knowledge Discovery & Data Mining, pp 1625–1635.
Zaman, G. S., Rahman, S., & Rahman, J. (2012). Postprandial lipemia in pre- and postmenopausal women. Journal of Natural Science, Biology, and Medicine, 3(1), 65–70.
Zeevi, D., Korem, T., Zmora, N., et al. (2015). Personalized nutrition by prediction of glycemic responses. Cell, 163(5), 1079–1094.
Zhao, X., Han, Q., Liu, Y., et al. (2016). The relationship between branched-chain amino acid related metabolomic signature and insulin resistance: A systematic review. Journal of Diabetes Research, 2016, 1–12.
Zivkovic, A. M., Wiest, M. M., Nguyen, U., et al. (2009). Assessing individual metabolic responsiveness to a lipid challenge using a targeted metabolomic approach. Metabolomics, 5, 209–218.
Acknowledgements
We thank the children and families of the COPSAC\({_{2000}}\) cohort for their contribution, and the clinical team at COPSAC for conducting the clinical testing.
Funding
Open access funding provided by OsloMet - Oslo Metropolitan University. This work was supported in part by the Research Council of Norway through project 300489 and Novo Nordisk Foundation Grant NNF19OC0057934.
Author information
Authors and Affiliations
Contributions
M.A.R., A.K.S. and E.A. designed the research; B.C. and M.A.R. curated the data; S.Y. carried out the data analysis; L.L. and E.A. validated the results; D.H., P.E., L.O.D., and M.A.R. interpreted the results; E.A., L.L., S.Y. and A.K.S. wrote the paper.
Corresponding author
Ethics declarations
Conflict of interest
Authors declare no Conflict of interest and no conflict of interest that are relevant to the content of this article.
Ethical approval
The COPSAC2000 study was conducted in accordance with the Declaration of Helsinki and was approved by the Copenhagen Ethics Committee (KF 01-289/96 and H-16039498) and the Danish Data Protection Agency (2015-41-3696).
Consent to participate
Both parents gave written informed consent before enrollment. At the COPSAC2000 18-year visit, the study participants gave written consent themselves.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yan, S., Li, L., Horner, D. et al. Characterizing human postprandial metabolic response using multiway data analysis. Metabolomics 20, 50 (2024). https://doi.org/10.1007/s11306-024-02109-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11306-024-02109-y