
Abstract
Despite much research on mimicry, little is known about the ecology of dynamic mimetic signals involving mimicry of multiple species. Some of the most conspicuous examples of phenotypically plastic mimicry are produced by oscine passerines, where vocal production learning enables some species to mimic multiple models and flexibly adjust what they mimic and when. While singing from a perch, male superb lyrebirds (Menura novaehollandiae) accurately imitate multiple songs and calls of over 20 species of bird. However, at key moments within their multimodal displays performed on display arenas on the forest floor, males mimic a small number of mobbing-alarm calls creating the acoustic illusion of a mixed-species mobbing flock (‘D-song’). Using observations from camera footage and a field-based playback experiment, we tested six hypotheses for alarm call model selection within D-song. Mimicked species were remarkably invariant, with 79% of D-song made up of imitations of just three different bird species. Males did not mimic the most common species in their general environment, but neither did they mimic rare species. Instead, males imitated the mobbing-alarm calls of heterospecific birds that foraged on or near the forest floor. Indeed, males primarily mimicked the alarm calls of heterospecific species that foraged alongside lyrebirds and were likely to appear together in experimentally-induced, terrestrial mobbing flocks. These findings support the hypothesis that males mimic a cue of a terrestrial predatory threat to lyrebirds, most likely to exploit the antipredator behaviour of female lyrebirds. Our study illustrates the importance of investigating the drivers of model selection in dynamic multi-model mimicry.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Mimicry – where an individual gains adaptive benefits by simulating the phenotype of a model – is widespread across taxa, functioning in many ecological contexts via a diverse range of mimetic signal structures and sensory modalities (Dalziell and Welbergen 2016b). The best-studied examples of mimicry involve static signals where an individual mimic is limited to mimicking just one, fixed phenotype (e.g., host eggs by common cuckoos, Cuculus canorus Davies 2015). However, some taxa mimic multiple models and flexibly adjust which models they mimic and when (Cheney et al. 2008). For example, the mimic octopus (Thaumoctopus mimicus) mimics the colour and shape of multiple sympatric species and can change between visual forms (models) within a matter of seconds (Norman et al. 2001). The jumping spider Portia fimbriata uses different mimetic vibratory signals to capture different types of prey (Jackson and Wilcox 1990; Tarsitano et al. 2000; Herberstein 2011; Nelson and Jackson 2011). Perhaps the most conspicuous and complex dynamic mimetic signals are those incorporated into the vocalisations of birds. Approximately 7–15% of oscine passerines are estimated to imitate sounds produced by other species, but estimates suffer from a paucity of research (Goller and Shizuka 2018). For the majority of avian vocal mimics, it is unclear why some models are mimicked and others are not (Dalziell et al. 2015).
Several species of bird regularly mimic the acoustic alarm calls given by prey species in response to a predator (reviewed in Dalziell et al. 2015). Mimicking alarm calls could facilitate interspecific communication about predators, thereby reducing the risk of predation for the mimic (Chu 2001; Wheatcroft and Price 2013; Goodale et al. 2014; Igic and Magrath 2014). Alternatively, individuals may mimic alarm calls deceptively to exploit the anti-predator behaviour of others (Igic et al. 2015). For example, fork-tailed drongos (Dicrurus adsimilis) mimic the alarm calls of heterospecifics in the absence of predators to steal food (Flower 2011; Flower et al. 2014). The drongo increases its success in stealing food from other species by tailoring its mimicry to the targeted victim (Flower et al. 2014).
Deceptive vocal mimicry of multiple alarm calls has recently been reported in the sexual display of the male superb lyrebird, Menura novaehollandiae (Dalziell et al. 2021). The superb lyrebird is a large oscine passerine that produces highly accurate and versatile vocal mimicry in the wild (Higgins et al. 2001; Zann and Dunstan 2008; Dalziell and Magrath 2012; Dalziell and Welbergen 2016a; Dalziell et al. 2021). Males regularly imitate multiple songs and calls of 20–25 different species of bird while singing from a perch (‘recital display: Dalziell et al. 2022). However, at two key points during sexual interactions with females – when females attempt to leave males without copulating, and during copulation itself – males always mimic a small subset of species in a discrete song-like sequence (‘D-song’: Dalziell et al. 2021). Males pattern mimetic elements in D-song to recreate the acoustic signature of an entire flock of alarm-calling birds. The complexity of this mimicry and its exclusive production in these particular sexual contexts suggests that males use D-song as a sensory trap (sensu Christy 1995; Arnqvist 2006 West-Eberhard 1979), seemingly manipulating an antipredator response in females to prevent them from terminating sexual interactions prematurely (Dalziell et al. 2021). Similar to sensory traps in other species (Bro-Jørgensen and Pangle 2010; Nakano et al. 2010) a male lyrebird may benefit by inducing a female to freeze or make her reluctant to leave the male and his display arena after she hears the male’s false cue of a nearby predator (Dalziell et al. 2021).
Lyrebird D-song mostly comprises mimicry of a specific class of alarm calls termed ‘mobbing’ alarm calls. Mobbing calls are common in passerines worldwide and are typically produced in response to a stationary or slow-moving predator, such as a perched hawk or terrestrial ambush predator (Marler 2004; Magrath et al. 2015; Carlson et al. 2018; Carlson and Griesser 2022). Mobbing calls have the distinctive, general property of attracting conspecific and heterospecific individuals and inciting mobbing behaviours (Caro 2005; Magrath et al. 2015). Male superb lyrebirds, however, imitate mobbing calls in the absence of a predator, and so mimic deceptively (Dalziell et al. 2021). In the lyrebird’s habitat, many different bird species produce mobbing or other alarm calls and natural mobbing choruses vary substantially in species composition and therefore acoustic structure. The results of a previous study suggested that lyrebird D-song is remarkably invariant, being dominated by the mobbing calls of just 3–4 model species (Dalziell et al. 2021). Given their extraordinary ability to mimic a vast array of avian, mammalian and anthropogenic sounds (Higgins et al. 2001), it is unclear why, during their song and dance display, male lyrebirds selectively mimic the mobbing alarm calls of this limited set of species.
In this study we examine six plausible explanations for the specific suite of model species that male lyrebirds mimic during song and dance displays. Male lyrebirds could simply mimic the alarm calls of heterospecifics that are common in the male’s general environment (i), or commonly encountered by male lyrebirds specifically around their terrestrial foraging grounds and display territories (ii). If so, then there should be a positive relationship between the species that feature in lyrebird mimicry and their relative abundance in either the lyrebird’s general environment or the lyrebird’s local microhabitat of the forest floor. Similarly, lyrebirds may preferentially mimic species with which they commonly associate while foraging (iii). Several species of small passerines are thought to benefit from ‘piloting’ lyrebirds by feeding on terrestrial arthropods flushed by lyrebirds digging in the ground or dismantling rotting wood (Smith 1988; Higgins et al. 2001). Alternatively, male lyrebirds may mimic the alarm calls of species that are rare in their environment, perhaps as a signal of the male’s mimetic prowess to the female (iv). If so, there should be a negative relationship between the abundance of heterospecific species in the environment and the probability with which they appear in lyrebird mimicry. It is also possible that male lyrebirds mimic particular mobbing alarm calls that are exceptionally reliable cues of predator presence to lyrebirds and other bird species (v). If so, then we would expect playback of mobbing alarm calls that lyrebirds mimic to attract a greater diversity and/or number of individuals relative to playback of species that lyrebirds do not mimic. Finally, lyrebirds may mimic typical participants of a terrestrial mobbing chorus to create a mobbing chorus that is particularly informative to ground-dwelling species such as lyrebirds (vi). If so, then the species mimicked by lyrebirds should be similar to the suite of species that mob terrestrial threats. Here, we systematically describe lyrebird mimicry in D-song and assess how well the six hypotheses outlined above explain the composition of mimetic sounds in the male lyrebird’s song and dance display.
Materials and methods
Study site
We conducted our study from late May to early August in 2014 and 2015 on a population of lyrebirds in the Blue Mountains National Park in NSW on Dharug and Gundungurra country, Australia. Specifically, we studied lyrebirds in a continuous tract of habitat within the Jamison and Kedumba Valleys near Katoomba (Fig. S1). The study area is characterised by tall forests dominated by Eucalyptus species with dense understories, interspersed with patches of warm temperate rainforest.
Study species and song
Lyrebirds are endemic to the wet forests of south-east Australia. During the winter mating season (May – July) males defend individual territories in which they display in an exploded, lek-like mating system. Both sexes are accurate and versatile vocal mimics (Zann and Dunstan 2008; Dalziell and Magrath 2012; Dalziell and Welbergen 2016a; Dalziell et al. 2021, 2022) In the wild, males mimic a diverse set of sounds, mostly imitations of other species of bird (Higgins et al. 2001). Of the distinct multicomponent displays all males perform (Dalziell et al. 2013, 2022; Dalziell and Welbergen 2022), two involve vocal mimicry. The common ‘recital’ display is usually performed from a perch and predominantly comprise imitations of multiple distinct vocalisations of 20–25 different species of bird (Dalziell et al. 2022 see Fig. 1; for a video example see https://macaulaylibrary.org/asset/475993). In contrast, the ‘dance’ display consist of four distinct song-types (song-types A – D), each accompanied by specific dance-like movements (Dalziell et al. 2013; Supplementary Video 1). Of the four song-types performed in dance displays, only the final vocalisation –‘D-song’ – is mimetic (Dalziell et al. 2013, 2022).

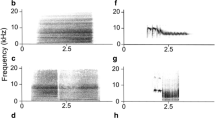
The alarm calls of three species of bird (panel A) regularly mimicked by male superb lyrebirds during D-song (panel B) and one species not mimicked by males during D-song (panel C). These four model sounds were used in the playback experiment (e.g., Fig. 3)
Assessing mimetic composition of lyrebird D-song
To identify mimicry in D-song, we compared lyrebird vocalisations and recordings of suspected model species by ear, and visually by inspecting spectrograms of model and suspected mimetic sounds using Raven Pro 1.5 (Cornell Lab of Ornithology). At least two observers classified each sound and the classification process continued until all observers agreed. We obtained recordings of mimetic D-song from audio-visual footage of the male lyrebirds performing song and dance displays on their display mounds. Audio-visual displays were recorded by motion-triggered trail cameras (Bushnell NatureView Cam HD, model #119,439) installed at lyrebird display mounds. These cameras were set to record sound and video for 60 s after triggering (the maximum possible duration for this camera model). For analysis, we selected recordings with low levels of background sound that contained a complete, uninterrupted sequence of D-song. D-song was considered completed after a male switched to another song-type or the male ceased singing naturally (without evidence of interruption: Dalziell and Magrath 2012; Dalziell et al. 2013). We arbitrarily selected one recording per male (N = 15) for analysis and excluded copulation events from this dataset.
Preliminary analysis indicated that some model species were mimicked much more often than others, both within and among lyrebird males. To examine hypotheses for this very particular distribution of mimicked species within lyrebird D-song, for each rendition of D-song we calculated the ‘relative abundance’ of each mimicked species within lyrebird mimicry as the total duration of each species divided by the total duration of D-song. The distribution of model species in lyrebird D-song was then compared to other ecologically defined species assemblages in the lyrebird’s environment (details below).
Characterising assemblages of potential model species
It is not possible to measure with high accuracy the acoustic environment to which lyrebirds are exposed, given that the variance in soundscapes among forest strata, across seasons and among years is very difficult to control for using conventional survey methods, including those we use here. All these sources of variation are relevant for lyrebirds as they are long-lived (up to at least 30 years: Higgins et al. 2001) and inhabit some of the world’s tallest forests, regularly singing from perches up to 20 m high (Dalziell et al. 2022). Accordingly, we made the pragmatic but theoretically and empirically informed assumption (Alquezar and Machado 2015; Darras et al. 2018) that the sound environment to which lyrebirds are exposed can be estimated by the abundances of bird species in the lyrebird’s environment (e.g., Morrison et al. 2021). We tailor the confidence of our conclusions to reflect the indirect nature of these tests.
We compared the mimetic composition of lyrebird D-song to the relative abundance of bird species in the lyrebird’s general environment (hypotheses i and iv). We defined species in the lyrebird’s general environment as species that can be seen or heard anywhere within the lyrebird’s habitat. We assessed the abundance of such species using 27 opportunistic surveys conducted between 29th May and 22nd July at our field site during the lyrebird’s winter breeding season, coinciding with the second year of recordings (2015). Observers (N = 1–3) counted the number of individuals of all bird species detected either visually or by sound, throughout the entire duration of each visit to the field site. These opportunistic surveys undertaken while conducting fieldwork covered 2.5 ± 1.8 km (mean ± SD), took 4.4 ± 1.8 h (mean ± SD), and started between 06:30 − 13:00. All surveys were conducted in the Jamison/Kedumba Valleys in the general vicinity of male lyrebird display territories and adjacent female nesting territories. We calculated a simple measure of ‘relative abundance’ for each species by calculating the number of individuals of each species detected divided by the total number of individual birds detected per survey. We then calculate the average abundance across surveys and the associated standard error. This method provided a basic measure of the avian assemblage, that while limited, was nonetheless sufficient to test whether lyrebird mimicry in D-song reflected the species assemblage in their general environment (hypothesis i) or whether lyrebirds mimicked rare species (hypothesis iv).
In the tall, dense forests of the lyrebird’s habitat, lyrebirds are more likely to encounter bird species that forage on or near the forest floor than foliage gleaners or nectar feeders that forage high in the canopy. Accordingly, we tested whether lyrebirds were more likely to mimic birds that foraged on or near the forest floor (ii, iv) by comparing the distribution of species within the lyrebird’s terrestrial microhabitat with the distribution of heterospecific model species mimicked by lyrebirds in D-song. We used two methods to characterise the avian assemblage within the lyrebird’s microhabitat. First, we determined the suite of species that occur on lyrebird display mounds (hereafter ‘mound surveys’) using footage captured on the trail cameras (215 × 60 s videos of heterospecific birds). Each mound (Nmounds = 24) originated from a different male in our study population and included mounds from 13 of the 15 males from which we sampled D-song. For each lyrebird mound we calculated the abundance of each species as the number of individuals of each species observed divided by the total number of heterospecific individuals observed. Second, using published literature (Garnett et al. 2013; Pavlacky et al. 2015; Menkhorst et al. 2017), we classified all the species of bird recorded in our general environment surveys and mound surveys into four different foraging microhabitats with the forest: ‘upper-story/canopy’; ‘understory’; ‘terrestrial’; and ‘generalist’ that comprises species that use all these forest strata.
Several small passerines are thought to have feeding associations with superb lyrebirds (reviewed in Higgins et al. 2001), and lyrebird males may be more likely to imitate these species than others (iii). Using our camera trap footage, we identified species that were filmed alongside a lyrebird on the mound (i.e., < 1.5 m from the lyrebird’s feet; Supplementary Video 2) and compared this list of species with the species mimicked by lyrebirds in D-song.
Playback experiment
We conducted a playback experiment to test two further hypotheses for the models within D-song: if male lyrebirds selectively mimic species that are particularly effective at generating mobbing flocks (v); and if male lyrebirds selectively mimic birds that contribute to terrestrial mobbing flocks (vi). To do this, we broadcast the mobbing alarm calls of four different oscine passerines sympatric with lyrebirds; two of which male lyrebirds nearly always imitated during D-song (eastern yellow robin Eopsaltria australis and white-browed scrubwren Sericornis frontalis), one that was sometimes imitated during D-song (yellow-throated scrubwren Sericornis citreogularis), and one that we have never recorded male lyrebirds imitating during D-song (grey fantail Rhipidura albiscapa: Table 1), but can be mimicked by female superb lyrebirds in the same population, during nest defence (Dalziell and Welbergen 2016a: Fig. 1; Table 1; Audio S2).
We carried out our experiment from June to August 2015. We constructed playback tracks from recordings of the mobbing alarm calls of each of the four species that we elicited through playback of conspecific calls and the presentation of a life-sized rubber red-bellied black snake Pseudechis porphyriacus (manufactured by Australian Geographic); a predator common in the Blue Mountains. Mobbing alarm calls were recorded throughout the Blue Mountains. As shown previously (Dalziell et al. 2021), male lyrebirds mimic the mobbing calls of heterospecifics, and these mimicked mobbing calls can be prompted using a life-sized rubber red-bellied black snake. Mobbing calls were recorded at 48 kHz 24-bit using a Marantz PMD 661 digital recorder with a Sennheiser ME67 shotgun microphone or a ME62 microphone set in a Telinga V2 parabola. Recordings were filtered under 1000 Hz and normalized. We recorded five different individuals of each species.
Each trial comprised 5 min of silent, pre-playback observation; 4.5 min of the stimulus and 5 min of silent post-playback observation. Observers noted all species within 15 m of the playback speaker during each of these three phases of the trial. Each stimulus comprised 5 sets of 30 s of alarm calls interspersed with 30 s of silence. The 30-second stimulus was constructed from a single call element repeated at a natural rate and the recording was broadcast at a volume within the natural range of the playback species (51 dB at 6 m). To ensure our experiment reflected the avian species that lyrebirds can encounter, we conducted each trial within 200 m of a lyrebird mound (and at least 100 m from any other trial location to limit pseudoreplication).
Playback experiments were conducted between 07:30 and 13:00 and we performed 10 replicates of each of the 4 species (40 trials in total), each in a unique location. To avoid order effects, we selected the track for a given playback replicate using a Latin square. Two observers stood 15 m away from the speaker and noted each bird observed to come within 15 m of the speaker. While the habitat was dense, two observers together had the visibility required to monitor the 15 m radius circle around the speaker. To minimise observer bias, each of the three observers did an approximately equal number of trials per species (3 or 4). Observers were ‘blind’ to the playback stimulus type before the onset of the playback sound (i.e., during site selection and setup, and pre-playback observation periods), whereon the stimulus type could be identified by ear.
Statistical analysis
All statistical analysis was carried out in JMP Pro 12 (v. 12.0.1, SAS Institute Inc) except for the tests for association, which were performed in SPSS (v. 24.0, IBM Cop.). All tests were two-tailed and significance was set at α < 0.05. Means are expressed ± SE. We compared the distribution of averaged ‘relative abundance’ per mimicked species in lyrebird mimicry to the relative abundance of available (potential) model bird species in the general environment (hypotheses i and iv), the microhabitat of the lyrebird’s mound (mound surveys) (ii), and the species that approach simulated mobbing calls during the playback experiment (vi). Mimic (lyrebird) and suspected model assemblage were compared by calculating the Kendall’s tau-b correlation coefficient of the association. This procedure measures the association between rank orders and corrects for ties. Descriptive statistics were used to assess whether males mimic feeding associates (iii).
We used one-way ANOVA models to test whether lyrebirds were more likely to mimic heterospecific bird species that foraged in similar forest strata to lyrebirds (ii) and to analyse the playback experiment (v). All ANOVA models were examined to ensure that the assumptions of ANOVA models were met. If Levene’s test indicated that the variances across factor levels were not equal, then we used Welch’s ANOVA, which is more robust to heterogeneity of variance than Fischer’s ANOVA. Exploratory analysis of the results from the playback experiment revealed one outlier almost 4 standard-deviations above the mean, but model inferences were qualitatively the same whether or not we included it. This point corresponded to one of 10 trials broadcasting yellow robin mobbing alarm calls. The unusually high number of birds that were attracted to the playback tape speaker (17) was probably due to the unusually high number that were within 15 m of the speaker before the mobbing call was broadcast (5). We considered these insufficient grounds to remove this point.
Results
Species mimicked by lyrebirds in D-song
Male superb lyrebirds mimicked multiple species’ alarm calls during the production of D-song (Fig. 1; Supplementary Videos 1–2), but the suite of mimicked species was remarkably similar across individual males (Table 1). We identified 10 different species mimicked by the 15 male superb lyrebirds that we examined in detail (Table 1). However, imitations of just three calls made up the majority of all recitations of D-song: the alarm calls of eastern yellow robins, brown thornbills, and white-browed scrubwrens (79.3% ± 4.6 of all mimicked calls; Fig. 3).

Relative abundance of bird species in (i) the general environment as assessed by point counts (white bars, N = 27 surveys); (ii) the lyrebird’s terrestrial microhabitat as assessed in ‘mound surveys’ using camera traps positioned on lyrebird display mounds (grey bars, 215 heterospecific individuals filmed on the mounds of 24 individual male lyrebirds, with 34 hours of footage); and (iii) lyrebird D-song (black bars, N = 15 songs from 15 males). Model species relative abundance was calculated as the total number of individuals per species as a proportion of the total number of individuals recorded (i) per survey and (ii) per male lyrebird
(i, iv) does lyrebird mimicry reflect the species assemblage in their general environment?
The distribution of mimicry in lyrebird D-song did not simply mirror the natural distribution of heterospecific bird species in the lyrebirds’ general environment (Fig. 3). Whereas we detected 52 species of bird (‘available models’) in our surveys of their habitat, the lyrebirds in our study mimicked regularly only 4 of these (7.7% of available models). However, despite this limited number of mimicked models, males tended to mimic more often species that were common in their environment (Kendall’s tau-b τb = 0.328, N = 52; P = 0.003).
(ii, iv) does lyrebird mimicry reflect the species assemblage of the lyrebird’s microhabitat?
Lyrebirds appeared to be more likely to mimic species that occupied a similar microhabitat to themselves, but the evidence was not straightforward. First, lyrebird D-song primarily comprised mimicry of species that, like lyrebirds, foraged in the shrub layer and/or on the forest floor (Welch’s ANOVA: F3,29.8 = 14.2, P < 0.0001: Fig. 4B). This was not simply an artefact of the distribution of species because canopy/upper story and generalist feeders were more commonly recorded in the environment (Welch’s ANOVA: F3,56 = 46.6, P < 0.0001: Fig. 4 A).

Abundance of avian species as classified by microhabitat (forest strata) in (A) general environment as surveyed by sight and sound (N = 27 surveys) and (B) lyrebird D-song mimicry measured as time micked (N = 15 songs from 15 males)
Second, male lyrebirds were marginally more likely to mimic species that were recorded on lyrebird display mounds (as determined from mound surveys using camera trap footage) compared to those that were recorded only in the general environment (as determined from the general environment surveys; Fisher’s exact test, P = 0.0447). Indeed, the two heterospecific species that were most commonly observed on mounds (yellow-throated scrubwrens and white-browed scrubwrens) were also imitated by lyrebirds during D-song (Fig. 3; Table 1). Overall, however, we did not find a relationship between abundance of species observed on mounds and those detected in lyrebird mimicry, (Kendall’s tau-b τb = -0.183, N = 18; P = 0.330), suggesting that additional factors are important for explaining the mimetic composition of D-song.
(iii) do lyrebirds mimic the species that forage with them?
There was also equivocal evidence that lyrebirds preferentially imitated species that commonly associated with them. In most of the mound videos that showed at least one species of scrubwren, a male lyrebird was also present (white-browed scrubwrens: 34 of 53; yellow-throated scrubwrens: 34 of 59), which is consistent with reports that these two species of scrubwren may have a feeding association with lyrebirds (Higgins et al. 2001). However, the three other species commonly imitated by lyrebirds (eastern yellow robins, brown thornbills, grey shrike-thrushes) were not filmed on lyrebird mounds (of a total of 34 h of videos).
(v) do lyrebirds mimic species that are particularly effective at generating mobbing flocks?
There was no evidence that the species imitated by lyrebirds were unusually effective at generating mobbing flocks. Playbacks of avian alarm calls were successful in attracting other birds (Table 2): the number of individuals present within 15 m of the playback speaker immediately after broadcasting recorded alarm calls was greater than the number of individuals present before playback experiments (ANOVA-with blocks, F1,39 = 11.0, P = 0.002; Fig. 2 A). However, there was no evidence that any of the mobbing calls of the four species broadcast during the experiment was more effective than the others at attracting individual birds (Fig. 2B, ANOVA F3,36 = 0.626, P = 0.602).

Number of individuals attracted within 15 m of the speaker after the onset of the playback track (A) across all playbacks of alarm calls (N = 40) and (B) by playback treatment of mobbing alarm calls (N = 10 per each): eastern yellow robin (EYRO); white-browed scrubwren (WBSC); yellow-throated scrubwren (YTSC); grey fantail (GRFT)
(vi) do lyrebirds mimic calls typical of terrestrial mobbing choruses?
Lyrebirds preferentially imitated species that participated in mobbing flocks. Of the 54 bird species that we detected in our surveys and using camera traps, the species imitated by lyrebirds were more likely to approach within 15 m of the playback speaker broadcasting mobbing calls (70%) than those not imitated by lyrebirds (16%; Fisher’s exact test, P = 0.0013). However, the distribution of species mimicked by lyrebirds during D-song (Table 1) and the species attracted to the playback were different (Table 2; Kendall’s tau b, τb = -0.073, N = 17 P = 0.719). Four of the seven species most often mimicked by lyrebirds during D-song (Fig. 3; Table 1) were attracted to the playback (Table 2). However, several species of honeyeater also commonly responded to playbacks of alarm calls, as did the canopy foraging striated thornbills, even though during D-song male lyrebirds in our study rarely mimicked honeyeaters and never imitated striated thornbills.
Discussion
Here we show that during their song and dance display, all male lyrebirds in our study mimicked a restricted suite of model species that formed only a small subset of the potential model species available in their environment. The set of mimicked species was similar among males, and the three top species mimicked – making up the bulk of the song – were the same as at a distant, discrete population of lyrebirds 640 km away (Dalziell et al. 2021), providing evidence for selection maintaining model choice. Our assessment of several hypotheses for the drivers of this discrete mimetic repertoire suggests that males preferentially imitate the mobbing alarm calls of relatively common heterospecific birds that occupy niches that overlap with lyrebirds (hypotheses ii, iii). Thus, we conclude that during their song and dance display, male superb lyrebirds imitate a complex environmental acoustic cue that is expected to be highly informative about terrestrial predatory threats to other lyrebirds.
Superb lyrebirds regularly imitate multiple models, typically of other species of bird, by vocally imitating sounds in their environment (Bell 1976; Zann and Dunstan 2008), and this remarkable mimetic ability has led to the long-held but erroneous assumption that males copy sounds indiscriminately (e.g., Collins 1802; Robinson 1991; Carty 2014). In contrast, our study shows that males are highly selective about which species they mimic during the D-song component of their multicomponent displays. The extreme mimetic versatility of male superb lyrebirds renders it unlikely that the restricted mimetic content of lyrebird D-song is constrained by the cognitive or physiological limitations of males, as is suspected to be the case in northern mockingbirds Mimus polyglottos (Gammon 2013; Gammon and Corsiglia 2019) and icterine warblers Hippolais icterina (Riegert and Jůzlová 2018). Male lyrebirds (Dalziell et al. 2022; Zann and Dunstan 2008), including those in this study (unpublished data), regularly mimic multiple songs and calls of 20–25 different species of bird during their daily performances of ‘recital’ displays, which make up more than 97% of male vocal output (Dalziell et al. 2022). Commonly mimicked sounds in recital displays range from the calls of the 0.8 kg yellow-tailed black cockatoo Zanda funerea to the songs and calls of small passerines more than 2 orders of magnitude smaller than lyrebirds (Zann and Dunstan 2008; Dalziell and Magrath 2012; Dalziell et al. 2022; Dalziell and Welbergen 2021; see Higgins et al. 2001). Furthermore, captive males show extreme mimetic versatility, and males have been recorded imitating complex anthropogenic sounds such as camera shutters, emergency vehicle sirens, chainsaws and human baby cries (e.g., Attenborough 1998; Dalziell and Welbergen 2021; see Higgins et al. 2001). The drivers of model selection in recital displays remain poorly understood (Zann and Dunstan 2008; Dalziell et al. 2022), and so is a fruitful area for further research. Nevertheless, the set of models mimicked during recital displays differ consistently and predictably from the set mimicked within the D-song component of dance displays (Dalziell et al. 2022). Thus, our study of D-song adds to a growing body of evidence that, despite a clear capacity to imitate a wide variety of sounds, male superb lyrebirds are highly selective about what they imitate and when.
Males perform D-song sparingly but at key moments of their courtship cycle—during assessment by potential mates and during copulation itself—and so D-song is implicated directly in male reproductive success (Dalziell et al. 2021, 2022). To better understand the drivers of mimetic D-song, we tested several hypotheses for the small suite of model species that make up this song. We found that D-song did not simply reflect the abundance of species in the lyrebird’s general environment (hypothesis i), and neither were males mimicking rare species (iv). Furthermore, males did not appear to mimic species that were particularly successful at generating mobbing flocks as would be expected if D-song functioned to generate a mobbing flock (v). Instead, males appeared to be mimicking species that were relatively common in the lyrebird’s microhabitat (ii), especially species that contribute to terrestrial mobbing flocks (vi).
There was some evidence that male lyrebirds mimicked species that are thought to have a feeding association with lyrebirds (hypothesis iii). Two species commonly mimicked by males in our study, the white-throated and the yellow-throated scrubwren, have often been observed feeding with lyrebirds (e.g., Smith 1988). Our camera trap footage showed these species do indeed forage while ‘piloting’ lyrebirds closely (see also Supplementary Videos 2–3). Two other species commonly imitated by lyrebirds, the eastern yellow robin and the grey shrike-thrush, are also reported to regularly or opportunistically forage with lyrebirds (Higgins et al. 2001), but we did not capture any footage of this behaviour. This absence of footage is not surprising because eastern yellow robins and grey shrike-thrushes spend very little time on the ground, and typically capture terrestrial prey via quick dashes to the ground from a low perch. Our cameras were focused on the cleared ground of lyrebird display mounds so they were unlikely to film these species that mainly reside in the forest understory.
One question arising from these results is how lyrebirds acquire mobbing flock mimicry and what role the mechanism plays in model selection. In birds that acquire their vocal mimicry via vocal production learning (sensu Vernes et al. 2021), an individual can develop vocal mimicry either by copying the model species directly or by copying the mimicry produced by conspecifics (Dalziell et al. 2015). There is good evidence that male Albert’s lyrebirds M. alberti learn their vocal mimicry from both the models directly and from other male lyrebirds (Putland et al. 2006; Backhouse et al. 2022) and both mechanisms may be at play in male superb lyrebirds too (e.g., Smith 1988). It seems unlikely social transmission alone would be sufficient to maintain the complex acoustic illusion of a mixed-species mobbing flock males produce, of which mobbing alarm calls is only one component (Dalziell et al. 2021). Furthermore, it is unclear how social conformity could result in the agreement in key model species in D-song between this population and the distant (640 km) Sherbrooke Forest population studied previously (Dalziell et al. 2021), given that uniformity in socially transmitted vocalisations is rarely maintained over such broad spatial scales (Catchpole and Slater 2008), and is even less likely in species with patchy habitat (Laiolo and Tella 2005) and sedentary life histories (Podos and Warren 2007; Catchpole and Slater 2008), like lyrebirds (Higgins et al. 2001; Backhouse et al. 2021). Instead, such similarity among allopatric populations is consistent with a species-wide functional role for mimicry of terrestrial mobbing flocks. Nevertheless, small differences in the suite of models mimicked by male lyrebirds between the Blue Mountains population studied here and the Sherbrooke Forest population studied previously (Dalziell et al. 2021) suggest a role for social transmission of mimicry. Specifically, while grey shrike-thrushes and ringtail possums are common in both study populations they were only mimicked by Blue Mountains and Sherbrook lyrebirds respectively.
Variation among males within the Blue Mountains population studied here suggests that males directly copy model species, at least in part. For example, only 3 of 15 males imitated the alarm calls of yellow-throated scrubwrens (Table 1; Fig. 3), perhaps because this species is restricted to the wettest patches of our study site. Similarly, just one male mimicked the alarm calls of multiple species of honeyeater, possibly reflecting the atypical location of his display mounds close to flowering shrubs favoured by honeyeaters. Learning vocal mimicry indirectly from other lyrebirds has the advantage that it would allow learning of the sexual display context along with the vocalisations, while copying heterospecifics directly would not. However, copying models directly allows individuals to flexibly adjust their vocal mimicry (Dalziell et al. 2015). During D-song male lyrebirds may mimic the alarm calls that are most typical of their region or even their display territory, and so tailor their mimicry to the specific microhabitat where they perform their sexual display. In this way males could enhance the efficacy of their mimicry by mimicking the alarm calls of species in the immediate surroundings of their display mounds. Determining the relative contributions of direct and indirect mechanism of vocal mimicry acquisition in the D-song of the superb lyrebird will require experiments and comparisons of D-song models among multiple different lyrebird populations.
Previous work has shown that with D-song, male lyrebirds generate an acoustic illusion of the presence of a predator by mimicking a mixed-species mobbing flock (Dalziell et al. 2021). D-song is proposed to be used by males to arm a ‘sensory trap’ (sensu West-Eberhard 1979; Christy 1995) to prevent fertile females prematurely terminating sexual encounters by manipulating female anti-predator responses (Dalziell et al. 2021). Our examination of the relationship between the acoustic environment and D-song adds weight to this hypothesis by suggesting that males are specifically mimicking the terrestrial alarm calls of species that can provide the most fitness-relevant information to female lyrebirds – i.e., those heterospecifics that forage on the ground or in the forest understory, including species that specifically and regularly forage alongside lyrebirds. However, how females respond to D-song and real mobbing flock mimicry will need to be determined before we can fully understand this astonishing mimetic signal.
Superb lyrebirds are only one of many bird species where males regularly incorporate imitations of heterospecific sounds into their sexual displays (Baylis 1982; Garamszegi et al. 2007; Catchpole and Slater 2008; Dalziell et al. 2015; Goller and Shizuka 2018). It has been common to assume that such highly versatile vocal ‘mimics’ imitate models passively (Robinson 1975; Hindmarsh 1986), and so such imitations would not conform to functional definitions of mimicry, whereby the mimic benefits from the receiver perceiving the similarity between the mimic and the model (Dalziell et al. 2015; Dalziell and Welbergen 2016b). Instead, our work shows that model selection in these sexual displays can be highly restricted and are likely based on functional ecological relationships between the receiver and the models.
Appendices
Supplementary Video 1
An audio-video recording of an adult male superb lyrebird performing a song and dance display on his display mound, finishing with vocal mimicry of a mixed-species mobbing flock (D-song). The male mimics heterospecific mobbing calls between 0:23 and 0:32 and again between 0:37 and 0:43. Species mimicked in the first bout of mimicry, from beginning to end, are eastern yellow robin, brown thornbill, and grey shrike-thrush. In the second bout of mimicry, the male mimics eastern yellow robin, brown thornbill, white-browed scrubwren, and yellow-throated scrubwren in that order. Video archived in Cornell Lab of Ornithology’s Macaulay Library, ML475994: https://macaulaylibrary.org/asset/475994.
Supplementary Video 2
An audio-video recording of an adult male superb lyrebird performing a song and dance display on his display mound, whilst nearby are two model species that are regularly mimicked by male lyrebirds during D-song. The male mimics heterospecific mobbing calls between 0:34 and 0:36 and again between 0:48 and 1:00. In the first bout of mimicry the male mimics the mobbing-alarm calls of the white-browed scrubwren. In the second bout of mimicry, the male mimics the mobbing alarm calls of white-browed scrubwren, yellow-throated scrubwren, and eastern yellow robin in that order. A white-browed scrubwren can be seen just left of the male’s mound from 0:18–0:28. A yellow-throated scrubwren can be seen entering the mound from the left of the male’s mound at 0:28 s. Both species were often filmed alongside male lyrebirds on their mounds.Video archived in Cornell Lab of Ornithology’s Macaulay Library, ML465925141: https://macaulaylibrary.org/asset/465925141.
Supplementary Table 1
Recordings of the model species mobbing calls used during playback experiments are archived in Cornell Lab of Ornithology’s Macaulay Library under the catalogue numbers listed below (see https://macaulaylibrary.org)
Macaulay Library catalogue number | Common name | Scientific name | Recordist |
|---|---|---|---|
118,237,411 | Eastern yellow robin | Eopsaltria australis | Taylor L. Crisologo |
206,299 | Eastern yellow robin | Eopsaltria australis | Taylor L. Crisologo |
206,298 | Eastern yellow robin | Eopsaltria australis | Taylor L. Crisologo |
119,296,171 | Eastern yellow robin | Eopsaltria australis | Sarah A. Dzielski |
206,278 | Eastern yellow robin | Eopsaltria australis | Taylor L. Crisologo |
206,294 | Grey fantail | Rhipidura albiscapa | Taylor L. Crisologo |
206,212 | Grey fantail | Rhipidura albiscapa | Taylor L. Crisologo |
206,523 | Grey fantail | Rhipidura albiscapa | Sarah A. Dzielski |
206,522 | Grey fantail | Rhipidura albiscapa | Sarah A. Dzielski |
206,521 | Grey fantail | Rhipidura albiscapa | Sarah A. Dzielski |
206,287 | Yellow-throated scrubwren | Sericornis citreogularis | Taylor L. Crisologo |
206,286 | Yellow-throated scrubwren | Sericornis citreogularis | Taylor L. Crisologo |
206,283 | Yellow-throated scrubwren | Sericornis citreogularis | Taylor L. Crisologo |
118,239,091 | Yellow-throated scrubwren | Sericornis citreogularis | Taylor L. Crisologo |
206,249 | Yellow-throated scrubwren | Sericornis citreogularis | Sarah A. Dzielski |
206,520 | White-browed scrubwren | Sericornis frontalis | Sarah A. Dzielski |
206,519 | White-browed scrubwren | Sericornis frontalis | Sarah A. Dzielski |
206,251 | White-browed scrubwren | Sericornis frontalis | Sarah A. Dzielski |
206,244 | White-browed scrubwren | Sericornis frontalis | Sarah A. Dzielski |
206,235 | White-browed scrubwren | Sericornis frontalis | Sarah A. Dzielski |
Supplementary Figure S1
The study site in the Jamison/Kedumba Valleys of the Blue Mountains NP. (A) Satellite image of the study site with the groups of sound-recorded male superb lyrebirds denoted by yellow markers labeled with the number recorded in each group (total males recorded = 15) (Google Earth, 2021). (B) A photo of the western side of the field site facing to the north-east (photo by Justin Welbergen).

Data Availability
We uploaded our survey data to eBird.org (Sullivan et al. 2009), a citizen science database that collects bird observations and sampling effort information from bird watchers around the world. Sound recordings of model species are archived in the Macaulay Library, Cornell Lab of Ornithology (https://www.macaulaylibrary.org/, for catalogue numbers see Supplementary Table 1). Data are available within the text or will be deposited in publicly available repositories.
Code Availability
All statistical analysis was carried out in JMP Pro 12 (v. 12.0.1, SAS Institute Inc) except for the tests for association, which were performed in SPSS (v. 24.0, IBM Cop.).
References
Alquezar RD, Machado RB (2015) Comparisons Between Autonomous Acoustic Recordings and Avian Point Counts in Open Woodland Savanna. Wilson J Ornithol 127:712–723
Arnqvist G (2006) Sensory exploitation and sexual conflict. Philosophical Trans Royal Soc Lond Ser B-Biological Sci 361:375–386
Attenborough D (1998) Signals and songs. In: Salisbury M (ed) The Life of Birds. BBC Natural History Unit, Bristol
Backhouse F, Dalziell A, Magrath H, Welbergen RD J, A (2022) Sequences of vocal mimicry performed by male Albert’s lyrebirds are socially transmitted and enhance acoustic contrast.Proc R Soc Lond, Ser B: Biol Sci289
Backhouse F, Dalziell AH, Magrath Robert D, Rice A, Crisologo N, Welbergen TL JA (2021) Differential geographic patterns in song components of male Albert’s lyrebirds. Ecol Evol 11:2701–2716
Baylis JR (1982) Avian vocal mimicry: its function and evolution. In: Kroodsma DE, Miller EH, Ouellet H (eds) Acoustic Communication in Birds. Academic Press, New York, pp 51–83
Bell K (1976) Song of the superb lyrebird in south-eastern new south wales, with some observations on habitat. Emu 76:59–63
Bro-Jørgensen J, Pangle WM (2010) Male topi antelopes alarm snort deceptively to retain females for mating. Am Nat 176:E33–E39
Carlson NV, Griesser M (2022) Chapter One - Mobbing in animals: A thorough review and proposed future directions. In S. Healy & J. Podos (Eds.), Advances in the Study of Behavior. Academic Press, pp. 1-41
Carlson NV, Healy SD, Templeton CN (2018) Mobbing Curr Biol 28:R1081–R1082
Caro T (2005) Antipredator Defenses in Birds and Mammals. University of Chicago Press, Chicago
Carty S (2014) Hear the amazing lyrebird who can mimic all the birds of the forest and even gives his best camera lens impression. In: Daily mail, Australia
Catchpole CK, Slater PJB (2008) Bird Song: Biological Themes and Variations, 2nd edn. Cambridge University Press, Cambridge
Cheney KL, Grutter AS, Marshall NJ (2008) Facultative mimicry: cues for colour change and colour accuracy in a coral reef fish. Proc R Soc Lond Ser B: Biol Sci 275:117–122
Christy JH (1995) Mimicry, mate choice, and the sensory trap hypothesis. Am Nat 146:171–181
Chu M (2001) Heterospecific responses to scream calls and vocal mimicry by phainopeplas (Phainopepla nitens) in distress. Behaviour 138:775–787
Collins D (1802) An account of the English Colony in New South Wales, vol 2. Cadell & Davies, London
Dalziell AH, Magrath RD (2012) Fooling the experts: accurate vocal mimicry in the song of the superb lyrebird, Menura novaehollandiae. Anim Behav 83:1401–1410
Dalziell AH, Maisey AC, Magrath RD, Welbergen JA (2021) Male lyrebirds create a complex acoustic illusion of a mobbing flock during courtship and copulation. Curr Biol 31:1970–1976 e4
Dalziell AH, Peters RA, Cockburn A, Dorland AD, Maisey AC, Magrath RD (2013) Dance choreography is coordinated with song repertoire in a complex avian display. Curr Biol 23:1132–1135
Dalziell AH, Welbergen JA (2016a) Elaborate mimetic vocal displays by female superb lyrebirds. Front Ecol Evol 4:1–13
Dalziell AH, Welbergen JA (2016b) Mimicry for all modalities. Ecol Lett 19:609–619
Dalziell AH, Welbergen JA (2021) ‘Bloody fool!’: why Ripper the musk duck, and many other talkative Aussie birds, are exciting biologists. The Conversation. The Conversation, Australia
Dalziell AH, Welbergen JA (2022) Male Superb Lyrebirds (Menura novaehollandiae) perform an ornate multimodal display immediately following copulation. Ibis 164:809–815
Dalziell AH, Welbergen JA, Igic B, Magrath RD (2015) Avian vocal mimicry: a unified conceptual framework. Biol Rev 90:643–668
Dalziell AH, Welbergen JA, Magrath RD (2022) Male superb lyrebirds mimic functionally distinct heterospecific vocalizations during different modes of sexual display. Anim Behav 188:181–196
Darras K, Batáry P, Furnas B, Celis-Murillo A, Van Wilgenburg SL, Mulyani YA, Tscharntke T (2018) Comparing the sampling performance of sound recorders versus point counts in bird surveys: A meta-analysis. J Appl Ecol 55:2575–2586
Davies N (2015) Cuckoo: cheating by nature. Bloomsbury Publishing
Flower TP (2011) Fork-tailed drongos use deceptive mimicked alarm calls to steal food. Proceedings of the Royal Society of London Series B-Biological Sciences 278:1548–1555
Flower TP, Gribble M, Ridley AR (2014) Deception by flexible alarm mimicry in an African bird. Science 344:513–516
Gammon DE (2013) How is model selection determined in a vocal mimic?: Tests of five hypotheses. Behaviour 150:1375–1397
Gammon DE, Corsiglia AM (2019) Mockingbirds imitate frogs and toads across North America. Behav Processes 169:103982
Garamszegi LZ, Eens M, Pavlova DZ, Aviles J, Moller AP (2007) A comparative study of the function of heterospecific vocal mimicry in European passerines. Behav Ecol 18:1001–1009
Garnett S, Franklin D, Ehmke G, VanDerWal JJ, Hodgson L, Pavey C, Reside AE, Welbergen JA, Butchart S, Perkins GC, Williams SE (2013) Climate Change Adaptation Strategies for Australian Birds. In. National Climate Change Adaptation Research Facility. Gold Coast, Queensland
Goller M, Shizuka D (2018) Evolutionary origins of vocal mimicry in songbirds. Evol Lett 2:417–426
Goodale E, Ratnayake CP, Kotagama SW (2014) Vocal mimicry of alarm-associated sounds by a drongo elicits flee and mobbing responses from other species that participate in mixed-species bird flocks. Ethology 120:266–274
Herberstein ME (2011) Spider Behaviour: Flexibility and Versatility. Cambridge University Press, Cambridge, United Kingdom
Higgins PJ, Peter JM, Steele WK (2001) Superb lyrebird (Menura novaehollandiae). Handbook of Australian, New Zealand and Antarctic Birds Volume 5: Tyrant-flycatchers to Chats. Oxford University Press, Melbourne, pp 142–173
Hindmarsh AM (1986) The functional-significance of vocal mimicry in song. Behaviour 99:87–100
Igic B, Magrath RD (2014) A songbird mimics different heterospecific alarm calls in response to different types of threat. Behav Ecol 25:538–548
Igic B, McLachlan J, Lehtinen I, Magrath RD (2015) Crying wolf to a predator: deceptive vocal mimicry by a bird protecting young. Proc R Soc Lond Ser B: Biol Sci 282:1–7
Jackson RR, Wilcox RS (1990) Aggressive mimicry, prey-specific predatory behaviour and predator-recognition in the predator-prey interactions of Portia fimbriata and Euryattus sp., jumping spiders from Queensland. Behav Ecol Sociobiol 26:111–119
Laiolo P, Tella JL (2005) Habitat fragmentation affects culture transmission: patterns of song matching in Dupont’s lark. J Appl Ecol 42:1183–1193
Magrath RD, Haff TM, Fallow PM, Radford AN (2015) Eavesdropping on heterospecific alarm calls: from mechanisms to consequences. Biol Rev 90:560–586
Marler P (2004) Bird calls: a cornucopia for communication. In: Marler P, Slabbekoorn H (eds) Nature’s Music: The Science of Birdsong. Elsevier Academic Press, San Diego, pp 132–177
Menkhorst P, Rogers DI, Clarke R, Davies J, Marsack N, Franklin P K (2017) The Australian Bird Guide. CSIRO Publishing, Clayton, Victoria
Morrison CA, Auniņš A, Benkő Z, Brotons L, Chodkiewicz T, Chylarecki P, Escandell V, Eskildsen DP, Gamero A, Herrando S, Jiguet F, Kålås JA, Kamp J, Klvaňová A, Kmecl P, Lehikoinen A, Lindström Ã, Moshøj C, Noble DG, Øien IJ, Paquet JY, Reif J, Sattler T, Seaman BS, Teufelbauer N, Trautmann S, van Turnhout CAM, Vořišek P, Butler SJ (2021) Bird population declines and species turnover are changing the acoustic properties of spring soundscapes. Nat Commun 12:6217
Nakano R, Takanashi T, Skals N, Surlykke A, Ishikawa Y (2010) Ultrasonic courtship songs of male Asian corn borer moths assist copulation attempts by making the females motionless. Physiol Entomol 35:76–81
Nelson X, Jackson R (2011) Flexibility in the foraging strategies of spiders. In: Herberstein ME (ed) Spider behaviour: flexibility and versatility. Cambridge University Press, Cambridge, pp 31–56
Norman MD, Finn J, Tregenza T (2001) Dynamic mimicry in an Indo-Malayan octopus. Proceedings of the Royal Society of London Series B-Biological Sciences 268:1755–1758
Pavlacky DC, Possingham HP, Goldizen AW (2015) Integrating life history traits and forest structure to evaluate the vulnerability of rainforest birds along gradients of deforestation and fragmentation in eastern Australia. Biol Conserv 188:89–99
Podos J, Warren PS (2007) The evolution of geographic variation in birdsong. Adv Study Behav 37:403–458
Putland DA, Nicholls JA, Noad MJ, Goldizen AW (2006) Imitating the neighbours: vocal dialect matching in a mimic-model system. Biol Lett 2:367–370
Riegert J, Jůzlová Z (2018) Vocal mimicry in the song of Icterine warblers (Hippolais icterina): possible functions and sources of variability. Ethol Ecol Evol 30:430–446
Robinson FN (1975) Vocal mimicry and the evolution of bird song. Emu 75:23–27
Robinson FN (1991) Phatic communication in bird song. Emu 91:61–63
Smith LH (1988) The Life of the Lyrebird. William Heinemann Australia, Melbourne
Sullivan BL, Wood CL, Iliff MJ, Bonney RE, Fink D, Kelling S (2009) eBird: A citizen-based bird observation network in the biological sciences. Biol Conserv 142:2282–2292
Tarsitano M, Jackson RR, Kirchner WH (2000) Signals and signal choices made by the araneophagic jumping spider Portia fimbriata while hunting the orb-weaving web spiders Zygiella xnotata and Zosis geniculatus. Ethology 106:595–615
Vernes SC, Kriengwatana BP, Beeck VC, Fischer J, Tyack PL, ten Cate C, Janik VM (2021) The multi-dimensional nature of vocal learning. Philosophical Trans Royal Soc B: Biol Sci 376:20200236
West-Eberhard MJ (1979) Sexual selection, social competition, and evolution. Proceedings of the American Philosophical Society:222–234
Wheatcroft D, Price TD (2013) Learning and signal copying facilitate communication among bird species. Proceedings of the Royal Society of London Series B-Biological Sciences 280:1471–2954
Zann R, Dunstan E (2008) Mimetic song in superb lyrebirds: species mimicked and mimetic accuracy in different populations and age classes. Anim Behav 76:1043–1054
Acknowledgements
We acknowledge the Traditional Owners of the land on which this work was conducted, the Dharug and Gundungurra people. We pay our respects to the Elders past and present. We thank the editors David Outomuro, Bibiana Rojas, and Matthew Symonds, and the two reviewers for helpful comments We would also like to thank I. Lovette and D. Bonter for valuable advice on manuscript drafts; J. McGowan, M. Medler, G. Moore, and J. Morano for advice on acoustic data collection and analysis; C. Parkinson and A. Thomas for assistance with the video data analysis; and D. Veronese, E. Timmiss and S. Yabsley for field assistance. We thank the Australian Bird and Bat Banding Scheme for the supply of bird bands.
Funding
This study was funded by the Cornell Lab of Ornithology’s Ivy Expeditions for funding the project (TLC, SAD and JRP). Additional support was provided by the Cornell Lab of Ornithology’s Macaulay Library (MSW), the Cornell Lab of Ornithology Rose Postdoctoral Fellowship Program (AHD), the Hawkesbury Institute for the Environment, Western Sydney University (AHD and JAW) and a Vice-Chancellor’s Research Fellowship from the University of Wollongong (AHD).
Open Access funding enabled and organized by CAUL and its Member Institutions
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest/Competing interests
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Ethics approval
All work was approved under Australian Animal Ethics legislation (#A10699) and Cornell University institutional policy (IACUC: #2009 − 0105). The work also adhered to the legal requirements for research on wild native birds in Australia (Australian Bird and Bat Banding Scheme and Office of Environment and Heritage NSW #SL101351).
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Crisologo, T.L., Dzielski, S.A., Purcell, J.R. et al. Selective alarm call mimicry in the sexual display of the male superb lyrebird (Menura novaehollandiae). Evol Ecol 37, 245–266 (2023). https://doi.org/10.1007/s10682-022-10200-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10682-022-10200-w