
Abstract
Drought stress can negatively impact crop yield and quality. Improving wheat yields under drought stress is a major objective of agronomic research. Glutamine synthetase (GS) is a key enzyme of nitrogen metabolism that is critical to plant growth and development in abiotic stress response. However, to date, no systemic characterization of the GS genes has yet been conducted in wheat and its close relatives. We identified a total of 15 GS genes in Triticum aestivum (2n = 6x = 42; AABBDD), as well as 9 GS genes in Triticum dicoccoides (2n = 4x = 28; AABB), 6 in Aegilops tauschii (2n = 2x = 14; DD), and 5 in Triticum urartu (2n = 2x = 14; AA). The 35 GSs were further clustered into five lineages according to the phylogenetic tree. Synteny analysis revealed that the three subgenomes in bread wheat retained extensive synteny between bread wheat and its three relative species. We identified three up-regulated TaGSs (Ta4A.GSe, Ta4B.GSe, and Ta4D.GSe) from transcriptome data after drought and salt stress. Ta4D.GSe was subsequently used for further functional studies, and its subcellular localization were determined in Arabidopsis protoplasts. Its overexpression in Arabidopsis enhanced drought tolerance by increasing the ability of scavenging of reactive oxygen species (ROS) and osmotic adjustment. We identified GS gene family in four wheat species and performed comparative analyses of their relationships, chromosome locations, conserved motif, gene structure, and synteny. The subcellular localization of Ta4D.GSe was detected and its drought tolerance function was demonstrated. Taken together, these findings provide insight into the potential functional roles of the GS genes in abiotic stress tolerance.
Key message
This report clearly shows detailed characterization of GS gene family in four wheat species and demonstrates that Ta4D.GSe plays an important role in enhancing drought tolerance by improving the scavenging of ROS and osmotic adjustment ability in Arabidopsis.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Abiotic and biotic stresses are major environmental threats that result in considerable losses in crop productivity worldwide. In response to various stress factors, plants commonly exhibit osmotic and oxidative stresses (Baillo et al. 2019). One of the common responses shown by plants to mitigate stresses is the synthesis and accumulation of organic solutes known as osmoprotectants, such as proline (Pro), glycine betaine, O-sulphate, choline, sugars and polyols (Iqbal et al. 2014). Pro is a highly soluble neutral compound and mainly studied in response to osmotic stress (Ma et al. 2019; Verbruggen et al. 2008). It can stabilize antioxidant system through osmotic adjustments and protecting the integrity of cell membranes, thereby diminishing the impacts of reactive oxygen species (ROS; Reddy et al. 2015). It also can directly neutralize ROS, and may scavenge ·OH through a reaction that converts this amino acid to γ-aminobutyric acid (Sharma and Dieta 2006; Hayat et al. 2012; Signorelli et al. 2014). Multiple studies have proven that the importance of elevated Pro level in several plants exposed to varied stresses (e.g., drought, salt, chilling, heat, metal/metalloid and UV-B radiations) (Iqbal et al. 2014; Szabados and Savoure 2010).
Two pathways for Pro biosynthesis have been proposed in plants: glutamate (Glu) and ornithine (Orn) pathways. In Glu pathway, the biosynthesis of Pro begins with the phosphorylation of Glu to form γ-glutamyl phosphate, which is reduced by the action of bifunctional enzyme Δ1-pyrroline-5-carboxylate synthetase (P5CS; EC2.7.2.11/1.2.1.41) to glutamic-5-semialdehyde (GSA), which is spontaneously cyclized into pyrroline-5-carboxylate (P5C). Finally, P5C is reduced to Pro by the enzymatic catalysis of Δ1-pyrroline-5-carboxylate reductase (P5CR; EC 1.5.1.2). In this pathway, the biosynthesis of Pro takes place in the cytosol and chloroplasts, and glutamate is mainly derived from the glutamine synthetase-glutamine oxoglutarate aminotransferase (GS-GOGAT) cycle. In Orn pathway, Orn is trans-aminated to GSA through the activity of ornithine δ-aminotransferase (δ-OAT; EC 2.6.1.13), and subsequently gets converted to Pro via P5C (Szabados and Savoure 2010). It has been proved that biosynthesis of Pro in Arabidopsis occurs exclusively via the Glu pathway (Funck et al. 2008). Therefore, the Orn pathway remains controversial. Under osmotic stress, the biosynthesis of Pro by Glu pathway through enhancing GS-GOGAT cycle is the dominant pathway (Rejeb et al. 2014).
In most plant species, GS exists in multiple enzyme forms with a single isoform in the chloroplast (GS2) and up to five isoforms in the cytosol (GS1) (Swarbreck et al. 2011). GS2 is mainly involved in assimilation of NH4+, which is originated from nitrate reduction and photorespiration (Pérez-Delgado et al. 2015). The function of GS1 is mainly involved in the transport of storage nitrogen during seed germination and the reuse of nitrogen during leaf senescence (Harrison et al. 2003). In addition, GS is involved in grain protein synthesis. Nigro et al. (2017) isolated and confirmed that GS2 and Fd-GOGAT were related to grain protein accumulation. Further detailed analysis of the GS2 promoter showed that NAC transcription factor was involved in regulating its expression. Habash et al. (2010) also showed that overexpression of GS could increase the biomass and yield of transgenic plants.
GS is important for osmotic stress tolerance in plants. Szabados and Savoure (2010) showed that Pro accumulated in plants mainly via the GS-GOGAT pathway under drought stress. The GS2 mutant of Lotus japonicus showed lower Pro accumulation and rehydration ability than did the wild-type under drought stress (Díaz et al. 2010). In addition, overexpression of GS1 and GS2 in tobacco resulted in a higher accumulation of sucrose, Pro, and chlorophyll, and an enhanced ability to scavenge ROS, thus improving tolerance to drought induced stress (Yu et al. 2020).
Previous studies suggested that GS is one of the important physiological indicators for plants to adapt to drought stress. Results from experiments with drought-sensitive and drought-tolerant wheat genotypes showed that GS and RuBisCO (Ribulose-1,5-bisphosphate carboxylase/oxygenase, EC 4.1.1.39) could be used as physiological indicators to detect drought adaptation in wheat (Nagy et al. 2013). Singh and Ghosh (2013) showed that the expression of OsGS2 and OsGS1;1 may be related to the drought tolerance of Khitish (a drought-tolerant rice variety) under drought stress.
In addition, GS is involved in maintaining carbon and nitrogen balance in plants. Drought stress could limit the absorption of inorganic nitrogen in plants, inhibit the synthesis of carbohydrate and protein in leaves, and promote the degradation of protein and carbohydrate, thus break the balance of carbon and nitrogen metabolism in plants (Xu and Zhou 2006). Notably, GS can effectively utilize organic nitrogen and participate in metabolic processes such as photorespiration, reduction of ammonia, and reassimilation of circulating ammonia, thus improve drought tolerance of plants (Kusano et al. 2011).
In wheat, GS isoenzymes can be divided into four subfamilies on the basis of their sequence homology and cellular location. The first subfamily consists of GS2a, GS2b, and GS2c, which are nuclear-encoded (on chromosome 2) and chloroplast-localized proteins. The second subfamily consists of GS1a, GS1b, and GS1c, the third consists of GSr1 and GSr2, and the fourth GSe1 and GSe2. GS1, GSr and GSe are also nuclear-encoded (on chromosome 6, 4, 4, respectively) and cytoplasm-localized proteins (Nigro et al. 2017; Bernard et al. 2008; Habash et al. 2007). Bernard and Habash (2009) showed that GS isozymes have different functions in nitrogen metabolism of wheat. TaGS1 (GS1.1) and TaGSr (GS1.2) were mainly involved in the reuse of nitrogen in senescent leaves. In summary, there are many studies on the relationship between GS activity and nitrogen use efficiency and yield, and studies on the response of GS to abiotic stress such as drought, salt and extreme temperature are increasing gradually. The molecular mechanism of GS resistance to abiotic stress has attracted more and more attention in recent years.
Here, we aim to carry out a comprehensive study on the molecular characterization, phylogenetic relationship, and expression profiling of wheat GS gene family from the four wheat species, Triticum aestivum (Ta), Triticum dicoccoides (Td), Aegilops tauschii (Aet), and Triticum urartu (Tu). In addition, we infer that TaGSs respond to drought and salt stress through transcriptome data. The overexpression of Ta4D.GSe in Arabidopsis was used to confirm the effectiveness on drought tolerance of Ta4D.GSe.
Materials and methods
Data search and sequence retrieval
The genome files and annotation gff3 files of T. aestivum L. (Chinese spring), Triticum turgidum ssp. dicoccoides (Zavitan), A. tauschii Coss. (AL8/78) and T. urartu. (G1812) were downloaded from the EnsemblPlants (http://plants.ensembl.org) and MBKBASE (http://www.mbkbase.org). The published GS protein sequences were obtained from NCBI database (https://www.ncbi.nlm.nih.gov/protein), including Oryza sativa (Os), Zea mays (Zm), Hordeum vulgare (Hv), Arabidopsis thaliana (At), T), and six microbial species or genera Mucor ambiguus (Ma), Isosphaera pallida (Ip), Leptolyngbya (Le), Phaeodactylibacter (Ph), Caldithrix abyssi (Ca) and Phaeodactylum tricornutum (Pt). All of the 45 published GS proteins were used as query sequence to scan the whole genome protein sequences of bread wheat and its relative species with BLAST algorithm for Proteins (BLASP) search (e-value < 1e−5).
Genome-wide identification and characterization of GS genes
All candidate GS protein sequences were identified using the NCBI Conserved Domain Database (CDD, https://www.ncbi.nlm.nih.gov/cdd) with the automatic model and default parameters (threshold = 0.01, maximum hits = 500) and confirmed in InterPro (http://www.ebi.ac.uk/interpro/). The conserved protein domains in confirmed GS proteins were filtered from the CDD results.
The localization on chromosomes of all GS genes were analyzed by TBtools (Chen et al. 2020) using the annotation gff3 files. The number of amino acids, molecular weight, isoelectric point, and grand average of hydropathicity of GS proteins were analyzed by ProtParam (Gasteiger et al. 2005) (https://web.expasy.org/protparam/). The subcellular localizations were obtained from the web-server CELLO v2.5 (Yu et al. 2006) (http://cello.life.nctu.edu.tw/).
Synteny analysis and phylogenetic construction of GS genes
For the synteny analysis, the whole genome protein sequences of four wheat species were first pairwise compared by BLAST, then the calculation of the collinearity examination of paralogous genes were performed with MCScanX (http://chibba.pgml.uga.edu/duplication/) in TBtools. Finally, synteny visualization was conducted by TBtools.
The phylogenetic tree was constructed using the maximum likelihood method with MEGA X software (Kumar et al. 2018). At first, the GS proteins were aligned by Clustal W with default parameters. Then, a maximum likelihood phylogenetic tree was constructed, using the Poisson model, with 1000 bootstrap replicates. We colored the tree by web-server ITOL (https://itol.embl.de/) afterwards.
Gene structure and conserved motif analysis
The exon–intron structures of GS genes were constructed by Gene Structure Display Server 2.0 (GSDS, http://gsds.cbi.pku.edu.cn/). MEME v5.1.0 (http://meme-suite.org/tools/meme) was employed to analyze the conserved motifs of GS genes. We used the following parameters: distribution of motif occurrences, 0 or 1 occurrence per sequence; motif width 6 to 50 bp; and maximum number of motifs, 24. Finally, the analyzed results were visualized by TBtools.
Expression analysis of TaGS genes
According to gene expression profiles of Qingmai6 under 15% (w/v) polyethylene glycol (PEG) 6000 and 200 mmol·L−1 NaCl treatment (unpublished data), the differential expression of TaGS genes before and after treatment were obtained and analyzed. Then the expression characteristics were normalized and displayed as a heatmap. To detect the candidates that show the highest expression yields, we further checked the expressions of Ta4A.GSe, Ta4B.GSe, and Ta4D.GSe in Qingmai6 leaves with RT-qPCR after PEG-6000 (20% w/v, 2 h) treatment. The tissue specific expression of Ta4D.GSe was also detected by quantitative reverse-transcription PCR (RT-qPCR) at Qingmai6 seedling stage and maturation stage. According to the expression characteristics of Ta4D.GSe, it was induced by abiotic stress (PEG and salt) in Qingmai6. The function of Ta4D.GSe in drought tolerance was verified by comparing its levels of expression in three drought-tolerant wheat varieties (Qingmai6, Lumai21, and Shanrong3) (Zhang et al. 2011; Peng et al. 2009) and one drought-sensitive (Chinese Spring) (Hao et al. 2015) when exposed to drought stress conditions. The wheat seedlings were grown in nutrient solution, a continuous 25℃ temperature, a photoperiod of 12/12 h, and 50% relative humidity were used in a growth chamber. Drought stress treatment was carried out by submerging wheat seedling roots in nutrient solution of 20% (w/v) PEG-6000 at three-leaf stage. Leaves of the seedlings were sampled at different time points (0, 12, 24, 48, and 72 h) after treatment and RT-qPCR was used to detect Ta4D.GSe expression in the different wheat varieties.
Total RNA was extracted with the Total RNA Extraction Kit (Solarbio), and then one microgram of total RNA was reverse transcribed using PrimeScript™ RT reagent Kit (Perfect Real Time, Takara). In the RT-qPCR, β-actin gene was used as internal reference gene. For RT-qPCR, SYBR Green system (Roche) was used. Each experiment was repeated at least three times, with each reaction performed in triplicates, and the relative expression values were analyzed with the 2−ΔΔCq method. All tests were two-sided and p < 0.05 was considered statistically significant. All the gene-specific primers used for amplifications or vector constructions were listed in Table S1.
Subcellular localization and overexpression of Ta4D.GSe in Arabidopsis thaliana
Following the procedures of Wu et al. (2009) and Chen et al. (2006), we cloned and fused the non-terminator coding sequences of Ta4D.GSe to a green fluorescent protein (GFP) into vector Super1300 and transfected 10 μg plasmid DNA to 2 × 104 protoplasts by transient expression in 3-week-old Arabidopsis mesophyll protoplast method. GFP fluorescence was observed with a confocal laser scanning confocal microscope TCSsp5II (Agilent). In addition, the full coding sequence of the Ta4D.GSe was cloned into vector pBI121 with CaMV 35S promoter for gene overexpression. The recombinant plasmid and empty vector were introduced into the Agrobacterium tumefaciens strain EHA105, which were then infiltrated into At wild-type (Col-0) plants for transformation (Clough and Bent 1998). The T3 generation of transgenic Ta4D.GSe overexpression lines (Ta4D.GSe-OE) were used for further phenotypic analysis. The empty vector transgenic lines were used as mock, to serve as control, and will be referred to as WT.
Drought tolerance assessment
Surface sterilized seeds of At and transgenic lines were planted on Murashige and Skoog medium (MS). All of those were vernalized at 4℃ in dark for 3 days, and then grown under sterile conditions with 22℃, at a 16 h light/8 h dark cycle. For relative germination rate detection, the MS medium was added in different concentrations (50 mM, 100 mM, 150 mM) of mannitol. After 5 days, the relative germination rates were counted. For the detection of survival rate, 10 days after germination, seedling were grown in chamber with an equal weight of dry commercial soil (PINDSTRUP): vermiculite (1:3, v:v) at 22℃ under 16 h light/8 h dark cycle and 70% relative air humidity. Two-week-old seedlings were exposed to drought by discontinuing irrigation for 7 days and then re-watered with 100 mL water, which was the drought–rewatering treatment cycle. After 3 drought treatment cycles, the survival rate was determined based on the method of Li et al. (2020). After ten days of drought treatment, the enzyme reagent boxes (Solarbio) were used to detect the activities of GS, superoxide dismutase (SOD, EC 1.15.1.1), peroxidase (POD, EC 1.11.1.7) and catalase (CAT, EC 1.11.1.6). In addition, soluble sugars content was determined by the anthrone assay (Wang et al. 2013), and ninhydrin spectrophotometric assay (Bates et al. 1973) was used to detect the Pro level. Each experiment described above was repeated at least three independent times. All tests were two-sided and p < 0.05 was considered statistically significant.
Results
Genome-wide identification and characterization of GS genes in bread wheat and its relatives
Blast searches were performed by querying Os, Zm, Hv and At GS protein sequences from different wheat genomes, and 6, 15, 9 and 5 candidate GS genes were found in Aet, Ta, Td and Tu, respectively. The characteristics of 35 GS proteins in four wheat species are shown in Table 1. The coding amino acid length of four wheat species was between 60 and 884, and the molecular weights ranged from 6.15 to 98.33 kDa. Two proteins TRIDC4AG008800.5 (7.05) and TRIDC6BG052800.5 (7.67) showed isoelectric point above 7, indicating that these proteins were alkalescent, while all others showed isoelectric point below 7 indicating that they all were acidulous. In addition, all GS proteins in four wheat species had a negative grand average of hydropathicity (GRAVY) score, indicating that they were hydrophilic in nature.
According to the genomic location of each member of the GS gene family, a chromosome location map was constructed to illustrate the distribution of the GS genes by TBtools (Fig. 1; Table S1). The GS genes existed on the chromosomes from the groups 1, 2, 4, and 6 in Ta, Td and Tu, with number of GS genes in each chromosome ranging from one to two, and majority were detected at the distal end of the chromosomes. However, GS genes were located on 1D, 4D, and 6D in Aet. No GS gene was located on the chromosomes from the groups 3, 5, and 7 of all four wheat species. The exception here is TuUnGSe was located on chromosome TuUn in Tu because of the incomplete genome sequence. The chromosome 6D of Aet had 3 genes, the largest number of GS genes found in a single chromosome. The numbers of GS genes distributed in the subgenome showed little difference. The GS numbers in A, B and D subgenome of Ta were all 5, while a total of 4 and 5 GS genes were located on subgenomes A and B of Td, respectively. What is more, the GS numbers in A subgenome of Tu and D subgenome of Aet were 5 and 6.

Chromosome distribution of GS genes of four wheat species. The vertical scale on the left showed the physical size of chromosomes and black lines indicated the position of genes. The gene names correspond to those in Table S1
Synteny analysis of GS genes among four wheat species
Among all the 15 GS genes of Ta, 4 TaGSs had intergenomic homologous genes in Aet, 9 homologous genes in Td and 4 homologous genes in Tu, respectively (Fig. 2). The synteny analysis illustrated that four Aet-GSs could be mapped to bread wheat D subgenomes on the same chromosomes with one on 1D, two on 4D, except for Aet6D.GS1/Ta6B.GS1 homologous gene pairs (Aet6D.GS1 on 6D, Ta6B.GS1 on 6B). Moreover, nine Ta/Td.GSs homologous gene pairs were located on the same chromosomes, with one on 1A, two on 4A, one on 6A, one on 1B, one on 2B, two on 4B and one on 6B. Furthermore, only two homologous gene pairs were found between Tu and Td, with one on 4A and one on 6A.

Synteny analyses of GS genes between Triticum aestivum, Triticum dicoccoides, Aegilops tauschii and Triticum urartu
Phylogenetic analysis of GS genes
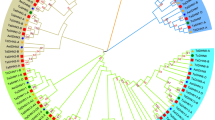
To study the phylogeny and subgroups of the GS family, an unrooted phylogenetic tree was constructed by using the 35 putative GS protein sequences from bread wheat and its relative species, and 4 OsGSs, 6 ZmGSs, 2 HvGSs, 5 AtGSs, 10 TaGSs and 17 GSs obtained from 6 micro-organisms in the literature (Table S2). Based on sequence analysis, phylogenetic studies and mapping data, Bernard et al. (2008) classified 10 TaGSs into four subfamilies: GS2, GS1, GSr, and GSe. However, in the phylogenetic tree, the 35 GSs were divided into five major clades, including 9 novel genotypes belonging to clade I, which show a high identity level to the GS genes in micro-organisms (Fig. 3). Based on the sequence of micro-organisms, clade I was separated from the GS family for the first time as a separate subfamily, including 4 TaGSs (TraesCS1A02G143000.1, TraesCS1B02G158600.1, TraesCS1D02G141800.1 and TraesCS6D02G065600.1), 2 TdGSs (TRIDC1AG021640.1 and TRIDC1BG025770.5), 2 AetGSs (AET1Gv20368100.2 and AET6Gv20169200.31), and 1 TuGSs (TuG1812G0195861400.01.T02). Therefore, the GS genes in clade I were expressed as GSm1 (Table S1). According to the published sequences of TaGSs, OsGSs, ZmGSs, HvGSs, and AtGSs, clade II belongs to the GS2 subfamily encoding nuclear gene for chloroplastic GS2 isoenzyme. Moreover, the other three clades (III, IV, and V) encode cytosolic GS1 isoenzymes (GSr, GSe, and GS1).

Phylogenetic tree of 80 GSs homolog proteins from different species. The GS proteins from 6 microbial species Mucor ambiguus (Ma), Isosphaera pallida (Ip), Leptolyngbya (Le), Phaeodactylibacter (Ph), Caldithrix abyssi (Ca) and Phaeodactylum tricornutum (Pt), and 4 model species Oryza sativa (Os), Zea mays (Zm), Hordeum vulgare (Hv), and Arabidopsis thaliana (At), and 10 Triticum aestivum (Ta). Five major clades were distinguished with colors
Sequence and structural analyses of GS genes and proteins
The gene structure of GSs genes was analyzed according to the gene annotation gff3 files. As illustrated in Fig. 4 and Table S3, 22 conserved motifs, with 11 to 50 amino acids, of GS genes were identified through the MEME v5.1.0, and most of motifs displayed similar patterns within the GS genes in four wheat species. For example, motif 8, 3, 21, 2, 7, 4, 1, 6, 5 were conserved in 19 GSs, and motif 21, 7, 4, 1, 6, 5 were retained as part of the combination in two GSs, Td4A.GSr and Td6A.GS1. Compared with GS1, GSr and GSe subfamily, motif 18 was specific to the GS2 branch. The number and type of conserved motifs were consistent in clade I, however, they were different from those in clade II to clade V. These results indicated that the GSs in different subfamilies had different conserved motif distributions, which might suggest a conserved function of different subfamilies. Gene structure analysis showed that a majority of the GS genes had more than one exon. In the same subfamily, the gene structure and conserved motif distribution were similar, indicating that the phylogenetic tree constructed in this study is accurate.

Phylogenetic, conserved motif and gene structure analyses of GS genes. A Phylogenetic analysis, B conserved motif, and C gene structure. UTR untranslated region, CDS coding sequence
RNA-seq expression profile of TaGS genes in abiotic stress
To identify the potential functions of TaGS genes in response to abiotic stress, the expression data under PEG and NaCl treatment were obtained from Qingmai6 RNA-Seq data, respectively. The expression profiles of TaGSs in leaf (0 h, 24 h, 48 h, 72 h) under abiotic stress were normalized to log2FPKM and performed with heatmap (Fig. 5). Under both PEG and NaCl treatment, the expression of Ta4A.GSe, Ta4B.GSe, and Ta4D.GSe from subfamily GSe were significantly up-regulated, and Ta4D.GSe was the most strongly up-regulated gene. The results of RT-qPCR in Qingmai6 under PEG treatment were consistent with the RNA-seq data shown in the heatmap (Fig. 6). The expression levels of Ta4D.GSe in Qingmai6 different tissues at seedling stage and maturation stage were compared. The highest expression levels were found in root and spikelet, respectively (Fig. S1), indicating that Ta4D.GSe function mainly in those tissues.

Heat map of the expression profiling of TaGS genes at different time under abiotic stress Green and red denote lower and higher expression levels, respectively. The lables 0 h, 24 h, 48 h, and 72 h indicate the time that passed after the PEG (A) and NaCl (B) treatment. Transcriptome expression of reads per kilobase per million mapped reads (rpkm) is the RNA-Seq expression unit

Expression levels of Ta4A.GSe, Ta4B.GSe, and Ta4D.GSe in leaves under drought stress. The relative expression levels were calculated by setting the expression value of Ta4A.GSe as 1. The relative expression values were calculated through the 2−ΔΔCq approach
To confirm that the expression pattern of Ta4D.GSe was associated with drought tolerance, RT-qPCR were used to detect it in drought-tolerant wheat varieties (Qingmai6, Lumai21, Shanrong3) and drought-sensitive variety (Chinese Spring) under PEG treatment. As shown in Fig. 7, Ta4D.GSe from drought-tolerant wheat varieties were more sensitive, being significantly up-regulated under PEG treatment. Thus, based on the analysis above, Ta4D.GSe was selected as representative gene for further functional investigation.

The expression pattern of Ta4D.GSe in different wheat varieties under PEG treatment. Drought-tolerant wheat varieties: Qingmai6, Lumai21, and Shanrong3. Drought-sensitive variety: Chinese Spring. The relative expression levels in each variety were calculated by setting the expression value at 0 h as 1. The relative expression values were calculated through the 2−ΔΔCq approach
Subcellular localization and ectopic overexpression analysis of Ta4D.GSe in Arabidopsis
The subcellular localization analysis showed that Ta4D.GSe was localized at the cytoplasm (Fig. 8), and the result was consistent with the subcellular localization prediction by CELLO v2.5 (Table 1). Compared to those of the WT, no significant difference were observed in biomass production under normal conditions, however, the germination rate and tolerance of seedlings under mannitol treatment and in response to repeated drought treatments were significantly improved (Fig. S2; Fig. 9), suggesting that the function of Ta4D.GSe is more evident following drought stress.

The subcellular localization of Ta4D.GSe. Scale bar 10 μm

Overexpression of Ta4D.GSe enhances drought resistance in Arabidopsis. WT, empty vector lines. A Phenotype of wild-type and homozygous transgenic lines grown under normal and drought stress conditions. B Phenotype of wild-type and homozygous transgenic lines under normal and 5 days after rewatering conditions. C The survival rate of wild-type and homozygous transgenic lines (Ta4D.GSe-OE) at 5 days after rewatering conditions
The activities of SOD, POD, and CAT of Ta4D.GSe-OE were significantly higher than that of WT before drought stress (Fig. S3). After 10 days of drought treatment, the enzymatic activities were significantly increased in both the Ta4D.Gse-OE and WT lines. The activities in SOD, POD, and CAT of Ta4D.GSe-OE were increased by 2.3, 1.2, and 1.8-fold, respectively, and in the WT, these enzyme’s activities were increased by 1.6, 1.1, and 1.6-fold, respectively, when compared with those of the untreated plants (Fig. S3b–d). The result indicated that Ta4D.GSe plays a significant role in conferring drought tolerance by improving the scavenging of ROS in Arabidopsis. In addition to the results above, soluble sugars and free-Pro accumulated in WT and Ta4D.GSe-OE and the accumulation in Ta4D.GSe-OE were much more significant than that in WT, after 10 days of drought treatment. The content of soluble sugars and free-Pro in Ta4D.GSe-OE were increased by 4.1 and 24.8-fold, respectively, and in WT were increased by 2.8 and 21.4-fold, respectively (Fig. S3e, f). The result showed that Ta4D.GSe could improve the osmotic adjustment ability in Arabidopsis through accumulate osmoregulating substances.
Discussion
Whole genome-wide screening and characterization of the GS gene family has been performed in several plants following the release of high-quality reference genomes. It has been generally accepted that GSs play a large role in nitrogen metabolism. For example, with the help of whole genome sequencing data, Populus trichocarpa was the first plant species in which the complete GS family was observed to be duplicated (Castro-Rodríguez et al. 2011). Liu et al. (2018) identified six GS genes from Gracilariopsis lemaneiformis genome through transcriptome data and all of these genes were divided into three clusters, and found that GSII might have a key role in the process of nitrogen metabolism. Moreover, Czyż et al. (2020) studied the complex characterization of narrow-leafed lupin GS gene family with advanced genomic resources and pointed out that sub-functionalization and/or regulatory rewiring played an important role in shaping the primary metabolic pathways of the extant carbon and nitrogen in some lineages. However, very little information about GS from Ta and its relatives is available. Nowadays, the genomes of Ta, Td, Aet, and Tu have been better sequenced for further understanding of wheat genomics (IWGSC 2018; Avni et al. 2017; Luo et al. 2017; Ling et al. 2018).
A previous study identified 10 GS genes in wheat using heterologous complementation and cloning, which is the first cloning and study of GS genes in wheat (Bernard et al. 2008). In this study, total of 15, 9, 6, and 5 GSs were identified in Ta, Td, Aet, and Tu, respectively (Table 1). In previous studies, plant GS genes were organized in 4 groups, 3 of which code for cytosolic isoforms (GS1) and 1 codes for the chloroplastic isoform (GS2). Our results indicate that the family members are organized in 5 groups, with clade III, IV, and V belonging to GS1 and clade II belonging to GS2, in addition, 9 genes belong to clade I showing a high identity level to the GS genes in micro-organisms (Fig. 3). Rodríguez et al. (2011) also found that some GS genes clustered with archaebacteria. The hexaploid wheat (Ta, BBAADD) established from Td (BBAA) and Aet (DD) less than 10,000 years ago (Marcussen et al. 2014; Feldman et al. 1995). The tetraploid wheat (Td) was produced through domestication by the wild tetraploid wheat T. turgidum ssp. dicoccoides (BBAA), which was formed via allotetraploidization from Tu (AA) and Aegilops speltoides (BB) about 0.5 million years ago (Marcussen et al. 2014; Dvorak et al. 2005). For the number of GSs in each isoform, Ta wheat had nearly 1.5 times as many as Td wheat and 3 times as many as Tu wheat (Table 1) and the GSs located in each genome corresponds one by one (Fig. 2). Furthermore, the synteny analysis illustrated that most TdGSs and AetGSs had intergenomic homologous genes in Ta, while two of five TuGSs had homologous genes in Td and Ta. These results also supported the evolutionary relationship between diploid, tetraploid and hexaploid wheat.
Numerous studies have shown that GS can regulate nitrogen metabolism in plants and affect development and growth (Migge et al. 2000; Oliveira et al. 2002; Thomsen et al. 2014). Bernard et al. (2008) also suggested that wheat cytosolic isozymes (GS1) played a major role in assimilating ammonia during the critical phases of remobilisation of nitrogen to the grain. However, the involvement of GS in tolerance to abiotic stress has rarely been investigated. The present study focused on TaGS genes that could play crucial roles in abiotic stress tolerance. Combining the transcriptome expression of TaGSs genes, Ta4D.GSe was selected to further understand its characteristics and functions. For the localization analysis, Ta4D.GSe was localized in cytoplasm (Fig. 8), such a result exactly meets the characteristic of cytosolic GS1 isoenzyme. Transcriptome data showed that Ta4D.GSe was expressed in almost all organs, particularly in spikelets (Fig. S1). In addition, Ta4D.Gse-OE showed a higher germination rate under mannitol stress and drought rehydration survival rate when exposed to repeated drought treatments (Fig. S2; Fig. 9), which were associated with the function of Ta4D.Gse for osmoregulation and ROS scavenging. Under drought treatment, Ta4D.GSe-OE lines accumulated much more soluble sugars and free Pro (Fig. S3), which could regulate osmotic pressure and protect the integrity of cell membranes as osmoprotectants. Futhermore, the free Pro could also directly neutralize ROS and might scavenge ·OH (Sharma and Dieta 2006; Hayat et al. 2012; Signorelli et al. 2014). In addition, GS could also resist stress by protecting the antioxidant system (Reddy et al. 2015). Before drought stress, there were small but significant differences in the activities of SOD, POD and CAT between Ta4D.GSe-OE and WT; however, the enzyme activities differed greatly between Ta4D.GSe-OE and WT, after drought stress (Fig. S3), indicating that Ta4D.Gse could stabilize antioxidant system, thereby diminishing the impacts of ROS. Taken together, these results suggested that Ta4D.GSe may be involved in drought tolerance. However, the detailed correlation between GSs and drought tolerance remains to be further verified.
Conclusions
In this study, 6 AetGSs, 15 TaGSs, 9 TdGSs and 5 TuGSs were identified and clustered into five lineages according to the phylogenetic tree. Particularly, according to the published sequence of micro-organisms nine novel GSs were found, and expressed them as GSm1. Then, their chromosome location, conserved motif, gene structure, and synteny were analyzed for understanding the gene family expansion and gene evolution. In addition, we used transcriptome data of Qingmai6 under abiotic stress conditions (drought and salinity) to identify the TaGSs expression profile, implying that Ta4A.GSe, Ta4B.GSe, and Ta4D.GSe might be involved in the response abiotic stress. Because of its high expression level, Ta4D.GSe was selected as representative gene for further functional investigation. The subcellular location of Ta4D.GSe to the cytoplasm was detected using confocal microscopy. Furthermore, its functions involved in abiotic stress were identified by inducing its overexpression in At. The results showed that, Ta4D.GSe plays an important role in conferring drought tolerance by improving the scavenging of ROS and the osmotic adjustment ability in Arabidopsis. Taken together, these findings provide insight into the potential functional roles of the TaGSs genes in abiotic stress tolerance.
Data availability
The datasets supporting the conclusions of the present study are included within this article (and its additional files). The authors are pleased to share any raw data upon request.
Consent for publication
Not applicable.
References
Avni R, Nave M, Barad O, Baruch K, Twardziok SO et al (2017) Wild emmer genome architecture and diversity elucidate wheat evolution and domestication. Science 357:93–97. https://doi.org/10.1126/science.aan0032
Baillo EH, Kimotho RN, Zhang ZB, Xu P (2019) Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 10:771. https://doi.org/10.3390/genes10100771
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207. https://doi.org/10.1007/bf00018060
Bernard SM, Habash DZ (2009) The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. N Phytol 182:608–620. https://doi.org/10.1111/j.1469-8137.2009.02823.x
Bernard SM, Moller AL, Dionisio G, Kichey T, Jahn TP, Dubois F, Baudo M, Lopes MS, Terce-Laforgue T, Foyer CH, Parry MA, Forde BG, Araus JL, Hirel B, Schjoerring JK, Habash DZ (2008) Gene expression, cellular localisation and function of glutamine synthetase isozymes in wheat (Triticum aestivum L.). Plant Mol Biol 67:89–105. https://doi.org/10.1007/s11103-008-9303-y
Castro-Rodríguez V, Garcia-Gutierrez A, Canales J, Avila C, Kirby EG, Canovas FM (2011) The glutamine synthetase gene family in Populus. BMC Plant Biol 11:119. https://doi.org/10.1186/1471-2229-11-119
Chen CJ, Chen H, Zhang Y, Thomas HR, Frank MH, He YH, Xia R (2020) TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant 13:1194–1202. https://doi.org/10.1016/j.molp.2020.06.009
Chen Y, Ji F, Xie H, Liang J (2006) Overexpression of the regulator of G-protein signalling protein enhances ABA-mediated inhibition of root elongation and drought tolerance in Arabidopsis. J Exp Bot 57:2101–2110. https://doi.org/10.1093/jxb/erj167
Clough SJ, Bent AF (1998) Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735–743. https://doi.org/10.1046/j.1365-313x.1998.00343.x
Czyż KB, Książkiewicz M, Koczyk G, Szczepaniak A, Podkowiński J, Naganowska B (2020) A tale of two families: whole genome and segmental duplications underlie glutamine synthetase and phosphoenolpyruvate carboxylase diversity in narrow-leafed lupin (Lupinus angustifolius L.). Int J Mol Sci 21:2580. https://doi.org/10.3390/ijms21072580
Díaz P, Betti M, Sanchez DH, Udvardi MK, Monza J, Marquez AJ (2010) Deficiency in plastidic glutamine synthetase alters proline metabolism and transcriptomic response in Lotus japonicus under drought stress. N Phytol 188:1001–1013. https://doi.org/10.1111/j.1469-8137.2010.03440.x
Dvorak J (2005) Tempos of gene locus deletions and duplications and their relationship to recombination rate during diploid and polyploid evolution in the Aegilops-Triticum alliance. Genetics 171:323–332. https://doi.org/10.1534/genetics.105.041632
Feldman M, Lupton FGH, Miller TE (1995) Wheats. Triticum spp. (Gramineae–Triticinae). In: Smartt J, Simmonds NW (eds) Evolution of crop plants. Longman Scientific and Technical Press, London, pp 184–192
Funck D, Stadelhofer B, Koch W (2008) Ornithine-δ-aminotransferase is essential for arginine catabolism but not for proline biosynthesis. BMC Plant Biol 8:40. https://doi.org/10.1186/1471-2229-8-40
Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins MR, Appel RD, Bairoch A (2005) Protein identification and analysis tools on the ExPASy server. In: Walker JM (ed) The proteomics protocols handbook. Springer protocols handbooks. Humana Press, Totowa, pp 571–607. https://doi.org/10.1385/1-59259-890-0:571
Habash DZ, Bernard S, Schondelmaier J, Weyen J, Quarrie SA (2007) The genetics of nitrogen use in hexaploid wheat: N utilisation, development and yield. Theor Appl Genet 114:403–419. https://doi.org/10.1007/s00122-006-0429-5
Habash DZ, Massiah AJ, Rong HL, Wallsgrove RM, Leigh RA (2010) The role of cytosolic glutamine synthetase in wheat. Ann Appl Biol 138:83–89. https://doi.org/10.1111/j.1744-7348.2001.tb00087.x
Hao P, Zhu J, Gu A, Lv D, Ge P, Chen G, Li X, Yan Y (2015) An integrative proteome analysis of different seedling organs in tolerant and sensitive wheat cultivars under drought stress and recovery. Proteomics 15:1544–1563. https://doi.org/10.1002/pmic.201400179
Harrison J, Crescenzo MAP, Sené O, Hirel B (2003) Does lowering glutamine synthetase activity in nodules modify nitrogen metabolism and growth of Lotus japonicus? Plant Physiol 133:253–262. https://doi.org/10.1104/pp.102.016766
Hayat S, Hayat Q, Alyemeni MN, Wani AS, Pichtel J, Ahmad A (2012) Role of proline under changing environments: a review. Plant Sign Behav 7:1456–1466. https://doi.org/10.4161/psb.21949
International Wheat Genome Sequencing Consortium (IWGSC), Appels R, Eversole K, Feuillet C, Keller B et al (2018) Shifting the limits in wheat research and breeding using a fully annotated reference genome. Science 361:eaar7191. https://doi.org/10.1126/science.aar7191
Iqbal N, Umar S, Khan NA, Khan MIR (2014) A new perspective of phytohormones in salinity tolerance: regulation of proline metabolism. Environ Exp Bot 100:34–42. https://doi.org/10.1016/j.envexpbot.2013.12.006
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35:1547–1549. https://doi.org/10.1093/molbev/msy096
Kusano M, Tabuchi M, Fukushima A, Funayama K, Diaz C, Kobayashi M, Hayashi N, Tsuchiya YN, Takahashi H, Kamata A, Yamaya T, Saito K (2011) Metabolomics data reveal a crucial role of cytosolic glutamine synthetase 1;1 in coordinating metabolic balance in rice. Plant J 66:456–466. https://doi.org/10.1111/j.1365-313X.2011.04506.x
Li X, Liu Q, Feng H, Deng J, Zhang R, Wen J, Dong J, Wang T (2020) Dehydrin MtCAS31 promotes autophagic degradation under drought stress. Autophagy 16:862–877. https://doi.org/10.1080/15548627.2019.1643656
Ling HQ, Ma B, Shi XL, Liu H, Dong LL, Sun H, Cao YH, Gao Q, Zheng SS, Li Y, Yu Y, Du HL, Qi M, Li Y, Lu HW, Yu H, Cui Y, Wang N, Chen CL, Wu HL, Zhao Y, Zhang JC, Li YW, Zhou WJ, Zhang BR, Hu WJ, van Eijk MJT, Tang JF, Witsenboer HMA, Zhao SC, Li ZS, Zhang AM, Wang DW, Liang CZ (2018) Genome sequence of the progenitor of wheat A subgenome Triticum urartu. Nature 557:424–428. https://doi.org/10.1038/s41586-018-0108-0
Liu XJ, Zhang QF, Huan ZY, Zhong MQ, Chen WZ, Du H (2018) Identification and characterization of glutamine synthetase isozymes in Gracilaria lemaneiformis. Aquat Bot 146:23–30. https://doi.org/10.1016/j.aquabot.2018.01.006
Luo MC, Gu YQ, Puiu D, Wang H, Twardziok SO, Deal KR, Huo NX, Zhu TT, Wang L, Wang Y, McGuire PE, Liu SY, Long H, Ramasamy RK, Rodriguez JC, Van SL, Yuan LX, Wang ZZ, Xia ZQ, Xiao LC, Anderson OD, Ouyang SH, Liang Y, Zimin AV, Pertea G, Qi P, Ennetzen JLB, Dai XT, Dawson MW, Muller HG, Kugler K, Rivarola-Duarte L, Spannagl M, Mayer KFX, Lu FH, Bevan MW, Leroy P, Li PC, You FM, Sun QX, Liu ZY, Lyons E, Wicker T, Salzberg SL, Devos KM, Dvorak J (2017) Genome sequence of the progenitor of the wheat D genome Aegilops tauschii. Nature 551:498–502. https://doi.org/10.1038/nature24486
Ma YL, Cao J, Chen QQ, He JH, Liu ZB, Wang JM, Li XF, Yang Y (2019) The kinase CIPK11 functions as a negative regulator in drought stress response in Arabidopsis. Int J Mol Sci 20:2422. https://doi.org/10.3390/ijms20102422
Marcussen T, Sandve SR, Heier L, Spannagl M, Pfeifer M et al (2014) Ancient hybridizations among the ancestral genomes of bread wheat. Science 345:1250092. https://doi.org/10.1126/science.1250092
Migge A, Carrayol E, Hirel B, Becker TW (2000) Leaf-specific overexpression of plastidic glutamine synthetase stimulates the growth of transgenic tobacco seedlings. Planta 210:252–260. https://doi.org/10.1007/PL00008132
Nagy Z, Nemeth E, Guoth A, Bona L, Wodala B, Pecsvaradi A (2013) Metabolic indicators of drought stress tolerance in wheat: glutamine synthetase isoenzymes and Rubisco. Plant Physiol Biochem 67:48–54. https://doi.org/10.1016/j.plaphy.2013.03.001
Nigro D, Fortunato S, Giove SL, Mangini G, Yacoubi I, Simeone R, Blanco A, Gadaleta A (2017) Allelic variants of glutamine synthetase and glutamate synthase genes in a collection of durum wheat and association with grain protein content. Diversity 9:52. https://doi.org/10.3390/d9040052
Oliveira IC, Brears T, Knight TJ, Clark A, Coruzzi GM (2002) Overexpression of cytosolic glutamine synthetase. Relation to nitrogen, light, and photorespiration. Plant Physiol 129:1170–1180. https://doi.org/10.1104/pp.020013
Peng ZY, Wang MC, Li F, Lv HJ, Li CL, Xia GM (2009) A proteomic study of the response to salinity and drought stress in an introgression strain of bread wheat. Mol Cell Proteomics 8:2676–2686. https://doi.org/10.1074/mcp.M900052-MCP200
Pérez-Delgado CM, Garcia-Calderon M, Marquez AJ, Betti M (2015) Reassimilation of photorespiratory ammonium in Lotus japonicus plants deficient in plastidic glutamine synthetase. PLoS ONE 10:e0130438. https://doi.org/10.1371/journal.pone.0156568
Reddy PS, Jogeswar G, Rasineni GK, Maheswari M, Reddy AR, Varshney RK, Kavi Kishor PB (2015) Proline over-accumulation alleviates salt stress and protects photosynthetic and antioxidant enzyme activities in transgenic sorghum [Sorghum bicolor (L.) Moench]. Plant Physiol Biochem 94:104–113. https://doi.org/10.1016/j.plaphy.2015.05.014
Rejeb KB, Abdelly C, Savoure A (2014) How reactive oxygen species and proline face stress together. Plant Physiol Biochem 80:278–284. https://doi.org/10.1016/j.plaphy.2014.04.007
Rodríguez VC, Garcia-Gutierrez A, Canales J, Avila C, Kirby EG, Canovas FM (2011) The glutamine synthetase gene family in Populus. BMC Plant Biol 11:119. https://doi.org/10.1186/1471-2229-11-119
Sharma SS, Dieta KL (2006) The significance of amino acids and amino acid-derived molecules in plant responses and adaptation to heavy metal stress. J Exp Bot 57:711–726. https://doi.org/10.1093/jxb/erj073
Signorelli S, Coitiño EL, Borani O, Monza J (2014) Molecular mechanisms for the reaction between ·OH radicals and proline: insights on the role as reactive oxygen species scavenger in plant stress. J Phys Chem B 118:37–47. https://doi.org/10.1021/jp407773u
Singh KK, Ghosh S (2013) Regulation of glutamine synthetase isoforms in two differentially drought-tolerant rice (Oryza sativa L.) cultivars under water deficit conditions. Plant Cell Rep 32:183–193. https://doi.org/10.1007/s00299-012-1353-6
Swarbreck SM, Defoin-Platel M, Hindle M, Saqi M, Habash DZ (2011) New perspectives on glutamine synthetase in grasses. J Exp Bot 62:1511–1522. https://doi.org/10.1093/jxb/erq356
Szabados L, Savoure A (2010) Proline: a multifunctional amino acid. Trends Plant Sci 15:89–97. https://doi.org/10.1016/j.tplants.2009.11.009
Thomsen HC, Eriksson D, Moller IS, Schjoerring JK (2014) Cytosolic glutamine synthetase: a target for improvement of crop nitrogen use efficiency? Trends Plant Sci 19:656–663. https://doi.org/10.1016/j.tplants.2014.06.002
Verbruggen N, Hermans C (2008) Proline accumulation in plants: a review. Amino Acids 35:753–759. https://doi.org/10.1007/s00726-008-0061-6
Wang JC, Xu H, Zhu Y, Liu QQ, Cai XL (2013) OsbZIP58, a basic leucine zipper transcription factor, regulates starch biosynthesis in rice endosperm. J Exp Bot 64:3453–3466. https://doi.org/10.1093/jxb/ert187
Wu FH, Shen SC, Lee LY, Lee SH, Chan MT, Lin CS (2009) Tape-Arabidopsis Sandwich—a simpler Arabidopsis protoplast isolation method. Plant Methods 5:16. https://doi.org/10.1186/1746-4811-5-16
Xu ZZ, Zhou GS (2006) Combined effects of water stress and high temperature on photosynthesis, nitrogen metabolism and lipid peroxidation of a perennial grass Leymus chinensis. Planta 224:1080–1090. https://doi.org/10.1007/s00425-006-0281-5
Yu CS, Chen YC, Lu CH, Hwang JK (2006) Prediction of protein subcellular localization. Proteins 64:643–651. https://doi.org/10.1002/prot.21018
Yu HD, Zhang YM, Zhang ZY, Zhang J, Wei YH, Jia XT, Wang XC, Ma XM (2020) Towards identification of molecular mechanism in which the overexpression of wheat cytosolic and plastid glutamine synthetases in tobacco enhanced drought tolerance. Plant Physiol Biochem 151:608–620. https://doi.org/10.1016/j.plaphy.2020.04.013
Zhang YQ, Zhang HS, Qi L, Liu YG, Li LY (2011) Effect of water stress on drought-resistance characteristics of wheat seedling with different fertilizer and water types. J Agric 1:1–7
Acknowledgements
We thank Steven Xu for critical proofreading and language editing.
Funding
This work was supported by the National Natural Science Foundation of China Grants (32101715); the Foundation of Research and Application of Whole Genome Selection in Wheat (2019LZGC016); the Wheat Innovation Team of Modern Agricultural Production Systems in Shandong Province; the High-Level Talents Project of Qingdao Agricultural University (663/1119057; 663/1120077); the State Key Laboratory of Crop Biology at Shandong Agricultural University (2020KF03).
Author information
Authors and Affiliations
Contributions
PM and HY designed the research; QS, XL, CG, and LZ conducted the research; QS and XL prepared the samples; HY and YY analyzed the data; HY wrote the draft; PM, WM, and XH made the revision of the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interest.
Ethical approval
Not applicable.
Informed consent
Not applicable.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
11103_2022_1287_MOESM2_ESM.tif
Supplementary file2 (TIF 1354 kb) Fig. S1 Expression pattern of Ta4D.GSe in different tissues. A Expression levels of Ta4D.GSe in seedling stage. B Expression levels of Ta4D.GSe in maturation stage. The relative expression levels in each tissue were calculated by setting the expression value of Ta4D.GSe in root as 1. The relative expression values were calculated through the 2−ΔΔCq approach
11103_2022_1287_MOESM3_ESM.tif
Supplementary file3 (TIF 12404 kb) Fig. S2 Overexpression of Ta4D.GSe improves the relative germination rate of Arabidopsis. A Germination of Arabidopsis treated with 150 mM mannitol. B–D The relative germination rate of Arabidopsis treated with mannitol at 50, 100, 150 mM
11103_2022_1287_MOESM4_ESM.tif
Supplementary file4 (TIF 5092 kb) Fig. S3 The differences of physiological indexes before and after treatment of drought in Arabidopsis. WT empty vector lines, OE Ta4D.GSe overexpression transgenic lines. A Identification of the glutamine synthetase (GS) activity in WT and OE before and after drought treatment. B–D Activity of SOD (B), POD (C), CAT (D) in Arabidopsis. E and F Changes of soluble sugars and proline content of Arabidopsis under drought treatment
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Yin, H., Sun, Q., Lu, X. et al. Identification of the glutamine synthetase (GS) gene family in four wheat species and functional analysis of Ta4D.GSe in Arabidopsis thaliana. Plant Mol Biol 110, 93–106 (2022). https://doi.org/10.1007/s11103-022-01287-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11103-022-01287-4