
Abstract
Most studies on habitat selection among animals are conducted at local scales, whereas reliable determination of species requirements at larger spatial scales can be problematic. We used data available for NATURA 2000 sites to determine the habitat requirements of two relatively widespread and common species—pine marten Martes martes and stone marten Martes foina—in Southern Europe. Using presence-absence data, we applied statistical models at two spatial scales. At the local scale (within the dispersal distances of the species), sites occupied by martens were compared with unoccupied sites using buffers of different sizes, whereas at the regional scale, unoccupied sites were selected randomly. To adjust for spatial autocorrelation of data, penalised quasi-likelihood approximations were used. Both species of martens demonstrated preferences for areas with higher proportions of forest cover and steeper terrain. At the local, but not at the regional, scale, pine martens occurred at lower elevations, whilst stone martens occurred at higher elevations. We found that climatic variables (mean temperature, precipitation) had no significant effect on the presence of the martens. The results of our analyses generally confirmed findings of previous studies on habitat selection of both marten species in Southern Europe. This demonstrates the utility of data collected for NATURA 2000 sites for use in various analyses such as conservation planning and evaluating the impact of climate change on the distribution of animal species.
Similar content being viewed by others


Introduction
Understanding habitat requirements is fundamental for improving species’ management and conservation strategies (Law and Dickman 1998; Debinski and Holt 2000; Boyce et al. 2003). Patterns of habitat selection depend on the scale of analysis: at the landscape scale, habitat selection is regulated by geological and historical processes, and human-induced influences, whereas at the fine scale, it depends on decisions made by individual organisms that directly affect their fitness (Mayor et al. 2009). Although results of analyses at broader spatial scales usually suffer from a lack of adequate replication and often present patterns rather than processes, they provide context for interpreting results obtained at smaller scales (Bissonette 1997).
Most studies of carnivore habitat selection have been conducted at small, local scales and only a few at national scales (Virgos et al. 2012). Reliable determination of species requirements at large spatial scales is difficult, mainly due to data collection limitations. The methods used to analyse species-habitat relationships vary and include observations (e.g. from museum records), telemetry (VHF and GPS), remote monitoring (camera traps) and the more recently developed non-invasive DNA sampling and molecular techniques (e.g. Wereszczuk and Zalewski 2015; Vergara et al. 2016; Balestrieri et al. 2016; O’Mahony 2017). Telemetry or remote monitoring data are usually collected from relatively small areas (e.g. Wereszczuk and Zalewski 2015). In contrast, observational data covering larger areas are often collected non-systematically and can be biased by various factors (e.g. by the distribution of observers). The data collected for NATURA 2000 (N2K) sites, available in standard data forms (SDF), provide a unique opportunity to analyse habitat selection at different spatial scales, as the data contains not only information about the occurrence of different species but also an extensive description of the site and its ecology. To test the suitability of these records for determining habitat requirements, we decided to use data on a pair of relatively widespread and common species—the European pine marten Martes martes and the stone marten Martes foina.
Pine and stone martens are closely related medium-sized mustelids. Their distribution extensively overlaps in continental Europe (Proulx et al. 2004); however, the stone marten occurs from Southern to Eastern Europe whereas the pine marten also occurs in Northern Europe, which suggests it is better adapted to cold climates. Both species are morphologically and ecologically similar, with comparable life history characteristics (Mead 1994; Sato et al. 2012; Monterroso et al. 2016). Despite many similarities, these species differ with respect to their environmental preferences (Virgos et al. 2012) and in extreme cases they use completely separate habitats (Wereszczuk and Zalewski 2015). Generally, the pine marten is considered a habitat specialist that primarily associates with forests and avoids the vicinity of humans (Clevenger 1994; Caryl et al. 2012; Lombardini et al. 2015; Wereszczuk and Zalewski 2015; but see Balestrieri et al. 2010), whereas the stone marten is more closely associated with agricultural, industrial and urban areas, with its occurrence less affected by human presence (Virgós et al. 2000; Rondinini and Boitani 2002). However, the stone marten has regional intraspecific differences in habitat choice, being less associated with human settlements in Southern than in Central Europe (Sacchi and Meriggi 1995). The pine marten density may decline, but the stone marten may accelerate expiation due to habitat loss, mainly deforestation (Caryl et al. 2012, Wereszczuk et al. 2017). Thus, both species’ current distribution may have been shaped by habitat preferences and anthropogenic factors in different ways (Proulx et al. 2004).
Our goal was to analyse the habitat preferences of these two sympatric mesocarnivores that potentially compete for various resources (e.g. food or shelter), using datasets collected in N2K areas. To do this, we compared data on the presence of both species in N2K areas with habitat type, climatic conditions and landscape features. It was hypothesised that the pine marten would prefer areas with higher forest cover, lower temperature and more steep terrain than the stone marten and, in contrast, that the occurrence of the stone marten would be more affected by climatic conditions (e.g. temperature).
Material and methods
Data sources
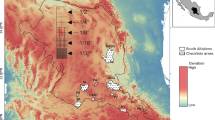
Analyses were based on N2K data—the European network of protected areas, accessible from the web page of the European Environment Agency (EEA). Descriptive data are available as MS Access and spatial data (borders of sites) as ESRI shape files (http://www.eea.europa.eu/data-and-maps/data/natura-2000). We searched databases for information about the presence/absence of both marten species in N2K sites of Southern Europe (the Iberian, Apennine and Balkan peninsulas), which included the following countries: Spain, Portugal, Italy, Greece, Bulgaria and Romania (Fig. 1). The analyses were restricted to these regions due to data availability, because for most of the remaining European countries, data on these species were not available or very scarce. We used SDFs published on the 12th of July 2013; thus, the marten occurrence data were collected before this date.

Occurrence of European pine marten Martes martes and stone marten Martes foina in NATURA 2000 sites in Southern Europe
For all analysed sites, the centroids were determined (defined as the centre of gravity of an object) and georeferenced in the grid standard of the European Terrestrial Reference System Lambert Azimuthal Equal-Area (ETRS LAEA) projected coordinate reference system. We used the Shuttle Radar Topography Mission (SRTM) Digital Terrain Model (90-m resolution) to attribute environmental and climatic variables to the centroids (http://srtm.csi.cgiar.org/). Meteorological data (for the period 2000–2009) were downloaded from http://spirits.jrc.ec.europa.eu/. The analyses were performed in ArcGIS for Desktop Advanced 10.1 and Quantum GIS 2.0.1.
Statistical methods
Analyses were based on the database of marten presence and environmental and climate variables for 4440 N2K sites: forest cover (%), slope (%) and elevation (m above sea level), mean (for the period 2000–2009) air temperature (°C) and yearly (for the period 2000–2009) mean rainfall (mm). Initial analyses revealed some degree of correlation among factors; thus, only two uncorrelated climatic variables (mean ambient temperature and precipitation) and two uncorrelated topographical variables (slope and elevation) were selected to test the influence of climate and landform features on marten presence. Slope was chosen as a proxy for human disturbance, as steeper terrains are usually less urbanised and visited by humans; forest cover has been reported as a major variable affecting the occurrence of both martens in Southern Europe (Virgos et al. 2012). We determined the effect of these variables on the occurrence of both marten species using generalised linear mixed models, with multivariate normal random effects and a logit link function for the binomial distribution. The response variable was coded 1 and 0, corresponding to the presence or absence of either the pine or stone marten. The area (km2) of each N2K site was used as an additional variable. Prior to analysis, continuous explanatory variables were log transformed, whereas variables expressed as percentages were arcsin transformed. In all models, the region (the Balkans, Iberia, Italy) was used as a random variable. Initially, we distinguished among deciduous, coniferous and mixed forests, but preliminary analyses did not find any significant effect of these variables on the occurrence of either marten species.
To examine patterns of spatial autocorrelation of model residuals, spatial correlograms were used. We plotted Moran’s I autocorrelation coefficient calculated for pairs of spatial observations against distance (5 km increment) using the ncf package (R Development Core Team 2013). Rousset and Ferdy (2014) suggested accounting for this effect when inferring fixed effects, even if autocorrelation appears non-significant. Generalised linear mixed models (GLMMs) were applied with spatially autocorrelated random effects and a variant of penalised quasi-likelihood approximations (PQL/L) for the estimation of fixed effects, as recommended for binary data (Rousset and Ferdy 2014). The Matérn autocorrelation function was used to fit the random effect of spatial distances. Initial values for correlation parameters (rho and nu are parameters of the Matérn family) were provided based on the model with the least data points (a buffer of 10 km radius; details provided in the next paragraph), whereas dispersion parameters were inferred from the current model. The significance of fixed effects was estimated using a log-likelihood ratio test. The most common procedure used to deal with spatial autocorrelation is based on penalised quasi-likelihood approximations (glmmPQL), but the assumptions underlying the autologistic regression used in this algorithm have been criticised recently (Betts et al., 2009; Rousset and Ferdy, 2014). Our preliminary analyses (results not reported) demonstrated that although penalised quasi-likelihood approximations produced similar results, the method suggested by Rousset and Ferdy (2014) was more conservative.
Analyses were based on presence/absence data and performed at two spatial scales—local and regional. At both scales, we compared N2K sites occupied by martens with random sites, where the animals were absent. The entire dataset contained over 4000 records, but only about 800 sites were occupied by martens. In order to balance the number of sites where martens were present with those where animals were absent, we applied different methods to select unoccupied areas. At the local scale, around the N2K sites where martens were present (occupied sites), we set buffers of radii 10, 15 and 20 km. These buffers encompassed various numbers of N2K sites, where the presence of martens was not confirmed by surveys (unoccupied sites). The number of unoccupied sites selected by the 10-km buffers was approximately equal to the number of occupied sites, but two times higher for the 15-km buffers and three times higher for the 20-km buffers. This means that on average there was one unoccupied site at a distance of 10 km from an occupied site, two at a range of 20 km and three at a range of 30 km. The range of the radii was within the dispersal distances of both species; thus, all unoccupied sites within the buffers could have potentially been settled by martens (Broquet et al. 2006).
At the regional scale, within the three regions (Iberian, Apennine and Balkan peninsulas), N2K sites unoccupied by martens were selected randomly. This was done by determining different numbers of unoccupied areas from all available sites within a given region. Analogous to analyses applied at local scales, the numbers of unoccupied sites were either equal to the number of occupied sites or two or three times higher than the number of occupied sites.
All analyses were done separately for the pine and stone marten. There were only a small number of sites simultaneously occupied by both species, and the initial model failed to converge using this dataset; thus, we excluded these sites from further analyses. For the spatial analyses, we used Quantum GIS ver. 1.7.4–Wrocław (Szczepanek 2013). All statistical analyses were performed using R software (R Development Core Team 2013), package spaMM (Rousset and Ferdy 2014).
Results
Residuals of all examined models were spatially autocorrelated. For the pine marten, Moran’s autocorrelation coefficients (Moran’s I) were significant and positive for distances up to 120 km (mean Moran’s I = 0.179, mean p = 0.014), and negative between 250 and 350 km (mean Moran’s I = − 0.083, mean p = 0.018), whereas above 350 km, autocorrelations were not significant. Similarly, for the stone marten, autocorrelation coefficients were significant and positive for distances up to 120 km (mean Moran’s I = 0.200, mean p = 0.003) and again between 320 and 480 km (mean Moran’s I = 0.068, mean p = 0.003), whereas between 200 and 300 km, they were negative (mean Moran’s I = − 0.044, mean p = 0.028). Generally, autocorrelation coefficients of model residuals were higher in the models constructed for the stone marten data, including the range above 500 km, most probably due to the higher number of records within each distance range. The climatic variables (mean ambient temperature and precipitation) were more spatially correlated than the environmental variables (proportion of area covered by forest, slope and elevation).
In all analyses, the area of N2K sites occupied by pine martens was significantly (p < 0.001) larger (on average two to three times) than the unoccupied sites (Table 1). In relation to all buffer sizes and random sites, these differences were highly significant (p < 0.001). At the local scale, the occurrence of the pine marten was positively affected by percentage forest cover (on average, 58.12 vs. 43.54%) and slope (on average, 19.97 vs. 17.04°), but negatively associated with elevation (on average, 660.15 vs. 706.01 m a.s.l.) (Table 2). Mean temperature and precipitation had no significant effect on the occurrence of this species. At the regional scale, we observed similar effects of forest cover and slope on pine marten occurrence, but the effect of elevation was opposite, i.e. pine martens more frequently occupied sites located at higher elevations than random sites (Table 1). The mean elevation of the occupied sites was 160 to 200 m higher than that of the unoccupied sites within the regions, but this effect was not significant (Table 3). Also, the effect of precipitation was significant at the regional scale (Table 3).
Regardless of the scale at which analyses were carried out, the sites occupied by the stone marten were significantly larger than the unoccupied ones (Tables 4 and 5). At the local scale, stone marten occurrence was significantly affected by the proportion of forest cover (except for the 10-km buffer) and slope (Table 4). At the regional scale, stone martens also preferred sites with a higher proportion of forest cover, but also those located at higher elevations and on less steep slopes, when compared with random sites (Table 5).
At the regional scale (in the 10-km buffer), sites occupied by stone martens were significantly smaller, less forested and less sloped than those occupied by pine martens (Table 6). Climate conditions at the sites occupied by these species were not significantly different (Table 6).
Across all buffers and for randomly selected sites, the proportion of forest cover, after adjusting for the effect of spatial autocorrelation, was positively affected by mean ambient temperature, mean precipitation, slope and elevation. The magnitude of this effect varied according to the spatial scale (local vs. regional) and species (sites associated with the pine or stone marten), but in all cases was significant. Sites with higher mean temperatures and precipitation as well as higher elevations and on more steep slopes tended to have higher forest cover.
Discussion
Our results largely confirmed the findings of previous studies on habitat selection of the pine and stone marten in Southern Europe, mainly in the Mediterranean region (Clevenger 1994; Sacchi and Meriggi 1995; Virgós et al. 2000; Balestrieri et al. 2010; Lombardini et al. 2015). The main weakness of this approach was the quality of the data, which may differ among regions and years, because it was based both on direct observations of focal species and literature sources. Similarly, data-quality issues need to be considered in citizen science, which is becoming a widely accepted scientific method (Dickinson et al. 2010). Despite some disadvantages, both approaches represent valuable tools for collecting large-scale data (Kallimanis et al. 2017).
The results of habitat preferences for both marten species were relatively consistent across different scales, but with increasing buffer size and number of random sites, there was a tendency for variables unimportant at local scales to increase in significance. In particular, slope and elevation were not different for locations near sites occupied by martens, but their difference became significant when more sites were included in the comparison. The tendency of martens to occur in more steep and hilly locations has also been previously demonstrated by Virgós et al. (2000).
According to Clevenger (1994), pine martens do not exhibit a marked preference for forested over non-forested habitats in the absence of competitors/predators. Prigioni et al. (2008) also pointed to interspecific competition as a significant factor affecting habitat selection by martens. Therefore, the preferences of these two species for more forested and steep locations can be explained by the avoidance of either competitors/predators or a preference for areas less intensively managed or populated. The former explanation seems to be more plausible for the stone marten, considering the common occurrence of this species in urban areas, where they avoid competitors, such as foxes or badgers (Prigioni et al. 2008). The preference of stone martens for higher elevations indicates that the distribution of this species in Southern Europe is not limited by climatic factors. On the other hand, this species can exploit areas with lower forest cover, characterised by a higher proportion of crevices and other types of shelter (Virgós et al. 2002; but see Sacchi and Meriggi 1995). This type of habitat is less suitable for the pine marten, which prefers slightly lower elevations, with higher proportions of forest; however, pine martens also inhabit areas located at higher altitude, when compared with random locations. A similar pattern has also been observed among pine martens from Sardinia, Italy (Lombardini et al. 2015).
N2K sites occupied by either species of marten were two to three times larger than unoccupied ones. Despite the stone marten being recognised as a habitat generalist, its presence was still affected by site size. Similar effects of a patch size on this species have been demonstrated by Virgós and García (2002). In contrast, the pine marten is considered a forest specialist (Clevenger 1994; Caryl et al. 2012; Lombardini et al. 2015); thus, it occupied N2K sites of even larger area. Also, it is likely that larger N2K sites are less disturbed by people; thus, they offer better conditions for both species of martens. In the study, we included only areas in the N2K network that are more or less natural ecosystems, and therefore, intensely urbanised and human-modified habitats were not analysed. Thus, we did not show the stone marten’s strong preferences for urban areas, as these have been found in other studies (Santos and Santos-Reis 2010; Vergara et al. 2016).
It cannot be excluded that similar preferences of the stone and pine marten for steeper and higher elevation locations, characterised by higher forest cover, are caused by different factors. In this way, both species may avoid competitors and predators, but whereas pine martens stay away from urban areas and arable land (Lombardini et al. 2015), stone martens indicate some preferences for these types of habitats (Prigioni et al. 2008; Vergara et al. 2016). The study of Rondinini and Boitani (2002), based on observations of radio-collared stone martens in the Apennines (Italy), indicated a higher proportion of woodlands and a lower proportion of arable land within their home ranges, compared with random locations. Whereas other studies in Southern Europe have shown that stone martens avoid forests and prefer areas with higher proportions of cultivated land (Sacchi and Meriggi 1995; Vergara et al. 2016), other studies have indicated the importance of mosaic habitats (forest, agriculture land and rural areas) for the stone marten (Virgós et al. 2000; Rondinini and Boitani 2002; Santos and Santos-Reis 2010). These discrepancies may result either from local differences in habitat preferences and availability of suitable environments or from the methods used for estimating the presence/absence of study species.
This comparison of site selection between the martens has provided additional insight into their habitat niche separation. N2K sites occupied only by pine martens were characterised by larger area, higher forest cover and more steep locations when compared with sites where only the stone marten occurred. The differences between species in habitat selection may reduce competition between them, but may also reflect differences in their behaviour and morphological adaptation. The pine marten is a more arboreal species than the stone marten, often climbing trees, moving in the tree canopy and using resting sites in the crown of trees (Zalewski 1997, Goszczyński et al. 2007). In general, pine martens have larger home ranges than stone martens (Zalewski et al. 2006; Herr 2008), and this may explain why the species occupies N2K sites with larger areas.
The results of this study generally agree with previous findings, and the additional new findings concerning habitat selection by both martens demonstrate the usefulness of freely available NATURA 2000 site data for large-scale analyses. In addition, it offers some novel information about habitat-species interactions in protected areas, and it is potentially suitable for other types of analyses. This type of data may be widely used, for example, in conservation planning and assessing the impact of climate change on species’ ranges.
References
Balestrieri A, Remonti L, Ruiz-González A, Gómez-Moliner BJ, Vergara M, Prigioni C (2010) Range expansion of the pine marten (Martes martes) in an agricultural landscape matrix (NW Italy). Mamm Biol 75:412–419
Balestrieri A, Bogliani G, Boano G, Ruiz-González A, Saino N, Costa S, Milanesi P (2016) Modelling the distribution of forest-dependent species in human-dominated landscapes: patterns for the pine marten in intensively cultivated lowlands. PLoS One 11:e0158203
Betts MG, Ganio LM, Huso MMP, Som NA, Huettmann F, Bowman J, Wintle BA (2009) Comment on “Methods to account for spatial autocorrelation in the analysis of species distributional data: a review”. Ecography 32:374–378
Bissonette JA (1997) Scale-sensitive ecological properties: historical context current meaning. In: Bissonette JA (ed) Wildlife and landscape ecology effects of pattern and scale. Springer New York, pp 3-31
Boyce MS, Mao JS, Merrill EH, Fortin D, Turner MG, Fryxell J, Turchin P (2003) Scale and heterogeneity in habitat selection by elk in Yellowstone National Park. Ecoscience 10:321–332
Broquet T, Johnson CA, Petit E, Thompson I, Burel F, Fryxell J (2006) Dispersal and genetic structure in the American marten Martes americana. Mol Ecol 15:1689–1697
Caryl FM, Quine CP, Park KJ (2012) Martens in the matrix: the importance of nonforested habitats for forest carnivores in fragmented landscapes. J Mamm 93:464–474
Clevenger AP (1994) Habitat characteristics of Eurasian pine martens Martes martes in an insular Mediterranean environment. Ecography 17:257–263
Debinski DM, Holt RD (2000) A survey and overview of habitat fragmentation experiments. Conserv Biol 14:342–355
Dickinson JL, Zuckerberg B, Bonter DN (2010) Citizen science as an ecological research tool: challenges and benefits. Annu Rev Ecol Evol S 41:149–172
Goszczyński J, Posłuszny M, Pilot M, Gralak B (2007) Patterns of winter locomotion and foraging in two sympatric marten species: Martes martes and Martes foina. Can J Zool 85:239–249
Herr J (2008) Ecology and behaviour of urban stone marten (Martes foina) in Luxembourg. PhD School of Life Sciences, University of Sussex, Sussex, pp. 1–144
Kallimanis AS, Panitsa M, Dimopoulos P (2017) Quality of non-expert citizen science data collected for habitat type conservation status assessment in Natura 2000 protected areas. Sci Rep 7:8873. https://doi.org/10.1038/s41598-017-09316-9
Law BS, Dickman CR (1998) The use of habitat mosaics by terrestrial vertebrate fauna: implications for conservation and management. Biodivers Conserv 7:323–333
Lombardini M, Cinerari CE, Murru M, Rosin AV, Mazzoleni L, Meriggi A (2015) Habitat requirements of Eurasian pine marten Martes martes in a Mediterranean environment. Mamm Res 60:97–105
Mayor SJ, Schneider DC, Schaefer JA, Mahoney SP (2009) Habitat selection at multiple scales. Ecoscience 16:238–247
Mead RA (1994) Reproduction in Martes. In: Buskirk SW, Harestad AS, Raphael MG, Powell RA (eds) Martens, sables and fishers: biology and conservation. Cornell University Press, Ithaca and London, pp 404–422
Monterroso P, Rebelo P, Alves PC, Ferreras P (2016) Niche partitioning at the edge of the range: a multidimensional analysis with sympatric martens. J Mamm 97:928–939
O’Mahony DT (2017) A species distribution model for pine marten (Martes martes) in the least forested region of Europe. Mamm Res 62:195–200
Prigioni C, Balestrieri A, Remonti L, Cavada L (2008) Differential use of food and habitat by sympatric carnivores in the eastern Italian Alps. Ital J Zool 75:173–184
Proulx G, Aubry K, Birks J, Buskirk S, Fortin C, Frost H, Krohn W, Mayo L, Monakhov V, Payer D, Saeki M, Santos-Reis M, Weir R, Zielinski W (2004) World distribution and status of the genus Martes in 2000. In: Harrison DJ, Fuller AK, Proulx G (eds) Martens and fishers (Martes) in human-altered environments: an international perspective. Springer, New York, pp 21–76
R Development Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria http://wwwR-projectorg/ (accessed 18 October 2016)
Rondinini C, Boitani L (2002) Habitat use by beech martens in a fragmented landscape. Ecography 25:257–264
Rousset F, Ferdy JB (2014) Testing environmental and genetic effects in the presence of spatial autocorrelation. Ecography 37:781–790
Sacchi O, Meriggi A (1995) Habitat requirements of the stone marten (Martes foina) on the Tyrrhenian slopes of the northern Apennines. Hystrix 7:99–104
Santos MJ, Santos-Reis M (2010) Stone marten (Martes foina) habitat in a Mediterranean ecosystem: effects of scale sex and interspecific interactions. Eur J Wildlife Res 56:275–286
Sato JJ, Wolsan M, Prevosti FJ, D’Elíad G, Begge C, Begge K, Hosodaf T, Campbellg KL, Suzuki H (2012) Evolutionary and biogeographic history of weasel-like carnivorans (Musteloidea). Mol Phyl Evol 63:745–757
Szczepanek T (2013) Systemy informacji przestrzennej z Quantum GIS Część I. Politechnika Krakowska, Kraków [in Polish]
Vergara M, Cushman SA, Urra F, Ruiz-Gonzalez A (2016) Shaken but not stirred: multiscale habitat suitability modeling of sympatric marten species (Martes martes and Martes foina) in the northern Iberian Peninsula. Landsc Ecol 31:1241–1260
Virgós E, Recio MR, Cortés Y (2000) Stone marten (Martes foina) use of different landscapes types in the mountains of central Spain. Mamm Biol 65:375–379
Virgós E, Tellería JL, Santos T (2002) A comparison on the response to forest fragmentation by medium-sized Iberian carnivores in central Spain. Biodivers Conserv 11:1063–1079
Virgós E, García FJ (2002) Patch occupancy by stone martens Martes foina in fragmented landscapes of central Spain: the role of fragment size isolation and habitat structure. Acta Oecol 23:231–237
Virgós E, Zalewski A, Rosalino LM, Mergey M (2012) Habitat ecology of Martes species in Europe: a review of the evidence. In: Aubry KB, Zielinski WJ, Raphael MG, Proulx G, Buskirk SW (eds) Biology and conservation of martens, sables, and fishers. A new synthesis. Cornell University Press, Ithaca and London, pp 255–266
Wereszczuk A, Zalewski A (2015) Spatial niche segregation of sympatric stone marten and pine marten—avoidance of competition or selection of optimal habitat? PLoS One 10:e0139852
Wereszczuk A, Leblois R, Zalewski A (2017) Genetic diversity and structure related to expansion history and habitat isolation: stone marten populating rural-urban habitats. BMC Ecology (in print)
Zalewski A (1997) Patterns of resting site use by pine marten Martes martes in Białowieża National Park (Poland). Acta Theriol 42:153–168
Zalewski A, Jędrzejewski W (2006) Spatial organisation and dynamics of pine marten Martes martes population in Białowieża Forest (E Poland) compared with other European woodlands. Ecography 29:31–43.
Acknowledgements
We thank Edyta Kozieł for inspiration and improving the English. We are grateful to Tomasz Diserens (www.englishpad.pl) for correcting the English.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical approval
This article does not contain any studies with animals performed by any of the authors.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Zub, K., Kozieł, M., Siłuch, M. et al. The NATURA 2000 database as a tool in the analysis of habitat selection at large scales: factors affecting the occurrence of pine and stone martens in Southern Europe. Eur J Wildl Res 64, 10 (2018). https://doi.org/10.1007/s10344-018-1168-z
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-018-1168-z