
Abstract
The Functional Response (FR) has been identified as a powerful predictive tool to forecast the ecological impacts of existing, emerging and future invasive alien species. In particular, the parameters of attack rate a and handling time h may be predictive of the ecological impacts of invaders when utilised in comparison with trophically analogous natives. However, researchers in many cases face somewhat contradictory impact predictions based on the use of one parameter or the other. Here, we thus propose a new metric, the Functional Response Ratio (FRR), which is simply a divided by h: that is, FRR = a/h. Given that high values of a and low values of h should associate with high impact, and vice versa, the FRR metric balances the information from both parameters. This also resolves contradictions when one parameter gives opposite predictions to the other. Using multiple examples obtained from the literature, we find that the FRR indeed resolves such contradictions and that values of FRR of invaders are consistently higher than those of natives, irrespective of experimental or environmental context. Accordingly, the use of FRR provides a novel and reliable metric for scientists, stakeholders and practitioners to predict the ecological impacts of existing, emerging and future invasive alien species across taxa and trophic groups.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
A distinct lack of methodological advancement has impeded the quantification of the ecological impacts of invasive alien species and thus the transformation of invasion biology into a more predictive science (Dick et al. 2014, 2017a, b, c). Despite an improved understanding in recent years of the various ecological impacts associated with invasive species (Simberloff et al. 2013; Kumschick et al. 2015), attempts to identify single and multiple species-specific traits that are predictive of ecological impact have repeatedly failed (Parker et al. 1999; Dick et al. 2014, 2017a, b, c). However, the use of classical ecological concepts, in particular the Functional Response (FR), Numerical Response (NR) and predator ‘switching’, has more recently been identified as powerful means to quantify the ecological impacts of invasive alien species (Dick et al. 2014, 2017a, b, c; Cuthbert et al. 2018a, b; Dickey et al. 2018; Joyce et al. 2019).
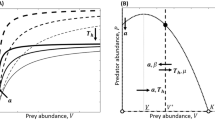
In particular, examinations of FRs, which define the relationship between resource availability and consumer uptake (Solomon 1949; Holling 1959), and generate, for example, estimated maximum feeding rates, have proven useful in deriving per capita impacts of invasive alien species when used comparatively with trophically analogous native species (e.g. Bollache et al. 2008; Dick et al. 2013, 2014, 2017a, b, c; Alexander et al. 2014). Centrally, the attack rate a and handling time h parameters of FRs offer insights into the ecological impacts of invaders, with a corresponding to impact at low prey/resource densities (i.e. initial FR slope) and h corresponding to impact through estimated maximum feeding rates (i.e. 1/h; FR asymptote). Whilst the utility of these two parameters in quantifications of ecological impact has been confirmed (Dick et al. 2014, 2017a, b, c), contradictory impact predictions can often emerge when assessment is based on one parameter or the other. To illustrate this, Fig. 1 shows that low a and high h (blue) indicate low impact, while high a and low h (red) indicate high impact; however, impact is more difficult to discern from these individual parameters in the yellow zones, which might also indicate intermediate impact. Thus, considering a and h as separate entities may impede robust assessment and prediction of invader impacts. In the present study, we thus present a novel metric, which combines the a and h parameters of the FR, increasing the rapidity, robustness and reliability of predictions of invader impact. We then validate the metric using published examples from the FR literature.

Hypothetical Functional Response (FR) ‘heat map’ demonstrating differences in ecological impact based on attack rates (a; initial slope of curve) and handling times (h; which inversely gives maximum feeding rates i.e. 1/h, curve asymptote) of invasive and native species. Increasing per capita impact is illustrated from bottom left to top right
We propose the Functional Response Ratio (FRR) as:
where a is the attack rate and h is the handling time from a FR curve.
We now present an example of the utility of FRR over individual FR parameters. Dick et al. (2013) used comparative FR parameters to quantify impacts of the invasive bloody red shrimp, Hemimysis anomala, relative to analogous native mysids. This invader is known to have large ecological impacts, driving some species of zooplankton to local extinction (e.g. Ricciardi et al. 2012). Towards one prey species, the invader displayed a lower attack rate compared to the native analogue (a: invader, 3.05; native, 3.65), but also a shorter handling time (h: invader, 0.01; native, 0.07) and hence higher maximum feeding rate. Here, the predictability of invader impact is contradictory, with the invader potentially classified as less impactful than the native based on the a parameter, but more impactful when the h parameter is considered. However, when our new FRR metric is used, higher predicted impact of the invader based on the combination of parameters is clear (FRR: invader, 305.00; native, 52.14). Similarly, towards another prey species, Barrios-O’Neill et al. (2014) found that H. anomala exhibited a higher attack rate compared to the native analogue (a: invader, 0.79; native, 0.19), but a longer handling time (h: invader, 0.14; native, 0.13) and hence lower feeding rate. This suggests that the invader is more impactful based on the a parameter, but less impactful based on the h parameter. Here, again, when FRR is applied, the predictability of impact in favour of the invasive species is highly evident (FRR: invader, 5.64; native, 1.46). Thus, FRR is a metric that is fully consistent with actual field impacts in this case, resolving contradictions of individual FR parameters, and we propose that FRR should provide a more robust and reliable quantification of invader ecological impact in general. That is, as the FRR metric captures both parameters, analyses based on FRR alone negate the need for interpretation of a or h in isolation.
To demonstrate the overall power of FRR in predicting the ecological impacts of invasive alien species, across multiple taxa and trophic groups, we collated paired native/invader a and h parameters from comparative FR studies, following the review and search terms of Dick et al. (2014). We selected invasive species with known high ecological impacts. Depending on data availability, either starting parameter estimates or means from bootstrapped re-sampling were used. Where individual studies derived multiple FRs for given invasive/native species across several experimental contexts (e.g. prey types, environmental variation), mean a and h values were discerned for each species prior to analysis across the given contexts. Similarly, if multiple analogous native species were presented as comparators in a given study against a single invader, the mean a and h values were obtained across all the native species used. Given the inherent differences in parameter estimation, only paired invasive/native species scenarios that used matched FR models were selected (i.e. Holling 1959; Rogers 1972). A paired two-sided Wilcoxon signed rank test was used to derive whether FRRs were significantly different between invaders and natives across all study systems at a significance level of p < 0.05.
Functional Response Ratios (FRR) were discerned for 13 invasive species across 17 studies, totalling 22 invader/native pairings (Table S1). We found FRRs to be very significantly higher for invasive species in comparison to native analogues (V = 249.00, p < 0.001; Fig. 2). Therefore, the new FRR metric is successful in differentiating ecologically damaging invasive species across a variety of studies spanning multiple ecological systems, habitats, taxa and trophic groups.

Mean (± SE) Functional Response Ratio (FRR: a/h) values for paired groups of trophically analogous invasive and native species (n = 22 pairs; see Table S1)
Whilst the use of comparative FRs has proven a powerful means of assessment and prediction of the ecological impact of invasive alien species (Dick et al. 2014, 2017a, c; but see Vonesh et al. 2017a, b; Dick et al. 2017b), it has often been unclear which estimated parameters from the FRs are most appropriate for use in species comparisons. Indeed, contradictions of impact prediction can emerge when the parameters are considered in isolation (see above). However, based on the amalgamation of the attack rate a and handling time h parameters of FR models, the use of FRR provides a novel approach to quantify the per capita impacts of invasive alien species. Further, in the present study, known ecologically damaging invasive species displayed significantly greater FRR (a/h) than trophically analogues native comparators. Thus, the use of FRR provides a powerful, yet simplistic blend of these integral FR parameters for species comparative purposes. Furthermore, the combination of FRRs (i.e. per capita impact) with consumer Numerical Response proxies (e.g. abundance) could enable improved derivation of invader Relative Impact Potential (RIP) at the population-level (Dick et al. 2017c; Dickey et al. 2018). Importantly, our results span multiple ecological systems and demonstrate applicability to impact predictions in any consumer-resource pairing where the focal parameters, or similar, can be estimated. Furthermore, we propose that FRR will be a useful probe into the key behavioural components of invaders that lead to ecological impact, such as those driven by invader ‘novel weapons’ and native prey ‘naïveté’. For example, we predict high levels of these will be characterised by FRs towards the top right of Fig. 1, while eco-evolutionary processes could lead to FRs towards the lower left of Fig. 1. We therefore propose the use of FRR as a means to amalgamate and compare the core parameters of FR models between invasive and native species and in multiple other contexts involving assessment of ecological impacts. This simple and rapid predictive tool will better inform researchers, managers, policy makers and practitioners as to the potential ecological impacts of existing, emerging and future invasive species.
References
Alexander ME, Dick JTA, Weyl OLF, Robinson TB, Richardson DM (2014) Existing and emerging high impact invasive species are characterized by higher functional responses than natives. Biol Lett 10:2–6
Barrios-O’Neill D, Dick JTA, Emmerson MC, Ricciardi A, MacIsaac HJ et al (2014) Fortune favours the bold: a higher predator reduces the impact of a native but not an invasive intermediate predator. J Anim Ecol 83:693–701
Bollache L, Dick JTA, Farnsworth KD, Montgomery WI (2008) Comparison of the functional responses of invasive and native amphipods. Biol Lett 4:166–169
Cuthbert RN, Dalu T, Wasserman RJ, Dick JTA, Mofu L, Callaghan A, Weyl OLF (2018a) Intermediate predator naïveté and sex-skewed vulnerability predict the impact of an invasive higher predator. Sci Rep 8:14282
Cuthbert RN, Dickey JWE, McMorrow C, Laverty C, Dick JTA (2018b) Resistance is futile: lack of predator switching and a preference for native prey predict the success of an invasive prey species. R Soc Open Sci 5:180339
Dick JTA, Gallagher K, Avlijas S, Clarke HC, Lewis SE et al (2013) Ecological impacts of an invasive predator explained and predicted by comparative functional responses. Biol Invasions 15:837–846
Dick JTA, Alexander ME, Jeschke JM, Ricciardi A, MacIsaac HJ (2014) Advancing impact prediction and hypothesis testing in invasion ecology using a comparative functional response approach. Biol Invasions 16:735–753
Dick JTA, Alexander ME, Ricciardi A, Laverty C, Downey PO et al (2017a) Functional responses can unify invasion ecology. Biol Invasions 19:1667–1672
Dick JTA, Alexander ME, Ricciardi A, Laverty C, Downey PO et al (2017b) Fictional responses from Vonesh et al. Biol Invasions 19:1677–1678
Dick JTA, Laverty C, Lennon JJ, Barrios-O’Neill D, Mensink PJ et al (2017c) Invader Relative Impact Potential: a new metric to understand and predict the ecological impacts of existing, emerging and future invasive alien species. J Appl Ecol 54:1259–1267
Dickey JWE, Cuthbert RN, Rea M, Laverty C, Crane K et al (2018) Assessing the relative potential ecological impacts and invasion risks of emerging and future invasive alien species. NeoBiota 40:1–24
Holling CS (1959) Some characteristics of simple types of predation and parasitism. Can Entomol 91:385–398
Joyce PWS, Dickey JWE, Cuthbert RN, Dick JTA, Kregting L (2019) Using functional responses and prey switching to quantify invasion success of the Pacific oyster, Crassostrea gigas. Mar Environ Res 145:66–72
Kumschick S, Gaertner M, Vilà M, Essl F, Jeschke JM et al (2015) Ecological impacts of alien species: quantification, scope, caveats and recommendations. Bioscience 65:55–63
Parker IM, Simberloff D, Lonsdale WM, Goodell K, Wonham M et al (1999) Impact: toward a framework for understanding the ecological effects of invaders. Biol Invasions 1:3–19
Ricciardi A, Avlijas S, Marty J (2012) Forecasting the ecological impacts of the Hemimysis anomala invasion in North America: lessons from other freshwater mysid introductions. J Great Lakes Res 38:7–13
Rogers D (1972) Random search and insect population models. J Anim Ecol 41:369–383
Simberloff D, Martin J, Genovesi P, Maris V, Wardle DA (2013) Impacts of biological invasions: what’s what and the way forward. Trends Ecol Evol 28:58–66
Solomon ME (1949) The natural control of animal populations. J Anim Ecol 18:1–35
Vonesh JR, McCoy M, Altwegg R, Landi P, Measey J (2017a) Functional responses can’t unify invasion ecology. Biol Invasions 19:1673–1676
Vonesh JR, McCoy M, Altwegg R, Landi P, Measey J (2017b) Rather than unifying invasion biology, Dick et al.’s approach rests on subjective foundations. Biol Invasions 19:1679–1680
Acknowledgements
RNC and PWSJ acknowledge funding from the Department for the Economy, Northern Ireland. JWED acknowledges funding from Inland Fisheries Ireland and the Department of Agriculture, Environment and Rural Affairs, Northern Ireland. NEC and JTAD are supported by the Irish Environmental Protection Agency research Grant 2015-NC-MS-4. Thanks also to the Natural Environment Research Council.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Cuthbert, R.N., Dickey, J.W.E., Coughlan, N.E. et al. The Functional Response Ratio (FRR): advancing comparative metrics for predicting the ecological impacts of invasive alien species. Biol Invasions 21, 2543–2547 (2019). https://doi.org/10.1007/s10530-019-02002-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-019-02002-z