
Abstract
Biodiversity and identification facilitate a prioritization of determination basis for plant evolutionary, phytogeography, biodiversity and conservation of plant genetic resources. High-throughput phenotyping platforms (HTPP) provide more attractive for genotyping for biodiversity evaluation. Descriptor lists include 51 attributes applied on all growth-stages of 53 C. olitorius accessions plus C. capsularis, C. trilocularis, C. tridens, and C. depressus. Three main groups were split. Variance decomposition was estimated within class of 68.87% and between classes of 31.13%. Major distance recorded 18.41 between class 1 (wild taxa only) and class three (18 accessions of C. olitorius). In contrast, the minor distance scored 8.3 between class 2 (35 accessions) and class 3 (18 accessions). Wild taxa with their rarity distribution and habitats prioritize highly conservation due to the association of evolutionary role and importance of genetic improvement. Conservation strategies applied to PGR prioritization are influenced by the distribution frequency and habitat specificity. The results are useful information on genetic diversity at the Egyptian Gene Bank, which could be used for C. olitorius genetic improvement programs and sustainable utilization.
Similar content being viewed by others


Introduction
The Corchorus (Family Malvaceae Juss.; formerly Tiliaceae) L., Sp. Pl. 1:529 (1753) is distributed in tropical and subtropical regions (Whitehouse et al. 2001). This genus (2n = 14; genome size of olitorius = – 1,250 Mb) has a broad sense of biodiversity, variability and adaptability. The Global Biodiversity Information Facility (http://www.gbif.org) has reported 215 species, subspecies, varieties, and forms belonging to the genus Corchorus. The precise number of good species, approximately 100 (Saunders 2006), is significantly C. olitorius and C. capsularis that commercially proliferated over the world (Sinha et al. 2011). Africa embraces about 50 species (Aluka 2006–2008: http://www.aluka.org), including olitorius and capsularis, and therefore represents the chief center of diversity and origin have been reported (Edmonds 1990; Plants of South Africa 2009: http://www.biodiversityexplorer.org/plants/malvaceae/corchorus.htm). They have various utilizations as raw material for industry of pulp, paper and fiber, and a source of biofuel (Paul 2008), dietary supplements and therapeutic uses (Chopra et al. 1986; Sen 2002; Hazra and Saha 2004; Natesh 2001). The primary five taxa of Corchorus are in Egypt, which are olitorius, tridens, capsularis, trilocularis, and depressus (Boulos 2000, 2009). The taxa of tridens, trilocularis, and depressus characterize resistance to the abiotic stresses. Its genetic diversity has been made safe because of its strong sexual incompatibility barrier between the most cultivated taxa of olitorius and capsularis and secondary gene pools (La Farge et al. 1997). Thus, this is due to very limited genetic variance regarding the adaptability to different agronomic environments and susceptibility to diseases and pests. The species olitorius, a leafy vegetable crop, is still cultivated widely in Egypt; however, not utilized for genetic improvement and breeding new lines/cultivars. Therefore, C. olitorius falls in a broad basket of underutilized species despite its nutritionally rich adaptability to low-input agriculture and sowing over 200 years ago in Egyptian agro-climatic districts (El-Wakeel 2007; Mahdy 2012). Only three off-types nominated Balady (meaning local), Falahy and Siwy have been identified, which vary in some clearly morphological attributes (El-Wakeel 2007). More collections with introgression new various Corchorus genetic resources (GR) will help breeders select and breed new promised ones with high-yielding (El-Wakeel 2007; Mahdy 2012). These collections will help to find more Corchorus mutants and off-types that have not been collected yet due to humans' lack of selection applications for a longer time (Mahdy 2012).
The high-throughput phenotyping platform (HTPP) provides more accurate information through a comprehensive valuation of morphological traits of interest (Finkel 2009). Various HTPP approaches give them more engaging for genotyping (Agarwal et al. 2008). It requires more available GRs for utmost crops and species to study the HTPP (Rizk and El-Sharabasy 2008; El-Sharabasy and Rizk 2019). The HTPPs have witnessed more continual progress in molecular marker technologies and diversity in technologies of array-based markers, which permit the identification of molecular markers appropriate for distinguishing various GRs. The Quantitative trait loci (QTLs) are used in genome selection in linking disequilibrium with at least a single marker (Goddard and Hayes 2007). The combination of genomic selection and high-throughput phenotyping has revolutionized genetic improvement programs by bringing superior accuracy in selecting the new promising genotypes (Ingvarsson and Street 2011). Similarly, the progress in the field of HTPPs made it possible to procure exact data for various attributes.
The assessment of Corchorus biodiversity conserved in the National Gene Bank (NGB) comprehensively evaluated the 51 morphological attributes. Corchorus GRs comprising 53 C. olitorius accessions and its wild relatives of C. tridens, C. trilocularis, and C. depressus that grows in Egypt subjected to the high-throughput phenotyping platform (HTPP) to make possible new promised varietal breeding in the future. We also attempt to highlight the genepools of wild Corchorus growing in Egypt and focus on taxonomic placement, which enables to utilize for various multi-purposes.
Materials and methods
Base line data collection
A total of 53 olitorius accessions and their wild relatives of tridens, trilocularis, capsularis, and depressus was collected from all naturalized agro- and ecological regions of Egypt. These were conserved under ex situ conditions at National Gene Bank (NGB), Agricultural Research Center (ARC), Egypt, and cultivated in the experimental field of National Gene Bank (NGB), Agricultural Research Centre (ARC), Giza, Egypt (Table S1). Species were validated for taxonomic identification via herbarium specimens at Herbarium of Botany Dept., NGB, Egypt. Proliferation was conducted on a board dimension 1 m × 0.5 m with a space of 0.2 m × 0.3 m. A distance of 0.5 m was set between two successive boards. Organic manure was applied at the time of plowing to fertilize the soil.
Corchorus genetic resources characterization
Descriptors including 51 attributes applied to all growth-stages of plant according to Loumerem and Alercia (2016), the Draft National Test Guidelines for testing Distinctness, Uniformity and Stability (DUS) for Jute (Kumar and Mahapatra 2004), quantitative attributes from AVRDC jute descriptors (2008), and various reports (Ghosh et al. 2012). All attributes follow Stearn (1995) terminologically. Attributes are listed in Table S2.
Observations were recorded on 10 random healthy in each accession at specified stage of plant growth when the attributes had been expressed fully. Quantitative attributes were scored as an average of 10-random accessions.
Plant phenotyping system
Plant phenotyping was performed with hand-held instruments to measure all growth stages of GRs differed genetically, from the soil line to the terminal shoot of the plant with a calibrated bar-coded ruler. A single plant for each accession was chosen visually to perform more detailed measurements as mentioned previously. The images of plants were captured by Kinect v2 camera (Kinect-v2, Microsoft, Redmond, WA, USA) which acquires RGB color (1920 × 1080 pixels), and depth (512 × 424 pixels), at a rate of up to 30 FPS. Plant phenotyping was done by high-throughput morphology-based phenotyping applied to living plants. The phenotyping was carried out on living plants according to field-based HTPP by employing the descriptors for Corchorus (AVRDC jute descriptors 2008; Ghosh et al. 2012; Kumar and Mahapatra 2004). They were provided and fed into the platform of the NGB database.
Data collection and analysis
Data were described morphologically and calculated average of all quantitative attributes. Data fed into the descriptive statistics with XLSTAT 2018 software to compute all attributes (Addinsoft 2022). The individuals’ coordinate was used to cluster accessions by Hierarchical Ascending Classification (Arabie et al. 1996; Everitt et al. 2001; Jobson 1992). A heat map was plotted according to (Hahne et al. 2008; Rokach and Maimon 2005) using the online program package (https://biit.cs.ut.ee/clustvis/; Metsalu and Vilo 2015) and XLSTAT Version 2014.5.03 (http://www.xlstat.com) based on Euclidean distance as the similarity measure and hierarchical clustering with complete linkage.
Results and discussions
Taxonomic position of Corchorus L., Sp. Pl. 1:529 (1753).
Five species in the stripe of Egypt as shown in are utilized for biodiversity of Corchorus using the HTPP. The status of those is reported in Table 1. Kingdom: Plantae; Division: Magnoliophyta; Subdivision: Angiosperm; Class: Magnoliopsida; Order: Malvales; Family: Malvaceae; Subfamily: Grewioideae; Tribe: Grewieae; Genus: Corchorus. The phylogenetic analyses clarify the taxonomic position (Whitlock et al. 2003). The genus Corchorus falls into the subfamily Grewioideae. Reviews strongly include the Oceanopapaver in a clade of Malvaceae, the Grewioideae (Malvaceae). Morphological attributes consist of stellate indumentum, mucilage canals, valvate sepals with stellate hairs on the adaxial surface, and pollen morphology.
A key of Corchorus species growing in Egypt
-
1. Capsule splitting into 5 valves
-
2. Capsules cylindric, tapering to a beak wich 5-10mm long; Seeds angular, irregularly ribbed, black; Leaves narrow-elliptic, oblong-ovate, or ovate-lanceolate, margins serrate with 2 basal serrations prolonged into 2 setae. C. olitorius
-
2. Capsules globose, flattened on top; Seeds oval pointed, concave on one surface, copper brown; Leaves lanceolate, glabrous or glabrescent, serrate, 2 lower teeth prolonged into fine pointed auricles, apex acuminate. C. capsularis
-
1. Capsule splitting into 3–4 valves.
-
3. Leave margin crenate …C. depressus.
-
3. Leave margin serrate.
-
4. Capsule when young, densely covered with shining palmate hairs, glabrous when mature; seed oblong-ovoid, blackish and pruinose, smooth.…. C. trilocularis.
-
4. Capsule glabrous to sparsely setulose, ending in 3 small spreading horns splitting into 3 valves ending in 3 small spreading horns; seeds angular, oblong, dull brown, finely reticulate……. C. tridens.
The synonymy information gives spacious use in Corchorus taxonomy and applicable genetic improvement. Lack of information on synonymy may lead to the hybridizing of the same species and posterior failure of the crop improvement programs (Sinha et al. 2011). Hinsley (2008) was synonymized the genus Antichorus Linn., Caricteria Scop., Coreta P. Bri., Ganja Reichb., Maerlensia Vell., Nettoa Baill., Oceanopapaver Guillaumin, Palladia Lam., Rhizanota Lour. ex Gomes, Riddelia Rafin., Scorpia Ewart & A.H.K. Petrie with the genus Corchorus L. Correspondingly, some Corchorus species synonymized with already validated species. Hinsley (2008) synonymized 87 validated Corchorus species in detail (further details: http://www.malvaceae.info/Synonymy/Synonymy.php?genus=Corchorus ).
Morphological characterization of Corchorus L.
Corchorus olitorius L., Sp. P1. 1:529 (1753).
Plant is an annual herbaceous reaching to 2.5 m, often suffrutescent. Stems are reddish, sparsely pilose to glabrescent. Leaves are elliptic to narrow-elliptic, oblong-ovate, or ovate-lanceolate, chartaceous, 4.5–15.0 cm long, 1–5 cm wide, margins serrate with 2 basal serrations prolonged into 2 setae 0.6–2.3 cm long, rounded at the base, acute to acuminate, prominently nerved above and beneath, with sparse setaceous hairs; petioles (0.6-)1.4–5.0 cm long, moderately covered with setaceous hairs on upper surface, stipules 0.5–1.6 cm long, glabrous. Inflorescences are in 1–2(3) flowered fascicles; peduncles are up to 8 mm long, glabrous; pedicels are 1-3 mm long, glabrous; bracts are up to 3 mm long, glabrous. Sepals are five, with 5.0–7.5 mm long and 1.5 mm wide, circulating at the basal margin. Petals are five, pale yellow, oblanceolate, with a short ciliate claw at the base, 5-7 mm long, 1-2 mm wide. Stamens are numerous. Ovary is cylindric, minutely setulose, and 3.5 mm long. Capsules are cylindric, 10-ribbed, straight or slightly curved on erect pedicels. 2–10 cm long, 0.4–0.8 cm wide, splitting into five valves tapering to a beak 5–10 mm long; young capsules are moderately pilose with adpressed simple hairs, mature capsules are smooth (often somewhat chequered), glabrous and often striped. Seeds are angular, (1.0-)1.5–2.0 mm long, irregularly ribbed, and black.
Corchorus trilocularis L., Syst. Nat. ed. 12(2):369 (1767).
Plant is an annual herb to 1 m, rarely to 1.5 m. Stems are erect or decumbent, often purplish, and moderately covered with spreading setulose pubescence when young; on older stems pubescence is confined to one side of the stem. Leaves are narrow-elliptic to elliptic, ovate or lanceolate, with pubescence on midrib and veins, 4–9(− 13) cm long, 0.6–3.0 cm wide, margins coarsely serrate (occasionally crenate), with a pair of basal setae, 0.6–10.0 mm (though these are often indistinct or absent), rounded or broadly cuneate at the base, acute to subacute; petioles are 0.3–2.0 cm long, covered with setaceous hairs; stipules are 6–12 mm long, pubescent. Inflorescences are 1–3 flowered cymes, and peduncles are 1-2 mm long, often forked; pedicels are 1–4 mm long; bracts are up to 3.3 mm long, ciliate. Sepals are 4–5, 5.5–6.3 mm long, 1 cm wide, dorsal surface pubescent, especially at the base, torulose. Petals are 4–5, obovate tapering to a short ciliate claw, 5–7 mm long, 2.0–2.5 mm wide. Stamens are numerous. Ovary is cylindric, 3 mm long, and shortly pubescent. Capsules cylindric, torulose, 2.5–7.3(− 9.2) cm long, 1.0–2.5(− 4.0) mm wide, held erect straight or slightly curved, splitting into 3, rarely 4, valves, glabrous when mature, covered with scattered forked or palmate hairs which are dense when young. Seeds are smooth, oblong-ovoid, blackish and pruinose, and 1 mm in diameter.
Corchorus tridens L., Mant. Pl., Alt. 2:566 (1771).
Plant is an annual herb, suberect or decumbent, up to 1 m tall. Stems are glabrous to sparsely pilose and red. Leaves are narrow-ovate, ovate or narrowly oblong, glabrous above; pubescent on midrib beneath and occasionally on the blade, 2.8–11.2 cm long, 0.6–2.0(3.0) cm wide, and margins are coarsely serrate, usually with a pair of basal setae, 2–9 mm long, rounded at the base, acute to subacute; petioles are 0.3–1.4 cm long, pilose especially on upper surface; stipules are up to 5 mm, glabrous. Inflorescences are in 1–4 flowers, glabrous fascicles; bracts are up to 4 mm long, glabrous. Sepals are 4–5 with long of 3.5–4.3 mm, and wide of 0.5–0.6 mm, glabrous or with odd hairs. Petals are in 4–5, obovate, tapering to a short ciliate claw, 3.5–4.8 mm long, 0.5–1.5 mm wide. Ovary is cylindric, 2–3 mm long, and papillose. Capsules are cylinders, 2.0–4.2 cm long, 1.0–1.5(2.0) mm wide, held erect, straight or slightly curved, splitting into 3 valves, glabrous to sparsely setulose, ending in 3 small spreading horns (c. 1 mm long). Seeds are angular, oblong, dull dark brown, up to 0.5 mm in diameter, and finely reticulate.
Corchorus depressus (L.) Stocks. C. Christensen in Dansk. Bot. Arkiv., 4:34 (1922); vide Chatterje in Kew Bull., 1948:372 (1949)
Plants are prostrate annual or perennial herbaceous, up to 30 cm long. Stems are mostly adpressed to grounded, glabrous to sparsely pubescent. Leaves are small, glabrous to sparsely setulose on veins; leaves are ovate to elliptic or oblong with a long of 0.3–2.5 cm and wide of 0.2–1.5 cm, without setae, margins are crenate to serrate, acute to rounded; petioles are 0.2–4 cm; stipules are up to 2 mm long, sparsely pubescent. Inflorescences are in 1–2(4) flowered glabrous fascicles, peduncles and are up 2 mm, curved outwards and downwards in fruit, glabrous; bracts are a long of 1.5–2.0 mm. Sepals are a long of 2-3 mm, glabrous. Stamens are 10–15. Ovary is densely covered with bulbous-based bristly hairs. Capsules are solitary or in 2’s, 4-valved 0.8–2.0 mm long, scabrid, ± straight to strongly bent and twisted, often buried in the ground. Seeds are a long of 1.0–1.5 mm, and very finely reticulate.
Corchorus capsularis L. Sp. Pl., 529 (1753).
Plant is herbaceous annual with a straight slender stem, 3–4 m in height under cultivation, branching near the top. Leaves are alternate, light green, leaf blade is lanceolate, glabrous or glabrescent, 5-5 cm wide, serrate, 2 lower teeth prolonged into fine pointed auricles, apex acuminate; petioles are 1–2 cm long; stipules are up to 10 mm long, linear, deciduous. Inflorescences are solitary or in few-flowered cymes, leaf-opposed, sessile or shortly pedunculate; pedicels are very short. Sepals are usually 5, free, narrow, and 4–5 mm long. Petals are usually 5, yellow, 4-5 mm long; short corona separates petals from insertion of stamens. Stamens are 10 to many, free, filaments, short; anthers are small, bi-lobed. Ovary is superior, 5-locular with numerous ovules. Style is short with flattened stigma. Capsules are globose, 0.8–2.0 cm in diameter, wrinkled, 10-ridged, flattened on top, dehiscing loculicidal into 5 valves, without transverse partitions between seeds. Seeds are small, 2-3 mm long, oval pointed, concave on one surface, and copper brown.
Polymorphism and variability of C. olitorius morphological criteria:
In this study, the 49 morphological descriptors points of view were explored for distinctness of olitorius accessions via HTPP (Table S2). It is notably that data showed a broad sense of polymorphism that the attributes of branching habit, color of branches, plant height, plant width, a diameter of the basal stem, stem color, stem diameter, blade color, stem angle, leaf shape, blade length, blade width, nature of leaf-margin, surface nature, pubescence of leaves, basal setae length, petiole color, petiole length, fruit diameter, surface between ridges, shape of apex, length of mature fruit, pedicel length, peak length, seed size, seed color, time of flowering, and time of seedy harvest maturity were polymorphic. The attributes of premature flowering resistance, life span, plant habit, stem texture, distribution of pubescence no. of nodes, leaf apex, leaf base, softness of leaf-margin, existence of leaf-basal setae, direction of setae, dehiscence of a pod, no. of locules, capsule shape, curvature of a mature capsule, direction of curvature, hairiness of capsule, seed diameter, seed shape, and glossiness of seed surface were monomorphic. For more distinctness of olitorius accessions, the derived data were subjected to the DUS-pie test (Fig. 1). Notably, the attributes of branch habit, plant height, stem color, leaf shape, stem angle, seed color, seed size, and time of flowering were the most distinct attributes.

DUS-Pie testing of 53-olitorius accessions. Illustration in outers indicates the ID accession number (see Table S1) present in the concerned state of polymorphic attributes
Agro-biodiversity and eco-geographic information are vital to conservation and utilization for various specialists (FAO 1994, 2006). These data, including plant descriptors, can help manage and utilize collections. Therefore, the HTPP assist scientists in improving their strategy and capacity to manage genebank and facilitate the exchange of the GRs and information (BI 2007; Gotor et al. 2008; Van Hintum 1993).
Corchorus GRs hierarchical clustering
Hierarchical clustering is classified and explores the accessions with their wild relatives. The illustration reflects a chromatic appraisal of the different accessions. It is based on Euclidian distances using agglomerative hierarchical clustering of Ward’s method, optionally went before by the k-means algorithm depending on the matrix size. The dendrogram is split into three main classes as depicted in Figure S1 (accessions are similarly colored in Table S2). First-class (left structure colored with olive green) clustered 18 accessions into a group. Second (right structure) is split into two sub-classes including 35 accessions (colored with red) that are close to wild species. Four wild species are grouped into a separated sub-class (colored with pink; Table S2).
The distance between the centroids of classes is calculated (Table 1). For more classification, variance decomposition is estimated within class of 68.87% and between classes of 31.13%. The major distance was recorded at 18.41 between class 1 (wild relatives: C. trilocularis, C. depressus, C. tridens, C. capsularis) and class three (18 accessions of C. olitorius). In contrast, the minor distance scored 8.3 between class 2 (35 accessions) and class 3 (18 accessions).
Data revealed via hierarchical clustering explored a chromatic appraisal of the different genetic resources according to Euclidian distances. Our results clustered all species as distinct taxa and defined interspecific relationships, though a slight noticeable variance. The possibilities were suggested to elucidate the biodiversity and habitat of Corchorus GRs. The HTPP as an extensive characterization of PGRs is a great challenge and opportunity in the future that document plant agro-/biodiversity on a broader scale worldwide.
The highest similarity was observed between trilocularis and tridens (81.67%). Both the cultivated species of Corchorus scored 90% similarity at the DNA level.
Configuration of C. olitorius accession perceptual mapping
Perceptual mapping is used to visually relations between objects and subjects to interpret a point represented.
A PCA biplot illustrated the impact of the studied 53 olitorius accession and 30 polymorphic characters (Fig. 2). Accessions 29, 31, 36, 39 & 53 grouped influenced by beak length (23), fruit diameter (18), surface between ridges (19), seed size (24), seed color (25), time of flowering (26), time of seed harvest maturity (27), stem diameter (7), leaf shape (10) and weight of total plant (29). For accession name see Table (S1) & for characters see Table (S2).

Perceptual Mapping (MDPREF Bi-plot) analysis illustrating the genetic distance of the 53-olitorius accession (numbers) and 30 polymorphic characters (Numbers in bracts), Configuration has been standardized prior to fitting. For accession and characters see Table (S1 and S2)
Heat maps of Corchorus spp. and their attributes
A cluster is categorized and explored by a group of attributes that correspond to accessions. The heat maps tool is based on Euclidian distances using ascendant hierarchical clustering, optionally went before by the k-means algorithm depending on the size of matrix. It reflects data in the permuted matrix where data values are swapped by matching color intensities.
A clustered data heat-map analysis of all attributes was applied to illustrate a chromatic appraisal of the different accessions. Analysis clustering of left structure is divided into two main groups. The first group (group A) correlates with the attributes of softness of leaf-margin, premature flowering resistance, capsule shape, seed shape, no. of locules, direction of curvature, fruit diameter, length of mature fruit, blade color, pedicel length, peak length, plant width, diameter of basal stem, existence of leaf-basal setae, glossiness of seed surface, basal setae length, petiole length, plant height, blade length, and width. Another group (group B) correlates with the rest attributes (Figure S2). It is notable that in each group, the most attributes are quantitative criteria.
On the other side, the top cluster analysis illustrates a chromatic appraisal of the different accessions is divided into two main groups (left to right); the first group includes the accessions of C. olitorius only that split into subgroups subsequently. Another group is separated into two main subgroups; the first sub-group includes 55 and 57 in a cluster and accession of 54 and 65 in another cluster.
The relationship between attributes with accessions that show significant variability from less to major may implicate various disciplines, including improvement programs, genetic diversity, speciation diversification, and agronomic and systematic disciplines.
The heat map findings strengthened the evidenced frequency of agro-/biodiversity. Finally, our results also presented that the HTPP significantly affected various variables, including agro-ecological and eco-geographical effects for diverse market destinations. These outcomes are in line with those reported (Métais et al. 2002; Tripodi et al. 2019). The HTPP makes available direct measurements and skips indirect measurements of biodiversity based on collection data, passport data, characterization and evaluation, or geographical origin (Métais et al. 2002).
Biodiversity status and conservation of Corchorus L. GRs in Egypt
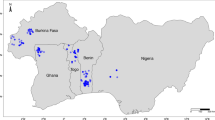
Global Biodiversity Information Facility (http://www.gbif.org) reported more than 200 taxonomical forms belonging to the corchorus. About a hundred are exactly so good species with the domesticated C. olitorius and C. capsularis. Other taxa are wild naturally. Most of those are extinct or endemic. Conservation of plant genetic resources (PGR) do a vital role against potential genetic erosion for quality and yield attributes in a cultivar and abiotic and biotic stresses resistance/tolerance. In Egypt, where 2200 species are widely distributed across phytogeographical and eco-geographical regions, PGRs should be conserved practically feasible or economically sound. Egypt involves three wild species stated rarity. Nonetheless, major impediments have been reported due to extreme variable taxa, eco-geographical distribution, genetic relationships, evolutionary associations, and center of origin being poorly reported, effectively contentious, and yet to be resolved. PGR conservation strategies prioritize primacy based on the distribution occurrence and habitat specificity (Ahemadullah and Nayar 1986). Corchorus tridens, C. trilocularis, and C. depressus with their rarity distribution and habitats (Table 2), prioritize highly conservation due to the association of evolutionary role and importance of genetic improvement. Because of climate change and anthropogenic activities such as urban sprawl and overgrazing, small prominent populations are more susceptible to genetic erosion. Similar lands like India and Ethiopia accentuate the problem's gravity, further emphasized in climate change-related vulnerability studies (Benor 2010, 2012; El-Zeina and Tonmoy 2015; Vollesen and Demissew 1995). Richness of taxa is a key biodiversity indicator based mainly on environmental factors that form a broad sense of microclimate in a certain landscape (Ellu and Obua 2005; Kharkwal et al. 2005). Distribution of Corchorus species across the country as clarifies the phenomenon. Egypt has already been documented as an important repository of 70 families (Boulos 2000, 2009).
In the HTPP, direct measurements of biodiversity are given priority over indirect measurements based on collection data, passport data, characterization, or geographic origin (Métais et al. 2002). Various efforts have been made in the past to document and state the variance among taxa Corchorus via the integrity of various approaches, including micro-and macro-morphology, photochemistry, and molecular markers (Mahdy 2012; Hossain et al. 2002, 2003; Qi et al. 2004; Basu et al. 2004; Roy et al. 2006; Haque et al. 2007; Mir et al. 2008b). Many reviews had determined and stated the biodiversity of Corchorus collections from diverse locations using morphological description (Mahdy 2012), karyotype (Mahdy 2012) Random Amplified Polymorphic DNA (RAPD; Qi et al. 2003b; Haque et al. 2007), Inter-Simple Sequence Repeat (ISSR; Mahdy 2012; Qi et al. 2003a), Amplified Fragment Length Polymorphism (AFLP; Hossain et al. 2002, 2003) and Simple Sequence Repeat (SSR; Basu et al. 2004; Akter et al. 2008; Mir et al. 2008a, b), Sequence Tagged Microsatellite (STMS; Qi et al. 2004; Roy et al. 2006).
Conclusions and future prospective
The relationship between attributes with accessions that show significant variability from less to major may implicate various disciplines, including improvement programs, genetic diversity, speciation diversification, and agronomic and systematic disciplines.
Biodiversity information facilitates the conservation of PGRs that shield against a potent genetic erosion. Corchorus tridens, C. trilocularis, and C. depressus, with their rarity distribution and habitats prioritize highly conservation due to the association of evolutionary role and importance of genetic improvement. It is necessary to conserve PGRs in a practical and economical sound. Though, main difficulty has been reported due to extreme variable taxa, genetic relationships, natural distribution, evolutionary associations, and center of origin that are inadequately recognized, effectively contentious and yet to be resolved. Conservation strategies applied to PGR prioritization are influences by the distribution frequency and habitat specificity. The significant germplasm is more prone to genetic erosion accounting for the loss of genetic diversity. Species richness is a key biodiversity indicator established on agro-/biodiversity factors. The HTPP is based on morphological measurements made available to highlight the direct and indirect indicators of Corchorus biodiversity. The integrity of various approaches and tools, including micro- and macro-morphology, photochemistry, and molecular markers, should be processed. Diversification is a plant depends mainly on: (1) genetic improving novel germplasm, (2) screening of the climate changing, (3) utilization of available genepools, (4) exchange the GRs, and (5) measurement of plant biodiversity. This can afford available opportunities to minimize exposure risks due to biotic/abiotic stresses including climate changes and loss of agro-/biodiversity. This is the first study describing morphological attributes of the entire accessions conserved in the National gene Bank (NGB) collections. The results obtained in this study serve as useful information on genetic diversity of Jew’s Mallow accessions at the National gene Bank (NGB), which could be used for C. olitorius genetic improvement programs. In final, it is notable in our data that the attributes of premature flowering resistance, branch habit, plant height, stem color, leaf shape, stem angle, seed color, seed size, and time of flowering were the most discrete attributes. It could say that those are very valuable for boosting the various platforms and disciplines.
Availability of data and materials
The germplasms used are available for distribution to researchers who are interested in them. Requests for material should be sent to National Gene Bank (NGB), Agricultural Research Center (ARC), 9 Gamaa St., PO Box: 12,619; Giza, Egypt.
References
Addinsoft (2022) XLSTAT statistical and data analysis solution. New York, USA. https://www.xlstat.com/en
Agarwal M, Shrivastava N, Padh H (2008) Advances in molecular marker techniques and their applications in plant sciences. Plant Cell Rep 27:617–631. https://doi.org/10.1007/s00299-008-0507-z
Ahemadullah M, Nayar MP (1986) Endemic plants of Indian region, vol I Botanical Survey of India, Howrah
Akter J, Islam MS, Sajib AA, Ashraf N, Haque S, Khan H (2008) Microsatellite markers for determining genetic identities and genetic diversity among jute cultivars. Aust J Crop Sci 1:97–107
Aluka (2006–2008) African plants. http://www.aluka.org. Accessed Jan 2009.
Arabie P, Hubert LJ, De Soete G (1996) Clustering and classification. Wold Scientific, Singapore
Basu A, Ghosh M, Meyer R, Powell W, Basak SL, Sen SK (2004) Analysis of genetic diversity in cultivated jute determined by means of SSR markers and AFLP profiling. Crop Sci 44:678–685
Benor S, Blattner FR, Demissew S, Hammer K (2010) Collection and ethnobotanical investigation of Corchorus species in Ethiopia: potential leafy vegetables for dry regions. Genet Resour Crop Evol 57:293–306
Benor S, Demissew S, Hammer K, Blattner FR (2012) Genetic diversity and relationships in Corchorus olitorius (Malvaceae) inferred from molecular and morphological data. Genet Resour Crop Evol 59:1125–1146. https://doi.org/10.1007/s10722-011-9748-8
Bioversity International (2007) Guidelines for the development of crop descriptor lists. Bioversity technical bulletin series. Bioversity International, Rome, Italy. http://www.bioversityinternational.org/nc/publications/publication/issue/developing_crop_descriptor_lists.html.
Boulos L (2009) Flora of Egypt. Revised Annonated Edition, Al Hadara Publishing, Egypt, Checklist
Boulos L (2000) Flora of Egypt. Al-Hadara Publishing, Cairo, Egypt
Chopra RN, Nayar SL, Chopra IC (1986) Glossary of Indian medicinal plants. Council of Scientific and Industrial Research, India
Edmonds JM (1990) Herbarium survey of African Corchorus species: Systematic and eco-geographic studies in crop gene pools. International Board of Plant Genetic Resources, Rome, Italy, pp 2–3.
Ellu G, Obua J (2005) Tree condition and natural regeneration in disturbed sites of Bwindi Impenetrable Forest National Park, southwestern Uganda. Trop Ecol 46(1):99–111
El-Sharabasy S, Rizk R (2019) Atlas of date palm in Egypt. FAO, Egypt
El-Wakeel HE (2007) The impact of development policies and human activities on plant genetic resources on Siwa Oasis. The capacity building program for Genetic Resources Policies Initiative (GRPI-Egypt), Bioversity International, 32–35
El-Zeina A, Tonmoy FN (2015) Assessment of vulnerability to climate change using a multi-criteria outranking approach with application to heat stress in Sydney. Ecol Indic 48:207–217
Everitt BS, Landau S, Leese M (2001) Cluster analysis, 4th edn. Arnold, London
FAO - Food and agriculture organization of the United Nations (2006) Guidelines for soil description, 4th edn. Food and Agriculture Organization of the United Nations, Rome
FAO - Food and agriculture organization of the United Nations/International plant genetic resources institute (1994) Genebank Standards. Food and agriculture organization of the United Nations/International plant genetic resources institute, Rome.
Finkel E (2009) With ‘phenomics’, plant scientists hope to shift breeding into overdrive. Science 325:380–381. https://doi.org/10.1126/science.325_380
Ghosh RK, Sreewongchai T, Nakasathien S, Phumichai C (2012) Phenotypic variation and the relationships among jute (Corchorus species) genotypes using morpho-agronomic traits and multivariate analysis. Aust J Crop Sci 7(6):830–842
Goddard ME, Hayes BJ (2007) Genomic selection. J Anim Breed Genet 124:323–330
Gotor E, Alercia A, Ramanatha V, Watts J, Caracciolo F (2008) The scientific information activity of bioversity International: the descriptor list. Genet Resour Crop Evol 55:757–772
Hahne F, Huber W, Gentleman R, Falcon S (2008) Bioconductor case studies. Springer, USA
Haque S, Begum S, Sarker RH, Khan H (2007) Determining genetic diversity of some jute varieties and accessions using RAPD markers. Plant Tiss Cult Biotechnol 17:183–191
Hazra SK, Saha A (2004) Unexploited potential of Indian Corchorus and Hibiscus for therapeutic uses and developing phytomedicines. In: proceedings of national seminar on diversified uses of jute and allied fiber crops, Barrackpore, Kolkata, India, 8–9 Jan 2004, pp 49–54
Hinsley SR (2008) Partial synonymy of Corchorus. http://www.malvaceae.info/Synonymy. Accessed Mar 2009.
Hossain MB, Haque S, Khan H (2002) DNA fingerprinting of jute germplasm by RAPD. J Biochem Mol Biol 35:414–419
Hossain MB, Awal A, Rahman MA, Haque S, Khan H (2003) Distinction between cold-sensitive and -tolerant jute by DNA polymorphism. J Biochem Mol Biol 36(5):427–432
Ingvarsson PK, Street NR (2011) Association genetics of complex traits in plants. New Phytol 189:909–922
Jobson JD (1992) Applied multivariate data analysis categorical and multivariate methods. Springer-Verlag, New York
Kharkwal G, Mehrotra P, Rawat YS, Pangtey YPS (2005) Phytodiversity and growth form in relation to altitudinal gradient in the Central Himalayan (Kumaun) region of India. Curr Sci 89(5):873–878
Kumar D, Mahapatra AK (2004) Draft National test guidelines for testing distinctness, uniformity and stability (DUS) for jute. Central Research Institute for Jute and Allied Fibers, Barrackopre, West Bengal, 1–14.
La Farge T, Friedman ST, Cock CG (1997) Improvement of fiber crops using genetics and biotechnology. In: Powell RM et al (eds) paper and composites from agro-based resources. CRC Lewis Publishers, Boca Raton, Fl, pp 39–59
Loumerem M, Alercia A (2016) Descriptors for jute (Corchorus olitorius L.). Genet Resour Crop Evol 63:1103–1111. https://doi.org/10.1007/s10722-016-0415-y
Mahdy EMB (2012) Genetic studies on some vegetable crops (Corchorus olitorius L. and Lactuca sativa L.). M.Sc. Thesis Fac of Agric Zagazig Univ Egypt
Métais I, Hamon B, Jalouzot R, Peltier D (2002) Structure and level of genetic diversity in various bean types evidenced with microsatellite markers isolated from a genomic enriched library. Theor Appl Genet 104:1346–1352
Metsalu T, Vilo J (2015) Clustvis: a web tool for visualizing clustering of multivariate data using principal component analysis and heatmap. Nucleic Acids Res 43(W1):W566–W570. https://doi.org/10.1093/nar/gkv468
Mir JI, Karmakar PG, Chattopadhyay S, Chaudhury SK, Ghosh SK, Roy A (2008a) SSR and RAPD profile based grouping of selected jute germplasm with respect to fibre fineness trait. J Plant Biochem Biotechnol 17:29–35
Mir RR, Rustgi S, Sharma S, Singh R, Goyal A, Kumar J, Gaur A, Tyagi AK, Sinha MK, Balyan HS, Gupta PK (2008b) A preliminary genetic analysis of fibre traits and the use of new genomic SSRs for genetic diversity in jute. Euphytica 161:413–427
Natesh S (2001) The changing global scenario of herbal drugs: role of botanist. Phytomorphology 51(3–4):75–96
Qi J, Zhou D, Wu W, Lin L, Fang P, Wu J (2003a) The application of RAPD technology in genetic diversity detection of Jute. Yi Chuan Xue Bao 30:926–932
Qi J, Zhou D, Wu W, Lin L, Wu J, Fang P (2003b) Application of ISSR technology in genetic diversity detection of jute. Ying Yong Sheng Tai Xue Bao 14:1473–1477
Qi JM, Zhou DX, Wu WR, Lin LH, Fang PP, Wu JM (2003c) The application of RAPD technology in genetic diversity detection of Jute. Yi ChuanXueBao Oct 10:926–932
Qi JM, Zhou D, Wu WR, Wu W, Lin L, Fang P, Wu JM, Wu J (2004) A comparison between RAPD and ISSR technology in detection of genetic diversity of jute. Sci Agric Sin 37:2006–2011
Rizk RM, El-Sharabasy SF (2008) A descriptor for date palm (Phoenix dactylifera l.) characterization and evaluation in gene banks. Plant Genet Res Newsl 150:42–44
Rokach L, Maimon O (2005) Clustering methods data mining and knowledge discovery handbook. Springer, NY
Roy A, Bandyopadhyay A, Mahapatra AK, Ghosh SK, Singh NK, Bansal KC, Koundal KR, Mohapatra T (2006) Evaluation of genetic diversity in jute (Corchrus spesies) using STMS, ISSR and RAPD markers. Plant Breed 125:292–297
Saunders M (2006) Recovery plan for the endangered native jute species, Corchorus cunninghamii F. Muell in Queensland (2001–2006). Natural Heritage Trust, Australia, 1–29.
Sen S (2002) Legacy of international health care: drug resistance, drug development and therapeutic status. Sci Cult 68:45–55
Sinha MK, Kar CS, Ramasubramanian T, Kundu A, Mahapatra BS (2011) Corchorus. In : Kole C (ed.) Wild crop relatives: genomic and breeding resources, industrial crops, Springer-Verlag Berlin Heidelberg. https://doi.org/10.1007/978-3-642-21102-7_2.
Stearn WT (1995) Botanical latin, 4th edn. David & Charles Publishers, Newton Abbot
Tripodi P, Ficcadenti N, Rotino GL et al (2019) Genotypic and environmental effects on the agronomic, health-related compounds and antioxidant properties of chilli peppers for diverse market destinations. J Sci Food Agric 99:4550–4560
Van Hintum TJL (1993) A computer compatible system for scoring heterogeneous populations. Genet Resour Crop Evol 40:133–136
Vollesen K, Demissew S (1995) Corchorus. In: Edwards S, Tadesse M, Hedberg I (eds) Flora of Ethiopia and Eritrea, vol 2. part 2. Canellaceae to Euphorbiaceae. The National Herbarium, Addis Ababa University and Department of Systematic Botany, Uppsala University, Addis Ababa and Uppsala, pp 154–158
Whitehouse C, Cheek M, Andrews S, Verdcourt B (2001) Flora of tropical East Africa, Tiliaceae & Muntingiaceae, 1–120
Whitlock BA, Karol KG, Alverson WS (2003) Chloroplast DNA sequences confirm the placement of Oceanopapaver within Corchorus (Grewioideae: (Malvaceae s.l., formerly Tiliaceae). Genome Biol Evol 164:35–41
Acknowledgements
We acknowledge Prof. Dr. Reda M. Rizk, consultant of PGR, Arab Organization for Agricultural development (AOAD), for his great efforts to identify the accessions.
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Contributions
Conceptualization: E.M.B.M; Methodology: E.M.B.M and H.A; Software: E.M.B.M; Validation, E.M.B.M; Formal analysis, E.M.B.M; Investigation: E.M.B.M. and H.A; Resources: E.M.B.M. and H.A; Data curation: E.M.B.M and H.A; Writing (original draft preparation): E.M.B.M; Writing (review and editing): E.M.B.M and H.A.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Ethics approval
No approval is required for the study.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Mahdy, E.M.B., Ahmad, H. A study of Corchorus L. diversity in Egypt using high-throughput phenotyping platform (HTPP): an Egyptian gene bank example. Genet Resour Crop Evol 70, 2009–2019 (2023). https://doi.org/10.1007/s10722-023-01551-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10722-023-01551-6