
Abstract
Background
Postprandial lipemia (PPL) is likely a risk factor for cardiovascular disease but these changes have not been well described and characterized in a large cohort. We assessed acute changes in the size and concentration of total and subclasses of LDL, HDL, and VLDL particles in response to a high-fat meal. Participants (n = 1048) from the Genetics of Lipid-Lowering Drugs and Diet Network (GOLDN) Study who ingested a high-fat meal were included in this analysis. Lipids were measured at 0 hr (fasting), 3.5 hr, and 6 hr after a standardized fat meal. Particle size distributions were determined using nuclear magnetic resonance spectroscopy. Analyses were stratified by baseline triglycerides (normal vs. elevated) and gender. The effect of PPL on changes in lipoprotein subclasses was assessed using repeated measures ANOVA.
Results
Postprandially, LDL-C, HDL-C, VLDL-C, and triglycerides increased regardless of baseline triglyceride status, with the largest increases in VLDL-C and TG; however, those with elevated triglycerides demonstrated larger magnitude of response. Total LDL particle number decreased over the 6-hour time interval, mostly from a decrease in the number of small LDL particles. Similarly, total VLDL particle number decreased due to reductions in medium and small VLDL particles. Large VLDL particles and chylomicrons demonstrated the largest increase in concentration. HDL particles demonstrated minimal overall changes in total particle number.
Conclusions
We have characterized the changes in LDL and VLDL particle number, and their subclass patterns following a high-fat meal.
Similar content being viewed by others


Background
Most studies use the measurement of fasting lipoproteins to assess cardiovascular disease (CVD) risk [1, 2]; however, humans spend most of the day in a postprandial state. Postprandial lipemia (PPL) is a physiological response occurring 2 to 12 hours after consuming a meal [3]. Gross lipid profile changes during PPL include increases in triglyceride (TG), very low-density lipoprotein cholesterol (VLDL-C), and chylomicron concentrations; decreases in high-density lipoprotein cholesterol (HDL-C) concentration; and little to no change in low-density lipoprotein cholesterol (LDL-C) concentration. Each of these lipoprotein classes is, however, heterogeneous and composed of various subclasses [4–6]. Lipoprotein particles are composed of different proportions of cholesterol ester (CE) and TG, and each subclass also has specific apolipoproteins on its surface for recognition of receptors and enzymes [7]. Constituent fractions of the lipoprotein profile can vary with respect to particle size, particle number, and particle concentrations [8]. The ingestion of a meal high in fat leads to distributional shifts of VLDL, LDL (including intermediate density lipoproteins (IDL)), and HDL particle sizes, numbers, and plasma concentrations [9]. The various particle sizes have also demonstrated both pro-atherogenic (large VLDL and small LDL) and anti-atherogenic (large LDL) effects [10]. Although much research examines the major lipoproteins and lipid particle sizes in relation to cardiovascular disease, only a few relatively small studies have used the interventional methodology described here to examine changes in lipoprotein distribution and subclasses in the postprandial state following a high-fat challenge [11–16].
Thus, we sought to describe in a large cohort, acute changes in lipoprotein (LDL, HDL, and VLDL) particle sizes and numbers following a high-fat meal and to determine if the lipoprotein particle changes would vary by baseline triglyceride concentrations and gender.
Results
Population characteristics
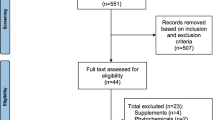
Table 1 shows the characteristics of the study participants according to gender and baseline triglyceride status. These 1048 subjects were part of a cohort of 1328 who were contacted, of whom 1123 participated (Figure 1). Of the 1048 subjects in the analysis, all were white, about half were female, and the mean age of the study group was 48.2 (18-87) years old (Table 1). The hyper-triglyceridemic individuals differed significantly from normal individuals in all baseline traits except smoking status.

GOLDN study recruitment.
Plasma lipid concentrations
Total LDL-C levels demonstrated decreases between 0 and 6 hours following the high-fat meal (113 to 110 mg/dL in the normo-triglyceridemic men; and no change among hyper-triglyceridemic men, p < 0.0001, Figure 2). Among women, LDL-C levels had small but significant (p < 0.0001) increases between 0 and 6 hours (103 to 107 mg/dL among normo-triglyceridemic women and 128 to 138 mg/dL among hyper-triglyceridemic women, Figure 2).

Low-density lipoprotein cholesterol response to high-fat meal, stratified by gender and baseline triglycerides.
Among both normo- and hyper-triglyceridemic men, HDL-C levels displayed a decrease at 3.5 hours but returned to baseline levels by 6 hours, with normo-triglyceridemic men having higher HDL-C concentrations than hyper-triglyceridemic men. Normo-triglyceridemic women also had a decrease in HDL-C levels at 3.5 hours followed by a return to levels slightly above baseline at 6 hours; however, hyper-triglyceridemic women did not demonstrate a decrease at 3.5 hours and at 6 hours increased above baseline (Figure 3). Similar to men, normo-triglyceridemic women had higher HDL-C levels than hyper-triglyceridemic women. These time and triglyceridemic effects were significant (p < 0.0001).

High-density lipoprotein cholesterol response to high-fat meal, stratified by gender and baseline triglycerides.
All groups had a similar pattern of statistically significant changes in VLDL-C levels in response to the high-fat meal: large increases at 3.5 hours were followed by a decrease at 6 hours; however, VLDL-C levels were still elevated above baseline at 6 hours (p < 0.0001). Further, the magnitude of the VLDL-C levels was higher among men than women and among hyper-triglyceridemic compared to normo-triglyceridemic individuals (Figure 4). Total triglycerides increased significantly from 83 to 145 mg/dL in the normo-triglyceridemic men and from 188 to 307 mg/dL in hyper-triglyceridemic men (p < 0.0001). Women displayed similar pattern of statistically significant changes (p < 0.0001, Figure 5).

Very low-density lipoprotein cholesterol response to high-fat meal, stratified by gender and baseline triglycerides.

Triglyceride response to high-fat meal, stratified by gender and baseline triglycerides.
LDL particle size subclasses
Hyper-triglyceridemic participants had higher baseline concentrations of all LDL particle subclasses compared to normo-triglyceridemic participants (Tables 2 and 3). Additionally hyper- and normo-triglyceridemic participants displayed similar patterns of change in LDL particle size concentrations. In both the normo- and hyper-triglyceridemic men and women, there was a small but significant (p < 0.0001) decrease in total LDL particle number following the high-fat meal (normo-triglyceridemic: 1212 to 1195 nmol/L in men, and 1086 to 1076 nmol/L in women); however, the concentrations of IDL and large LDL particles demonstrated significant increases (IDL, normo-triglyceridemic: 28 to 43 nmol/L in men and 22 to 41 nmol/L in women, p < 0.0001; large LDL, normo-triglyceridemic: 406 to 452 nmol/L in men and 457 to 527 nmol/L in women, p < 0.0001), and the concentration of small LDL particles demonstrated a significant decrease (normo-triglyceridemic: 787 to 703 nmol/L in men and 611 to 518 nmol/L in women, p < 0.0001). This large decrease in small LDL particle concentration is responsible for the overall decrease in total LDL particle number (p < 0.0001). Despite significant (p < 0.0001) PPL changes, LDL particle size did not demonstrate substantial changes.
HDL particle size subclasses
In response to a high-fat meal, normo- and hyper-triglyceridemic men displayed similar patterns of change in HDL particle size concentration distributions; however, the magnitudes of these patterns differed in hyper-compared to normo-triglyceridemic participants. In men, there was no effect on large HDL particle concentrations, but significant increases in the concentration of medium HDL particles (3 to 7 mmol/L in normo-triglyceridemics and 2 to 5 mmol/L in hyper-triglyceridemics, p < 0.0001) as well as significant decreases in small HDL particle concentrations (21 to 18 mmol/L in normo-triglyceridemics and 24 to 21 mmol/L in hyper-triglyceridemics, p < 0.0001). Furthermore, total HDL particle number significantly increased among men in response to the high-fat meal (p < 0.0001), which can be attributed to the large increases in medium HDL particle size concentrations.
Among both normo- and hyper-triglyceridemic individuals, women responded differently to the high-fat meal than men. Normo-triglyceridemic women demonstrated a significant decrease in total HDL particle number (p < 0.0001), while there was no overall change among hyper-triglyceridemic women in response to the meal (33 to 32 mmol/L in normo-triglyceridemic and 35 mmol/L in hyper-triglyceridemic). Additionally, these subgroups demonstrated different responses for large HDL particle concentrations where normo-triglyceridemic women displayed significant increases (8 to 9 mmol/L), and hyper-triglyceridemic women displayed significant decreases in large HDL particle concentration (6 to 5 mmol/L, p = 0.0367). For both normo- and hyper-triglyceridemic women, medium and small HDL particle concentrations demonstrated similar responses to the high-fat meal: medium HDL particle concentrations had significant increases (5 to 10 mmol/L for both subgroups, p < 0.0001), and small HDL particle concentrations had significant decreases (20 to 13 mmol/L for normo-triglyceridemics and 24 to 18 mmol/L for hyper-triglyceridemics, p < 0.0001). Similar to LDL size, no substantial change in HDL size was demonstrated in response to the high-fat meal despite its statistical significance (p < 0.0001).
VLDL particle size subclasses
Results of the PPL VLDL particle size subclass analysis were similar in pattern for men and women, but the magnitude of change and concentration values were different, with women, on average, having lower concentrations. Total VLDL particle number decreased significantly among both normo- and hyper-triglyceridemics (among men, 48 to 42 nmol/L for normo- and 100 to 83 nmol/L for hyper-triglyceridemics, p < 0.0001). Large VLDL particle concentrations increased significantly (1 to 3 nmol/L among normo-triglyceridemic men and 6 to 10 nmol/L among hyper-triglyceridemic men, p < 0.0001). Medium and small VLDL particle concentrations demonstrated significant decreases (p < 0.0001 for both men and women). Finally, VLDL particle size significantly increased (p < 0.0001) among normo- and hyper-triglyceridemic men and women.
Discussion
The present study focused on the PPL response to an acute (1-time) high-fat meal using data derived from the Genetics of Lipid-Lowering Drugs and Diet Network (GOLDN) Study. We sought to describe lipoprotein particle subclass number, size, and concentrations stratified by gender and baseline triglyceride status. Although the pattern of change observed for both men and women was qualitatively similar, the magnitude of these changes appeared to be more exaggerated in men compared to women, especially for VLDL and TG response. Similar to the response demonstrated by gender, the pattern of change was qualitatively similar for normo- and hyper-triglyceridemic individuals; however, the magnitude was more exaggerated among hyper-triglyceridemic than normo-triglyceridemic individuals.
Specifically, in terms of lipoprotein particles for both normo- or hyper-triglyceridemic individuals, we observed little change in total particle number over the 3 time points (up to 6 hours after the meal) for either total LDL or HDL particle number. In contrast, for normo-triglyceridemic subjects, total VLDL particle number appeared to rise above baseline at 3.5 hours and drop considerably below baseline at 6 hours after the high-fat meal. This pattern was considerably different in hyper-triglyceridemic individuals where, in both men and women, we observed a precipitous drop relative to baseline which continued from the 3.5 to the 6 hour time point.
Despite expected baseline differences between normo- and hyper-triglyceridemic individuals, there was a reduction in small LDL particle concentrations secondary to the PPL challenge. In normo-triglyceridemic individuals there was a persistent decline from baseline, while in hyper-triglyceridemic men and women small LDL particle concentration appeared to plateau.
The major pattern we observed among HDL particle subclasses was a significant shift to more medium HDL particles and fewer small HDL particles with a non-significant increase in HDL-C (p > 0.05 for men and women). Decewicz et al [9] observed decreases in medium HDL particles in a 73 case - 73 control study of a 1-year dietary lifestyle intervention; this contrasts to the observed increase in medium HDL observed among the 546 women and 502 men studied in the GOLDN cohort after an acute high-fat meal. It is likely that sample size and diet (Ornish diet low in fat [9] vs high fat (GOLDN)) are at the root of these differences in response. In a study examining the difference in particle distribution between diabetics and non-diabetics by Colhoun et al [10], diabetics had a lower concentration of small HDL and a larger concentration of large HDL and larger HDL sizes [10], a finding similar to ours; however, the Colhoun et al study was a descriptive study of fasting particle distributions and did not include a fat-loading intervention.
In our study, the total number of VLDL particles decreased during the postprandial period which was due to an increase in large VLDL and chylomicrons along with more substantial decreases in medium and small VLDL subclasses. A decrease in medium VLDL and total VLDL particles was also observed among the diabetic subjects in the study by Colhoun et al [10]. Our study contained a small proportion of diabetics, thus rendering further stratification by diabetic status unfeasible. Furthermore, decreases in small/medium VLDL particles were demonstrated among women during the therapeutic lifestyle change diet by Li et al [11]. The distributional shift in VLDL particle subclasses was consistent and somewhat expected since large VLDL and chylomicrons accept triglycerides in an effort to quickly repackage and remove them from the circulation [7]; however, the small reduction in total VLDL particles is unexpected, especially since VLDL-C increased in response to the acute high-fat meal.
During the PPL response to a high-fat meal, many changes occur during lipoprotein metabolism. It has been shown that HDL becomes enriched with TG, leading to increased removal of HDL from the plasma, thereby decreasing HDL-C [17]. Also the transfer of cholesterol ester (CE), the main component of HDL, is increased due to the increased activity of cholesteryl ester transfer protein (CETP) during PPL [18], and CE is preferably transferred to chylomicrons [17], all of which are demonstrated in our study by a postprandial decrease in HDL-C and increase in chylomicrons. Similarly CE from LDL is also transferred to chylomicrons during PPL [18]. Hepatic lipase (HL) activity also increases during PPL, thus catabolizing both HDL and LDL into smaller, denser HDL and LDL particles [17, 19]; however, increases in smaller, denser HDL and LDL were not clearly demonstrated in our population after a high-fat meal. Since it is generally believed that smaller LDL particles are more atherogenic [20], further characterization and understanding of the lipoprotein fluctuations that occur due to an acute high-fat meal may enhance our understanding of the relationship of lipoprotein particle subclasses to the increased risk for CVD.
This study differs from the majority of previous studies examining dietary influences on PPL that were conducted in smaller study populations, utilized different dietary interventions, and did not report changes in LDL particle subclasses [11, 12]. For example, Li et al [11] studied 33 subjects and showed that therapeutic lifestyle changes focused on diet decreased IDL among men, but not women. This is opposite to what we observed, and this is likely due to differences in the dietary intervention of Li et al [11] who utilized a diet low in total fat, saturated fat, and cholesterol, compared to the GOLDN intervention which was high in total fat and cholesterol. Additionally, random variation could explain differences since the Li et al [11] study evaluated 33 subjects compared to the 1048 reported from GOLDN.
The strengths of this study are its large sample size, the use of a standardized acute fat load, and the use of NMR technology to characterize particle size subclasses. To our knowledge, it is the largest PPL study conducted to date. Our study did not measure serum apolipoprotein B and is not a CVD outcome or mechanistic study; therefore, we do not know for certain how or why these lipoprotein particle distributional shifts occur or how they might impact CVD risk. Nonetheless, among hyper-triglyceridemic individuals, the prolonged return to baseline levels may add to an already elevated atherogenic profile [18].
Additionally, we did not obtain information on the activity level of enzymes and proteins that are known to be activated during the PPL response, such as CETP and HL [18]. Specific knowledge of how these enzymes and proteins respond during an acute high-fat meal challenge would greatly enhance the understanding of our findings as well as further the understanding of lipoprotein metabolism. We only examined the acute response to 1 high-fat meal, but since humans are in a non-fasting postprandial state for a majority of the day [21], the changes that we demonstrated may actually never return to baseline except during the latter phases of sleep. Thus, it may be the length of time that these changes in lipoprotein particle fluctuations exist that is important in atherogenesis. Another possible mechanism for atherogenesis of the particle subclasses is the fluctuations in the TG:CE content of the particles [18]. Since each lipoprotein particle subclass is composed of varying proportions of TG and CE, it may be that the changes in those proportions are atherogenic. Lastly, these findings may not extend to other racial groups, as our study population was all white.
Conclusions
Postprandial lipemia persists for at least 6 hours following a high-fat meal. We confirmed that men differ from women in the PPL response, and that the PPL response varies based on baseline triglyceride profile. We demonstrated an increase in IDL particles, especially in normo-triglyceridemic women, and an increase in triglycerides in both men and women. Large HDL also changed in hyper-triglyceridemic women (17% decrease was observed), which is interesting in view of the clinically observed greater risk of CVD in hyper-triglyceridemic women compared to men [22]. Overall, VLDL particles decreased but triglyceride-rich large VLDL and chylomicrons (with their propensity to generate atherogenic remnants) increased dramatically; however, the actual magnitude of change in total VLDL particles was relatively small. As our results demonstrate, the lipoprotein fluctuations are distinct, and some are not intuitive based on our existing knowledge of lipids and lipoprotein metabolism. Finally, our findings provide further mechanistic insight into the changes in lipoprotein metabolism occurring after an acute high-fat meal.
Methods
Study population
The Genetics of Lipid-Lowering Drugs and Diet Network (GOLDN) study population consisted of 189 families who were recruited from 3-generational pedigrees from the NHLBI Family Heart Study Minneapolis, MN, and Salt Lake City, UT, field centers. The overarching intent of GOLDN was to identify loci contributing to phenotypic variation in response to a high-fat meal and fenofibrate therapy. The Institutional Review Boards at the University of Alabama, the University of Minnesota, the University of Utah, and Tufts University approved the study protocol.
Inclusion and exclusion criteria
Participants had to meet the following criteria: White (other racial groups were excluded by design since the recruitment sources were extant cohorts of white families from Utah and Minnesota), male or female, ≥ 18 years of age, fasting triglycerides (TGs) < 1500 mg/dL, willingness to participate in the study and attend the scheduled clinic exams, part of a family with at least 2 members in a sibship, AST and ALT tests within normal range, and creatinine ≤ 2.0. Exclusion criteria for the fenofibrate intervention included the following: history of liver, kidney, pancreas, or gall bladder disease or malabsorption; pregnancy or women of childbearing potential not using an acceptable form of contraception; insulin use; known hypersensitivity to fenofibrate; pancreatitis within 12 months prior to enrollment; and current use of warfarin and/or nutraceuticals. Subjects using hypolipidemic drugs were asked to consult with their physicians to determine whether they could be taken off medications for 4 weeks prior to and for the duration of the study; informed consent for drug withdrawal was obtained. Only subjects who remained untreated with any prescription or over-the-counter hypolipidemic medication were eligible.
Study design
Participants underwent 2 interventions: an acute fat loading dietary challenge and a short-term (3 weeks) open-label intervention trial of fenofibrate. The specific methodology of GOLDN is reported elsewhere [23, 24]. For this analysis, we report LDL, HDL, and VLDL particle subclass concentrations, size and number in response to an acute standardized high fat challenge.
The high-fat challenge followed the protocol of Patsch et al [25]. The caloric intake of the intervention meal was determined by body surface area, containing 700 kilocalories per m2 of body surface area (2.93 MJ/m2 body surface area). The meal composition was 83% of calories from fat, 14% from carbohydrates, and 3% from protein. The meal was formulated to have a cholesterol content of 240 mg and a polyunsaturated:saturated fat ratio of 0.06. Based on these guidelines, the average individual ingested 175 mL of heavy whipping cream (39.5% fat) combined with 7.5 mL powdered, instant, non-fat, dry milk, and blended with ice. To increase palatability of the drink, 15 mL of chocolate- or strawberry-flavored syrup was also added. Participants had 15 minutes to ingest this meal and were required to fast a minimum of 8 hours prior to the meal. Immediately before ingestion, we drew blood samples on all participants (0 hr, fasting), and then again at 3.5 and 6 hours after the high-fat meal.
Biochemical assays
Serum and EDTA-anticoagulant tubes were collected and processed using a standardized protocol, aliquoted, and stored at -70°C until time of use. Analysis was completed on all stored samples at the end of the study and all samples for an individual were processed in the same batch to reduce measurement error. Triglyceride measurements were calculated using the glycerol-blanked enzymatic method on the Roche COBAS FARA centrifugal analyzer (Roche Diagnostics Corporation). Hyper-triglyceridemia was defined as a baseline triglyceride ≥ 150 mg/dL. HDL-C was calculated using the same procedure as TG measurement after precipitation of non-HDL cholesterol with magnesium/dextran. LDL-C measurement employed a homogeneous direct method (LDL Direct Liquid Select™ Cholesterol Reagent; Equal Diagnostics) on a Hitachi 911 Automatic Analyzer.
Lipoprotein subclass analysis
Proton nuclear magnetic resonance (NMR) spectroscopy was used to determine LDL, HDL, and VLDL particle size subclass concentrations (Liposcience, Raleigh, NC). Measured amplitudes after exposure to a 400 MHz magnet and known distinct lipid methyl group NMR spectroscopic profiles were used to determine the subclass concentrations [8, 26–30]. Concentrations of LDL and VLDL subclasses are reported as nanomoles per liter and HDL is reported as mmol/L. The different lipoprotein particles correspond to the following average size in nanometers (nm): intermediate density lipoprotein (IDL), 23-27 nm; large LDL, 21.2-23 nm; small LDL, 18-21.2 nm; large HDL, 8.8-13 nm; medium HDL, 8.2-8.8 nm; small HDL, 7.3-8.2 nm; large VLDL, 60-200 nm; medium VLDL, 35-60 nm; and small VLDL, 27-35 nm [8, 11, 31]; relative size is depicted in Figure 6.

Nuclear magnetic resonance-derived low-density lipoprotein (LDL), high-density lipoprotein (HDL), and very low-density lipoprotein (VLDL) particle size subclasses.
Statistical methods
All analyses were conducted separately for men and women and stratified by baseline hyper-versus normo-triglyceride levels (TG > 150 vs TG ≤ 150) due to significant interactions between gender and baseline triglyceride concentrations in analyses for LDL, HDL, and VLDL. To determine the significance of differences in percentages between male and female normo- and hyper-triglyceridemic individuals, we used the Pearson c2 and Fisher exact tests. To compare crude means, we used ANOVA and Student's t-test. Results are expressed as least squares mean (standard error). We examined changes in subclass particle size concentrations that occurred during the dietary fat challenge (baseline, 3.5 hours, and 6 hours) using repeated-measures ANOVA in which the family was modeled as a random variable. All statistical calculations were performed using SAS 9.1 (SAS Institute, Inc.).
Abbreviations
- ALT:
-
alanine transaminase
- AST:
-
aspartate transaminase
- CETP:
-
cholesteryl ester transfer protein
- CVD:
-
cardiovascular disease
- GOLDN:
-
Genetics of Lipid-Lowering Drugs and Diet Network Study
- HDL-C:
-
high-density lipoprotein cholesterol
- HL:
-
hepatic lipase
- IDL-C:
-
intermediate-density lipoprotein
- LDL-C:
-
low-density lipoprotein cholesterol
- NMR:
-
nuclear magnetic resonance (spectroscopy)
- PPL:
-
post-prandial lipemia
- TG:
-
triglyceride
- VLDL-C:
-
very low-density lipoprotein cholesterol
References
, : Executive Summary of The Third Report of The National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, And Treatment of High Blood Cholesterol In Adults (Adult Treatment Panel III). JAMA. 2001, 285: 2486-2497. 10.1001/jama.285.19.2486
Wilson PW, D'Agostino RB, Levy D, Belanger AM, Silbershatz H, Kannel WB: Prediction of coronary heart disease using risk factor categories. Circulation. 1998, 97: 1837-1847.
Roche HM, Gibney MJ: The impact of postprandial lipemia in accelerating atherothrombosis. J Cardiovasc Risk. 2000, 7: 317-324.
Cromwell WC, Otvos JD: Low-density lipoprotein particle number and risk for cardiovascular disease. Curr Atheroscler Rep. 2004, 6: 381-387. 10.1007/s11883-004-0050-5
Otvos JD, Jeyarajah EJ, Cromwell WC: Measurement issues related to lipoprotein heterogeneity. Am J Cardiol. 2002, 90: 22i-29i.
Mudd JO, Borlaug BA, Johnston PV, Kral BG, Rouf R, Blumenthal RS, Kwiterovich PO: Beyond low-density lipoprotein cholesterol: defining the role of low-density lipoprotein heterogeneity in coronary artery disease. J Am Coll Cardiol. 2007, 50: 1735-1741. 10.1016/j.jacc.2007.07.045
Karpe F: Postprandial lipoprotein metabolism and atherosclerosis. J Intern Med. 1999, 246: 341-355. 10.1046/j.1365-2796.1999.00548.x
Otvos JD: Measurement of lipoprotein subclass profiles by nuclear magnetic resonance spectroscopy. Clin Lab. 2002, 48: 171-180.
Decewicz DJ, Neatrour DM, Burke A, Haberkorn MJ, Patney HL, Vernalis MN, Ellsworth DL: Effects of cardiovascular lifestyle change on lipoprotein subclass profiles defined by nuclear magnetic resonance spectroscopy. Lipids Health Dis. 2009, 8: 26- 10.1186/1476-511X-8-26
Colhoun HM, Otvos JD, Rubens MB, Taskinen MR, Underwood SR, Fuller JH: Lipoprotein subclasses and particle sizes and their relationship with coronary artery calcification in men and women with and without type 1 diabetes. Diabetes. 2002, 51: 1949-1956. 10.2337/diabetes.51.6.1949
Li Z, Otvos JD, Lamon-Fava S, Carrasco WV, Lichtenstein AH, McNamara JR, Ordovas JM, Schaefer EJ: Men and women differ in lipoprotein response to dietary saturated fat and cholesterol restriction. J Nutr. 2003, 133: 3428-3433.
James RW, Pometta D: Postprandial lipemia differentially influences high density lipoprotein subpopulations LpAI and LpAI, AII. J Lipid Res. 1994, 35: 1583-1591.
Chen YD, Swami S, Skowronski R, Coulston A, Reaven GM: Differences in postprandial lipemia between patients with normal glucose tolerance and noninsulin-dependent diabetes mellitus. J Clin Endocrinol Metab. 1993, 76: 172-177. 10.1210/jc.76.1.172
Groot PH, van Stiphout WA, Krauss XH, Jansen H, van Tol A, van Ramshorst E, Chin-On S, Hofman A, Cresswell SR, Havekes L: Postprandial lipoprotein metabolism in normolipidemic men with and without coronary artery disease. Arterioscler Thromb. 1991, 11: 653-662. 10.1161/01.ATV.11.3.653
Syvanne M, Talmud PJ, Humphries SE, Fisher RM, Rosseneu M, Hilden H, Taskinen MR: Determinants of postprandial lipemia in men with coronary artery disease and low levels of HDL cholesterol. J Lipid Res. 1997, 38: 1463-1472.
Couillard C, Bergeron N, Prud'homme D, Bergeron J, Tremblay A, Bouchard C, Mauriege P, Despres JP: Gender difference in postprandial lipemia: importance of visceral adipose tissue accumulation. Arterioscler Thromb Vasc Biol. 1999, 19: 2448-2455. 10.1161/01.ATV.19.10.2448
Cohn JS: Postprandial lipemia and remnant lipoproteins. Clin Lab Med. 2006, 26: 773-786. 10.1016/j.cll.2006.07.003
Chung BH, Liang P, Doran S, Cho BH, Franklin F: Postprandial chylomicrons: potent vehicles for transporting cholesterol from endogenous LDL+HDL and cell membranes to the liver via LCAT and CETP. J Lipid Res. 2004, 45: 1242-1255. 10.1194/jlr.M300350-JLR200
Berneis KK, Krauss RM: Metabolic origins and clinical significance of LDL heterogeneity. J Lipid Res. 2002, 43: 1363-1379. 10.1194/jlr.R200004-JLR200
Cromwell WC, Otvos JD, Keyes MJ, Pencina MJ, Sullivan L, Vasan RS, Wilson PW, D'Agostino RB: LDL Particle Number and Risk of Future Cardiovascular Disease in the Framingham Offspring Study - Implications for LDL Management. J Clin Lipidol. 2007, 1: 583-592. 10.1016/j.jacl.2007.10.001
Lopez-Miranda J, Williams C, Lairon D: Dietary, physiological, genetic and pathological influences on postprandial lipid metabolism. Br J Nutr. 2007, 98: 458-473. 10.1017/S000711450774268X
Hokanson JE, Austin MA: Plasma triglyceride level is a risk factor for cardiovascular disease independent of high-density lipoprotein cholesterol level: a meta-analysis of population-based prospective studies. J Cardiovasc Risk. 1996, 3: 213-219. 10.1097/00043798-199604000-00014
Corella D, Arnett DK, Tsai MY, Kabagambe EK, Peacock JM, Hixson JE, Straka RJ, Province M, Lai CQ, Parnell LD, Borecki I, Ordovas JM: The -256T > C polymorphism in the apolipoprotein A-II gene promoter is associated with body mass index and food intake in the genetics of lipid lowering drugs and diet network study. Clin Chem. 2007, 53: 1144-1152. 10.1373/clinchem.2006.084863
Lai CQ, Arnett DK, Corella D, Straka RJ, Tsai MY, Peacock JM, Adiconis X, Parnell LD, Hixson JE, Province MA, Ordovas JM: Fenofibrate effect on triglyceride and postprandial response of apolipoprotein A5 variants: the GOLDN study. Arterioscler Thromb Vasc Biol. 2007, 27: 1417-1425. 10.1161/ATVBAHA.107.140103
Patsch JR, Miesenbock G, Hopferwieser T, Muhlberger V, Knapp E, Dunn JK, Gotto AM, Patsch W: Relation of triglyceride metabolism and coronary artery disease. Studies in the postprandial state. Arterioscler Thromb. 1992, 12: 1336-1345. 10.1161/01.ATV.12.11.1336
Ordovas JM, Cupples LA, Corella D, Otvos JD, Osgood D, Martinez A, Lahoz C, Coltell O, Wilson PW, Schaefer EJ: Association of cholesteryl ester transfer protein-TaqIB polymorphism with variations in lipoprotein subclasses and coronary heart disease risk: the Framingham study. Arterioscler Thromb Vasc Biol. 2000, 20: 1323-1329. 10.1161/01.ATV.20.5.1323
Tsai MY, Georgopoulos A, Otvos JD, Ordovas JM, Hanson NQ, Peacock JM, Arnett DK: Comparison of ultracentrifugation and nuclear magnetic resonance spectroscopy in the quantification of triglyceride-rich lipoproteins after an oral fat load. Clin Chem. 2004, 50: 1201-1204. 10.1373/clinchem.2004.032938
Otvos JD, Jeyarajah EJ, Bennett DW, Krauss RM: Development of a proton nuclear magnetic resonance spectroscopic method for determining plasma lipoprotein concentrations and subspecies distributions from a single, rapid measurement. Clin Chem. 1992, 38: 1632-1638.
Staels B, Vu-Dac N, Kosykh VA, Saladin R, Fruchart JC, Dallongeville J, Auwerx J: Fibrates downregulate apolipoprotein C-III expression independent of induction of peroxisomal acyl coenzyme A oxidase. A potential mechanism for the hypolipidemic action of fibrates. J Clin Invest. 1995, 95: 705-712. 10.1172/JCI117717
Staels B, Dallongeville J, Auwerx J, Schoonjans K, Leitersdorf E, Fruchart JC: Mechanism of action of fibrates on lipid and lipoprotein metabolism. Circulation. 1998, 98: 2088-2093.
Ikewaki K, Tohyama J, Nakata Y, Wakikawa T, Kido T, Mochizuki S: Fenofibrate effectively reduces remnants, and small dense LDL, and increases HDL particle number in hypertriglyceridemic men - a nuclear magnetic resonance study. J Atheroscler Thromb. 2004, 11: 278-285. 10.5551/jat.11.278
Acknowledgements
We thank the GOLDN families for their participation in this research.
This study was supported by NIH Heart, Lung, and Blood Institute grant U01 HL72524, Genetic and Environmental Determinants of Triglycerides. Dr. Mary K. Wojczynski was supported by the UAB Statistical Genetics Post-Doctoral Training Program, T32HL07275057.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
MKW conducted the statistical analyses and drafted the manuscript. AO and EKK provided critical input within their respective areas of expertise during analysis, drafting, review, and revision. SPG, PNH, MYT, RJS, and JMO were critical participants in study design and provided input within their respective areas of expertise during analysis, drafting, review, and revision. DKA was the primary designer of the study and oversaw and contributed to all stages of analysis and authoring. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is published under license to BioMed Central Ltd. This is an Open Access article is distributed under the terms of the Creative Commons Attribution License ( https://creativecommons.org/licenses/by/2.0 ), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Wojczynski, M.K., Glasser, S.P., Oberman, A. et al. High-fat meal effect on LDL, HDL, and VLDL particle size and number in the Genetics of Lipid-Lowering drugs and diet network (GOLDN): an interventional study. Lipids Health Dis 10, 181 (2011). https://doi.org/10.1186/1476-511X-10-181
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/1476-511X-10-181