
Abstract
Cacao mucilage exudates (CMEs), byproducts of cacao beans fermentation, are effluents rich in sugars and low-molecular-weight organic acids. In this study, we utilized CMEs to formulate culture media for the biosynthesis of polyhydroxyalkanoates (PHAs) using a native Bacillus megaterium strain (B2). Our investigation followed the adaptation process of B2 to the CMEs-based media, and we closely monitored the development of the microorganism in terms of consumption of acids and sugars. Upon successful adaptation to the CMEs-based media, we transferred the strain to a 4-L batch bioreactor to evaluate the impact of operational variables on PHAs production. We examined the microorganisms’ yield, productivity, and growth kinetics using the CME-based media. Our results showed a product yield of 0.98 g PHA/g carbon source, with an accumulation percentage of 57% and a productivity increase of 33% compared to PHA produced from residual glycerol using the same microorganism. The biopolymer's structure was confirmed to be of a polyhydroxybutyrate homopolymer (PHB) type using various molecular characterization techniques, including Matrix-Assisted Laser Desorption/Ionization mass spectrometry (MALDI-TOF-MS), infrared spectroscopy (IR), elemental composition (EC), ultraviolet–visible spectroscopy (UV–VIS), and thermal analysis (TGA, DSC).
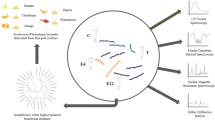
Graphical Abstract

Similar content being viewed by others


Explore related subjects
Find the latest articles, discoveries, and news in related topics.Avoid common mistakes on your manuscript.
Introduction
In 2022, worldwide production of synthetic polymers reached 390 million tons [1]. Unfortunately, most of these plastics do not degrade naturally, and their manufacture releases significant amounts of carbon dioxide into the atmosphere. One possible solution to this pollution problem is polyhydroxyalkanoates (PHAs), which are biodegradable polyesters derived from renewable sources. PHAs have thermoplastic and elastomeric properties like conventional synthetic polymers derived from petrochemical sources [2] These polymers are produced by various microorganisms as carbon and energy reserves [3]. Interestingly, the structure of PHAs can be influenced by the composition of the growth media [4].
PHAs have received significant attention for their applications in medicine [5] [6]agriculture [7], and food science [8]. However, the primary disadvantage of PHA production is its economic viability. PHA biosynthesis requires a carbon source, typically involving low molecular weight organic substrates such as monosaccharides and alcohols. Due to the high cost of these substrates, current research efforts are focused on using agro-industrial residues as carbon sources for large-scale PHA production [9]. Alternative carbon sources such as beet molasses [10], crude glycerol [11], and cheese whey [12] have been considered for PHAs production. Some of these residues allow PHAs production in high yields and productivities like those obtained with conventional carbon sources such as glucose, sucrose, and fructose[13].
Based on data from the International Cacao Organization, the global production of cacao beans in the 2020/2021 season reached 4.84 million tons [14]. In 2019, Colombia was ranked eleventh among cacao bean-producing countries, with a total yield of 69 thousand tons [15]. As is typical with other agro-industrial commodities, cacao bean production generates a substantial amount of residual biomass resulting from the harvesting and processing of cacao fruits. The discarded waste materials contain a high concentration of organic compounds, which, if left untreated, can have adverse effects on the environment and pose health hazards for both humans and crops. It is worth remarking that the circularity of cacao fruit to cacao beans is currently only 8 to 10% [16].
The presence of a white mucilaginous layer (or pulp) with high sugar content in fresh cacao beans is a crucial factor in promoting the growth of beneficial microorganisms during the fermentation process. This process is vital for deactivating the seed embryo and producing aroma precursors that determine the final product's flavor and quality [17, 18] Cacao mucilage exudates are liquid remnants that emerge from the cacao pulp during fermentation, comprising between 3 and 5% of the cacao fruit's weight. Fresh cacao mucilage exudate (CME) is extracted via cold pressing of fresh cacao beans prior to fermentation, whereas spent cacao mucilage exudate (SCME) is produced during fermentation. As a result, fresh CME has a high sugar (glucose, fructose, and sucrose) and mineral content, while SCME contains low-molecular-weight organic acids and residual sugars. Our research group recently reported the utilization of fresh CME as a culture medium to produce bacterial cellulose (BC) both in laboratory and pilot-plant scales [19]. The residual medium from BC production (RMBC) still contains substantial amounts of simple sugars (57.4 g L−1) rendering it a feasible substrate for producing biopolymers such as PHAs. Moreover, CMEs, particularly the spent effluent, are deemed waste products with adverse environmental effects owing to their high COD (chemical oxygen demand). The production of biopolymers using residual biomass from cacao bean processing can increase crop circularity while decreasing its environmental footprint.
The selection of microorganisms plays a crucial role in the production of PHAs and their potential applications. Gram-negative bacteria are particularly desirable due to their ability to accumulate higher amounts of PHAs compared to Gram-positive bacteria. However, it should be noted that PHAs extracted from Gram-negative microorganisms cannot be utilized in biomedical applications owing to the risk of immunogenic reactions caused by residual lipopolysaccharides (LPSs) that may co-extract with the biopolymer. Despite their lower yields, Gram-positive bacteria remain viable options for PHA production, particularly in the field of biomedical applications [20]. One such strain is B. megaterium B2, which was originally isolated by our research group [11]. These non-pathogenic, obligate aerobic microorganisms are Gram-positive and have the remarkable ability to tolerate high temperatures while accumulating PHA as a carbon and energy reserve, all without the need for specific nutritional requirements. Additionally, B. megaterium B2 can grow using low-cost substrates, making it a promising candidate for future PHA production [11].
The consumption demands of natural resources have caused a significant increase in the worldwide materials footprint. Between 2000 and 2017, there was a 70% increase in the total amount of natural resources extracted, reaching 92 billion metric tons [21]. In this context, enhancing materials' circularity in agro-industrial crops is critical for a low-carbon economy. Our focus is on improving Colombia's cacao production circularity by increasing biomass usage. To achieve this goal, we present the use of SCME and RMBC in developing a culture medium for PHAs biosynthesis using a native strain of B. megaterium B2. We investigated the B2 adaptation process to the CME-based media and scaled up the process to 4L reactors to study the influence of operational variables on PHAs production, yield, productivity, and microorganism kinetics.
Materials and Methods
Microorganism
Bacillus megaterium B2 is a native bacterial strain isolated from residual glycerol, a byproduct of palm oil transesterification for biofuel production in Colombia, by the Centro de Estudios e Investigaciones Ambientales (CEIAM) of Universidad Industrial de Santander as previously reported by Moreno et al. [11]. B. megaterium B2 was frozen in nutrient broth (peptone 15 g.L−1, yeast extract 3 g.L−1, and NaCl 6 g.L−1) plus glycerol (20%) as a cryoprotectant.
Substrates and Culture Media
Various culture media were tested: strain adaptation media, inoculum production media, and PHA production media. These were formulated with model substrates and CME-based substrates (SCME and RMBC) plus a basic salt medium. Initially, two standard culture media were tested as substrates for B. megaterium B2 using the basic salt medium (BSM) containing fructose (Merck, Darmstadt, Germany) “medium a” or glacial acetic acid (Merck, Darmstadt, Germany) “medium b”. These substrates were chosen as model compounds, considering the CME composition. Two CME-based culture media were formulated using SCME “medium c” and RMBC “medium d” as substrates. All culture media were heat sterilized in an autoclave (Tuttnauer 3850 EL) at 120 °C and 125 psi for 20 min. The sterilized media were stored at − 9 °C in airtight containers for further assays. The detailed media composition is shown in Table 1.
SCME, corresponding to 3 ± 0.2% of the cacao fruit weight, was collected as a lixiviate during the first two days of cacao beans fermentation, and RMBC was collected after fifteen days of bacterial cellulose batch production with CME-based media, as reported by Saavedra-Sanabria et al. [19]. The composition of SCME and RMBC was determined using standard protocols. Total solids in SCME and RMBC were determined by the AOAC 932.12 method (AOAC Official Method 932.12 Solids Soluble in Fruits and Fruit Products Refractometer Method). Moisture, ash, and protein percentages were determined using gravimetric methods AOAC 931.04, (AOAC Official Method 931.04 Moisture in Cacao Products Gravimetric Method), AOAC 972.15 (AOAC Official Method 972.15 Ash of Cacao Products) and AOAC 970.22 (AOAC Official Method 970.22 Total Nitrogen in Cacao Products). Low molecular weight acids (acetic, lactic, and citric), simple sugars (sucrose, fructose, and glucose), and total sugars measurements were performed by High-Performance Liquid Chromatography (HPLC). pH at 20 °C was measured with a pH meter (METTLER-TOLEDO AG 8603 Schwerzenbach) equipped with a glass electrode calibrated using buffer solutions of known hydrogen ion activity.
Strain Adaptation
A two-step strain adaptation was required considering that B. megaterium B2 was used to glycerol as a carbon source [11]. Initially, culture media 1a and 1b (Table 1) were tested to determine the microorganism performance under high sugar and acid concentrations. The tolerance of B. megaterium B2 to develop in SCME- or RMBC-containing culture media (1c, 1d, Table 1) was also tested. We conducted the adaptation tests in triplicate using a 250 mL batch reactor with a working volume of 50 mL. The inoculation was carried out with 1 mL of the B. megaterium B2 strain, and the culture was incubated aerobically at 34 °C ± 2 with constant shaking at 200 rpm for 24 h.
After 24 h, the growth of B. megaterium B2 was examined through plate count (surface spread method) using serial dilutions in a solid culture medium as described by Herbert [22]. The solid medium contained the same culture media plus agar–agar as a solidifying agent. Samples of 1 mL were collected hourly and inoculated onto the corresponding agar. Bacterial growth on the solid medium was regarded as a positive adaptation.
PHA Production Tests
After the adaptation stage was concluded, laboratory-scale bioprocesses were carried out in a 7.5-L reactor with an operational volume of 4 L using the culture media 3 containing model substrates (3a and 3b) and the CME-based culture media with substrates (3c and 3d) as shown in Table 1.
The bioprocess for PHA production began with strain activation. A cryopreserved vial containing 1 mL of B megaterium B2 was added to an Erlenmeyer with 50 mL of nutrient broth to prepare a pre-inoculum. The mixture was incubated at 34 °C ± 2 with constant shaking at 200 rpm for approximately 6–10 h until the microorganism reached the exponential growth stage. The inoculum was prepared by adding 4 mL of the pre-inoculum to an Erlenmeyer with 200 mL of medium 2 containing substrates 2a,2 b, 2c, 2d as described in Table 1. The mixture was incubated under the same pre-inoculum conditions. The total inoculum volume (204 mL) was transferred to the reactor containing 4 L of culture media 3 with the corresponding substrate used for PHA production. The following experimental conditions were kept constant during the fermentation process for bacterial comfort: pH 7.0 ± 0.2, temperature 34 °C ± 2, and dissolved oxygen (100%). The variables monitored during the bioprocess were biomass production, PHA production, and substrate consumption. For CME-based culture media the substrate consumption was determined in terms of acids and sugars consumption. All tests were performed by triplicate.
Experimental Design Assays
A response surface methodology utilizing a simplex-centroid mixture experimental design was employed to identify the optimal conditions for PHA production using culture media with various SCME:RMBC ratios, as shown in Table 2. Throughout the bioprocess, biomass production, PHA production, and substrate consumption were the measured variables, with all tests being conducted in triplicate. The response variables of product-substrate yield (Yp/s), product-biomass yield (Yp/x), volumetric productivity (Qp), and product accumulation (%) were determined based on intracellular PHA content relative to dry biomass, as per Eqs. 1 through 4.
where P = Product concentration in the sample, P0 = Product concentration at Zero ℎour, S = Substrate concentration in the sample, S0 = Substrate concentration at Zero ℎour, X = Biomass concentration in the sample, X0 = Biomass concentration at Zero ℎour, t = Hours
Statistical analysis
The Simplex-Centroid Design of the Response Surface Methodology consists of a triangle representing all possible mixtures of SCME, RMBC and BSM, in CME-based culture media. The statistical analysis of multiple regression to evaluate the effect of SCME and RMBC on CME-based culture media was carried out using Minitab software and required an analysis of variance (α = 0.05) of linear and quadratic regression models to select the most significant (p < 0.05) and best fitting (R2 > 0.85) model with which the response surface was constructed to determine the ranges of the three components that optimize Qp, Yp/s and Yp/x.
Analytical Methods
PHA Extraction
PHA extraction requires cell lysis because the biopolymer is accumulated intracellularly. PHA was extracted from the dried sediment using a 10 mL aqueous solution of sodium dodecyl sulfate (SDS, 0.5% w/v). This mixture was sonicated for 15 min at a temperature of 70 °C and thereafter centrifuged for 15 min at 7000 rpm to obtain the PHA [23]. The supernatant was discarded, and the PHA was washed twice with a mixture of H2O:EtOH (4:1) to eliminate impurities and cell debris. The sample was then dried at a temperature of 60 °C for 24 h and weighed to determine PHA production. PHA production was defined as the difference in weight difference between the empty tube and the tube filled with the pellet.
Quantification of Sugars and Acids
High-Performance Liquid Chromatography (HPLC) was performed using an Agilent 1100 series equipment, fitted with an automatic injector, a SUPELCOGEL ion exchanger C-610H (30 cm × 7.8 mm ID) column, and a refractive index detector (RID). Injection volumes of 20 μL, a mobile phase of 0.1% H3PO4 aqueous solution with a flow rate of 0.4 mL min−1, and oven and RID temperatures of 30 °C and 35 °C were used for HPLC analysis. Sugars and acids quantification was performed by external calibration using stock solutions (60 g.L−1) of glucose, fructose, and sucrose standards, and stock solutions (5 g.L−1) of acetic, citric, and lactic acids. The HPLC technique was used to characterize CME-based substrates and substrate consumption in the fermentations.
pH Measurement
pH at 20 °C was measured with a pH meter (METTLER-TOLEDO AG 8603 Schwerzenbach) equipped with a glass electrode calibrated using buffer solutions of known hydrogen ion activity.
Biomass and Product Quantification
Biomass production is reported as dry cell weight (DCW) [24]. The gravimetric method AOAC 931.04 was used to determine PHA production, while HPLC was used to track substrate consumption. 30 mL samples were collected every two hours and centrifuged at 7000 rpm for 15 min at 4 °C. The supernatant was recovered and stored at 4° C for substrate consumption measurements. The pellet was dried in an oven at 60 °C for 24 h for DCW determinations [24]. Biomass production was calculated as the weight difference between the empty tube and the tube containing the pellet.
PHA Characterization
Molecular characterization of PHA was performed by MALDI-TOF mass spectrometry. MS experiments were conducted on an UltrafleXtreme MALDI TOF-TOF instrument (Bruker Daltonics, Billerica, MA) equipped with a 1 kHz Smart Beam Nd:YAG laser (355 nm). Mass spectra were obtained in positive/reflectron and positive/linear ion modes, with delayed extraction set at 100 ns, an accelerating voltage of 25 kV. Instrument calibration was performed using a standard peptide mixture (leu-enkephaline, bradykinin, bombesin, and renin substrate) and polythiophene purchased from Sigma Aldrich (St. Louis, MO). A mixture of the PHA sample, matrix trans-2-[3-(4-tert-Butylphenyl)-2-methyl-2-propenylidene] malononitrile (DCTB), and Tetrahydrofuran (THF) were mixed using a pestle on an agate mortar to facilitate homogenization. A small sample fraction was deposited onto a MALDI stainless steel target. For each sample, 2000 spectra were accumulated. Mass spectrometric analysis was carried out using the FlexAnalysis software (Bruker Daltonics, Billerica MA, USA).
Additionally, PHA´s functional group analysis was carried out using a Shimadzu IR Prestige-21 Fourier Transform Infrared (IR-ATR) instrument with a 4000–500 cm−1 spectral range. Elemental composition was performed on a Thermo Scientific Model Flash 2000 analyzer. The PHA absorption spectrum was taken on a Thermo Scientific Evolution 300 spectrophotometer using a THF solution of the biopolymer. The thermal properties of the biopolymer were tested by differential scanning calorimetry (DSC) using a Mettler Toledo DSC 3 + equipment. 5.3 mg of sample was weighed and subjected to a heating ramp of 20.00 K/min from − 50 to 250 °C, using nitrogen as a carrier gas, with a flow rate of 50.0 mL/min, and a stabilization period of − 50.0 °C, 5.0 min air 50.0 mL/min. Thermogravimetric analysis (TGA) was performed in a Mettler Toledo TGA/DSC STARe instrument. 19.361 mg of sample was weighed and heated with a ramp of 20 °C/min up to 400 °C.
Results and Discussion
Substrate Characterization
Table 3 shows the composition of the SCME and RMBC which, compared to raw CME, are both acidic and contain a high concentration of dissolved organic matter in the form of sugars, acids, and proteins. Using HPLC analysis, we were able to quantify the amount of low molecular weight carbohydrates in RMBC and SCME. The original CME had a total carbohydrate content of up to 17.18 ± 6.44 wt % with average concentrations of fructose, glucose, and sucrose at 62.48 g.L−1, 58.80 g.L−1, and 19.43 g.L−1, respectively. As previously reported, the total carbohydrates and individual sugar content in CME can vary depending on factors such as cacao clone, season, and weather conditions such as rainfall [16]. As anticipated, SCME and RMBC display lower levels of sugar concentration in comparison to the original CME. SCME contains 17.1 and 29.2 g.L−1 of glucose and fructose, whereas RMBC has 22.3 and 25.4 g.L−1 of the same sugars. In RMBC, a small amount of sucrose (9.7 g.L−1) is still present, however, it is absent in SCME.
The lower sugar concentrations found in SCME and RMBC are a result of microbial populations consuming carbohydrates during cacao bean fermentation or BC production. It is interesting to note that the relatively high sucrose content remaining in RMBC after BC production can be attributed to the inability of the bacteria involved in cellulose biosynthesis to effectively degrade disaccharides, leading to the consumption of simple sugars such as glucose and fructose [25]. Conversely, during cacao bean fermentation, a wide range of environmental microorganisms with the necessary enzymatic machinery to metabolize complex sugars and convert them to glucose and fructose participate in the process. Despite the previous processes, the residual sugar amount in both SCME and RMBC is sufficient to fuel a subsequent bioprocess, as intended in this contribution.
Citric and acetic acid were present in RMBC and SMCE, with lactic acid additionally detected in the latter. Interestingly, the acid composition suggests that RMBC is derived from aerobic processes and SCME from anaerobic fermentation processes, which aligns with previous research [25]. It is worth noting that the acid concentration was higher in both RMBC and SCME compared to the original CME, where citric acid and acetic acid were present at 2.10 g.L−1 and 0.40 g.L−1, respectively. Furthermore, RMBC contained higher protein levels than SCME, which were both higher than CME's protein content of 0.29 g.L−1. The increase in protein content in CME is likely due to the presence of microbial biomass residues. Thus, with a high organic matter content, RMBC and SCME could be used as substrates for biopolymer biosynthesis. We hypothesize that B. megaterium B2 could potentially use these materials as a carbon source for PHA biosynthesis, accumulating it intracellularly as a carbon and energy reserve.
We selected fructose and acetic acid as model substrates for the standard culture media formulation during the adaptation stage. Fructose is the most abundant sugar in SCME and RMBC as shown in Table 2. On the other hand, despite being the most abundant in each medium, citric acid was not selected as a model substrate because it could affect PHA production by favoring the tricarboxylic acid (TCA) cycle pathway (Fig. 1). As depicted in Fig. 1, in the metabolic pathway for PHA biosynthesis the assimilated carbon is primarily utilized by glycolysis and the TCA cycle, while the remaining carbon is directed toward other metabolites, such as polymer production [26]. Therefore, the utilization of citric acid alone as a carbon source may cause metabolic inhibition for PHA production by favoring the TCA pathway [27]. Consequently, acetic acid was chosen as the model substrate since it is the second most abundant and is present in both substrates.

Metabolic pathway for PHA synthesis. Adapted from [28]
Strain Adaptation
Microbial growth can be divided into three phases: adaptation or lag phase, growth or exponential phase, and stationary phase. Figure 2 shows B. megaterium B2 growth plots using the model substrates (fructose, acetic acid) and the cacao fruit liquid residues (RMBC and SCME) for 16 h. By examining growth plots, we can estimate the length of the exponential phase, which is when the microorganism reaches its maximum growth rate. The duration of this phase is crucial in determining the incubation time for pre-inoculum preparation. For fructose and acetic acid, the exponential stage lasted 7 and 11 h, respectively, while for RMBC and SCME, it lasted 12 and 14 h. The growth of B. megaterium B2 was affected by the substrate model used, as depicted in Fig. 2a. Notably, biomass production was higher with acetic acid as compared to fructose.

Bacterial growth plots (as log CFU/ml) for B. megaterium B2 using model substrates a fructose and acetic acid and b cacao fruit residual substrates (RMBC and SCME). The bottom images depict B. megaterium B2 growth for 5, 6, 7, and 8 h on a solid medium using fructose as a substrate
This finding is in line with previous research conducted by Vu et al. [29] where a B. megaterium strain showed increased growth (2.5 OD600 nm) in a medium supplemented with acetic acid than in a medium supplemented with glucose (2.3 OD600 nm). This is because low-molecular-weight acids serve as direct metabolic precursors in the PHA biosynthetic pathway [30]. Vishnuvardhan et al. [31] reported the same effect for B. megaterium OU303A, which accumulated 62.43% of PHB in the presence of propionic acid, in contrast with 58.63% in the presence of glucose.
The growth rate of B. megaterium B2 was lower with fructose as the substrate as compared to acetic acid. However, the microorganism was able to adapt to the substrate successfully, albeit with a longer adaptation period. A microorganism's positive adaptation to a substrate involves its ability to modify its metabolic machinery to consume the available substrate. B. megaterium B2 was adapted to glycerol as a substrate and had not undergone a carbon source change process previously. Based on the results presented in Fig. 2a, we conclude that B. megaterium B2 has positively adapted to the model substrates.
After observing positive adaptation to the model substrates, we proceeded with the adaptation to RMBC and SCME, as illustrated in Fig. 2b. We noted that the exponential phase commenced faster with SCME, like the observed growth with the acetic acid model substrate. The concentration of total acids in SCME is higher (18.8 g.L−1) than in RMBC (6.8 g.L−1). The faster growth in SCME and acetic acid suggests a shorter latency phase with these substrates compared to RMBC and fructose. Based on the plots in Fig. 2b, we can affirm that B. megaterium B2 positively adapted to the SCME and RMBC substrates. The growth behavior of B megaterium with the cacao fruit liquid waste substrates could be due to a synergistic effect, between acids and sugars, on bacterial metabolism, unlike the effects of each carbon source individually. In a similar vein, Vishnuvardhan et al. observed that the B. megaterium OU303A strain accumulated higher amounts of PHA when propionic acid was added to the growth media containing glucose and glycerol than when the carbon sources were used separately. The addition of VFA (volatile fatty acids) supports PHA synthesis [31].
PHA Production
The dynamics of substrate consumption during 24 h of fermentation is shown in Fig. 3a. The microorganism demonstrated the ability to metabolize all substrates, albeit to varying degrees. For instance, B. megaterium B2 consumed 40% of fructose added to the model substrate culture medium. Noticeably, the consumption behavior for the culture medium with acetic acid as a model substrate follows a sinusoidal periodic shape with crests at 0, 8, and 16 h and valleys at 4 and 12 h (see inset in Fig. 3a). This trend indicates the ability of the microorganism to metabolize and produce acetic acid as a metabolic pathway byproduct. Thus, acetic acid acts as a metabolic intermediate and a substrate, considering the culture media's poor nutritional conditions. Figure 3b and c show low-to-negligible biomass and PHA production when using acetic acid, as a carbon source. The cells with acetic acid as a carbon source experienced starvation and used their PHA reserves as an alternative source for deficient growth [32].

a Substrate consumption (g.L−1), b Bacterial growth (g.L−1 DCW), c Biopolymer production (PHA g.L−1) by B. megaterium B2 versus time. The insert shows the pellet and the supernatant after sample centrifugation
When it comes to the cacao liquid residues (SCME and RMBC), the microorganism exhibits two distinct consumption patterns during the fermentation process, as depicted in Fig. 3a. The sugar consumption profile follows a sigmoidal curve with an adaptation phase or lag phase of five hours for RMBC, followed by rapid substrate consumption after ten hours. This pattern is consistent with the exponential growth of the microorganism presented in Fig. 3b, indicating the behavior of a primary metabolite as explained by Shuler and Kargi [33]. Thus, after 24 h of fermentation, B. megaterium B2 consumed 86% of the total sugars present in the culture medium containing RMBC. Conversely, in the SCME-containing culture medium, sugar consumption followed a linear pattern, with B. megaterium B2 consuming 50% of the total sugars after 24 h. These results lead us to conclude that CME-based culture media provide the necessary nutrients for the microorganism's growth and biopolymer production.
After achieving the microorganism's successful adaptation to all substrates, we proceeded to investigate PHA production in a batch mode using a 7.5 L bioreactor. Figure 3b depicts the biomass production of B. megaterium B2 with all substrates considered in this work. SCME resulted in higher biomass production (3.35 g.L−1), followed by RMBC (3.06 g.L−1), fructose (0.56 g.L1), and acetic acid (0.54 g.L−1). The plots of PHA production vs. time in Fig. 3c show a pattern comparable to that of biomass production in Fig. 3b, indicating that PHA acts as the primary metabolite of the fermentation process. This type of metabolite exhibits concurrent substrate consumption and product formation [33]. Additionally, the data in Fig. 3c indicates that PHA production dramatically declines when model substrates are employed, as opposed to cacao fruit liquid residues. The accumulation of PHAs in the B. megaterium B2 cells reached 72% for RMBC and 63% for SCME, in contrast with 4% and 30% for acetic acid and fructose, respectively.
PHA Characterization
Figure 4 shows the positive mode reflectron MALDI-MS spectrum, in the range m/z 1000 to 6000, for a PHA sample produced by B. megaterium B2 using DCTB as a matrix. Unlike proteins, MALDI showed PHA signals as sodium adducts because the biopolymer does not acquire charge through protonation. Typically, MALDI analysis of polymers (of both natural and synthetic origin) requires cationizing agents, such as Na+, K+, and Ag+ ions [34, 35].

PHA mass spectrum in reflectron mode using MALDI-TOF
MALDI MS data for the PHA sample shows single-charged periodic signals in the medium-range mass region with a mass difference of 86 Da. A PHA with a repeating unit of 86 Da can be assigned the molecular formulae (C4H6O2)n; thus, revealing a homopolymer of the PHB type as the primary product of B. megaterium B2 metabolism. To confirm the molecular formulae assignment, we used isotopic pattern analysis. The insert in Fig. 4 shows the isotopic pattern for the polymeric structure corresponding to (C4H6O2)24-Na+ at m/z 2105. The agreement between the theoretical and the experimental isotopic patterns confirms the biopolymer's identity. Also, from the molecular weight distribution, we calculated average molecular mass values of Mn = 1704.25, Mw = 1837.88, and polydispersity (PD) of 1.08.
The IR spectrum in Fig. 5 displays signals for the characteristic functional groups in PHB. For instance, the strong band at 1724 cm−1 corresponds to the C=O stretching vibration of the carbonyl group in esters [36]. The signal at 1542 cm−1 corresponds to the C–O stretching vibration. The signals at 1455 cm−1 and 1376 cm−1 are the asymmetric bending vibrational modes of the methyl (–CH3) and methylene (–CH2) groups [37]. The bands at 1270, 1231, and 1142 cm−1 are characteristic of the asymmetric stretching vibration of the C–O–C bond in the amorphous phase, the asymmetric stretching of the C–C–O bond in the crystalline phase, and the rocking of methylene CH2 in the C–C–O backbone in the crystalline phase respectively [38, 39]. The characteristic C–O bond stretching signal in esters appears at 1056 cm−1 [40]. The signals around 2923 and 2852 cm−1 are assigned to the symmetric and asymmetric axial deformations of the methylene –CH2 groups in the PHB chain; these signals were previously reported [36]. Also, the broad signal at 3300 cm−1 indicates moisture in the sample.

a FT-IR spectrum, b theoretical and experimental elemental composition, c UV–vis spectrum, d DSC curve, and e thermal degradation TGA and DTG curves of PHB from cacao fruit liquid residues
Figure 5b compares PHB's theoretical and experimental elemental composition according to the chemical formula (C4H6O2)n. The differences between the expected and actual results arise mainly from nitrogen in the sample. Impurities, such as proteins and cell debris left on the polymer after extraction, might be responsible for the presence of nitrogen (up to 6.7 wt %) in the PHB sample. The presence of organic molecules containing nitrogen, thus affects the percentages of carbon, oxygen, and hydrogen. Figure 5c shows the UV–vis absorption properties of the polymer. The UV region of the spectrum exhibits a band at 210 nm assigned to electronic transitions of the type π → π*, characteristic of carbonylic compounds [41]. Figure 5d shows the PHB's thermal behavior. The DSC melting curve for the PHB sample shows a sharp melting peak (endothermic event) with an onset of 165.6 °C corresponding to the polymer's melting point (Tm). Giedraitytė et al. [42] reported a similar value of 168.8 °C for PHB produced by Geobacillus sp using glucose as a carbon source. Also, Drusilla et al. reported a range of 161.71 to 165.09 °C for the melting point of PHB produced by Burkholderia cepacia BPT1213 using residual glycerol (2 to 9% (v/v)) as a carbon source [43]. We observe a straight line for the melting peak, indicating a relatively pure sample. However, the exothermic bump after the endothermic peak suggests melting followed by decomposition, probably caused by impurities in the PHB, as discussed above. The PHB produced by B megaterium B2 exhibit a heat of fusion (△Hm) of 74.33 J/g, like that reported in the literature. For instance, PHB produced from residual glycerol exhibits a heat of fusion in the range of 50.36 to 69.04 J/g [43]. Oliveira et al. [44] reported a △Hm of 66.9 J/g for PHB produced by fermentation with Cupriavidus necátor using sugar cane residues and a △Hm of 77.6 J/g for a commercial PHB sample.
The TGA plot, Fig. 5e, shows the thermal decomposition of a PHB sample under an air atmosphere and a heating rate of 20 °C/min. There are two main mass-loss events; the first is associated with solvent loss and the second with polymer decomposition. The latter corresponds to the process of random chain cleavage at the C=O and C-O ester bonds by β-elimination, leading to the formation of crotonic acid [45]. The absence of shoulders in the TGA profile suggests a single type homopolymer with no impurities or copolymers. PHB's main decomposition event ranges from 250 to 400 °C with an overall mass loss of 70.01%. Polymer decomposition starts at 258.20 °C, with a maximum decomposition rate at 270.62 °C (DTG). Depolymerization reactions and hydrolysis occur at this stage, resulting in low molecular weight products. Previous studies reported single-stage thermal degradations above 270 °C for pure PHB [46]. Pradhan et al.[45] reported a maximum decomposition temperature of 248 °C for PHB synthesized by B. megaterium from fructose as a carbon source.
Effect of Culture Media Composition on the Bioprocess Performance
The collective outcomes of the experimental design are presented in Table 4. Microorganism growth and PHA production are considered positive adaptations. B. megaterium B2 demonstrated metabolic adaptation to culture media 4, 6, 7, 8, 9, and 10. However, no growth was observed in culture media 1, 2, and 3, which corresponded to 100% SCME, RMBC, and basic salt medium, respectively. The lack of growth in these media could be attributed to the absence of essential nutrients. Similarly, the microorganism did not thrive in culture media 5 (50:50% v/v RMBC:SCME), which lacked the basic salt medium. Thus, it can be concluded that the micronutrients provided by the BSM are crucial for B. megaterium B2 development. Culture media 7 (33:33:33% v/v BSM:RMBC:SCME) yielded the highest biomass (65.89 g.L−1) and PHA accumulation (34.25 g.L−1).
Regarding bioprocess performance, culture media 7 (33:33:33% v/v BSM:RMBC:SCME) displays the highest rate of production per hour (0.50 g.L−1.h−1) in comparison to the other media we tested. Conversely, mixture 9, made up of 17:67:17% v/v BSM:RMBC:SCME, demonstrated the highest PHA production per gram of substrate at 0.97 g/g, but exhibited low volumetric productivity of only 0.19 g.L−1.h−1. Media 6, 8, and 10, which contained the highest SCME proportions, delivered the lowest PHA yields and productivities, potentially due to the acid composition of the SCME. As previously discussed, an excessive concentration of citric acid in the substrate can lead to metabolic inhibition in the PHA production pathway (see Fig. 1) [27]. Therefore, the assimilated carbon source in SCME was utilized for biomass production and survival, rather than for product generation. Concerning substrate consumption, B. megaterium B2 utilized 95.54% of the carbon source in medium 8 and 68.06% in medium 7. From our observations, we can conclude that RMBC has a positive impact on both biomass and PHA yield. In culture media where high proportions of RMBC are present (i.e., media 4, 7, and 9), we notice an increase in yields, PHA accumulation, and productivity. This behavior can be attributed to the favorable content of low molecular weight carbohydrates in RMBC, which aids in the PHA production pathway [47].
The experimental design analysis was conducted using the Minitab® Statistical Software [48]. Contour graphs in Fig. 6 were created based on the statistical analysis from the product yield in substrate, product in biomass, and volumetric productivity. The contour plots 6b and 6c demonstrate that a binary mix between BSM and RMBC has a significant impact on Yp/s , and \({Q}_{p}\) with an extension to the central point. On the other hand, the contour plot 6d indicates that Yp/x is significantly influenced by a binary mixture of BSM and SCME with an extension to the central point. As expected, the lowest values for the response variables are found in the corners of the triangle representing the pure culture media. Therefore, we can conclude that culture media 7, which contains equal volumetric amounts of BSM:RMBC:SCME, provides the best outcome for the response variables and should be considered for future research.

Statistical analysis for a simplex experimental design and mixing contour plots for b \({Q}_{p}\), c \(Y_{{{\raise0.5ex\hbox{$\scriptstyle p$} \kern-0.1em/\kern-0.15em \lower0.25ex\hbox{$\scriptstyle s$}}}}\) , and d \(Y_{{{\raise0.5ex\hbox{$\scriptstyle p$} \kern-0.1em/\kern-0.15em \lower0.25ex\hbox{$\scriptstyle x$}}}}\)
In Table 5, we compare the performance of our process with several authors who used other B. megaterium strains, Mohanrasu et al. [49], Dañez et al. [50], Ali et al. [51], and Manikandan et al. [52] reached PHA productivity values between 0.08 and 5.82 g.L−1.h−1 and yields between 0.001 and 0.140 g.g−1. We observed a maximum PHA productivity of 0.50 g.L−1.h−1 and yields 0.63 g.g−1 with culture media 7, which compares favorably with the data available in the literature.
To our knowledge, there are no studies on cacao liquid wastes as substrates for PHA production with B megaterium. However, various agro-industrial residues have been tested for PHA production. For instance, Sukruansuwan et al. [53] used residues from the canned pineapple industry for the growth of Cupriavidus necátor A-04 growth and PHA production. Also, Manikandan et al. [52] employed carob pods and Ralstonia eutropha for PHA biosynthesis; and Ali et al. [51] used cantaloupe waste extract and a B. megaterium strain (TISTR 1814) isolated from soil.
On another note, comparison of PHA production (g.L−1) with other works is also illustrative. Mohanrasu et al. [49], Manikandan et al. [52], Sukruansuwan et al. [53], and Manikandan et al. [52]report PHA productions of 5.65 g.L−1, 5.82 g.L−1, 8.16 g.L−1, and 12.2 g.L−1respectively. These values are much higher than the productions reported in this work. However, the volumetric productivities of the literature reports cited above are much lower, indicating that a higher PHA production is achieved at the expense of longer fermentation times which translates into higher operational and energy expenses. We consider that the production of biopolymers using cacao fruit liquid residues is a strategy that can generate positive environmental, economic, and social impacts by increasing crop circularity, particularly in tropical countries such as Colombia.
Conclusions
The findings of this study make a significant contribution to the cacao industry and the advancement of novel materials. The bioprocess implemented effectively incorporates liquid residues of the cacao fruit processing (SCME and RMBC) into synthesizing biodegradable plastics utilizing the strain Bacillus megaterium B2.
Statistical analysis shows that maximum response variables (Qp, Yp/x and Yp/s) are individually influenced by binary mixtures RMBC:BSM and SCME:BSM, and that equal volumetric amounts of BSM:RMBC:SCME (33:33:33), provide the best outcome for all the response variables.
Biopolymer characterization using MS-MALDI-TOF and IR techniques confirmed its structure as a polyhydroxybutyrate (PHB) type. Further analysis using elemental analysis, UV–VIS, TGA, and DSC confirmed its thermal and absorption properties.
Data Availability
Regarding Data Availability, the authors have not included a data availability in the manuscript, however the data will be made available on request sent to the corresponding author.
References
PlasticsEurope (2022) Plastics – the Facts. 81
Rakkan T, Chana N, Sangkharak K (2023) Utilization of textile wastewater as a substrate for polyhydroxyalkanoate (PHA) and enhanced production by mutant enterobacter. J Polym Environ 31:677–687. https://doi.org/10.1007/s10924-022-02563-6
Al Battashi H, Al-Kindi S, Gupta VK, Sivakumar N (2021) Polyhydroxyalkanoate (PHA) production using volatile fatty acids derived from the anaerobic digestion of waste paper. J Polym Environ 29:250–259. https://doi.org/10.1007/s10924-020-01870-0
Chen GQ, Wu Q (2005) The application of polyhydroxyalkanoates as tissue engineering materials. Biomaterials 26:6565–6578. https://doi.org/10.1016/j.biomaterials.2005.04.036
dos Santos AJ, Oliveira Dalla Valentina LV, Hidalgo Schulz AA, Tomaz Duarte MA (2017) From Obtaining to degradation of PHB: material properties. Part I Ing Cienc 13:269–298
Pryadko AS, Botvin VV, Mukhortova YR et al (2022) Core-shell magnetoactive PHB/gelatin/magnetite composite electrospun scaffolds for biomedical applications. Polymers (Basel). https://doi.org/10.3390/polym14030529
Winnacker M (2019) Polyhydroxyalkanoates: recent advances in their synthesis and applications. Eur J Lipid Sci Technol 121:1–9. https://doi.org/10.1002/ejlt.201900101
Mizuno K, Ohta A, Hyakutake M et al (2010) Isolation of polyhydroxyalkanoate-producing bacteria from a polluted soil and characterization of the isolated strain Bacillus cereus YB-4. Polym Degrad Stab 95:1335–1339. https://doi.org/10.1016/j.polymdegradstab.2010.01.033
Vega-Castro O, León E, Arias M et al (2021) Characterization and production of a polyhydroxyalkanoate from cassava peel waste: manufacture of biopolymer microfibers by electrospinning. J Polym Environ 29:187–200. https://doi.org/10.1007/s10924-020-01861-1
Ertan F, Keskinler B, Tanriseven A (2021) Exploration of cupriavidus necator ATCC 25207 for the production of poly(3-hydroxybutyrate) using acid treated beet molasses. J Polym Environ 29:2111–2125. https://doi.org/10.1007/s10924-020-02020-2
Moreno P, Yañez C, Cardozo NSM et al (2015) Influence of nutritional and physicochemical variables on PHB production from raw glycerol obtained from a colombian biodiesel plant by a wild-type Bacillus megaterium strain. N Biotechnol 32:682–689. https://doi.org/10.1016/j.nbt.2015.04.003
Suhazsini P, Keshav R, Narayanan S et al (2020) A Study on the Synthesis of Poly (3-hydroxybutyrate-co-3-hydroxyvalerate) by Bacillus megaterium utilizing cheese whey permeate. J Polym Environ 28:1390–1405. https://doi.org/10.1007/s10924-020-01687-x
Mohammed S, Ray L (2022) Polyhydroxyalkanoate recovery from newly screened Bacillus sp. LPPI-18 using various methods of extraction from Loktak Lake sediment sample. Journal of Genetic Engineering and Biotechnology. https://doi.org/10.1186/s43141-022-00392-7
(ICCO) International Cocoa Organization (2021) Cocoa year 2020/2021
FEDECACAO (2022) Producción mundial de cacao en grano por continentes. Cifras periodo cacaotero (Año Cacaotero: 2020/2021). https://www.fedecacao.com.co/post/a%C3%B1o-cacaotero-2020-2021-el-de-mayor-producci%C3%B3n-de-cacao-en-la-historia-de-colombia. Accessed 17 Oct 2022
Vergara-Mendoza M, Martínez GR, Blanco-Tirado C, Combariza MY (2022) Mass Balance and Compositional Analysis of Biomass Outputs from Cacao Fruits. Molecules. https://doi.org/10.3390/molecules27123717
Rojas F, Sacristan E (2013) Guía ambiental para el Cultivo de cacao. Ministerio de Agricultura y Desarrollo Rural
Luna F, Crouzillat D, Cirou L, Bucheli P (2002) Chemical composition and flavor of Ecuadorian cocoa liquor. J Agric Food Chem 50:3527–3532. https://doi.org/10.1021/jf0116597
Saavedra-Sanabria OL, Durán D, Cabezas J et al (2021) Cellulose biosynthesis using simple sugars available in residual cacao mucilage exudate. Carbohydr Polym 274:1–12. https://doi.org/10.1016/j.carbpol.2021.118645
Choonut A, Prasertsan P, Klomklao S, Sangkharak K (2020) Study on mcl-PHA production by novel thermotolerant gram-positive isolate. J Polym Environ 28:2410–2421. https://doi.org/10.1007/s10924-020-01779-8
UN. United Nations (2022) The global materials footprint. /sdgs/report/2019/goal-12/. Accessed 10 Oct 2022
Herbert RA (1990) Methods for enumerating microorganisms and determining biomass in natural environments. Methods Microbiol 22:1–39. https://doi.org/10.1016/S0580-9517(08)70238-1
Arikawa H, Sato S, Fujiki T, Matsumoto K (2017) Simple and rapid method for isolation and quantitation of polyhydroxyalkanoate by SDS-sonication treatment. J Biosci Bioeng 124:250–254. https://doi.org/10.1016/j.jbiosc.2017.03.003
Herbert D, Phipps PJ, Strange RE (1971) Chemical analysis of microbial cells. Methods Microbiol 5:209–344. https://doi.org/10.1016/S0580-9517(08)70641-X
Madigan MT, Martinko JM, Parker J (2015) Brock Biología de los microorganismos, 14th edn. Prentice Hall, Madrid
Hendry JI, Prasannan C, Ma F et al (2017) Rerouting of Carbon Flux in a Glycogen Mutant of Cyanobacteria Assessed via Isotopically Non-Stationary 13 C Metabolic Flux Analysis. Biotechnol Bioeng 114:2298–2308. https://doi.org/10.1002/bit.26350/abstract
Monshupanee T, Chairattanawat C, Incharoensakdi A (2019) Disruption of cyanobacterial γ-aminobutyric acid shunt pathway reduces metabolites levels in tricarboxylic acid cycle, but enhances pyruvate and poly(3-hydroxybutyrate) accumulation. Sci Rep. https://doi.org/10.1038/s41598-019-44729-8
Tsuge’ T, (2002) Metabolic Improvements and Use of Inexpensive Carbon Sources in Microbial Production of Polyhydroxyalkanoates. J Biosci Bioeng. https://doi.org/10.1016/S1389-1723(02)80198-0
Vu DH, Wainaina S, Taherzadeh MJ et al (2021) Production of polyhydroxyalkanoates (PHAs) by Bacillus megaterium using food waste acidogenic fermentation-derived volatile fatty acids. Bioengineered 12:2480–2498. https://doi.org/10.1080/21655979.2021.1935524
Szacherska K, Oleskowicz-Popiel P, Ciesielski S, Mozejko-Ciesielska J (2021) Volatile fatty acids as carbon sources for polyhydroxyalkanoates production. Polymers (Basel) 13:1–21. https://doi.org/10.3390/polym13030321
Vishnuvardhan Reddy S, Thirumala M, Mahmood SK (2009) Production of PHB and P (3HB-co-3HV) biopolymers by Bacillus megaterium strain OU303A isolated from municipal sewage sludge. World J Microbiol Biotechnol 25:391–397. https://doi.org/10.1007/s11274-008-9903-3
Fradinho JC, Oehmen A, Reis MAM (2014) Photosynthetic mixed culture polyhydroxyalkanoate (PHA) production from individual and mixed volatile fatty acids (VFAs): Substrate preferences and co-substrate uptake. J Biotechnol 185:19–27. https://doi.org/10.1016/j.jbiotec.2014.05.035
Shuler M, Kargi F (2002) Bioprocess Engineering : Basic Concepts, 2nd edn. Prentice Hall, Izmir
Chen R, Li L (2001) Lithium and transition metal ions enable low energy collision-induced dissociation of polyglycols in electrospray ionization mass spectrometry. J Am Soc Mass Spectrom 12:832–839
Memboeuf A, Vékey K, Lendvay G (2011) Structure and energetics of poly(ethylene glycol) cationized by Li+, Na+, K+ and Cs+: a first-principles study. Eur J Mass Spectrom 17:33–46
Quines LKM, Ienczak JL, Schmidt M et al (2015) Extraction of poly(3-hydroxybutyrate), produced by Cupriavidus necator, with propylene carbonate. Quim Nova 38:214–220. https://doi.org/10.5935/0100-4042.20140297
Xiao N, Jiao N (2011) Formation of polyhydroxyalkanoate in aerobic anoxygenic phototrophic bacteria and its relationship to carbon source and light availability. Appl Environ Microbiol 77:7445–7450. https://doi.org/10.1128/AEM.05955-11
Martínez-Herrera RE, Alemán-Huerta ME, Almaguer-Cantú V et al (2020) Efficient recovery of thermostable polyhydroxybutyrate (PHB) by a rapid and solvent-free extraction protocol assisted by ultrasound. Int J Biol Macromol 164:771–782. https://doi.org/10.1016/j.ijbiomac.2020.07.101
Porter M, Yu J (2011) Monitoring the in situ crystallization of native biopolyester granules in Ralstonia eutropha via infrared spectroscopy. J Microbiol Methods 87:49–55. https://doi.org/10.1016/j.mimet.2011.07.009
Cervantes-Uc JM, Catzin J, Vargas I et al (2014) Biosynthesis and characterization of polyhydroxyalkanoates produced by an extreme halophilic bacterium, Halomonas nitroreducens, isolated from hypersaline ponds. J Appl Microbiol 117:1056–1065. https://doi.org/10.1111/jam.12605
Pretsch E, Bühlmann P, Badertscher M (2009) Structure Determination of Organic Compounds, edition 4t. Springer, Berlín - German
Giedraitytė G, Kalėdienė L Purification and characterization of polyhydroxybutyrate produced from thermophilic Geobacillus sp. AY 946034 strain. Journals of the Lithuanian Academy of Sciences - CHEMIJA
Drusilla Wendy YB, Nor Fauziah MZ, Siti Baidurah Y et al (2022) Production and characterization of polyhydroxybutyrate (PHB) BY Burkholderia cepacia BPT1213 using waste glycerol as carbon source. Biocatal Agric Biotechnol. https://doi.org/10.1016/j.bcab.2022.102310
Oliveira FC, Dias ML, Castilho LR, Freire DMG (2007) Characterization of poly(3-hydroxybutyrate) produced by Cupriavidus necator in solid-state fermentation. Bioresour Technol 98:633–638. https://doi.org/10.1016/j.biortech.2006.02.022
Pradhan S, Dikshit PK, Moholkar VS (2018) Production, ultrasonic extraction, and characterization of poly (3-hydroxybutyrate) (PHB) using Bacillus megaterium and Cupriavidus necator. Polym Adv Technol 29:2392–2400. https://doi.org/10.1002/pat.4351
Nery TBR, dos Santos ZIG, José NM (2018) Development and characterization of polyhydroxybutyrate and banana fiber biocomposites. Revista Materia. https://doi.org/10.1590/s1517-707620180004.0591
Fadipe TO, Jamil N, Lawal AK (2021) Biosynthesis and Characterization of Poly-(3)-hydroxyalkanoic Acid by Bacillus megaterium SF4 Using Different Carbohydrates. Microbial Polymers. Springer, Singapore, pp 109–129
Minitab LLC (2021) Minitab 18
Mohanrasu K, Rao RGR, Dinesh GH et al (2020) Optimization of media components and culture conditions for polyhydroxyalkanoates production by Bacillus megaterium. Fuel. https://doi.org/10.1016/j.fuel.2020.117522
Dañez JCA, Requiso PJ, Alfafara CG et al (2020) Optimization of Fermentation Factors for Polyhydroxybutyrate (PHB) Production Using Bacillus megaterium PNCM 1890 in simulated Glucose-Xylose Hydrolysates from agricultural residues. Philipp J Sci 149:163–175
Ali HH, Rehman ZU, Basheer S et al (2020) Prodcution of Polyhydroxy Butyrate (PHB) from Soil Bacterium (Bacillus megaterium TISTR 1814) with Cantaloupe Waste Extract as Potential Carbon Source. J Med life Sci. https://doi.org/10.31580/pjmls.v3i1.1293
Manikandan NA, Pakshirajan K, Pugazhenthi G (2020) A closed-loop biorefinery approach for polyhydroxybutyrate (PHB) production using sugars from carob pods as the sole raw material and downstream processing using the co-product lignin. Bioresour Technol. https://doi.org/10.1016/j.biortech.2020.123247
Sukruansuwan V, Napathorn SC (2018) Use of agro-industrial residue from the canned pineapple industry for polyhydroxybutyrate production by Cupriavidus necator strain A-04. Biotechnol Biofuels. https://doi.org/10.1186/s13068-018-1207-8
Ahmady-Asbchin S, Rezaee H, Safari M et al (2020) Production and optimization of polyhydroxybutyrate (PHB) from Bacillus megaterium as biodegradable plastic. Eur J Biol Res 10:26–34. https://doi.org/10.5281/zenodo.3711400
Acknowledgements
We thank Guatiguará Technological Park and the Central Research Laboratory Facility at Universidad Industrial de Santander for infrastructural support. We acknowledge financial support from Ministerio de Ciencia, Tecnología e Innovación (Minciencias – Colombia) Grant 70652, Program 1102-852-70114 (2021), and SGR-Gobernación de Santander Grant BPIN 2020000100251. The funding sources had no involvement in study design, data collection/analysis/interpretation, writing, or deciding where to submit the article for publication.
Funding
Open Access funding provided by Colombia Consortium. Ministerio de Ciencia, Tecnología e Innovación (Minciencias–Colombia) Grant 70652, Program 1102-852-70114 (2021), and SGR-Gobernación de Santander Grant BPIN 2020000100251.
Author information
Authors and Affiliations
Contributions
SJSR: Investigation, Methodology, Validation, Visualization, Writing—Original Draft. MAGS: Investigation, Methodology, Validation, Visualization, Writing—Original Draft. MQS: Conceptualization, Formal Analysis, Supervision, Project Administration, Data Curation, Writing—Original Draft CBT: Conceptualization, Resources, Funding Acquisition. MYC: Conceptualization, Formal Analysis, Supervision, Writing—Review and Editing, Funding Acquisition.
Corresponding authors
Ethics declarations
Competing Interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Quintero-Silva, M.J., Suárez-Rodríguez, S.J., Gamboa-Suárez, M.A. et al. Polyhydroxyalkanoates Production from Cacao Fruit Liquid Residues Using a Native Bacillus megaterium Strain: Preliminary Study. J Polym Environ 32, 1289–1303 (2024). https://doi.org/10.1007/s10924-023-03018-2
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10924-023-03018-2