
Abstract
The influence of lignite mining on vegetation constitutes a key issue due to the role of plants in restoring and maintaining the ecological balance of ecosystems. In this context, the identification of its impact on the functioning of silver birch (Betula pendula Roth) as a species often colonizing disturbed habitats takes on particular importance. Therefore, we aimed to determine the changes in nutrient content in silver birch overgrowing a spoil heap and in the vicinity of a fly ash settling pond and power plant. For this purpose, plant tissues (fine and coarse roots, stemwood, bark, coarse and fine branches, leaves) and soil samples (0–10, 10–20, 20–40, 20–40 cm deep) were examined. The basic soil characteristics were determined, along with the N, P, K, Ca, Mg, S, Fe, Mn, Cu, and Zn contents of the soil and plant samples. The soils varied in terms of soil pH and were poor in total organic carbon and other elements. The plant nutrient content varied strongly across the analyzed tissues, with the leaves usually containing the most and the stemwood the least nutrients. Statistical analysis indicated significant differences between the control vs spoil heap (particularly in Mn, S, and Mg) and the stand close to the settling pond (particularly in Ca, Mn, P, K, and S). We found that the chemical properties of the spoil heap and fly ash originating from the lignite mining operations are likely factors influencing nutrient accumulation in silver birch trees.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Lignite (brown coal) is an important fuel in the production of electricity and heat in many European countries. In Poland, it is extracted from several large, opencast mines near Bełchatów, Konin, Adamów, Turów, and Sieniawa. The amount of lignite mined in Poland in 2017 was 61 million tons, equaling approximately one-third of Germany’s production. Up to 2021, there had been expected reduction in lignite mining over the next years. However, considering the present political and economic situation in the world, this is no longer so reliable.
Lignite mining and processing constitutes one of the most important ecological and economic problems of the last several decades. Opencast mines cover large areas of strongly degraded environment, including the lithosphere, soils, hydrology, vegetation, and landscapes. In addition, contaminants can be dispersed over large distances. Among such pollutants, lignite dust is typical of mining activity, whereas ash and several other products originate from lignite burning (Kosztowniak et al., 2016; Pandey et al., 2014; Vrablik et al., 2017). These substances are carriers of nutrients, along with potentially toxic elements and various organic and inorganic contaminants. Acid mine drainage is another inherent phenomenon of areas affected by lignite mining (Fyson et al., 2006), as it affects the quality of water reservoirs. For instance, in the Lausitz area (Germany), approximately 170 lignite pit lakes have a pH range of 2.4 to 3.4 (Fyson et al., 2006). The water in the pits is also characterized by high concentrations of iron (Fe) (up to 800 mg dm–3), which occurs as sulfides (mostly pyrite and marcasite) as accessory minerals in the lignite. It should be noted that sulfides can also be a source of trace elements.
The influence of lignite dust and coarse fragments on the soil environment and the plant–soil system is multifaceted and ambiguous. Several studies have demonstrated that it can have a positive effect on the soil organic carbon content and pore structure, and can slow mineralization (Kołodziej et al., 2020; Sekhohola et al., 2013). Some authors (e.g., Kwiatkowska-Malina, 2015) have also tested the ability of humic acids extracted from lignite to improve soil quality. However, there is also some evidence that lignite used as a soil amendment negatively influences soil system. Frouz et al. (2005) showed that some lignite mine spoils inhibit the reproduction of pot worms (Enchytraeus crypticus), mainly due to their low pH (< 4) and/or high salinity. Simmler et al. (2013) found that high doses of lignite introduced to soil reduced the growth of ryegrass. Only a few studies have monitored the effects of lignite on soil microbial processes. Several authors (Bekele et al., 2015; Rumpel et al., 2001; Tran et al., 2015) have suggested that lignite can be utilized by soil microorganisms as a carbon (C) source, but the efficiency of this process is limited. Qin and Leskovar (2018) proved that lignite‐derived humic substances promote plant root growth and soil microbiota populations, which are essential for improving plant–microbial interactions under water-deficit stress. Baumann et al. (2005) showed that lignite can promote the relative abundance of arbuscular mycorrhizal fungi (AMF), which form symbiotic root associations with plants. This may be because, among rhizosphere microorganisms, the response of fungi to increasing C content is clearer than that of bacteria because fungal mycelia can accelerate the turnover cycle of the fungal C metabolism, while bacteria generally need much more time to turn over the C (Staddon et al., 2003).
Former lignite mining sites are reclaimed using various techniques. Areas of excavation and spoil heaps are usually leveled and afforested. Scots pine (Pinus sylvestris L.) and silver birch (Betula pendula Roth) have been commonly used for this purpose due to their high tolerance of such site conditions (Warfvinge & Svedrup, 1995), including aluminum (Al) stress (Clegg & Gobran, 1995) and contamination (Eltrop et al., 1991; Marguí et al., 2007; Ulbrichová et al., 2005). Knowledge on the feedbacks between soils and vegetation is crucial for the successful reclamation of degraded areas. Aspects related to nutrient cycling, including their uptake, bioaccumulation in fresh biomass, return to the soil via litterfall, and release during litter decomposition, are key importance. Rapid nutrient cycling is particularly important in nutrient-poor sites, which are often represented by degraded post-mining areas. The choice of silver birch is more suitable in this context than Scots pine. This is mainly due to the characteristics of the litterfall from these trees. Scots pine litter results in a strongly acidified, nutrient-poor biomass saturated with resins, which makes it resistant to decomposition (Jonczak, 2011). By contrast, silver birch litterfall is rich in essential nutrients and microelements (Brandtberg et al., 2004; Carnol & Bazgir, 2013; Jonczak et al., 2023), and the process of its decomposition is rapid (Hynynen et al., 2010; Shorohova & Kapitsa, 2016). The choice of tree species in the reclamation of degraded soils has another important aspect––phytoremediation. Silver birch is considered a hyperaccumulator of zinc (Zn) (Dmuchowski et al., 2012; Marguí et al., 2007). The latest studies by Rustowska (2022, 2024) and Jonczak et al. (2023) have also shown a strong accumulation of manganese (Mn) and copper (Cu). Their highest concentrations are typically recorded in the leaves (Jonczak et al., 2023; Rustowska, 2022).
Considering the importance of silver birch in the reclamation of areas degraded by lignite mining, and the insufficiency of our knowledge on the feedbacks in the soil–silver birch system in such areas, we undertook a broad study on nutrient accumulation in the biomass of that tree. The study covered three stands, including a spoil heap near a mine pit, a stand near a power plant dust settling pond, and a control stand. We expected large differences in the contents of the studied nutrients in the silver birch biomass related to the location and site characteristics. Moreover, we hoped that this study would contribute to a better understanding of the ecology of this species under unfavorable conditions, and its suitability for the reclamation of soil in degraded areas.
2 Material and Methods
2.1 Study Area
The first geophysical exploration for lignite, near Bełchatów (central Poland), began during World War II (Ratajczak & Hycnar, 2017). Continued exploration resulted in lignite being found at a depth of 127 m below ground level in 1960. The thickness of the lignite deposit was estimated at 70 m. In 1975, after intense exploration works, the Presidium of the Government decided to start lignite operations and to build a thermal power plant. The lignite deposits near Bełchatów are of the tectonic type (Ratajczak & Hycnar, 2017). They occur within Mesozoic formations in the Łódź Basin (with a W–E orientation). The length and width of the deposits was estimated at 6.5 and 2.5 km, respectively. Based on the geology, the lignite deposits have been divided into three areas––Szczerców, Bełchatów, and Kamieńsk. The first two are currently being exploited. The boundary between the Szczerców and Bełchatów areas is represented by the Dębina salt dome, whereas the Kamieńsk and Bełchatów areas are divided by the Widawka Fault. In general, the lithostratigraphic profile in the study area allows the distinction of three major parts: (a) Quaternary sediments; (b) a Cenozoic lignite series; and (c) a Mesozoic base. The Quaternary sediments (35–80 m thick) are typically represented by sands, clays, gravels, fluvioglacial sands, and organic sediments. The parent rocks identified in the Mesozoic base are mostly limestones. In the Cenozoic lignite series, lignite, gray fine-grained sands, clays, lake chalks, sandstones, and marls have been identified (Wagner & Słomka, 2000). In the Bełchatów mine, the lignite is exploited by means of multi-bucket, wheeled excavators, the material transported using a belt conveyor (Borcz & Kozioł, 2015). Various technologies are used to take out the overburden and parent rocks, which are relatively resistant to crushing (Kozioł et al., 2011).
Our study was performed on three silver birch stands, including one on a reclaimed spoil heap formed from the overburden material, one in the vicinity of a fly ash settling pond, and one located approximately 8 km NE of the spoil heap and lignite opencast mine, which acted as the control (Fig. 1). Although, the stands were comparable in age (30–34 years), they varied in terms of density and diameter (Table 1).

Location of studied stands
2.2 Soil and Biomass Sampling and Analysis
Silver birch trees and the soils were sampled once in June 2022. Biomass samples were collected from 10 average trees per stand, and included second-order roots, first-order roots, stemwood, and bark at a height ≈130 cm, and first-order branches, second-order branches, and leaves from the central part of the crown. The biomass samples were dried at 65 °C, then milled into powder, and analyzed. The total organic C (TOC), total nitrogen (N) and total sulfur (S) were determined by dry combustion (Vario MacroCube, Elementar, Germany), while the contents of phosphorus (P), potassium (K), calcium (Ca), magnesium (Mg), Fe, Mn, Cu, and Zn were obtained by inductively coupled–plasma atomic-emission spectrometry (Avio 200, Perkin Elmer, USA) after sample digestion in 65% nitric acid (HNO3) using a microwave digestion oven (Ethos Up, Milestone, Italy).
The soil samples were collected from close to each tree from depths of 0–10, 10–20, 20–40, and 40–80 cm using a 3 cm–diameter corer. One core was taken from the central part of the stand for the purposes of describing and classifying the soil. The soil samples were air-dried and passed through a 2.0 mm–mesh sieve. The < 2.0-mm fraction was used for the physical properties analyses. Additionally, part of the homogenized sample was milled into powder for chemical analysis. The particle size distribution was determined using the mixed pipette and sieve method. The Polish Soil Science Society (PTG) classification of textural fractions and groups was applied (PTG, 2009). The soil pH was determined using the potentiometric method (Mettler Toledo SevenDirect SD23 equipped with an InLab ExpertPro ISM electrode, Switzerland) in a soil:water proportion of 1:10. The total C (TC), N, and S contents were determined by dry combustion (Vario MacroCube, Elementar, Germany). The total inorganic C (TIC) was analyzed using the volumetric Scheibler method, and the TOC was calculated as TC – TIC. Approximately 0.3 g of soil sample was digested in 7.5 ml of 38% hydrochloric acid (HCl) and 2.5 ml of 65% HNO3. The solution was transferred into a 50-ml falcon tube, topped up with deionized water to 50 ml, and then filtered (through a hard paper filter). The contents of P, K, Ca, Mg, Fe, Mn, Cu, and Zn were determined by inductively coupled–plasma optical emission spectroscopy. Only high-quality, pure per-analysis reagents were used for biomass and soil sample digestion.
2.3 Statistical Analysis
Statistical evaluation of the results included obtaining the mean values and standard deviations for the biomass and soil samples. The statistical significances of the differences between the mean values were tested using the Kruskal–Wallis test. Principal component analysis (PCA) was applied to identify major sources of variability in the studied datasets.
3 Results
3.1 Basic Characteristics of the Soils
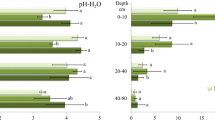
The studied soils represented three reference groups, according to the WRB 2022 (International Union of Soil Sciences, 2022) classification, that developed under the various impacts of natural and anthropogenic factors. These were a semi-natural Dystric Brunic Arenosol, representing the control location, an Albic Podzol (Arenic, Eutric), located in the vicinity of the settling pond, characterized by a natural or close-to-natural sequence of genetic horizons, albeit threatened by contamination from the settling pond, and a Spolic Technosol (Arenic, Eutric, Ochric), developed from the overburden material that comprised the spoil heap. The soils were characterized by a sandy texture, with the content of the sand fraction ranging from 91.5 to 99.6% (Table 2). The pH values varied among the stands and with depth (Fig. 2), being strongly acidic or acidic at the settling pond and control locations, showing low variability with depth. The observed pH values are typical of the sandy soils of Poland under forest vegetation. The soil of the spoil heap was acidic at the top, but with the pH strongly increasing with depth. This tendency indicates the leaching of basic cations and carbonates from the surface layers of anthropogenic materials. This process is typical under percolative water regimes, which can be additionally accelerated by forest vegetation. The studied soils were generally poor in TOC, despite the observed differences among the stands (Fig. 2).

pH and TOC content in the studied soils (the same letters indicate no statistically significant differences between the stands at a significance level of p < 0.05 based on the Kruskal–Wallis test)
3.2 Soil Nutrient Content
Although having comparable textural characteristics, soil nutrient content was significantly different among the three sites (Table 3, Fig. 3). On the heap stand, soils were deficient in N and P, while being much more abundant in K, Ca, and Mg when compared to the settler and control locations. The differences were also apparent in the micronutrients. Generally, the contents of S, Mn, Zn, and Cu were low (close to natural), considering the proximity of the sources of pollution (opencast mine, settler, power plant).

Mean ± SD soil nutrient content at depths of 0–10, 10–20, 20–40, and 40–80 cm (the same letters indicate no statistically significant differences between the stands at a significance level of p < 0.05 based on the Kruskal–Wallis test)
3.3 Nutrient Content in Silver Birch Biomass
The nutrient distribution in the silver birch biomass varied strongly among the tissues and stands (Fig. 4). The highest concentrations were mainly noted in the leaves and the lowest in the stemwood. The only exceptions to this tendency were Fe and Mn. The trends for these nutrients were not fully clear, however large differences were noted between the stands. Also, relatively high contents of Ca, Cu, and Zn were recorded in the bark, branches, and roots. In the roots, there were also high concentrations of Fe. Based on the mean values of the tissues, the N contents were the highest among all the major nutrients in the silver birch biomass (2,337.5–24,786.3 mg kg–1), followed by Ca (720.2–16,418.4 mg kg–1), K (325.1–7,892.0 mg kg–1), Mg (164.3–2,899.2 mg kg–1), and P (68.9–1,651.1 mg kg–1). Micronutrients occurred in amounts of 7.3–2,341.3 mg kg–1 Mn, 54.7–1,246.5 mg kg–1 S, 10.8–1,114.4 mg kg–1 Fe, 18.0–179.3 mg kg–1 Zn, and 2.8–9.7 mg kg–1 Cu.

Mean ± SD nutrients content in birch biomass (the same letters indicate no statistically significant differences between the stands at a significance level of p < 0.05 based on the Kruskal–Wallis test). RII––second-order roots, RI––first-order roots, S––stemwood, B––bark at a height ≈130 cm, BrI––first-order branches, BrII––second-order branches, and L––leaves
3.4 Principal Component Analysis (PCA)
A PCA analysis, based on the major soil characteristics and nutrient content of the soils and silver birch biomass, allowed us to identify the major sources of variability in, and the interrelationships between, the studied variables in a silver birch–soil system (Fig. 5). An analysis of the contributions of the variables in the model revealed no dominant role by any of them, highlighting the complicated nature of the studied system. However, the correlation matrix of the variables indicated strong feedbacks between the soil characteristics and nutrient contents in the silver birch biomass. These were both positive and negative correlations. Based on the projection of the cases on the plane of factors, separate clusters for each study plot are clearly visible, suggesting differences between them, considering the variables used for the analysis (Fig. 6).

PCA analyses of individual silver birch tissues and soil characteristics. RII––second-order roots, RI––first-order roots, S––stemwood, B––bark at a height ≈130 cm, BrI––first-order branches, BrII––second-order branches, and L––leaves

PCA analyses of individual silver birch tissues (projection of cases on the plane of factors). RII––second-order roots, RI––first-order roots, S––stemwood, B––bark at a height ≈130 cm, BrI––first-order branches, BrII––second-order branches, and L––leaves
4 Discussion
Site conditions, affected by anthropopressure, are often subject to strong fluctuations, which can be reflected in the plant nutrient status. For silver birch, this has been illustrated by several authors (e.g., Daugaviete et al., 2015; Hytönen et al., 2014; Kříbek et al., 2020; Pająk et al., 2016; Sitko et al., 2022). In the present study, the influence of the lignite operations was confirmed by the PCA analysis (Fig. 6), which clearly showed different effects on the nutrient composition in the silver birch tissues from each stand. This highlights the complex character of this process, in which the nutrient availability for plants is controlled by certain factors, particularly soil pH (Barrow & Hartemink, 2023). As we found, the soil nutrient profiles of the investigated stands were not homogeneous (Fig. 2). While we expected the soils of the settling pond and spoil heap locations to be acidic, as is typical of Podzols and Arenosols (Brożek & Zwydak, 2010), we found the soil of the heap to be alkaline in the deeper layers. This contrasts with the reports by Pająk et al. (2023) and Wójcik et al. (2012), who noted a strongly acidic reaction at this site, whereas it corresponds with the observations of Pająk and Krzaklewski (2006). A varying pH in the overburden was observed by Gruszczyński (2015) and Pudełko and Chodak (2020). Such discrepancies are common, being an effect of the heterogeneity of the deposited material (Wójcik & Krzaklewski, 2009). As mentioned, soil pH determines nutrient uptake and its accumulation in the plant biomass, along with the nutrient concentration in the substrate (Cataldo & Wildung, 1978), and the plant’s requirements for a given nutrient (Clark, 1983). Considering the environmental factors, either directly or indirectly shaped by anthropization, the influence of pH on the nutritional status of plants can be highly differentiated, depending also on the type of the nutrient.
Nitrogen and P are dominant limiting nutrients in plant growth, influencing productivity and the overall functioning of terrestrial ecosystems (Perring et al., 2008), especially those associated with industrial/post-industrial sites, which are often extremely poor in biogenic elements (Pająk et al., 2023; Pietrzykowski et al., 2013), as was also noted in this study (Fig. 3). The soil N and P contents were considerably lower than in the control (Fig. 3), as well as in other nutrient-poor, sandy soils under silver birch stands (Rustowska, 2022, 2024). This explains the limited accumulation of P in the birch biomass in the settler location. The comparable biomass P contents in the heap and control locations were probably influenced by a higher soil reaction increasing its uptake. In contrast to P, the N content in the birch biomass was not reduced in any stand in the mining zone, and was generally similar to the reports by other authors for various sites (Jonczak et al., 2023; Kuznetsova et al., 2010; Rustowska, 2022, 2024; Uri et al., 2007). This might be explained by the symbiotic interaction of the birch roots with AMF facilitating N uptake from the soil (Smith & Smith, 2011), promoted by their abundance due to the lignite (Baumann et al., 2005). Moreover, an increase in N content in the aboveground plant tissues, particularly the leaves (Fig. 3), might have been caused by the emission of N oxides (NOx) from the openpit mining (Huertas et al., 2012).
In terms of K, significantly lower contents were recorded in the birch trees growing near the ash settler pond and power plant in relation to the control (Fig. 3). This is possibly due to the typically small amounts of K that occur in Podzols (Brożek & Zwydak, 2010) (in this study, the K was lower than in the Arenosol of the control stand), as well as the lignite fly ash from the Bełchatów power plant (Bąk et al., 2023; Uzarowicz et al., 2018).
The fly ash from the Bełchatów lignite mine is characterized by a high Ca content (Giergiczny & Michniewicz, 1991; Peukert et al., 1986). As reported by Hycnar et al. (2017) and Uzarowicz et al. (2018), this is due to the presence of significant amounts of CaCO3-rich lake chalk in the combusted coal (Hycnar et al., 2017). What is more, the combustion lignite rich in Ca can lead to the formation of other Ca mineral phases, such as gehlenite (Ca2Al2SiO7), ye’elimite [Ca4Al6(SO4)], anhydrite (CaSO4), and lime (CaO), which have all been found in the Bełchatów fly ash (Bąk et al., 2023). Also, the soil of the heap stand contained higher contents of Ca than the control stand, which, as reported by Pająk and Krzeklewski (2006), is due to the presence of CaCO3 in these formations. Based on the above, the increased Ca content in the birch biomass growing in the vicinity of the mine was not surprising (Fig. 3). The Ca content was considerably higher than in birches in urban areas (Modrzewska et al., 2016), and also higher than in silver birch growing on post-mining spoil heaps formed from waste rock from the coal mine in Katowice (Sitko et al., 2022). The increased accumulation recorded in this study was probably influenced by the higher pH of the overburden materials (5.8 ± 0.4–8.1 ± 0.7) compared to that recorded by Sitko et al. (2022) (3.68 ± 0.06).
The Mg contents in some of the birch tissues varied from the control only at the heap location (Fig. 3), which differed in the soil reaction (Fig. 2). As confirmed by PCA analysis (Fig. 4), the increased uptake of Mg is positively correlated with soil pH, this being a significant factor determining the availability of this nutrient to plants (Hailes et al., 1997), which explains the observed trend.
Sulfur is typically accumulated by plants from the soil in the form of sulfates, although it can be also absorbed from the air as S dioxide (SO2) (Linzon et al., 1979) and hydrogen sulfide (H2S), released during the pyrolysis of lignite (Zhang & Yani, 2011). The presence of S in these forms is particularly common in industrial areas through its emission by several activities, such as smelting, combustion, and mining (Gordon & Gorham, 1963; Katz et al., 1939; Linzon, 1958, 1965), thus contributing to the elevated contents of this nutrient in plants. Frazer (1935) highlighted that some coniferous plants growing on industrial sites exhibited significantly higher levels of S compared to their counterparts growing in non-polluted areas. Also, Katz et al. (1939) observed that the S concentrations in coniferous foliage were the highest near a lead smelter, gradually decreasing with distance from this. A similar tendency was noted in our study, with the S contents in the silver birch tissues being usually significantly higher in both the heap and settler stands than in the control (Fig. 4).
The large-scale mining and burning of coal often poses a risk of soil contamination by trace elements (Dang et al., 2002; Rout et al., 2013). However, studies conducted at the Bełchatów lignite mine site have indicated that both the combustion waste and the overburden heap do not show elevated concentrations of these (Hycnar et al., 2017; Stolecki, 2005). We can confirm these reports––the contents of Cu and Zn in the soils of the studied stands were low and, according to legal regulations (Regulation of the Minister of the Environment of 1 September 2016 on the Method of the Contamination Assessment of the Earth Surface), they did not exceed permissible norms. Therefore, the impact of lignite mining activities on the accumulation of these micronutrients by silver birch was negligible (Fig. 4), and their contents did not differ from those presented by Rustowska (2022) from Arenosol stands. Contrastingly to Cu and Zn, Mn exhibited significant differences between the heap and the settler stands vs the control stand in all tissues, showing a considerable decrease in concentration in the stands that experienced anthropogenic impacts (Fig. 4). The accumulation of Mn is primarily dependent on soil pH, which is the most effective in acidic soils (Kabata-Pendias & Pendias, 1999). Reimann et al. (2007) revealed that birches will accumulate Mn in their leaves up to 4,888 mg kg−1 in soils with low pH. This explains the higher content of Mn in the birches growing in the control stand. At the settler location, the Mn content was considerably lower than at the control stand, despite the acidic reaction of the soil. This seems to be related to the extremely low Mn contents in the soils of the settler and heap stands, which had much lower concentrations than in other soils under silver birch stands reported in the literature (Jonczak et al., 2023; Rustowska, 2022; Sitko et al., 2022). Deficiency in Mn is prevalent among plants that thrive in soils originating from parent materials with low Mn contents, as well as in soils that have undergone extensive leaching (Ivanov et al., 2022). The other explanation of the observed differences between the stands can be the impact of stand density. As presented in the work of Machava and Barna (2005), Mn content in in the foliage of dominant beech was the highest on plots with the highest density, which is in line with our study. Positive correlation of the amount of accumulated nutrients in different parts of Scots pine with stand density indicated also Węgiel et al. (2018).
According to Hycnar et al. (2017), fly ash from the Bełchatów power plant contains a number of mineral forms of Fe, including hematite (Fe2O3), magnetite (Fe3O4), wüstite (FeO), goethite (FeO(OH)), lepidocrocite (FeO(OH)), and pyrrhotite (FeS), indicating the possible capture of this nutrient from dust by birch, and an increase in the Fe content of the sites close to the settling pond and power plant. However, we found no such trend (Fig. 3), the statistical analysis showing significant differences between the heap and control stands, but the trend being unclear.
The distribution of nutrients accumulated in trees is dependent on the specific nutrient demands and functions of each organ (Martins et al., 2019; Miller, 1995; Tromp, 1983). In addition, in deciduous trees, this is controlled by the retranslocation process, which is an important strategy for preserving the loss of nutrients from senescent leaves (Jonczak et al., 2023; Scalon et al., 2017). This tissue usually contains the highest content of nutrients, followed by the branches, bark, or roots, while stemwood has the lowest (Alifragis et al., 2001; Sharma & Sharma, 2013; Vos et al., 2023). In this study, the distribution of the majority of the nutrients showed a similar pattern (see Supplementary material), which was also consistent with the findings reported for silver birch by Novák et al. (2017) and Rustowska (2022, 2024).
5 Conclusions
Based on the obtained data, our hypothesis that the site characteristics in the lignite mining zone would be reflected in the nutritional status of the silver birch stands seems to have been confirmed for the majority of nutrients. However, depending on the nutrient and the site conditions, the effect was diverse. Statistical analysis demonstrated that silver birch growing on the heap comprising overburden material deposited during lignite mining had increased contents of, particularly, S, followed by Fe and Mg, while the birches growing in the vicinity of the fly ash settling pond and power plant showed a capacity to retain mainly Ca and S in the majority of their tissues. We found lower accumulations of some nutrients, especially Mn, which was considerably decreased in both of the polluted stands compared to the control. Additionally, P and K accumulation in the zone under the impact of fly ash was reduced in relation to the undisturbed forest stand. In the case of Fe, although significant differences were present in some cases, the tendency was not clear. The impact of lignite mining operations on the accumulation of Cu and Zn was negligible. The distribution of nutrients was typical for tree species, with the leaves being the most abundant tissue (except for Fe), while the stemwood was the poorest. Overall, the results of this study, in tandem with the anthropogenically altered nature of the site, indicate the complexity of the issue of nutrient accumulation in silver birch trees growing in the investigated area, in which soil-related parameters and the nutrient composition of the fly ash seem to have play important roles. Our findings expanded knowledge in the field of silver birch ecology of the areas affected by lignite opencast mining, and they have the potential to be applied to the effective reclamation and restoration of degraded areas.
Data Availability
All data supporting the findings of this study are available within the paper and its Supplementary Information.
References
Alifragis, D., Smiris, P., Maris, F., Kavvadias, V., Konstantinidou, E., & Stamou, N. (2001). The efect of stand age on the accumulation of nutrients in the aboveground components of an Aleppo pine ecosystem. Forest Ecology Management, 141(3), 259–269. https://doi.org/10.1016/s0378-1127(00)00334-0
Bąk, A., Bazan, P., Pławecka, K., & Łach, M. (2023). Geopolymers based on fly ash from the bełchatów power plant. Materials Proceedings, 13, 17. https://doi.org/10.3390/materproc2023013017
Barrow, N. J., & Hartemink, A. E. (2023). The effects of pH on nutrient availability depend on both soils and plants. Plant and Soil, 487, 21–37. https://doi.org/10.1007/s11104-023-05960-5
Baumann, K., Schneider, B. U., Marschner, P., & Huttl, R. F. (2005). Root distribution and nutrient status of mycorrhizal and non-mycorrhizal Pinus sylvestris L. seedlings growing in a sandy substrate with lignite fragments. Plant and Soil, 276, 347–357. https://doi.org/10.1007/s11104-005-5300-y
Bekele, A., Roy, J. L., & Young, M. A. (2015). Use of biochar and oxidized lignite for reconstructing functioning agronomic topsoil: Effects on soil properties in a greenhouse study. Canadian Journal of Soil Science, 95, 269–285. https://doi.org/10.4141/cjss-2014-008
Borcz, A., & Kozioł, W. (2015). Systemy maszynowe w górnictwie odkrywkowym. Cz. 1. Maszyny w górnictwie polskim”. Nowoczesne Budownictwo Inżynieryjne, 4, 102–107. (in Polish).
Brandtberg, P. O., Bengtsson, J., & Lundkvist, H. (2004). Distributions of the capacity to take up nutrients by Betula spp. and Picea abies in mixed stands. Forest Ecology and Management, 198, 193–208. https://doi.org/10.1016/j.foreco.2004.04.012
Brożek, S., & Zwydak, M. (2010). Atlas gleb leśnych Polski. Centrum Informacyjne Lasów Państwowych. (in Polish).
Carnol, M., & Bazgir, M. (2013). Nutrient return to the forest floor through litter and throughfall under 7 forest species after conversion from Norway spruce. Forest Ecology and Management, 309, 66–75. https://doi.org/10.1016/j.foreco.2013.04.008
Cataldo, D. A., & Wildung, R. E. (1978). Soil and plant factors influencing the accumulation of heavy metals by plants. Environmental Health Perspectives, 27, 149–159. https://doi.org/10.1289/ehp.7827149
Clark, R. B. (1983). Plant genotype differences in the uptake, translocation, accumulation and use of mineral elements required for plant growth. Plant and Soil, 72, 175–196.
Clegg, S., & Gobran, G. R. (1995). Effects of aluminium on growth and root reactions of phosphorus stressed Betula pendula seedlings. Plant and Soil, 168–169, 173–178. https://doi.org/10.1007/BF00029326
Dang, Z., Liu, C., & Haigh, M. J. (2002). Mobility of heavy metals associated with the natural weathering of coal mine spoils. Environmental Pollution, 118, 419–426.
Daugaviete, M., Korica, A. M., Silins, I., Barsevskis, A., Bardulis, A., Bardule, A., Spalvis, K., & Daugavietis, M. (2015). The use of mineral nutrients for biomass production by young birch stands and stands vitality in different forest growing conditions. Journal of Environmental Science and Engineering, B4, 177–189. https://doi.org/10.17265/2162-5263/2015.04.002
Dmuchowski, W., Baczewska, A. H., & Gozdowski, D. (2012). Silver birch (Betula pendula Roth) as zinc hyperaccumulator. In J. Nriagu, J. Pacyna, P. Szefer, B. Markert, S. Wuenschmann, & J. Namieśnik (Eds.), Heavy metals in the environment (pp. 19–25). Maralte BV.
Eltrop, L., Brown, G., Joachim, O., & Brinkmann, K. (1991). Lead tolerance of Betula and Salix in the mining area of Mechernich/Germany. Plant and Soil, 131, 275–285. https://doi.org/10.1007/BF00009459
Frazer, J. (1935). Sulphur content of plants. Plant Physiology, 10, 529.
Frouz, J., Krištůfek, V., Bastl, J., KalčÍk, J., & VaňkovÁ, H. (2005). Determination of toxicity of spoil substrates after brown coal mining using a laboratory reproduction test with Enchytraeus crypticus (Oligochaeta). Water, Air, & Soil Pollution, 162, 37–47. https://doi.org/10.1007/s11270-005-5991-y
Fyson, A., Nixdorf, B., & Kalin, M. (2006). The acidic lignite pit lakes of Germany-Microcosm experiments on acidity removal through controlled eutrophication. Ecological Engineering, 28, 288–295. https://doi.org/10.1016/j.ecoleng.2006.06.012
Giergiczny, Z., & Michniewicz, E. (1991). Badanie procesów zestalania zaczynów z popiołów bełchatowskich, Górnictwo Odkrywkowe, 1. (in Polish)
Gordon, A. G., & Gorham, E. (1963). Ecological aspects of air pollution from an iron-sintering plant at Wawa, Ontario. Canadian Journal of Botany, 41, 1063.
Gruszczyński S. (2015). Accumulation of organic carbon in reclaimed soils. Dumping grounds: Bełchatów, Machów, Turów. Infrastructure and Ecology of Rural Areas, IV/4, 1417–1436. http://dx.medra.org/10.14597/infraeco.2015.4.4.102. (in Polish).
Hailes, K. J., Aitken, R. L., & Menzies, N. W. (1997). Magnesium in tropical and subtropical soils from north-eastern Australia. II. Response by glasshouse-grown maize to applied Magnesium. Soil Research, 35, 629–641. https://doi.org/10.3390/agronomy12112693
Huertas, J. I., Camacho, D. A., & Huertas, M. E. (2012). Standardized emissions inventory methodology for open pit mining areas. Environmental Science and Pollution Research, 19, 2784–2794.
Hycnar, E., Jończyk, M. W., & Ratajczak, T. (2017). Popioły lotne i iły beidellitowe z Bełchatowa jako składniki mieszanin samozestalających się. Zeszyty Naukowe Instytutu Gospodarki Surowcami Mineralnymi i Energią Polskiej Akademii Nauk, 100, 37–48. (in Polish).
Hynynen, J., Niemistö, P., Viherä-Aarnio, A., Brunner, A., Hein, S., & Velling, P. (2010). Silviculture of birch (Betula pendula Roth and Betula pubescens Ehrh.) in northern Europe. Forestry, 83(1), 103–119. https://doi.org/10.1093/forestry/cpp035
Hytönen, J., Saramäki, J., & Niemistö, P. (2014). Growth, stem quality and nutritional status of Betula pendula and Betula pubescens in pure stands and mixtures. Scandinavian Journal of Forest Research, 29(1), 1–11. https://doi.org/10.1080/02827581.2013.838300
Ivanov, Y. V., Pashkovskiy, P. P., Ivanova, A. I., Kartashov, A. V., & Kuznetsov, V. V. (2022). Manganese deficiency suppresses growth and photosynthetic processes but causes an increase in the expression of photosynthetic genes in scots pine seedlings. Cells, 11(23), 3814. https://doi.org/10.3390/cells11233814
IUSS Working Group WRB. (2022). World reference base for soil resources. International soil classification system for naming soils and creating legends for soil maps (4th ed). International Union of Soil Sciences (IUSS).
Jonczak, J. (2011). Struktura, dynamika i właściwości opadu roślinnego w 110-letnim drzewostanie bukowym z domieszką sosny i świerka. Sylwan, 155(11), 760–768. (in Polish).
Jonczak, J., Oktaba, L., Chojnacka, A., Pawłowicz, E., Kruczkowska, B., Oktaba, J., & Słowińska, S. (2023). Nutrient fluxes via litterfall in silver birch (Betula pendula Roth) stands growing on post-arable soils. European Journal of Forest Research, 142, 981–996.
Kabata-Pendias, A., & Pendias H. (1999). Biogeochemia pierwiastków śladowych. Wydawnictwo Naukowe PWN, Warszawa. (in Polish).
Katz, M., Ledingham, G. A., & McCallum, A. W. (1939). Effects of sulphur dioxide on vegetation. National Research Council of Canada.
Kołodziej, B., Bryk, M., & Otremba K. (2020). Effect of rockwool and lignite dust on physical state of rehabilitated postmining soil. Soil and Tillage Research, 199, 104603.
Kosztowniak, E., Ciężka, M., Zwoździak, A., & Górka, M. (2016). OC/EC from PM10 in the vicinity of Turów lignite open-pit mine (SW Poland): Carbon isotopic approach. Atmospheric Pollution Research, 7, 40–48. https://doi.org/10.1016/j.apr.2015.07.003
Kozioł, W., Machniak, Ł, Sośniak, E., & Chałupka, R. (2011). Technologia wydobycia trudno urabialnych skał nadkładowych na przykładzie kopalni węgla brunatnego “Bełchatów.” Górnictwo i Geoinżynieria, 35, 165–180. (in Polish).
Kříbek, B., Míková, J., Knésl, I., Mihaljevič, M., & Sýkorová, I. (2020). Uptake of trace elements and isotope fractionation of Cu and Zn by birch (Betula pendula) growing on mineralized coal waste pile. Applied Geochemistry, 122, 104741. https://doi.org/10.1016/j.apgeochem.2020.104741
Kuznetsova, T., Rosenvald, K., Ostonen, I., Helmisaari, H.-S., Mandre, M., & Lõhmus, K. (2010). Survival of black alder (Alnus glutinosa L.), silver birch (Betula pendula Roth.) and Scots pine (Pinus sylvestris L.) seedlings in a reclaimed oil shale mining area. Ecological Engineering, 36(4), 495–502. https://doi.org/10.1016/j.ecoleng.2009.11.019
Kwiatkowska-Malina, J. (2015). The influence of exogenic organic matter on selected chemical and physicochemical properties of soil. Polish Journal of Soil Science, 58(2), 173–180.
Linzon, S. N. (1965). Sulphur dioxide injury to trees in the vicinity of petroleum refineries. Forestry Chronicle, 417, 245–250.
Linzon, S. N. (1958). The Influence of smelter fumes on the growth of white pine in the sudbury region. Ontario Department Lands and Forests, Ontario Department Mines, Toronto, pp. 1–45.
Linzon, S. N., Temple, P. J., & Pearson, R. G. (1979). Sulfur concentrations in plant foliage and related effects. Journal of the Air Pollution Control Association, 29(5), 520–525. https://doi.org/10.1080/00022470.1979.10470822
Machava, J., & Barna, M. (2005). The influence of stand density on Mn and Fe concentrations in beech leaves. Journal of Forest Science, 51(5), 225–236. https://doi.org/10.17221/4593-JFS
Marguí, E., Queralt, I., Carvalho, M. L., & Hidalgo, M. (2007). Assessment of metal availability to vegetation (Betula pendula) in Pb-Zn ore concentrate residues with different features. Environmental Pollution, 145, 179–184. https://doi.org/10.1016/j.envpol.2006.03.028
Martins, T. G. V., Rocha, M. F. V., Nieri, É. M., Melo, LAd., Silva, MLd. S., & Silva, D. SNd. (2019). Nutrient accumulation in eucalyptus bark at different population densities. Revista Brasileira De Engenharia Agrícola E Ambiental, 23(1), 40–46. https://doi.org/10.1590/1807-1929/agriambi.v23n1p40-46
Miller, H. G. (1995). The influence of stand development on nutrient demand, growth and allocation. Plant and Soil, 168, 225–232. https://doi.org/10.1007/BF00029332
Modrzewska, B., Kosiorek, M., & Wyszkowski, M. (2016). Content of some nutrients in Scots pine, silver birch and Norway maple in an urbanized environment. Journal of Elementology, 21(1), 149–157. https://doi.org/10.5601/jelem.2015.20.2.845
Novák, J., Dušek, D., Kacálek, D., & Slodičák, M. (2017). Nutrient content in silver birch biomass on nutrient-poor, gleyic sites. Zpravy Lesnickeho Vyzkumu, 62(3), 135–141.
Pająk, M., Halecki, W., & Gąsiorek, M. (2016). Accumulative response of Scots pine (Pinus sylvestris L.) and silver birch (Betula pendula Roth) to heavy metals enhanced by Pb-Zn ore mining and processing plants: Explicitly spatial considerations of ordinary kriging based on a GIS approach. Chemosphere, 168, 851–859. https://doi.org/10.1016/j.chemosphere.2016.10.125
Pająk, M., & Krzaklewski, W. (2006). Selected chemical properties of initial soils on the outside spoil bank of the “Bełchatów” mine. Soil Science Annual, 57(1/2), 158–163.
Pająk, M., Woś, B., Likus-Cieślik, J., Bobik, I., Otremba, K., & Pietrzykowski, M. (2023). Effect of properties of technogenic soils developed from different lignite spoil heap materials on the growth of Scots pine Pinus sylvestris L. Sylwan, 167(2), 61–77. https://doi.org/10.26202/sylwan.2023017
Pandey, B., Agrawal, M., & Singh, S. (2014). Assessment of air pollution around coal mining area: Emphasizing on spatial distributions, seasonal variations and heavy metals, using cluster and principal component analysis. Atmospheric Pollution Research, 5, 79–86. https://doi.org/10.5094/APR.2014.010
Perring, M. P., Hedin, L. O., Levin, S. A., McGroddy, M., & Mazancourt, Cd. (2008). Increased plant growth from nitrogen addition should conserve phosphorus in terrestrial ecosystems. Proceedings of the National Academy of Sciences, 105(6), 1971–1976. https://doi.org/10.1073/pnas.0711618105
Peukert, J., Thiel, A., & Kania J. (1986). Wykorzystanie popiołu lotnego z węgla brunatnego Bełchatów. Cz. II. Popiół jako surowiec „niski” do produkcji klinkieru portlandzkiego”. Cement Wapno Gips, 1. (in Polish).
Pietrzykowski, M., Krzaklewski, W., & Woś, B. (2013). Zawartość pierwiastków śladowych (Mn, Zn, Cu, Cd, Pb, Cr) w liściach olsz (Alnus sp.) zastosowanych jako gatunki fitomelioracyjne na składowisku odpadów paleniskowych. Zeszyty Naukowe. Inżynieria Środowiska/Uniwersytet Zielonogórski, 151(31), 26–34. (in Polish).
PTG. (2009). Klasyfikacja uziarnienia gleb i utworów mineralnych – PTG 2008. Roczniki Gleboznawcze – Soil Science Annual, 60(2), 5–17. (in Polish).
Pudełko, A., & Chodak, M. (2020). Estimation of total nitrogen and organic carbon contents in mine soils with NIR reflectance spectroscopy and various chemometric methods. Geoderma, 368, 11430606. https://doi.org/10.1016/j.geoderma.2020.114306
Qin, K., & Leskovar, D. (2018). Lignite‐derived humic substances modulate pepper and soil‐biota growth under water deficit stress. Journal of Soil Science and Plant Nutrition, 181(5). https://doi.org/10.1002/jpln.201800078
Ratajczak, T., & Hycnar E. (2017). Kopaliny towarzyszące w złożach węgla brunatnego. Geologiczno-surowcowe aspekty zagospodarowania kopalin towarzyszących. Wydawnictwo Instytutu Gospodarki Surowcami Mineralnymi i Energią Polskiej Akademii Nauk (in Polish).
Regulation of the Minister of the Environment of 1 September 2016 on the Method of Assessing the Pollution of the Earth’s Surface. J. Laws 2016, item 1395.
Reimann, C., Arnoldussen, A., Finne, T. E., Koller, F., Nordgulen, Ø., & Englmaier, P. (2007). Element contents in mountain birch leaves, bark and wood under different anthropogenic and geogenic conditions. Applied Geochemistry, 22, 1549–1566.
Rout, T. K., Masto, R. E., Ram, L. C., George, J., & Padhy, P. K. (2013). Assessment of human health risks from heavy metals in outdoor dust samples in a coal mining area. Environmental Geochemistry and Health, 35(3), 347–356.
Rumpel, P., Grootes, M., & Kögel-Knabnerc, I. (2001). Characterization of the microbial biomass in lignite-containing mine soils by radiocarbon measurements. Soil Biology and Biochemistry, 33(14), 2019–2021. https://doi.org/10.1016/S0038-0717(01)00122-5
Rustowska, B. (2022). Long-term wildfire effect on nutrient distribution in silver birch (Betula pendula Roth) biomass. Soil Science Annual, 73(2), 149943. https://doi.org/10.37501/soilsa/149943
Rustowska, B. (2024). Nutrient distribution and bioaccumulation in silver birch (Betula pendula Roth) biomass grown in nutrient-poor soil. Trees. https://doi.org/10.1007/s00468-024-02492-y
Scalon, M. C., Wright, I. J., & Franco, A. C. (2017). To recycle or steal? Nutrient resorption in Australian and Brazilian mistletoes from three low phosphorus sites. Oikos, 126, 32–39. https://doi.org/10.1111/oik.03455
Sekhohola, L. M., Igbinigie, E. E., & Cowan, A. K. (2013). Biological degradation and solubilisation of coal. Biodegradation, 24, 305–318.
Sharma, B., & Sharma, K. (2013). Nutrient accumulation in various plant parts of dominant tree species of three different localities. Pakistan Journal of Biological Sciences, 16(18), 965–968. https://doi.org/10.3923/pjbs.2013.965.968
Shorohova, E., & Kapitsa, E. (2016). The decomposition rate of non-stem components of coarse woody debris (CWD) in European boreal forests mainly depends on site moisture and tree species. European Journal of Forest Research, 135, 593–606. https://doi.org/10.1007/s10342-016-0957-8
Simmler, M., Ciadamidaro, L., Schulin, R., Madejón, P., Reiser, P., Clucas, L., Weber, P., & Robinson, B. (2013). Lignite reduces the solubility and plant uptake of cadmium in pasturelands. Environmental Science & Technology, 47(9), 4497–4504. https://doi.org/10.1021/es303118a
Sitko, K., Opała-Owczarek, M., Jemioła, G., Gieroń, Z., Szopiński, M., Owczarek, P., Rudnicka, M., & Małkowski, E. (2022). Effect of drought and heavy metal contamination on growth and photosynthesis of silver birch trees growing on post-industrial heaps. Cells, 11, 53. https://doi.org/10.3390/cells11010053
Smith, S., & Smith, F. (2011). Roles of arbuscular mycorrhizas in plant nutrition and growth: New paradigms from cellular to ecosystem scales. Annual Review of Plant Biology, 62(1), 227–250. https://doi.org/10.1146/annurev-arplant-042110-103846
Staddon, P. L., Ramsey, C. B., Ostle, N., Ineson, P., & Fitter, A. H. (2003). Rapid turnover of hyphae of mycorrhizal fungi determined by AMS microanalysis of C-14. Science, 300, 1138–1140. https://doi.org/10.1126/science.1084269
Stolecki, L. (2005). Wpływ sposobów składowania odpadów paleniskowych w wyrobisku końcowym odkrywki Bełchatów na środowisko wodne. Praca doktorska, Politechnika Wrocławska, Wydział Geoinżynierii, Górnictwa i Geologii, Wrocław. (in Polish).
Tran, C. K. T., Rose, M. T., Cavagnaro, R. T., & Patti, A. F. (2015). Lignite amendment has limited impacts on soil microbial communities and mineral nitrogen availability. Applied Soil Ecology, 95, 140–150. https://doi.org/10.1016/j.apsoil.2015.06.020
Tromp, J. (1983). Nutrient reserves in roots of fruit trees, in particular carbohydrates and nitrogen. Plant and Soil, 71, 401–413. https://doi.org/10.1007/BF02182682
Ulbrichová, I., Podrázský, V., & Slodičák, M. (2005). Soil forming role of birch in the ore mts. Journal of Forest Science, 51, 54–58. https://doi.org/10.17221/11850-JFS
Uri, V., Vares, A., Tullus, H., & Kanal, A. (2007). Above-ground biomass production and nutrient accumulation in young stands of silver birch on abandoned agricultural land. Biomass and Bioenergy, 31(4), 195–204. https://doi.org/10.1016/j.biombioe.2006.08.003
Uzarowicz, Ł, Kwasowski, W., Śpiewak, O., & Świtoniak, M. (2018). Indicators of pedogenesis of Technosols developed in an ash settling pond at the Bełchatów thermal power station (central Poland). Soil Science Annual, 69(1/2018), 49–59.
Vos, A. E. M., Boer, D., Vries, Ouden, J., & Sterckz, F. J. (2023). Aboveground carbon and nutrient distributions are hardly associated with canopy position for trees in temperate forests on poor and acidified sandy soils. Forest Ecology and Management, 529, 120731. https://doi.org/10.1016/j.foreco.2022.120731
Vrablik, P., Wildova, E., & Vrablikova, J. (2017). The effect of brown coal mining on the environment and health of the population in Northern Bohemia (Czech Republic). International Journal of Clean Coal and Energy, 06, 1–13. https://doi.org/10.4236/ijcce.2017.61001
Wagner, M., & Słomka T. (2000). Charakter petrograficzny i warunki sedymentacji wybranych kompleksów litologicznych z profilu miocenu w złożu węgla brunatnego Bełchatów. Wydawnictwo Instytutu Gospodarki Surowcami Mineralnymi i Energią PAN (in Polish).
Warfvinge, P., & Svedrup, H. (1995). Critical loads of acidity to Swedish Forest Soils. Methods, data and results. Reports in Environmental Engineering and Ecology, 5, 1–127. Chemical Engineering, Lund University. Lund, Sweden.
Węgiel, A., Bielinis, E., & Polowy, K. (2018). Macronutrient stocks in Scots pine stands of different densities. Forests, 9, 593. https://doi.org/10.3390/f9100593
Wójcik, J., & Krzaklewski, W. (2009). Forestation as a Method of the Remediation of Soilless Areas of the Lignite Mine Turów. Gospodarka Surowcami Mineralnymi, 25(3), 171–187. (in Polish).
Wójcik, J., Pająk, M., Kowalik, S., & Krzaklewski, W. (2012). Changes of the properties of initial soil within 10 years since the accomplishment of reclamation measures on toxically acid grounds of the outer dump in the “Bełchatów” Lignite Mine. Przegląd Górniczy, 8, 61–68. (in Polish).
Zhang, D., & Yani, S. (2011). Sulphur transformation during pyrolysis of an Australian lignite. Proceedings of the Combustion Institute, 33, 1747–1753. https://doi.org/10.1016/j.proci.2010.07.074
Author information
Authors and Affiliations
Contributions
Beata Rustowska: conceptualization, data curation, formal analysis, investigation, methodology, validation, visualization, writing—original draft, writing—review & editing; Jerzy Jonczak: investigation, validation, supervision; Artur Pędziwiatr: investigation.
Corresponding author
Ethics declarations
Ethical Responsibilities of Authors
All authors have read, understood, and have complied as applicable with the statement on “Ethical responsibilities of Authors” as found in the Instructions for Authors.
Competing Interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rustowska, B., Jonczak, J. & Pędziwiatr, A. Nutrient Accumulation in Silver Birch (Betula pendula Roth) Biomass in a Lignite Mining Area. Water Air Soil Pollut 235, 440 (2024). https://doi.org/10.1007/s11270-024-07254-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11270-024-07254-7