
Abstract
Ponds are key elements for ecosystem functions in urban areas. However, little is known about pond biodiversity changes over time and the drivers underlying such changes. Here, we tested whether past species assemblages, land cover and pond environmental change influence pond macroinvertebrate species richness and temporal beta diversity. We also compared spatial and temporal beta diversity, and investigated species-specific colonization and extinction rates over time. We sampled for presence of Odonata and Trichoptera (larvae), and Coleoptera and Hemiptera (larvae and adults) species in 30 ponds in Stockholm, Sweden, in 2014 and 2019. Past species richness was the best predictor of current species richness, whereas temporal changes in land cover and pond environment were not significantly related to current species richness. No correlations between temporal beta diversity and land cover or pond environmental changes were detected. However, ponds showed large changes in their temporal beta diversity, with equal contributions from species gains and losses. The probability of species colonizing and going extinct from ponds revealed that more common species were more likely to colonize a pond, while uncommon species were more likely to go extinct in a pond. Within our 5-year study, we found (i) highly similar spatial and temporal beta diversity, (ii) that past species richness is a good predictor of current species richness; however, the same does not hold true for species composition. The high dynamics of urban pond communities suggest that a large number of ponds are required to maintain high species richness at a landscape level.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It has been predicted that about 68% of the human population will be living in urban areas by 2030 (United Nations 2018). This trend reinforces the importance of urban green spaces, as they provide different ecosystem services, such as flood protection, water purification and improved mental and physical health (Blicharska and Johansson 2016; Garrett et al. 2019; Jim and Cheng 2006; Hassall 2014; Maragno et al. 2018). Urban green spaces can also have high biodiversity, sometimes including unique and red-listed species (Planchuelo et al. 2020). For better management of ecosystem services in urban environments, we need information on how environmental, spatial and temporal factors affect biodiversity. While numerous studies have focused on these three factors separately or in paired comparisons in urban areas (Aronson et al. 2014; Beninde et al. 2015; Jimenez et al. 2022; Nava-Díaz et al. 2022), very few have focused on all of them simultaneously in the same study system.
In addition to green spaces, urban centers also harbor blue spaces. Urban blue spaces include rivers, canals, lakes, reservoirs, wetlands, and ponds (Hill et al. 2017b; Oertli and Parris 2019; Smith et al. 2021). In recent years, there has been an increase in the number of studies on how biodiversity in blue spaces is affected by local and landscape variables. Three major findings have emerged from studies on ponds in the urban environment. First, biodiversity is affected by pond-related environmental variables such as plant community composition and water chemistry (Heino et al. 2017a; Hill et al. 2019; Perron et al. 2021). Second, the surrounding landscape (portrayed by land cover variables) in the urban area also affects biodiversity, and ponds with a higher proportion of green area and a low proportion of built-up areas around them usually have higher biodiversity (Hamer and Parris 2011; Heino et al. 2017a). Third, connectivity patterns also seem to play a role (Hill et al. 2017b). For example, Hyseni et al. (2021) showed that blue and green connectivity are important variables affecting community composition in ponds. However, since changes in land cover variables in the landscape can cascade down to the physical and biotic environment of ponds, it is also valuable to know how changes in these land cover variables affect biodiversity in ponds across time. A recent study, for example, showed that an increasing amount of developed area affected functional but not compositional turnover of bird species assemblages in an urban area (Petersen et al. 2022), but studies on how land use changes in urban areas affect pond diversity are rare.
Macroinvertebrates are an important part of urban pond diversity, generally harboring hundreds of species across urban landscapes (Heino et al. 2017a; Thornhill et al. 2017). A set of urban ponds can be viewed as a metacommunity (Leibold et al. 2004), where species composition is determined jointly by the pond environment (i.e., niche-based processes such as environmental filtering) and dispersal between ponds (Chase and Myers 2011; Heino 2013; Hill et al. 2017a). Despite being important, niche-based processes have been shown to not fully explain community composition (Heino et al. 2015) as, for example, species are unlikely to occupy all suitable locations within a metacommunity due to dispersal limitation or stochastic factors (Bispo et al. 2017; but see Siqueira et al. 2012). Community change might also occur due to stochastic processes (random extinctions and colonizations). Under such neutral processes, extinction and colonization of patches should be linearly related to population size (Hubbell 2001). Divergence from niche-based or neutral processes could potentially identify unusually persistent or non-persistent species in an urban landscape (Jabot et al. 2020). For an effective management of urban pond biodiversity, knowledge of such species-specific differences would be valuable.
By sampling the same blue spaces over time, one can test whether past community assemblage and environmental variables are predictors of the current species diversity. Studies from non-urban areas have indeed shown that the past community structure can explain a large part of the currently observed community structure (Oliveira et al. 2020). However, an increase in time intervals between samplings should result in a lower congruence between current and past community composition (Ruhí et al. 2012). Nevertheless, by sampling the same localities over time, it is possible to explore how temporal changes in pond biodiversity are determined by local environmental variables, land cover variables and stochastic processes. Also, the comparison between past and current communities allows us to evaluate how important colonization history is for the currently observed biodiversity in urban ponds. Such information would provide information on whether past biodiversity or environmental variables are the most important in driving biodiversity patterns across the urban landscape. Understanding such dynamics is valuable for the management of freshwater ecosystems when it comes to predicting effects of land-use changes on biodiversity (Lindholm et al. 2021).
Many biodiversity measures that can be used to understand the drivers of biodiversity patterns across space and time (Anderson et al. 2011) in urban ponds. Here, we used two measures. Species richness (alpha diversity) is the simplest measure with a clear definition, while beta diversity is more complex. In its simplest form, beta diversity can be defined as the turnover of species assemblages from place to place or time to time (Soininen 2010). However, this turnover can be quantified and partitioned in different ways (e.g., Baselga 2010; Cardoso et al. 2014; Legendre 2019) to understand the underlying ecological causes of community compositional difference between sites. Such detailed investigations can uncover the mechanisms that maintain regional species and community composition, thereby aiding urban planning and conservation efforts (Socolar et al. 2016; Liu et al. 2022).
Here, we aimed to test the effects of environmental changes (both changes in the local pond environment and land cover around ponds) on (i) species richness and (ii) temporal beta diversity of macroinvertebrate communities. In addition, we aimed to (iii) compare temporal beta diversity (temporal differences in community composition within ponds; e.g. Lindholm et al. 2021) to spatial beta diversity (spatial differences in community composition between ponds; e.g. Lindholm et al. 2020). Finally, we aimed to (iv) investigate species-specific responses over time to understand the ecological mechanisms underpinning diversity changes, and determine which species persist at both local community and metacommunity levels.
Methods
Study area
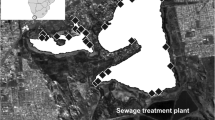
We sampled the same 30 ponds in the city of Stockholm 5 years apart, in 2014 and 2019 (Fig. 1). The spatial extent of the study area covers approximately 300 km2. Sampling was done in May or June and the same method was used for sampling all ponds in both years. Ponds were defined as natural or man-made water bodies with an area between 2 m2 and 2 ha and holding water for at least 4 months of the year (Biggs et al. 2005). In order to create a temporal dataset from which to gain novel insights using a new analysis workflow, we compiled data from previous studies of urban biodiversity in these ponds (Blicharska et al. 2016, 2017; Heino et al. 2017a; Hyseni et al. 2021), and complemented it with a new set of variables capturing land cover change.

Map showing the distribution of the 30 studied ponds (white dots) in northern Stockholm, Sweden. The orange, green, yellow, and dark-blue colors represent artificial surfaces, forests, grasslands, and water, respectively
Data collection
The data we analyzed in this study comprised the following insect groups: Odonata (larvae), Trichoptera (larvae), Coleoptera (larvae and adults) and Hemiptera (larvae and adults). Many studies, including ours, assume that these taxa are representative of the overall much larger biodiversity of aquatic insects found in pond settings, especially because they consist of a variety of functional groups (Hassall et al. 2011; Sánchez-Fernándes et al. 2006; but see Heino 2010 and Westgate et al. 2014). However, we note that some taxa that were not sampled, may be more sensitive to environmental change and one would then underestimate the effect of changing environmental conditions. Six subsamples were taken in each pond at a depth of 20–30 cm with a bottom scoop net (diameter of 20 cm, mesh size of 1.5 mm). These six subsamples included a variety of shoreline-representative microhabitats. A 1 m length of the bottom was swept with the net eight times (back and forward) composing each subsample. Then, we pooled all sub-samples from a given pond to avoid pseudoreplication in our analyses described below.
All insects were stored in 70% ethanol and then classified to the species level in the laboratory. For some specimens, species-level identification was not possible. These species were classified at the genus or family level, and were regarded as separate taxa in the analyses. Larval forms of Coenagrion pulchellum (Linnaeus, 1758) and Coenagrion pulchella (Vander Linden, 1825) are difficult to differentiate and were, therefore, treated as one species in the analysis (Coenagrion puella/pulchellum). Specimens belonging to the genus Lestes (Odonata) were identified to species level in 2019 but only to genus level in 2014. Consequently, we grouped all these records into Lestes sp. Several species of the order Trichoptera could not be distinguished as well. In the analyses, four species in Limnephilidae were regarded as Limnephilidae (Limnephilus affinis Curtis 1834, Limnephilus incisus Curtis 1834, Limnephilus luridus Curtis 1834, and Limnephilus ignavus McLachlan 1865), and Oligotricha striata (Linnaeus 1759) and Oligotricha lapponica (Hagen 1864), were treated as one taxon (O. stricta/lapponica). Nymphs of the family Corixidae and the genus Notonecta were each treated as a taxon.
In each pond, we measured pH, total nitrogen (TN), total phosphorus (TP), total organic carbon (TOC), and water depth. Furthermore, we visually categorized different types of vegetation cover in the terrestrial and littoral interface (bushes, bareground, floating, and emergent vegetation) by using ordinal variables varying from zero (no cover for a given type) to ten (representing 100% cover for a given type in the sampling unit; for more details on methods, see Blicharska et al. 2017 and Hyseni et al. 2021).
We estimated land cover variables using data from Copernicus Land Monitoring Service (https://land.copernicus.eu/, CLMS 2023). This service did not have land cover data for 2014 and 2019. We, therefore, used data for the years 2015 and 2018 in our analysis. We extracted raster files for impervious surfaces, grasslands, and forests (products: IMCC_1518_020m_E47N40_03035_v010, GRAC_1518_020m_E47N40_03035_v010, and TCCM_1518_020m_E47N40_03035_v010). Forest cover showed little temporal variation across ponds (mean: 0, range: 0–2%) and we excluded this land cover category in further analyses. CLMS classifies land cover with images from Sentinel-1 and Sentinel-2 satellites at a 20 m resolution (CLMS, 2023). We built a 250 m-buffer around each pond and quantified the number of pixels (20 m × 20 m) belonging to each land cover category. Then, we divided the number of pixels per category by the total number of pixels in a buffer area resulting in a relative measure varying from 0 to 100%. We are aware that the 2018 data are based on a higher resolution (10 m × 10 m), which improves the detection of habitats. However, this is primarily a problem when focusing on absolute cover changes, but presumably of minimal significance when comparing cover changes among ponds in a limited spatial area.
Data analysis
Our analyses did not include tests on how the variation in land cover or environmental pond variables explained spatial species diversity patterns. This type of analysis has already been performed on the same data in previous studies (Blicharska et al. 2016, 2017; Heino et al. 2017b; Hyseni et al. 2021). Here, we focus on how temporal changes in environmental variables affect species diversity, while spatial variation in diversity is included for comparison.
The presented analyses were performed including genus and family level taxa. We also ran all analyses excluding taxa identified to family and genus levels taxa to test whether these high-level taxa had a large impact on the results. Despite the large number of taxa (27), these subset analyses showed very similar results (see R code, https://doi.org/10.5281/zenodo.10215976). In addition, if only the seven taxa at the family level taxa were excluded, the results were also nearly identical.
All data analyses were performed using the statistical software R, version 4.2.1 (R Core team 2022). Sample-based rarefaction curves were plotted to evaluate how much of the species pool our sampling covered. Curves were calculated using the R package iNEXT (Chao et al. 2014; Hsieh et al. 2020). To test whether species richness changed between 2014 and 2019, and to compare spatial and temporal variation of species richness (i.e., variance partitioning of within- and between-pond variation), we fitted a linear mixed-model with time as a fixed factor (excluded for variance partitioning) and pond as a random factor using the lme4 package (Bates et al. 2015). An F-test was used to test the time effect (lmerTest package; Kuznetsova et al. 2017) and if the spatial and temporal variance components were unequal. Temporal beta diversity was evaluated using multiple metrics. First, we used the adespatial R package (ver. 0.3, Dray et al. 2022) to calculate the Temporal Beta diversity Index (TBI), as described by Legendre (2019). We used the Jaccard dissimilarity index in TBI to quantify species composition differences between 2014 and 2019 for each pond. We tested for unusual changes in TBI values for each pond with permutations tests based on 9999 permutations. We applied the Holm correction for multiple testing. TBI also provides the contributions of gains and losses of species to dissimilarities (Legendre 2019). Hence, this approach is designed to identify ponds where large community changes have taken place, and to provide information on the relative importance of species gains and losses for temporal species turnover.
Second, we applied Baselga (2010) partitioning of beta diversity (here called total beta diversity, i.e., Jaccard dissimilarity index) into species turnover and nestedness components of community variation using the R package betapart (Baselga et al. 2023). Turnover is the replacement of species across the landscape (here, between ponds) or between time points (2014 and 2019), and nestedness occurs when species-poor communities are subsets of species-rich communities across ponds or over time. These metrics were calculated using the R package betapart (Baselga et al. 2023). To compare spatial and temporal beta diversity for total, turnover, and nestedness, we calculated spatial beta diversity as the mean of pairwise dissimilarities between pond i and all other ponds in 2014, and temporal beta diversity is the pairwise dissimilarity between 2014 and 2019 for each pond. The spatial beta diversity in 2014 and 2019 were nearly identical (Table 1), thus we used 2014 as the baseline for comparing it to the temporal beta diversity.
To estimate the degree of environmental change for each pond, we followed Legendre (2019), and calculated the standardized Euclidean distance between the years 2014 and 2019 using the pond variables (pH, TN, TP, TOC, pH, depth, bushes, bare ground, floating and emergent vegetation cover). We also used the standardized Euclidean distance between the years 2014 and 2019 to estimate the change in the land use variables (grassland, and impervious surfaces). Ponds with a higher standardized Euclidean distance indicate a higher change in their environmental variables from 2014 to 2019 (or 2015 to 2018 for land cover), while those with distances closer to zero indicate that their environmental variables were similar between 2014 and 2019.
The relationship between changes in land cover and pond environment, as well as pond area, with macroinvertebrate species richness and temporal beta diversity, were assessed using structural equation modeling (SEM; Shipley 2016). SEM is a technique that allows to assess both direct and indirect relationships between variables in a single analysis (Shipley, 2016) and we used the R package piecewiseSEM for model fitting (Lefcheck 2016). The assessed causal diagram hypothesizes that changes in land cover variables influence changes in macroinvertebrates species richness and turnover (beta diversity), either indirectly by changing the local pond environment, or directly. For example, we expected that increase in impervious surface and a decrease in tree and grassland cover would affect environmental change in the pond themselves (in water and along the shore) between the years. With only 30 ponds in the dataset, only a few predictors can be included in the model to avoid overfitting, and we therefore used the above described Euclidean distances for land cover change and pond environment change (i.e. two change variables). The first SEM focused on species richness, where the final response variable was species richness in 2019. By including species richness in 2014 as a predictor in our model we: (i) tested whether the spatial patterns in species richness in 2014 were kept in 2019, and (ii), whether the other predictors influenced species richness change between 2014 and 2019. The second SEM tested the effect on temporal beta diversity (total, and the two components species gains and species loss). These SEMs included pond area to account for the potential effects of habitat area on species richness. To test for spatial autocorrelation in richness, temporal beta diversity, and model residuals, we used the Moran’s I autocorrelation index. This test was performed with the R package ape (Paradis and Schliep 2019).
To test for overall community change between the years and identify which species changed over time, we used the R package mvabund (Wang et al. 2022). Multivariate analyses with many rare species can be hard to interpret, especially if species-specific changes are of interest. Here, we aimed to investigate whether a set of more common species changed between years and, therefore, we removed species with only five or fewer observations over both years. We fitted a multivariate logistic regression model (complementary log-log link) with the presence/absence species matrix as the response and time as the predictor. Ridge regularization was applied to account for the correlation between variables (Warton 2008), and the model was evaluated using score statistics and restricted permutations using the permute package (Simpson 2022) to account for repeated measurements. In addition to the unadjusted p-values, we also give p-values that account for multiple testing.
To investigate species persistence at both local community and metacommunity levels, we used changes in species presence/absence as proxies for pond colonization/extinction. Our sampling did not encompass the whole pond, and it is not possible to distinguish between the effect of not finding the species and true absences. Hence, the processes of pond colonization and extinction need to be interpreted cautiously in our study. To estimate local extinction and colonization probabilities of species, we applied bivariate odds-models described by Yee and Dirnböck (2009) implemented in the VGAM R package (Yee 2010). In short, this approach uses two time points as response variables and estimates the odds ratio for each species. If the odds ratio is equal to one for a species, the two points are independent and a species’ local colonization/extinction probability is the marginal probability (named ps) of being present/absent at a pond. If the odds ratio is much greater than one, colonization and extinction probability is lower at time point two compared to the independent scenario. Likewise, an odds ratio close to zero indicates a higher colonization and extinction probability, which would result in a higher species turnover.
Macroinvertebrate and pond environmental data, and R script are available at Zenodo https://doi.org/10.5281/zenodo.10215976 (Granath et al. 2023). Land cover data from Copernicus can be found at https://land.copernicus.eu/pan-european/high-resolution-layers under expert products.
Results
Sampled communities were dominated by rare species (rare at the landscape scale, i.e., with species occurring only in a few ponds), and species accumulation curves suggested that all species had not been detected (Fig. 2). Merging the two sampling years suggests that the complete species pool has not been sampled, but it is clear that a larger number of samples would be needed to achieve this as the curve is slowly reaching its asymptote. In total, we recorded 117 taxa (88 identified to species level), and 56 were recorded in both years. The two most common taxa (determined to species or sister-species level) were Coenagrion puella-pulchellum, Odonata, (found in 31 of 60 samples) and Haliplus ruficollis (De Geer, 1774), Coleoptera, (found in 30 samples), and they were repeatedly detected at 9 and 8 ponds, respectively (Fig. 3).

Sample-based rarefaction curves for 2014, 2019, and both years together

The ten most recurring taxa between the surveys and the number of ponds where the species was found in both 2014 and 2019. Note that genera, and particularly the family Corixidae, likely include several species not identified in this study
Mean species richness did not change over the study period (Table 1, F1,29=0.50, P = 0.49). The proportion of spatial (for 2014 and 2019) and temporal variation of species richness were 49% and 51%, respectively. We did not detect a difference in spatial variation (between ponds variance) of species richness between the two sampling years (F1,29=0.80, P = 0.54). The temporal beta diversity index (TBI) was large, and on average only 15% of the species occurred in both years, with equal gain and loss of species over time. Species gains and losses represented 52% and 48% of the dissimilarity coefficient between 2014 and 2019, respectively. No ponds had significant TBI values (i.e., all P > 0.05) between the 2014 and 2019 sampling after adjusting for multiple testing, and without such adjustments four ponds had a p-value between 0.03 and 0.05. Hence, there was weak evidence that ponds showed unusual changes between the two years examined.
To compare spatial and temporal beta diversity, we applied the concept of turnover, nestedness and total dissimilarity (sensu Baselga 2010). Our results indicated that spatial and temporal beta diversity were very similar regardless of the component (i.e., turnover, nestedness and total dissimilarity) investigated (Fig. 4). In addition, spatial beta diversity was almost identical at the two sampling years (Table 1).

Boxplot of the turnover and nestedness components of beta diversity. Spatial data is represented by mean pairwise Jaccard dissimilarity for each pond in 2014 (n = 30), and temporal data by pairwise Jaccard dissimilarity between 2014 and 2019 for each pond (n = 30)
Structural equation modeling
Local (pond) environmental variables showed large change over time and average land cover change (absolute % change) for grassland was − 3% (range: -33 to 19%), and for impervious land − 2% (range: -13 to 1%) (Supplementary Information Figure S1). Our SEM for species richness in 2019 had weak support for most paths (Supplementary Information Table S1; Fig. 5). As expected, previous richness (year 2014) influenced richness in 2019, where a higher richness in 2014 resulted in a higher richness in 2019. Temporal change in land cover or pond environment, and pond area, tended to result in lower richness, but these effects were not detected statistically (all P > 0.05). We found no support for the SEM testing change in total temporal beta diversity (Supplementary Information Table S1, Fig. 6). This was also true for the two components of temporal beta diversity (species gain, species loss; results not shown).

Direct and indirect effect of previous species richness, pond area, temporal change of pond (local) environment, and temporal change of land cover variation on pond species richness in 2019 assessed by Structural Equation Modeling. A solid line means that the path coefficient was statistically significant (P < 0.05). Numbers close to arrows indicate standardized coefficients

Direct and indirect effect of pond area, temporal change of pond (local) environment, and temporal change of land cover variation on temporal beta diversity (2014–2019) in ponds assessed by Structural Equation Modeling. All path coefficients (P > 0.10). Numbers close to arrows indicate standardized coefficients
There was no strong evidence for spatial autocorrelation in temporal beta diversity for total (P = 0.05, Moran’s I = 0.08), gain (P = 0.47), loss (P = 0.93), or changes in species richness (all P = 0.21). The residuals of beta diversity SEM showed weak spatial autocorrelation (P = 0.03, Moran’s I = 0.10).
Species responses
After selecting species with six or more observations, we retained 30 species in the multivariate analysis. Species composition differed between years (score = 48, P < 0.001). Taxa driving this difference were (Unadjusted P-values, P < 0.05): Coenagrion hastulatum (Charpentier, 1825), Coenagrion puella/pulchellum, Corixidae, Graphoderus sp., Haliplus heydeni (Wehncke, 1875), Hydaticus ruficollis (Hope 1838), Hydroporus palustris (Linnaeus, 1761), Ilyocoris cimicoides (Linnaeus, 1758), Libellulidae, Notonecta sp., Rhantus sp. After adjusting the p-values for multiple comparisons the following taxa were still significant: Corixidae, Haliplus heydeni, Libellulidae, Notonecta sp.
Examining the probability of species colonizing and going extinct from ponds showed that more common species (i.e., higher probability that the species is found at any pond across years, here called ps) were more likely to colonize a pond (Fig. 7). This relationship was close to the expected odds ratio of 1, meaning that the sampling response of a species in 2019 (presence/absence) was independent of the response in 2014. The pattern for extinction from ponds was slightly different. Although many species fell along the odds ratio 1 line, most species showed an odds ratio above 1, meaning a lower probability of going extinct if they were present in 2014. Particularly, Aeshna grandis (Linnaeus, 1758), Hydrochara caraboides (Linnaeus, 1758), and Noterus crassicornis (Müller, 1776) showed low extinction probability despite being rather rare in the landscape (see lower left corner of Fig. 7B). They all had a high odds ratio (> 22), meaning that the species were persistent in the few ponds they were detected in 2014. In contrast, many species showed no temporal correlation, i.e., the species were lost from all ponds where they occurred in 2014. This was common for all the rare species, but also a few more common species (here, ‘common’ is defined as found in at least 5 ponds across the years) showed this pattern. These species have an extinction probability of 1 (Fig. 7B). Examples are the species Haliplus heydeni, that disappeared from 15 ponds between 2014 and 2019, Hydroporus palustris that was lost from 7 ponds, or Coenagrion hastulatum and Limnephilus binotatus (Curtis, 1834) that were both lost from 6 ponds. However, it should be noted that it can be difficult to differentiate between real extinctions and low detection rates of species with few individuals, i.e., species that are naturally rare or difficult to sample.

Local (A) colonization and (B) extinction probabilities for macroinvertebrate species sampled repeatedly in ponds in 2014 and 2019, and with at least five observations (30 species). Dashed lines represent an odds ratio of 1, and correspond to complete independence of the species responses at the two time points, i.e., the probability of extinction or colonization of a species at a specific pond is not related to the species’ presence/absence in 2014. ps is the marginal probability of being present/absent at a pond. The three species that diverge clearly from 1:1 line in panel B (low ps and low extinction probability) are Aeshna grandis, Hydrochara caraboides, and Noterus crassicornis, and are indicated by filled points. Species with an extinction probability of 1.0 were detected only in 2014. Note that we used changes in the presence/absence of species in our samples as proxies for pond colonization/extinction, and that undersampling can affect our results
Discussion
In this study, we explored how temporal changes in land cover and local environmental variables, past species assemblages, explained the current species assemblages in urban ponds. This is one of the few studies that focuses on urban environments and analyses on how temporal variation in predictor variables predicts current species diversity. We found that only previous species richness significantly influenced current species richness. However, none of the three predictors affected temporal beta diversity. These results suggest that within the time frame and spatial context examined here, urban planners can make somewhat reliable predictions on species richness changes over a short time frame. However, it will be difficult to predict the species composition variation.
Our results showed that spatial and temporal variation of both species richness and temporal beta diversity were similar. This result fits with the idea of Soininen (2010) who suggested that turnover in space and time is driven by similar intrinsic (e.g. body size) and extrinsic (e.g. pond size) factors. We also found that the temporal and the spatial beta diversity was high and that the turnover and nestedness presented similar contributions to the total beta diversity considering the spatial or temporal scale. Similar results have been found in other wetland studies. In a study on rural coastal wetland fish and macroinvertebrate communities, Langer et al. (2016) found that spatial and temporal beta diversity had high variation. Although there should be no a priori reasons for why urban and rural areas should differ in patterns of beta diversity, we note that the pattern of a similarity in turnover rate in space and time can be similar in an urban pond landscape.
Our structural equation modeling showed that 2014 species richness had a significant effect on 2019 species richness. Conversely, temporal environmental change was not linked to species richness change between 2014 and 2019. However, possible within-season variation in, for example, water chemistry can mask long-term changes and thereby lowering the power to detect links between community changes and environmental changes. Temporal beta diversity was not affected by any of the predictor variables. These results suggest that stochastic colonization and local extinctions may be driving the metacommunity structure in these urban ponds. The relatively small spatial extent of our study and the high diversity of dispersal modes of macroinvertebrates may favor the random colonization and extinction dynamics of pond communities (Bilton et al. 2001; Heino et al. 2017a; Leibold and Chase 2018). Other recent studies have also found that past species assemblage is a good predictor of species diversity (Gálvez et al. 2020), and in some of these studies it was a better predictor than the other environmental variables explored (Oliveira et al. 2020; Ortega et al. 2021). However, the predictive power of past assemblages should probably decrease with an increasing time interval between samples because of turnover events (Tanner et al. 1996). Some support for this was found in Ortega et al. (2021) where the relationship between past and current species assemblages decreased with a longer time interval between samplings. The predictive power of past species assemblages should also decrease at the initial stages of succession, which are likely to show fast turnover rates. In addition, studies have found that after an initial increase in species richness of newly established ponds, species richness levels off after a couple of years (Marchetti et al. 2010; Miguel-Chinchilla et al. 2014). All our study ponds were older than five years and can be one reason for the high correspondence between past and current biodiversity patterns.
The result that temporal changes in land cover variables did not affect temporal beta diversity corresponds to results found in birds in urban areas (Petersen et al. 2022). Our study and Petersen et al. (2022) investigated a fairly short time frame, spanning a range of five or six years. Since the majority of the macroinvertebrates in our study have a short generation time, it is interesting to note that past species richness still can predict current species richness to some extent, albeit a different species assemblage. This is also interesting considering the temporal changes in local variables, and despite these changes, spatial richness patterns were similar between 2014 and 2019.
Even though the general pattern in species richness and temporal beta diversity was not affected significantly by the majority of the predictors we used, we found some interesting species-specific patterns. First, we found that the more common species at a landscape scale likely had a higher probability of colonizing a pond compared to the less common ones. This suggests that a large part of the community assemblages observed can be related to dispersal-assembled communities, which is a result of stochastic colonization and extinction processes (Hubbell 2001). However, niche selection is probably also occurring in the ponds system as has been shown in other pond study systems (Chase 2007; Dubart et al. 2022).
Second, we found a negative relationship between species commonness and the probability of extinction. This pattern is expected from a stochastic extinction process. However, in contrast to colonization, where most of the species’ colonization probabilities were close to the 1:1 line, the relationship for extinction had some outliers. Although these outlier species (Aeshna grandis, Hydrochara caraboides, and Noterus crassicornis) were present in a few ponds in 2014, they still occurred in these ponds during the second survey year. We do not have enough understanding of the niches of these species to explain this pattern, but one potential explanation is that they are habitat specialists and thus are restricted to ponds with certain environmental characteristics.
Third, we also found that seven species had a very high extinction probability in the ponds where they were found in the first year of sampling. One explanation for this finding could be that they are habitat specialists, and when the environment changes too much they go extinct. For instance, by 2019, the water beetle Haliplus heydeni was no longer present in 15 of the ponds where it had been in 2014. This species feeds on filamentous algae (Holmen 1987), and perhaps such algae occur only sporadically in the ponds across the area studied. On the other hand, the species is described as a habitat generalist (Holmen 1987), suggesting that it should not be sensitive to habitat changes.
In our study, we focused on species presence at the landscape scale and did not consider species local abundance. Not all species have the same population size, nor is dispersal capacity limited solely by the number of occupied localities. Divergent patterns of for example colonization/extinction can therefore be attributed to differences in species abundance (high abundance increases the probability of dispersal) or dispersal capacity (body size and wing length). Future analysis and studies may be able to incorporate such intricacies. The local abundance of a species is also important for our ability to detect it. It should be noted that rare species at the landscape scale (few ponds occupied) can be more difficult to find because of the low number of individuals in ponds where they occur. Consequently, we were unable to distinguish between the effect of not finding the species and true absences for species with few individuals. Hence, our results regarding processes of colonization and extinction need to be interpreted cautiously.
Our study highlights the need for temporal studies focusing on changes in species assemblages as well as changes in environmental variables in an urban pond landscape. The patterns we found here might also be found in other urban green and blue areas, such as garden ponds, parks or gardens. It would be valuable for city planners to know at which time interval the prediction of past species assemblages can be used to predict future species assemblages, and to what extent land-cover change has minimal impact on species assemblages as found here. Future long-term biodiversity studies in urban areas can provide such knowledge. Additionally, our study shows that urban ponds are highly dynamic in space and time, arguing for the preservation of a large number of ponds to maintain landscape-level species richness and the underlying processes.
Data availability
Macroinvertebrate and pond environmental data, and R script are available at Zenodo https://doi.org/10.5281/zenodo.10215976 (Granath et al. 2023). Land cover data from Copernicus can be found at https://land.copernicus.eu/pan-european/high-resolution-layers under expert products.
References
Anderson MJ, Crist TO, Chase JM, Vellend M, Inouye BD, Freestone AL, Sanders NJ, Cornell HV, Comita LS, Davies KF, Harrison SP, Kraft NJ, Stegen JC, Swenson NG (2011) navigating the multiple meanings of β diversity: a roadmap for the practicing ecologist. Ecol Lett 14:19–28
Aronson MFJ, La sorte FA, Nilon CH et al (2014) A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc R Soc 281:20133330. https://doi.org/10.1098/rspb.2013.3330
Baselga A (2010) Partitioning the turnover and nestedness components of beta diversity. Glob Ecol Biogeogr 19:134–143
Baselga A, Orme D, Villeger S, De Bortoli J, Leprieur F, Logez M, Martinez-Santalla S, Martin-Devasa R, Gomez-Rodriguez C, Crujeiras R (2023) Betapart: partitioning beta diversity into turnover and nestedness components. R package version 1.6. https://CRAN.R-project.org/package=betapart
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting Linear mixed-effects models using lme4. J Stat Softw 67:1–48. https://doi.org/10.18637/jss.v067.i01
Beninde J, Veith M, Hochkirch A (2015) Biodiversity in cities needs space: a meta-analysis of factors determining intra-urban biodiversity variation. Ecol Lett 18:581–592. https://doi.org/10.1111/ele.12427
Biggs J, Williams P, Whitfield M, Nicolet P, Weatherby A (2005) 15 years of pond assessment in Britain: results and lessons learned from the work of pond conservation. Aquat Conserv Mar Freshw Ecosyst 15:693–714
Bilton DT, Freeland JR, Okamura B (2001) Dispersal in Freshwater invertebrates. Ann Rev Ecol Syst 32:159–181. https://doi.org/10.1146/annurev.ecolsys.32.081501.114016
Bispo PC, Balzter H, Malhi Y, Slik JWF, Santos JR, Rennó CD et al (2017) Drivers of metacommunity structure diverge for common and rare amazonian tree species. PLoS ONE 12:e0188300. https://doi.org/10.1371/journal.pone.0188300
Blicharska M, Andersson J, Bergsten J, Bjelke U, Hilding-Rydevik T, Johansson F (2016) Effects of management intensity, function and vegetation on the biodiversity in urban pond. Urban for Urban Green 20:103–112
Blicharska M, Andersson J, Bergsten J, Bjelke U, Hilding-Rydevik T, Thompson M, Östh J, Johansson F (2017) Is there a relationship between socio-economic factors and biodiversity in urban ponds? A study in the city of Stockholm. Urban Ecosyst 20:1209–1220
Blicharska M, Johansson F (2016) Urban ponds for people and by people. In: Francis R, Millington J, Chadwick MA (eds) Urban landscape ecology: science, policy and practice. Routledge, pp 164–180
Cardoso P, Rigal F, Carvalho JC, Fortelius M, Borges PAV, Podani J, Schmera D (2014) Partitioning taxon, phylogenetic and functional beta diversity into replacement and richness difference components. J Biogeogr 41:749–761
Chao A, Gotelli NJ, Hsieh TC et al (2014) Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol Monogr 84:45–67
Chase JM (2007) Drought mediates the importance of stochastic community assembly PNAS 104(44):17430–17434. https://doi.org/10.1073/pnas.0704350104
Chase JM, Myers JA (2011) Disentangling the importance of ecological niches from stochastic processes across scales. Phil Trans R Soc B 366:2351–2363. https://doi.org/10.1098/rstb.2011.0063
Copernicus Land Monitoring Service (CLMS) (2023). https://land.copernicus.eu/
Dray S, Bauman DS, Blanchet D, Borcard G et al (2022) adespatial: Multivariate Multiscale Spatial Analysis. R package version 0.3–19
Dubart M, Pointier JP, Jarne P, David P (2022) Niche filtering, competition and species turnover in a metacommunity of freshwater molluscs. Oikos 9:e09157. https://doi.org/10.1111/oik.09157
Garrett JK, White MP, Huang J et al (2019) Urban blue space and health and wellbeing in Hong Kong: results from a survey of older adults. Health Place 55:100–110. https://doi.org/10.1016/j.healthplace.2018.11.003
Gálvez Á, Aguilar-Alberola JA, Armengol X, Bonilla F, Iepure S, Monrós JS, Olmo C et al (2020) Environment and Space Rule, but Time also matters for the Organization of Tropical Pond Metacommunities. Front Ecol Evol 8:558833
Granath G, Hyseni C, Bini LM, Heino J, Ortega JCG, Johansson F (2023) Data: disentangling drivers of temporal changes in urban pond macroinvertebrate diversity. Zenodo. https://doi.org/10.5281/zenodo.10215976
Hamer AJ, Parris KM (2011) Local and landscape determinants of amphibian communities in urban ponds. Ecol Appl 21:378–390
Hassall C (2014) The ecology and biodiversity of urban ponds. Wiley Interdiscip Rev Water 1:187–206. https://doi.org/10.1002/wat2.1014
Hassall C, Hollinshead J, Hull A (2011) Environmental correlates of plant and invertebrate species richness in ponds. Biodivers Conserv 20:3189–3222
Heino J (2010) Are indicator groups and cross-taxon congruence useful for predicting biodiversity in aquatic ecosystems? Ecol Ind 10:112–117
Heino J (2013) Does dispersal ability affect the relative importance of environmental control and spatial structuring of littoral macroinvertebrate communities? Oecologia 171:971–980. https://doi.org/10.1007/s00442-012-2451-4
Heino J, Alahuhta J, Ala-Hulkko T, Antikainen H, Bini LM, Bonada N, Datry T, Erős T, Hjort J, Kotavaara O, Melo AS, Soininen J (2017a) Integrating dispersal proxies in ecological and environmental research in the freshwater realm. Environ Rev 25:334–334
Heino J, Bini LM, Andersson J, Bergsten J, Bjelke U, Johansson F (2017b) Unravelling the correlates of species richness and ecological uniqueness in a metacommunity of urban pond insects. Ecol Ind 73:422–431
Heino J, Melo AS, Siqueira T, Soininen J, Valanko S, Bini LM (2015) Metacommunity organisation, spatial extent and dispersal in aquatic systems: patterns, processes and prospects. Freshw Biol 60:845–869
Hill MJ, Biggs J, Thornhill I, Briers RA, Gledhill DG, White JC, Wood PJ, Hassall C (2017a) Urban ponds as an aquatic biodiversity resource in modified landscapes. Glob Change Biol 23:986–999
Hill MJ, Heino J, Thornhill I, Ryves DB, Wood PJ (2017b) Effects of dispersal mode on the environmental and spatial correlates of nestedness and species turnover in pond communities. Oikos 126:1575–1585
Hill MJ, Heino J, White JC, Ryves DB, Wood PJ (2019) Environmental factors are primary determinants of different facets of pond macroinvertebrate alpha and beta diversity in a human-modified landscape. Biol Conserv 237:348–357
Holmen M (1987) The aquatic Adephaga (Coleopter) of Fennoscandia and Denmark. I. Gyrinidae, Haliplidae, Hygrobiidae and Noteridae. Fauna Ent Scand 20:1–168
Hsieh TC, Ma KH, Chao A (2020) iNEXT: iNterpolation and EXTrapolation for species diversity. R package version 3.0.0
Hubbell SP (2001) The Unified Neutral Theory of Biodiversity and Biogeography. Princeton University Press
Hyseni C, Heino J, Bini LM, Bjelke U, Johansson F (2021) The importance of blue and green landscape connectivity for biodiversity in urban ponds. Basic Appl Ecol 57:129–145
Jabot F, Laroche F, Massol F et al (2020) Assessing metacommunity processes through signatures in spatiotemporal turnover of community composition. Ecol Lett 23:1330–1339
Jim CY, Cheng WY (2006) Recreation–amenity use and contingent valuation of urban greenspaces in Guangzhou, China. Landsc Urban Plan 75:81–96
Jimenez MF, Pejchar L, Reed SE, McHale MR (2022) The efficacy of urban habitat enhancement programs for conserving native plants and human-sensitive animals. Landsc Urban Plan 220:104356
Kuznetsova A, Brockhoff PB, Christensen RHB (2017) lmerTest package. Tests in linear mixed effects models. J Stat Softw 82:1–26
Langer TA, Murray BA, Pangle KL, Uzarski DG (2016) Species turnover drives beta-diversity patterns across multiple spatial and temporal scales in Great Lake Coastal Wetland Communities. Hydrobiologia 777:55–66
Lefcheck JS (2016) piecewiseSEM: Piecewise structural equation modeling in R for ecology, evolution, and systematics. Methods Ecol Evol 7:573–579
Legendre P (2019) A temporal beta-diversity index to identify sites that have changed in exceptional ways in space–time surveys. Ecol Evol 9:3500–3514
Leibold MA, Chase JM (2018) Metacommunity Ecology. Princeton University Press, Princeton
Leibold MA, Holyoak M, Mouquet N, Amarasekare P, Chase JM, Hoopes MF, Holt RD, Shurin JB, Law R, Tilman D, Loreau M, Gonzalez A (2004) The metacommunity concept: a framework for multi-scale community ecology. Ecol Lett 7:601–613. https://doi.org/10.1111/j.1461-0248.2004.00608.x
Lindholm M, Alahuhta J, Heino J, Toivonen H (2020) No biotic homogenisation across decades but consistent effects of landscape position and pH on macrophyte communities in boreal lakes. Ecography 43:294–305
Lindholm M, Alahuhta J, Heino J, Toivonen H (2021) Temporal beta diversity of lake plants is determined by concomitant changes in environmental factors across decades. J Ecol 109:819–832
Liu Z, Zhou T, Heino J, Castro DMP, Cui Y, Li Z, Wang W, Chen Y, Xie Z (2022) Land conversion induced by urbanization leads to taxonomic and functional homogenization of a river macroinvertebrate metacommunity. Sci Total Environ 825:153940
Maragno D, Gaglio M, Robbi M, Appiotti F, Fano EA, Gissi E (2018) Fine-scale analysis of urban flooding reduction from green infrastructure: an ecosystem services approach for the management of water flows. Ecol Model 386:1–10. https://doi.org/10.1016/j.ecolmodel.2018.08.002
Marchetti MP, Garr M, Smith ANH (2010) Evaluating wetland restoration success using aquatic macroinvertebrate assemblages in the Sacramento Valley, California. Restor Ecol 18:457–466
Miguel-Chinchilla L, Boix D, Gascón S, Comín FA (2014) Macroinvertebrate biodiversity patterns during primary succession in man made ponds in north-eastern Spain. J Limnol 73:428–440
Nava-Díaz R, Zuria I, Pineda-López R (2022) Taxonomic, phylogenetic and functional diversity of bird assemblages in urban green spaces: null model analyses, temporal variation and ecological drivers. Front Ecol Evol 9:795913
Oertli B, Parris KM (2019) Review: toward management of urban ponds for freshwater biodiversity. Ecosphere 10:e02810. https://doi.org/10.1002/ecs2.2810
Oliveira FJM, Lima-Junior DP, Bini LM (2020) Current environmental conditions are weak predictors of fish community structure compared to community structure of the previous year. Aquat Ecol 54:729–740. https://doi.org/10.1007/s10452-020-09771-z
Ortega JCG, Geijer J, Bergsten J, Heino J, Herrmann J, Johansson F, Bini LM (2021) Spatio-temporal variation in water beetle assemblages across temperate freshwater ecosystems. Sci Total Env 792:148071. https://doi.org/10.1016/j.scitotenv.2021.148071
Paradis E, Schliep K (2019) Ape 5.0: an environment for modern phylogenetics and evolutionary analyses in R. Bioinformatics 35:526–528
Perron MA, Richmond IC, Pick FR (2021) Plants, water quality and land cover as drivers of Odonata assemblages in urban ponds. Sci Total Environ 773:145467
Petersen TK, Speed JD, Grøtan V, Frøyen YK, Austrheim G (2022) Urbanisation and land-cover change affect functional, but not compositional turnover of bird communities. Urban Ecosyst 25:1679–1698
Planchuelo G, von der Kowarik I, von der Lippe M (2020) Endangered plants in novel urban ecosystems are filtered by strategy type and dispersal syndrome, not by spatial dependence on natural remnants. Front Ecol Evol Urban Ecology. https://doi.org/10.3389/fevo.2020.00018
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ruhí A, Herrmann J, Gascón S, Sala J, Geijer J, Boix D (2012) Change in biological traits and community structure of macroinvertebrates through primary succession in a man-made Swedish wetland. Freshw Sci 31:22–37. https://doi.org/10.1899/11-018.1
Shipley B (2016) Cause and correlation in biology: a user’s guide to path analysis, structural equations and causal inference with R. Cambridge university press
Simpson G (2022) permute: Functions for Generating Restricted Permutations of Data. R package version 0.9-7
Siqueira T, Bini LM, Roque FO, Marques Couceiro SR, Trivinho-Strixino S, Cottenie K (2012) Common and rare species respond to similar niche processes in macroinvertebrate metacommunities. Ecography 35:183–192
Smith N, Georgiou M, King AC, Tieges Z, Webb S, Chastin S (2021) Urban blue spaces and human health: a systematic review and meta-analysis of quantitative studies. Cities 119:103413. https://doi.org/10.1016/j.cities.2021.103413
Sánchez-Fernándes D, Abellán P, Mellado A, Velasco J, Millán A (2006) Are water beetles good indicators of biodiversity in Mediterranean aquatic systems? The case of the Segura river basin (Spain). Biodivers Conserv 15:4507–4520
Socolar JB, Gilroy JJ, Kunin WE, Edwards DP (2016) How should beta-diversity inform biodiversity conservation? Trends Ecol Evol 31:67–80
Soininen J (2010) Species turnover along abiotic and biotic gradients: patterns in space and time? Bioscience 60:433–439
Tanner JE, Hughes TP, Connell JH (1996) The role of history in community dynamics: a modelling approach. Ecology 77:108–177. https://doi.org/10.2307/2265660
Thornhill I, Batty L, Death RG, Friberg NR, Ledger ME (2017) Local and landscape scale determinants of macroinvertebrate assemblages and their conservation value in ponds across an urban land-use gradient. Biodivers Conserv 26:1065–1086
United Nations (2018) The World’s Cities in 2018 Data Booklet (ST/ESA/SER.A/417). In Department of Economic and Social Affairs, Population Division. https://www.un.org/en/events/citiesday/assets/pdf/the_worlds_cities_in_2018_data_booklet.pdf
Wang Y, Naumann U, Eddelbuettel D, Wilshire J, Warton D (2022) mvabund: Statistical Methods for Analysing Multivariate Abundance Data. R package version 4.2.1
Warton DI (2008) Penalized normal likelihood and ridge regularization of correlation and covariance matrices. J Am Stat Assoc 103:340–349
Westgate MJ, Barton PS, Lane WP, Lindenmayer DB (2014) Globalmeta-analysis reveals low consistency of biodiversity congruence relationships. Nat Commun 5:3899
Yee TW (2010) The VGAM Package for Categorical Data Analysis. J Stat Softw 32:1–34. https://doi.org/10.18637/jss.v032.i10
Acknowledgements
This work was supported by grant 201800538 from FORMAS to F. Johansson. We thank Ruth Hobro, Jan Pröjts, Jonas Roth, Hans Erik Wanntorp, for help with species determination. We would also like to thank Björn Almqvist, Arvid de Jong, August Lundholm, and Roberta Hedberg for help with sampling of ponds.
Funding
This work was supported by FORMAS (grant no. 201800538) to F. Johansson.
Open access funding provided by Uppsala University.
Author information
Authors and Affiliations
Contributions
Conceptualisation: FJ, GG, CH; Analyses: GG; Writing first draft: GG, FJ; Writing and editing: GG, FJ, CH, LMB, JH, JCGO. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Granath, G., Hyseni, C., Bini, L.M. et al. Disentangling drivers of temporal changes in urban pond macroinvertebrate diversity. Urban Ecosyst (2024). https://doi.org/10.1007/s11252-023-01500-2
Accepted:
Published:
DOI: https://doi.org/10.1007/s11252-023-01500-2