
Abstract
Background
Fires are a natural occurrence in African savannas yet little is known about how fires affect soil hydrological properties over time. This study aimed to determine the short-term (1–9 months post fire) and long-term (> seven decades fire exclusion) impact of experimental fires on soil hydrology in a fire-prone African savanna. This study also investigated how soil hydrological properties may vary under different vegetation physiognomies.
Methods
Saturated hydraulic conductivity (K) was determined using a modified falling head method through macropores (KMacro) while unsaturated K was measured using a tension disc infiltrometer (Ψ = 30 mm) to reflect the meso- and micro-pore conductivity (KMeso+Micro). Soil hydrophobicity was measured using the water droplet penetration time (WDPT) test.
Results
For the finer soil pores, fires significantly reduce KMeso+Micro in the short-term one month post-burn but returns to Pre-burn KMeso+Micro rates nine months later. After decades of fire suppression, KMacro is significantly faster compared to burned soils while KMeso+Micro was significantly slower through smaller pores compared to soils burned annually. KMeso+Micro is slowest under shrubs in fire suppression plots compared to other vegetation physiognomies. Fires did not induce hydrophobicity in the short-term nor the long-term.
Conclusion
Whilst fire reduces KMeso+Micro in the short-term, soils recovered nine months later, indicating that prescribed burning does not lead to lasting detrimental impacts on soil hydrological properties. Interestingly, decades of active fire suppression had either a positive or negative effect on the soil’s ability to allow water to infiltrate through the soil surface depending on the pore size and possible susceptibility to bioclogging and biocrusts.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Fires are an important natural and widespread phenomenon in many ecosystems globally. It is a critical driver in fire-prone landscapes such as African savannas, where it maintains healthy and functional ecosystems (Van Wilgen et al. 2000; Bond and Keeley 2005). Fires may induce short-term, long-term or permanent changes to soil properties depending on the fire intensity and frequency, fire type (wildfire versus prescribed fire), vegetation type as well as soil conditions (Doerr and Cerdá 2005; Certini 2005). In light of the growing practice of prescribed burning around the world for the reduction in wildfire risk (Furlaud et al. 2018), for biodiversity conservation and ecosystem services (Bond 2005) and for experimental reasons (Stoof et al. 2011), there is an increasing need to study the effects of prescribed burning and their varying fire intensities and frequencies on soil hydrological properties.
Due to over-utilization and poor land management practises world-wide, soils are under threat which has led to widespread soil degradation (Bone et al. 2010; Eijsackers et al. 2014; Gura and Mnkeni 2019). Since soils are pivotal in maintaining functional ecosystems and it is a non-renewable resource on human time scale (Jenny 1980), it is important to investigate how prescribed fires may affect soil properties in fire-prone landscapes. Considering that Africa is dubbed ‘The Fire Continent’ due to the extent of fire occurrence across the continent (Goldammer and Mutch 2001; Archibald et al. 2010), and that the majority of South Africa’s land surface falls within a fire-prone biome, it is particularly relevant that the impact of burning on soil hydrological properties over time are determined. The reduced capacity of soils to allow for water penetration and infiltration could prove problematic in a landscape in which extreme rainfall events are common or where it may increase under future conditions. When rainfall intensity exceeds the ability of the soil to allow for infiltration, overland flow and surface runoff occurs (Sidle et al. 2007), leading to increased risk of flooding and/or erosion. As such, some studies have previously raised concerns about fires accelerating soil erosion and land degradation in South Africa (Meadows and Hoffman 2002; Strydom and Savage 2016).
In other fire-prone ecosystems such as the coniferous forests in North America and Europe, Mediterranean woodlands, and Australian eucalyptus forests, prescribed fires affected soil hydrology, increased hydrophobicity, decreased soil stability and increased erosion which has a cascading impact on ecosystem resilience (Robichaud 2000; Doerr et al. 2004; Fonseca et al. 2017; Thomaz 2018). A reduction in infiltration capacity may be attributed to changes in macropore properties and/ or fire-induced hydrophobicity (or soil water repellency) which has also been documented in Eucalypt forests in Australia (Doerr et al. 2004; Nyman et al. 2010, 2014), Eucalypt plantations in Portugal (Keizer et al. 2008), and Mediterranean forests in Spain (Doerr and Cerdá 2005). As a result of the extreme heat generated during burning, and the potential release of water-repellent compounds from burning vegetation coating soil particles, hydrophobicity can increase soil erodibility which may also be aided by soil texture whereby coarser soils are more susceptible to fire-induced hydrophobicity (Ravi et al. 2006).
Vegetation cover and soil properties have variable effects on soil infiltration rates. Soil infiltration is a function of texture, soil porosity and structure, which is facilitated by biological activities (Cerdà and Robichaud 2009). Some studies propose that slower infiltration rates may be due to vegetation removal by fire rendering soil surfaces vulnerable to direct sunlight and other process such as raindrop impact and splash erosion (DeBano 2000; Assouline 2004; Cerdà and Robichaud 2009; Larsen et al. 2009; Jordán et al. 2016). Vegetation leads to a soil litter layer which promotes bioactivity, soil aggregation, water storage, and macro- and micro-pore development (Cerda and Robichaud, 2009).
In South African semi-arid savannas, it is unclear how regular burning impacts a soil’s ability to allow water to enter and move through the soil profile. Research on similar sandy, coarse-grained soils by Magomani and van Tol (2019) found that prescribed annual burning decreased saturated hydraulic conductivity and water conducting macroporosity compared to unburned soils but soils burned every 3 years had greater saturated hydraulic conductivity and water conducting macroporosity. A laboratory experiment by Mills and Fey (2004) found that soils burned annually with prescribed burning had slower infiltration rates than unburned soils in Kruger National Park (KNP), South Africa. However, Riddell et al. (2012) found that unsaturated hydraulic conductivity on annually-burned granitic soils in KNP was greater than on long-term unburned soils which they attribute to the loss of organic residue on soil particles due to regular annual prescribed burning. None of these aforementioned studies quantified these prescribed fire-induced impacts on soil hydrology in relation to time since last fire. Therefore, it is unclear whether fire effects are immediate or for how long they persist post-fire.
The work presented here aimed at investigating the short-term effect of fire (directly after and nine months after a prescribed fire) and long-term effect of fire exclusion (seven decades of fire suppression) on soil hydrological properties such as soil hydraulic conductivity (K) and hydrophobicity in granitic soils of southern Kruger National Park, South Africa. Considering the indirect changes to vegetation structure following decades of fire exclusion, and the possible interactive effects of fire and plant physiognomies, where possible we also investigated how these soil hydrological properties may differ under large trees (> 10 m), under shrubs (3–5 m) and in open, grassy areas on annual burn plots and decadal fire exclusion plots. Our hypotheses are that (1) prescribed fires will reduce soil K in the short-term (1 month post fire) but will recover to similar soil K rates observed in pre-burn conditions in less than one year post-fire due to ash dispersal, ash degradation and vegetation regrowth, (2) that soils under trees will have faster soil K rates than open, grassy areas due to soils below canopies being protected against the elements (direct sunlight, raindrop impact) after being denuded by fire, and (3) that burning will induce soil hydrophobicity in the short-term due to direct soil heating after a prescribed fire and altering the chemical and physical structure of the soil.
Materials and methods
Study Area and Experimental Design
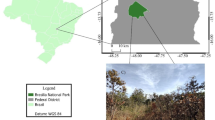
The study area is situated within the Kruger National Park (KNP) in north-eastern South Africa (-25.127° S; 31.389° E), as shown in Fig. 1. This large conservation area covers approximately 20 000 km2 and is located within the savanna biome. This study was conducted in the southern section of the park, characterized by a wooded savanna dominated by Marula (Sclerocarya birrea) trees, along with shrubs such as Red bushwillow (Combretum apiculatum), and Silver cluster leaf (Terminalia sericea). Between the woody vegetation, an expansive grass layer exists primarily dominated by Erogrostis rigidior, Urochloa mosambicensis and Panicum maximum. The region experiences a long-term average annual rainfall of approximately 550 mm, with the majority of precipitation occurring during the wet season, from December to March. Climate is typically characterized by hot, humid summer with high temperatures (average daily maximum temperature of 34℃ and milder winters with maximum daily temperatures of approximately 27℃ (Kennedy and Potgieter 2003).

Map of KNP with Skukuza study site circled in black (top). An example showing differences in vegetation (bottom) on the Annual burn (Aug1) and No Burn (Fire exclusion) experimental plots (Smit et al. 2010)
Fire is recognized as an ecosystem driver and consequently used as a management tool in KNP in order to promote biodiversity as well as for safety reasons around infrastructure. For nearly 70 years, KNP has maintained one of the world’s largest and longest-running fire experiments. This experiment began in 1954 in order to evaluate the impact of different fire frequencies and season of burn on savanna vegetation dynamics (Trollope et al. 1998; Biggs et al. 2003). These experimental burn plots are roughly 7 ha each (450 × 150 m) and primarily located on the crests. In the Skukuza section of the park, topography is characterized by gentle undulating hillslopes with majority classified as having a gradient = 0–6%. Around Skukuza, there are four burn string replicates which are underlain by granite-derived sandy soils belonging to the Hutton, Clovelly, and Glenrosa soil forms (Venter and Govender 2012). These South African soil taxonomic classifications can be related to the World Reference Base as Chromic Cambisols, Haplic Cambisols and Cutanic Leptosols for Hutton, Clovelly and Glenrosa, respectively (van Huyssteen 2020). For the purposes of this study, we focused on the No Burn plots, which remained unburned for nearly 70 years, and the Annual plots, which were subjected to burning annually in August, in each of the four replicates.
Vegetation and experimental fire characteristics
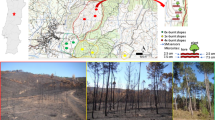
For each burn plot, all aboveground herbaceous biomass within 0.5 m x 0.5 m quadrats were clipped (n = 9 per plot). Samples were placed in a drying oven at 65 ℃ for 72 h and dry mass used to estimate grass fuel loads. Byram’s (1959) fireline intensity was estimated for four experimental fires applied in August 2021 taking into account grass fuel load, fire rate of spread and heat yield (constant). Weather parameters such as air temperature, relative humidity and wind speed was measured before burning commenced.
Soil sampling and analyses
Undisturbed soil cores (0–5 cm) were collected across the Annual and No Burn plots (n = 24) and analysed for bulk density (Blake and Hartge 1986). Soil samples (0–10 cm) were analysed for soil pH (KCl; n = 120) with a pH meter and soil particle size distribution was measured (n = 120) using the hydrometer method (Bouyoucos 1962). Soil fractions (i.e. sand, silt and clay) were used to classify soils according to the U.S. Department of Agriculture (USDA) soil texture classification system (USDA 1951). Total C was measured at 0–10 cm soil depths (n = 120) using a LECO CNS TruSpec Analyser (LECO Corporation, USA) on the Annual burn (before burning) and No Burn plots (data was sourced from published literature by Strydom et al. 2024).
Saturated K was measured in-situ using a modified, falling head method (Bouwer and Rice 1976; van Tol et al. 2012):
whereby L = thickness of the soil horizon; t = time until infiltration completed; and h0 and h1 = head of water above the soil surface before and after the test, respectively.
Unsaturated hydraulic conductivity (K) was measured in-situ at the soil surface using a tension disc infiltrometer under three different tensions, i.e. Ψ = 5 mm, 30 mm and 60 mm but only Ψ = 30 mm was used as described below. This custom-designed tension infiltrometer had a disc diameter of 8.4 cm (Ankeny et al. 1991). Unsaturated K measurements were conducted in-situ under large trees (Sclerocarya birrea), under shrubs (Combretum apiculatum) and in open, grassy areas until steady state was reached. Saturated K was measured with a double-ring infiltrometer only under large trees and in open, grassy areas due to impracticalities of the method used to measure saturated K, i.e. the equipment was too large to fit under a shrub. Both the saturated and unsaturated K measurements were conducted in-situ and on the soil surface which is exposed to burning at random locations within the burn plots. Macropore water conductivity was calculated by using the difference between saturated K and unsaturated K at Ψ = 30 mm (Van Tol et al. 2012). Thus, macropore conductivity accounts for water flowing through pores with a diameter of > 1 mm (Luxmoore 1981) and hereafter referred to as KMacro. Unsaturated K at Ψ = 30 mm accounts for water conductivity through pores with a diameter of < 1 mm (Van Tol et al. 2012; Patra et al. 2022) and hereafter will be referred to as KMeso+Micro.
To detect a short-term impact of fire on infiltration rates, KMacro was measured before a fire, one month and nine months post-fire (6 samples per plot X 3 time since fire treatments X 4 replicates; n = 72). Similarly, the short-term effect of burning on KMeso+Micro was assessed (9 samples per plot X 2 fire treatments X 4 replicates; n = 72). The long-term fire impact was assessed by measuring KMacro on soils burned every year (on Annual Burn plot ~ 10 months after previous fire) and soils on a plot where fires have been excluded (No Burn plot) for nearly 70 years (6 samples per plot X 2 fire treatments X 4 replicates; n = 48). The same was conducted to assess the long-term effect of burning on KMeso+Micro (9 samples per plot X 2 fire treatments X 4 replicates; n = 72). To determine the effect of varying vegetation physiognomy on soil infiltration, KMacro was measured beneath trees canopies and open, grassy areas (3 samples per physiognomy type X 2 physiognomy types X 2 fire treatments X 4 replicates; n = 48). KMeso+Micro was measured beneath tree canopies, shrubs and in open, grassy areas (3 samples per physiognomy type X 3 physiognomy types X 2 fire treatments X 4 replicates; n = 72).
Hydrophobicity was assessed in situ using the Water Droplet Penetration Time (WDPT) test directly at the soil surface at randomly-selected locations in the plots (Letey 1969). To investigate the short-term impact of burning, WDPT tests were conducted a few minutes before an experimental burn (Pre-burn) compared to measurements immediately after the burn (Post-burn) (5 droplets per sample point X 9 sample points per plot X 2 fire treatments X 3 replicates; n = 270). To examine the effect of long-term annual burning in comparison to decadal fire suppression, WDPT tests were conducted on the Annual Burn plot (~ 10 months since the previous fire) and No Burn plot (5 droplets per sample point X 9 sample points per plot X 2 fire treatments X 3 replicates; n = 270). Soils are classified as hydrophobic when WDPT is greater than five seconds (Doerr et al. 2004).
Statistical analyses
All statistical analyses were performed in R (R Core Team 2022) using the ggplot2, FSA, tidyverse, rstatix, conover.test, and fansi packages. The normality of the data was assessed using the shapiro.test (p ≥ 0.05). If data were not normally distributed, a log-transformation was attempted. To assess the effect of time since last fire occurrence and fire exclusion on soil KMacro and KMeso+Micro, log-transformation did not yield normality, leading to the use of a non-parametric kruskal.test (p < 0.05). If significant differences were detected, the multiple pair-wise comparison conover.test with the Benjamini-Hochberg adjustment was employed for post-hoc analysis (Conover and Iman 1981; Benjamini and Hochberg 1995; Dinno and Dinno 2017). The effect of vegetation (under trees vs. open, grassy areas) on KMacro was tested using a paired, non-parametric wilcox.test. The kruskal.test (p < 0.05) was applied to test for significant differences in KMeso+Micro in soils under trees, under shrubs and in open, grassy areas. If significant differences were observed, conover.test was also used to identify where those differences were detected. Short-term and long-term changes in hydrophobicity were analysed using a paired non-parametric wilcox.test.
Results
Vegetation parameters, fire characteristics and soil properties
Characteristics of the experimental fire conducted in August 2021, vegetation parameters and soil properties are presented in Table 1. As expected, grass fuel loads are greatest on the No Burn plots where fires have been excluded for nearly 70 years. Fire intensity was greatest on the Mbiyamiti replicate which could be attributed to the higher wind speeds facilitating a faster rate of fire spread. Results from the soil texture analysis classified all soils as Loamy Sand with > 80% sand content and soil pH ranged from 5.1 to 5.7. Total C on annually burned soils were consistently lower than their neighbouring fire suppressed (No Burn) plots.
Time since fire on macropore (KMacro) and, meso- and micro-pore (KMeso+Micro) hydraulic conductivity
KMacro was compared across soils following different durations since last fire occurrence (Fig. 2a). The non-parametric Kruskal-Wallis test was conducted and found a significant difference in KMacro among the soils (H (3) = 15.148, p < 0.01). A multiple pair-wise comparison Conover-Iman test revealed that while there was no statistically significant decrease in KMacro in the short-term (pre-burn, one month postburn and nine months post burn; p > 0.05), fire exclusion on the No Burn plot led to significantly faster KMacro (p < 0.01).
Soil KMeso+Micro rates were compared across various time since last fire occurrence (Fig. 2b). A non-parametric Kruskal-Wallis test found a significant difference in KMeso+Micro among the soils (H (3) = 11.959, p < 0.01). A subsequent multiple pair-wise post hoc test Conover-Iman test was employed to further explore these differences. The results revealed statistically significant differences in KMeso+Micro between pre-burn soils and one month post-burn (p < 0.05) alluding to a short-term immediate fire impact. Nine months post-burn, KMeso+Micro increases to rates similar to the pre-burn measurements (p > 0.05) but is statistically different to the KMeso+Micro measured one month post-burn (p < 0.01). This illustrates that while there is an immediate reduction in KMeso+Micro one month after a prescribed fire, within nine months KMeso+Micro recovers to rates observed before the fire. The results also suggested a long-term fire effect on soil KMeso+Micro due to statistically significant differences (p < 0.01) found between soils burned annually (Pre-burn and nine months post burn) and soils where fire have been excluded for nearly 70 years (No Burn).

Boxplots showing the effect of time since last fire and fire exclusion on KMacro (a) and KMeso+Micro (b). Median and mean values denoted by the horizontal line and solid square respectively, with first and third quartiles represented by the box and vertical lines depicting the range. Statistically significant differences denoted as * (p < 0.05) and ** (p < 0.01). 1 m − 1 month, 9 m − 9 months
Vegetation effect on meso- and micro-pore (KMeso+Micro) hydraulic conductivity
The effect of different vegetation physiognomic types on macro pore (KMacro), and meso- and micro-pore (KMeso+Micro) hydraulic conductivity was assessed as illustrated in Fig. 3. If not accounting for fire treatment, a non-parametric paired Wilcoxon signed-rank test found no significant difference KMacro between soils under large trees and soils in open, grassy areas (p > 0.05). However, when looking at the combined interaction between vegetation and prescribed fire, a Kruskal-Wallis test found a significant difference (H (3) = 8.8623, p < 0.05). A Conover-Iman test revealed significant differences between open areas on Burn vs. open areas on No Burn plots, under trees on Burn vs. open areas on No Burn plots, and between soils under trees on Burn vs. trees on No Burn plots (see Fig. 3 (a)). A Kruskal-Wallis test found a significant difference in KMeso+Micro among soils beneath different vegetation structures (H (2) = 6.449, p < 0.05). Subsequently, a multiple pair-wise comparison Conover-Iman test was conducted to further investigate these observed differences. KMeso+Micro was always lower under the No Burn treatment compared to the Burn treatments, but this was only statistically significant below shrubs on the No Burn plot, compared to soils measured under shrubs, under trees and in open areas on the Annual Burn plot (p < 0.05).

Boxplots showing the KMacro (a) and KMeso+Micro (b) across various vegetation structure types in annually burned and unburned soils. The horizontal line within each box represents the median value, with the box encompassing the first and third quartiles. Mean values are indicated by black squares. Statistically significant differences are denoted as * (p < 0.05)
Short- and long-term effect of fire on soil hydrophobicity
The effect of prescribed burning on soil hydrophobicity in both the short- and long-term was assessed by comparing the time taken for water droplets to penetrate the soil surface (Fig. 4). A non-parametric paired Wilcoxon signed-rank test found no significant difference in water droplet penetration time in the short-term between pre- and post-burn conditions (p > 0.05). However, in the long-term, a statistically significant increase in water droplet penetration time was evident in the unburned soils (although not considered as hydrophobic), where fires had been excluded for approximately 70 years (No Burn), in comparison to soils on the Annual burn plot (V = 1794; p < 0.001).

Effect of fire on water droplet penetration time, considering both short-term (pre- and immediately post-fire) and long-term effects. Bars represent the mean duration for water to penetrate the soil surface, with error bars indicating the standard error. Statistically significant differences were denoted as *** (p < 0.001)
Discussion
Prescribed fires have stronger short-term impacts on K Meso+Micro than K Macro, but only K Meso+Micro is negatively impacted by long-term fire exclusion
African savanna fires affect soil water conductivity through varying pore sizes differently. Hydraulic conductivity through larger pores spaces (KMacro) are not significantly affected by prescribed fires in the short-term. However, in the long-term following nearly 70 years of fire exclusion, KMacro is significantly faster than on burned soils. This is similar to what Riddell et al. (2012) found at the fire exclusion site in the sandier and wetter region to the south-west of KNP. We postulate that since KMacro accounts for water moving through macropore spaces, preferential pathways such as grass roots would facilitate increase KMacro on the No Burn plot. Table 1 illustrates the presence of higher grass fuel load on the No Burn than Annual burn plot. It is widely understood that vegetation roots (as well as soil fauna, e.g. termites) create preferential flow paths which influences soil hydraulic conductivity (Mitchell et al. 1995; Morales et al. 2010; Zhang et al. 2017).
Fires affect KMeso+Micro through smaller pore spaces both in the short- and long-term. Shortly after an experimental fire, a significant decrease in KMeso+Micro at the soil surface was observed (p < 0.05). The fire induced a ~ 35% reduction in KMeso+Micro one month after the burn compared to Pre-burn soils. However, nine months post-fire, soil KMeso+Micro returned to rates similar to those observed before the prescribed fire, indicating soil resilience in response to savanna fires. Immediately post-fire, we believe that KMeso+Micro is reduced due to ash deposition on the soil surface which inhibits water movement down the soil profile. Post-fire studies conducted in Pinus-dominated landscapes in North America found that ash played a significant role in covering soils surfaces and clogging preferential pathways (Onda et al. 2008; Balfour and Woods 2013; Balfour et al. 2014), which Gabet and Bookter (2011) attributed to the fine particle size of ash fragments clogging pore spaces in sandy, coarse-grained soils which is a similar soil environment in our study. Considering that KMeso+Micro refers to water moving through soil pores with diameters smaller than ~ 1 mm (Hu et al. 2009; Patra et al. 2022) and that more than 80% of ash produced from vegetation fires is smaller than 1 mm in diameter (Bodí et al. 2014), it is plausible that ash particles could clog preferential pore spaces inhibiting KMeso+Micro.
Interestingly, when fires are actively suppressed for decades, there is a long-term negative impact on KMeso+Micro. Our findings suggest that soils with long-term fire exclusion, exhibit a significantly lower KMeso+Micro (p < 0.05) compared to soils which experience annual burning. Unlike meso- and micro-pore clogging by ash particles after a fire, we believe that meso- and micro-pores are also susceptible to clogging by microbial activity; a phenomenon known as bioclogging (Cunningham et al. 1991; Morales et al. 2010). Nearly 70 years of fire exclusion may have allowed for significant microbial proliferation including that of bacteria, fungi and lichens (Belnap et al. 2001). This is further supported by higher total C observed in the No Burn plot compared to the Annual burn plot (see Table 1; Strydom et al. 2024).
Vegetation physiognomy have varying effects on K Meso+Micro
Whether burned or unburned, KMacro under trees did not differ significantly compared to open, grassy areas. However, the combined effect of fire and vegetation plays a role which may be due to the overriding effect of prescribed burning on KMacro (as shown in Fig. 3 (a). One of the most surprising findings was that KMeso+Micro under shrubs in the No Burn plot was the lowest compared to under large trees or in open, grassy areas. KMeso+Micro under shrubs in the No Burn plots were significantly slower compared to all three vegetation physiognomies (i.e. trees, shrubs and open areas) on the burned soils. Furthermore, there was no significant difference in KMeso+Micro under tree canopies or in open, grassy areas whether soils were annually burned or where fires were excluded for decades. It may be plausible that since shrub canopies are closer to ground level, the resulting microclimate and increased litter inputs create an environment that favours the development of biocrusts which could influence water movement through pore spaces less than 1 mm diameter. This is further supported by significantly greater soil C which was previously measured at the soil surface under shrubs on these long-term, unburned plots (Strydom et al. 2024). Biological crusts, also known as biocrusts, refer to cyanobacteria, lichens, fungi, algae, and mosses which live on or within the soil surface and are quite widespread across South Africa (Belnap et al. 2001; Mills and Fey 2003; Laker and Nortjé 2019). In eastern South Africa, Mills and Fey (2003) observed that these biocrusts thrive under moist soil conditions and often found below canopies. Light conditions below trees and shrubs vary depending on tree structure including height, density and diameter of overhead crown canopy (Belsky and Canham 1994). Baumgartner et al. (2015) found that the average lowest branch height for tall savanna trees such as Sclerocarya birrea ranges between 2 and 3 m whereas the lowest branch height for shrubs such as Combretum apiculatum ranges between 0.4 and 0.6 m. This variation in tree structure allows for direct light penetration onto soil surfaces to be greater under tall trees because they effectively only suppress direct sunlight in the middle of the day but are exposed during morning and afternoon periods. Therefore, since shrubs have a different growth structure to taller trees, their low-hanging canopies create ideal microsite conditions to support biological activity by reducing irradiance, enhancing soil moisture and reducing soil temperatures (O’Connor 1995; Vadigi and Ward 2013). Furthermore, tall single-stemmed trees are accessible to hooved herbivores that while grazing below tall trees may play a role in disturbing and breaking up crusts more than under less accessible areas below shrubs. Thus, considering the increase in woody vegetation in response to fire suppression, we postulate that increased presence of biocrusts in the No Burn plot may influence localized soil hydrological properties (Warren 2001; Kidron et al. 2021).
Prescribed fires do not lead to soil hydrophobicity in the short-term or long-term
Unlike what we had hypothesized, the prescribed experimental fires did not induce hydrophobicity in the short-term immediately following a fire nor in the long-term after 70 years. It is assumed that since these savanna fires have fast rates of fire spread (Govender et al. 2006), there is not enough contact time between the fire and the soil surface to allow for heat transfer which could induce hydrophobicity (Strydom et al. 2019). Even though an increase in water droplet penetration time was observed on the No Burn plot where fires have been suppressed for nearly 70 years compared to soils which are burned annually, it does not lead to hydrophobicity. According to Doerr et al. (2004), water penetration time needs to exceed 5 s before being considered as hydrophobic. Lastly, it is worth noting that timing of sampling is important. The pre- and post-burn hydrophobicity is lower than the values measured on the Annual burn plot even though it is from the same plot. It is thought that this variation may be linked to when sampling was done because pre- and post-burn measurements were done immediately before and after the experimental fire (August 2021 – mid dry season), whereas the Annual plot was measured in May 2021 at the start of the dry season. Thus, due to the dryness of the soil, adsorption may be a factor during the sampling campaign in August 2021.
Conclusion
Typical of highly complex ecosystems such as African savannas, this study highlights complex interactions between prescribed fires, vegetation dynamics and soil hydrological properties. Soils in our study site are resilient to frequent burning with short-lived effects on KMeso+Micro which recover within the first year post-fire. Long-term fire suppression leads to slower KMeso+Micro which might be attributed bioclogging within those minute pore spaces. The experimental fires did not lead to hydrophobicity in the short- nor long-term. It is recommended that future studies include further investigation to disentangle the relationship between fire, vegetation changes, biocrust proliferation and soil hydrology, and also to explore the effect of wildfires, often with different fire characteristics than prescribed fires (e.g. higher intensity) on soil hydrology. Due to the size of equipment used, it was not possible to measure KMacro under shrubs, and we recommend future studies exploring alternative methods that would allow for these measurements to be taken.
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
Ankeny MD, Ahmed M, Kaspar TC, Horton R (1991) Simple field method for determining unsaturated hydraulic conductivity. Soil Sci Soc Am J 55:467–470
Archibald S, Scholes RJ, Roy DP et al (2010) Southern African fire regimes as revealed by remote sensing. Int J Wildland Fire 19:861. https://doi.org/10.1071/WF10008
Assouline S (2004) Rainfall-induced soil surface sealing: a critical review of observations, conceptual models, and solutions. Vadose Zone J 3:570–591
Balfour VN, Doerr SH, Robichaud PR (2014) The temporal evolution of wildfire ash and implications for post-fire infiltration. Int J Wildland Fire 23:733–745
Balfour VN, Woods SW (2013) The hydrological properties and the effects of hydration on vegetative ash from the Northern Rockies, USA. CATENA 111:9–24
Baumgartner SA, Treydte AC, Grant CC, van Rooyen J (2015) Can diverse herbivore communities increase landscape heterogeneity? Comparing wild and domestic herbivore assemblages in a South African savanna. Perspectives in Plant Ecology. Evol Syst 17:34–43. https://doi.org/10.1016/j.ppees.2014.11.002
Belnap J, Büdel B, Lange OL (2001) Biological Soil crusts: characteristics and distribution. Biological Soil crusts: structure, function, and management. Springer, Berlin, Heidelberg, pp 3–30
Belsky AJ, Canham CD (1994) Forest gaps and isolated Savanna Trees. Bioscience 44:77–84. https://doi.org/10.2307/1312205
Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Stat Soc: Ser B (Methodol) 57:289–300. https://doi.org/10.1111/j.2517-6161.1995.tb02031.x
Biggs R, Biggs HC, Dunne TT et al (2003) Experimental burn plot trial in the Kruger National Park: history, experimental design and suggestions for data analysis. Koedoe 46. https://doi.org/10.4102/koedoe.v46i1.35
Blake GR, Hartge KH (1986) Bulk density. In: Klute A (ed) Methods of soil analysis, Part 1—physical and mineralogical methods, Second. American Society of Agronomy, Inc., Wisconsin, USA, pp 363–375
Bodí MB, Martin DA, Balfour VN et al (2014) Wildland fire ash: production, composition and eco-hydro-geomorphic effects. Earth Sci Rev 130:103–127
Bond WJ (2005) Large parts of the world are brown or black: a different view on the ‘Green World’hypothesis. J Veg Sci 16:261–266
Bond WJ, Keeley JE (2005) Fire as a global ‘herbivore’: the ecology and evolution of flammable ecosystems. Trends Ecol Evol 20:387–394
Bone J, Head M, Barraclough D et al (2010) Soil quality assessment under emerging regulatory requirements. Environ Int 36:609–622. https://doi.org/10.1016/j.envint.2010.04.010
Bouwer H, Rice RC (1976) A slug test for determining hydraulic conductivity of unconfined aquifers with completely or partially penetrating wells. Water Resour Res 12:423–428. https://doi.org/10.1029/WR012i003p00423
Bouyoucos GJ (1962) Hydrometer method improved for making particle size analyses of soils. Agron J 54:464–465
Byram GM (1959) Combustion of forest fuels. In: Davis KP (ed) ‘Forest fire: control and use. McGraw-Hill, New York, pp 61–89
Cerdà A, Robichaud PR (2009) Fire effects on soil infiltration. In: Haigh MJ (ed) Fire effects on soils and restoration strategies. Science Publishers, New Hampshire, U.S.A, pp 81–103
Certini G (2005) Effects of fire on properties of forest soils: a review. Oecologia 143:1–10. https://doi.org/10.1007/s00442-004-1788-8
Conover WJ, Iman RL (1981) Rank transformations as a bridge between parametric and nonparametric statistics. Am Stat 35:124–129
Cunningham AB, Characklis WG, Abedeen F, Crawford D (1991) Influence of biofilm accumulation on porous media hydrodynamics. Environ Sci Technol 25:1305–1311. https://doi.org/10.1021/es00019a013
DeBano LF (2000) The role of fire and soil heating on water repellency in wildland environments: a review. J Hydrol 231–232:195–206. https://doi.org/10.1016/S0022-1694(00)00194-3
Dinno A, Dinno MA (2017) Package ‘conover. test’. Conover-Iman test of multiple comparisons using rank sums. https://cran.r-project.org/package=conover.test
Doerr SH, Blake WH, Shakesby RA et al (2004) Heating effects on water repellency in Australian eucalypt forest soils and their value in estimating wildfire soil temperatures. Int J Wildland Fire 13:157–163
Doerr SH, Cerdá A (2005) Fire effects on soil system functioning: new insights and future challenges. Int J Wildland Fire 14:339. https://doi.org/10.1071/WF05094
Eijsackers H, Swartjes FA, van Rensburg L, Maboeta MS (2014) The need for attuned soil quality risk assessment for non-western humans and ecosystems, exemplified by mining areas in South Africa. Environ Sci Policy 44:174–180. https://doi.org/10.1016/j.envsci.2014.07.017
Fonseca F, de Figueiredo T, Nogueira C, Queirós A (2017) Effect of prescribed fire on soil properties and soil erosion in a Mediterranean mountain area. Geoderma 307:172–180
Furlaud JM, Williamson GJ, Bowman DM (2018) Simulating the effectiveness of prescribed burning at altering wildfire behaviour in Tasmania, Australia. Int J Wildland Fire 27:15–28
Gabet EJ, Bookter A (2011) Physical, chemical and hydrological properties of Ponderosa pine ash. Int J Wildland Fire 20:443–452
Goldammer JG, Mutch R (2001) Global forest fire assessment: 1990–2000. FAO, Rome
Govender N, Trollope WSW, Van Wilgen BW (2006) The effect of fire season, fire frequency, rainfall and management on fire intensity in savanna vegetation in South Africa: fire intensity in savanna. J Appl Ecol 43:748–758. https://doi.org/10.1111/j.1365-2664.2006.01184.x
Gura I, Mnkeni PNS (2019) Crop rotation and residue management effects under no till on the soil quality of a haplic cambisol in Alice, Eastern Cape, South Africa. Geoderma 337:927–934. https://doi.org/10.1016/j.geoderma.2018.10.042
Hu W, Shao M, Wang Q et al (2009) Temporal changes of soil hydraulic properties under different land uses. Geoderma 149:355–366
Jenny H (1980) Soil genesis with ecological perspectives. Springer-Verlag, New York
Jordán A, Zavala LM, Granged AJ et al (2016) Wettability of ash conditions splash erosion and runoff rates in the post-fire. Sci Total Environ 572:1261–1268
Keizer JJ, Doerr SH, Malvar MC et al (2008) Temporal variation in topsoil water repellency in two recently burnt eucalypt stands in north-central Portugal. CATENA 74:192–204. https://doi.org/10.1016/j.catena.2008.01.004
Kidron GJ, Veste M, Lichner L (2021) Biological factors impacting hydrological processes: pecularities of plants and biological soil crusts. J Hydrol Hydromech 69:357–359
Laker MC, Nortjé GP (2019) Review of existing knowledge on soil crusting in South Africa. Adv Agron 155:189–242
Larsen IJ, MacDonald LH, Brown E et al (2009) Causes of Post-fire Runoff and Erosion: Water Repellency, Cover, or Soil Sealing? Soil Sci Soc Am J 73:1393–1407. https://doi.org/10.2136/sssaj2007.0432
Letey J (1969) Measurement of contact angle, water drop penetration time, and critical surface tension. In: Proceedings of the Symposium on Water-Repellent Soils, May 1968. University of California, pp 43–47
Luxmoore RJ (1981) Micro-, meso-, and macroporosity of soil. Soil Sci Soc Am J 45(3):671
Magomani MI, van Tol JJ (2019) The impact of fire frequency on selected soil physical properties in a semi-arid Savannah Thornveld. Acta Agric Scand B — Soil Plant Sci 69:43–51. https://doi.org/10.1080/09064710.2018.1495253
Meadows ME, Hoffman MT (2002) The nature, extent and causes of land degradation in South Africa: legacy of the past, lessons for the future? Area 34:428–437. https://doi.org/10.1111/1475-4762.00100
Mills AJ, Fey MV (2003) Declining soil quality in South Africa: effects of land use on soil organic matter and surface crusting. South Afr J Sci 99(9):429–436
Mills AJ, Fey MV (2004) Frequent fires intensify soil crusting: physicochemical feedback in the pedoderm of long-term burn experiments in South Africa. Geoderma 121:45–64. https://doi.org/10.1016/j.geoderma.2003.10.004
Mitchell AR, Ellsworth TR, Meek BD (1995) Effect of root systems on preferential flow in swelling soil. Commun Soil Sci Plant Anal 26:2655–2666. https://doi.org/10.1080/00103629509369475
Morales VL, Parlange J-Y, Steenhuis TS (2010) Are preferential flow paths perpetuated by microbial activity in the soil matrix? A review. J Hydrol 393:29–36. https://doi.org/10.1016/j.jhydrol.2009.12.048
Nyman P, Sheridan G, Lane PNJ (2010) Synergistic effects of water repellency and macropore flow on the hydraulic conductivity of a burned forest soil, south-east Australia. Hydrol Process 24:2871–2887. https://doi.org/10.1002/hyp.7701
Nyman P, Sheridan GJ, Smith HG, Lane PN (2014) Modeling the effects of surface storage, macropore flow and water repellency on infiltration after wildfire. J Hydrol 513:301–313
O’Connor TG (1995) Acacia karroo invasion of grassland: environmental and biotic effects influencing seedling emergence and establishment. Oecologia 103:214–223
Onda Y, Dietrich WE, Booker F (2008) Evolution of overland flow after a severe forest fire, Point Reyes, California. CATENA 2:13–20
Patra S, Kaushal R, Singh D et al (2022) Surface soil hydraulic conductivity and macro-pore characteristics as affected by four bamboo species in North-Western Himalaya, India. Ecohydrol Hydrobiol 22:188–196
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria 55:275–286
Ravi S, D’Odorico P, Herbert B et al (2006) Enhancement of wind erosion by fire-induced water repellency. Water Resour Res 42:1–9. https://doi.org/10.1029/2006WR004895
Riddell ES, Khan A, Mauck B et al (2012) Preliminary assessment of the impact of long-term fire treatments on in situ soil hydrology in the Kruger National Park. Koedoe 54(1):1–7. https://doi.org/10.4102/koedoe.v54i1.1070
Robichaud PR (2000) Fire effects on infiltration rates after prescribed fire in Northern Rocky Mountain forests, USA. J Hydrol 231:220–229. https://doi.org/10.1016/S0022-1694(00)00196-7
Sidle RC, Hirano T, Gomi T, Terajima T (2007) Hortonian overland flow from Japanese forest plantations—an aberration, the real thing, or something in between? Hydrol Process 21:3237–3247
Smit IPJ, Asner GP, Govender N et al (2010) Effects of fire on woody vegetation structure in African savanna. Ecol Appl 20(7):1865–1875
Stoof CR, Vervoort RW, Iwema J et al (2011) Hydrological response of a small catchment burned by experimental fire. Hydrol Earth Syst Sci Dis 8:4053–4098.https://doi.org/10.5194/hessd-8-4053-2011
Strydom T, Riddell ES, Rowe T et al (2019) The effect of experimental fires on soil hydrology and nutrients in an African savanna. Geoderma 345:114–122. https://doi.org/10.1016/j.geoderma.2019.03.027
Strydom S, Savage MJ (2016) A spatio-temporal analysis of fires in South Africa. S Afr J Sci 112(11–12):1–8
Strydom T, Smit IPJ, van Tol JJ (2024) Short and long-term fire effects on soil C and N in an African savanna. Geoderma Regional e00802
Thomaz EL (2018) Interaction between ash and soil microaggregates reduces runoff and soil loss. Sci Total Environ 625:1257–1263. https://doi.org/10.1016/j.scitotenv.2018.01.046
Trollope WSW, Trollope LA, Biggs HC et al (1998) Long-term changes in the woody vegetation of the Kruger National Park, with special reference to the effects of elephants and fire. Koedoe 41:103–112
USDA (1951) Soil survey manual. Soil Survey division staff: soil conservation service volume handbook 18. U.S. Department of Agriculture
Vadigi S, Ward D (2013) Shade, nutrients, and grass competition are important for tree sapling establishment in a humid savanna. Ecosphere 4(11):1–27. https://doi.org/10.1890/ES13-00239.1
van Huyssteen C (2020) Relating the South African soil taxonomy to the World reference base for soil resources. UJ, South Africa
Van Tol JJ, Le Roux PAL, Hensley M (2012) Pedotransfer functions to determine water conducting macroporosity in South African soils. Water Sci Technol 65:550–557. https://doi.org/10.2166/wst.2012.885
Van Wilgen BW, Biggs H, O’regan SP, Mare N (2000) Fire history of the savanna ecosystems in the Kruger National Park, South Africa, between 1941 and 1996. S Afr J Sci 96:167–178
Venter FJ, Govender N (2012) A geomorphic and soil description of the long-term fire experiment in the Kruger National Park, South Africa. Koedoe 54(1):1–10. https://doi.org/10.4102/koedoe.v54i1.1037
Warren SD (2001) Synopsis: influence of biological soil crusts on arid land hydrology and soil stability. In: Belnap J, Lange OL (eds) Biological soil crusts: structure, function, and management. Springer Berlin Heidelberg, Berlin, Germany, pp 349–360
Zhang Y, Niu J, Zhang M et al (2017) Interaction between Plant Roots and Soil Water Flow in Response to Preferential Flow paths in Northern China. Land Degrad Dev 28:648–663. https://doi.org/10.1002/ldr.2592
Acknowledgements
The authors would like to acknowledge South African National Parks and Scientific Services for maintaining these long-term experimental burn plots. We also extend our gratitude to Khensani Mkhonto, Duduzile Mzimba, Shedron Mukhumo and Samukelisiwe Linda for their assistance in the field during data collection.
Funding
Open access funding provided by University of the Free State. Funding was provided by South African National Parks for Tercia Strydom to cover logistical costs relating to fieldwork. Funding was also provided by the Department of Forestry, Fisheries and the Environmental (DFFE) – Natural Resource Management and National Research Foundation (NRF) Incentive Funding allocated to Johan J. van Tol.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by Tercia Strydom. The first draft of the manuscript was written by Tercia Strydom and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing Interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Giacomo Certini.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Strydom, T., Smit, I.P. & van Tol, J.J. The effect of time since last fire occurrence on selected soil hydrological properties in a South African savanna. Plant Soil (2024). https://doi.org/10.1007/s11104-024-06763-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11104-024-06763-y