
Abstract
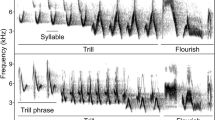
Among the broad diversity of songbird vocalisations, song can serve a wide range of different functions depending on the species and context. In many species, aggressive motivation has often been linked with the use of fast repeated series of elements typically referred to as trills. However, only a few studies explored the role of this specific component in species with a large repertoire and high song complexity. Here, we investigate the potential role of trills in the territorial vocal response of males Thrush Nightingale (Luscinia luscinia), a species with complex songs characterised by the frequent use and diversity of their trills. We performed playback experiments simulating territorial intrusion to test if trills signal aggressive motivation in this species. If so, we expected tested males to respond by changing their trill rate or frequency of trill use in songs, and/or using different trill types than before the stimulation. Contrary to our expectation, males did not modify their trill rate or differed in trill type use before, during or after playback. There was a tendency for decrease in trill duration and the number of elements in the trills during the playback stimulation, reflecting to some extent the overall use of shorter songs by males during that stage of the experiment. Altogether, our study does not support the role of trills as motivation signals for territory defence in Thrush Nightingales, but advocates for more research on their potential roles in signalling other information.
Zusammenfassung
Beim Sprosser, einem Singvogel mit komplexem Gesang, spielen Triller wohl keine Rolle bei der Antwort von Männchen auf ein Eindringen ins Revier
Als Teil der vielfältigen Lautäußerungen von Singvögeln kann Gesang eine Reihe unterschiedlicher Funktionen erfüllen, abhängig von der jeweiligen Vogelart und Situation. Bei vielen Arten ist die Motivation zum Angriff oftmals mit der Benutzung schneller, wiederholter Serien von typischerweise als Triller bezeichneten Elementen in Verbindung gebracht worden. Nur wenige Studien haben jedoch die Rolle dieser spezifischen Gesangskomponente bei Arten mit einem breiten Gesangsrepertoire und hoher Gesangskomplexität erforscht. Hier untersuchen wir die potenzielle Rolle von Trillern bei der Territorialantwort männlicher Sprosser (Luscinia luscinia), einer Art mit komplexen Gesängen, die durch den häufigen Gebrauch vielfältiger Triller gekennzeichnet sind. Wir haben Playback-Experimente durchgeführt, die das Eindringen eines Rivalen ins Revier simulierten, um zu testen, ob Triller bei dieser Vogelart die Angriffsmotivation signalisieren. In diesem Fall erwarteten wir, dass die getesteten Männchen ihre Trillerrate oder die Häufigkeit des Trillergebrauchs in ihren Gesängen ändern und/oder andere Trillertypen als vor der Simulation benutzen sollten. Entgegen unserer Erwartung änderten die Männchen jedoch weder ihre Trillerrate, noch unterschieden sich die Trillertypen, die sie vor, während und nach dem Playback benutzten. Es bestand eine Tendenz, die Trillerdauer sowie die Anzahl verschiedener Elemente in den Trillern während der Playback-Simulation zu verringern, was zu einem gewissen Grad den Gebrauch kürzerer Gesänge durch die Männchen in dieser Phase des Experiments widerspiegelt. Insgesamt stützt unsere Studie die Rolle von Trillern als Motivationssignale für die Revierverteidigung bei Sprossern nicht, sondern befürwortet die weitere Untersuchung ihrer Rolle beim Übermitteln anderer Informationen.






Similar content being viewed by others

Availability of data and materials
The datasets generated and analysed during the current study will be available online (Dryad data repository). All experimental recordings used in the analysis have been deposited in the Animal Sound Archive at the Museum for Natural History in Berlin and are available under the CC-BY-SA Licence at the following https://doi.org/10.7479/b478-jj11
Code availability
The custom code (R script) used for the statistical analysis is available from the corresponding author on request.
Change history
30 April 2021
A Correction to this paper has been published: https://doi.org/10.1007/s10336-021-01889-7
References
Akçay Ç, Arnold JA, Hambury KL, Dickinson JL (2016) Age-based discrimination of rival males in western bluebirds. Anim Cogn 19:999–1006. https://doi.org/10.1007/s10071-016-1004-3
Baker TM, Wilson DR, Mennill DJ (2012) Vocal signals predict attack during aggressive interactions in black-capped chickadees. Anim Behav 84:965–974. https://doi.org/10.1016/j.anbehav.2012.07.022
Ballentine B (2009) The ability to perform physically challenging songs predicts age and size in male swamp sparrows, Melospiza georgiana. Anim Behav 77:973–978. https://doi.org/10.1016/j.anbehav.2008.12.027
Ballentine B, Hyman J, Nowicki S (2004) Vocal performance influences female response to male bird song: an experimental test. Behav Ecol 15:163–168. https://doi.org/10.1093/beheco/arg090
Balsby TJS, Hansen P (2010) Element repertoire: change and development with age in whitethroat Sylvia communis song. J Ornithol 151:469–476. https://doi.org/10.1007/s10336-009-0481-4
Bartsch C, Wenchel R, Kaiser A, Kipper S (2014) Singing onstage: female and male common nightingales eavesdrop on song type matching. Behav Ecol Sociobiol 68:1163–1171. https://doi.org/10.1007/s00265-014-1727-6
Botero CA, Rossman RJ, Caro LM, Stenzler LM, Lovette IJ, de Kort SR, Vehrencamp SL (2009) Syllable type consistency is related to age, social status and reproductive success in the tropical mockingbird. Anim Behav 77:701–706. https://doi.org/10.1016/j.anbehav.2008.11.020
Briefer E, Aubin T, Lehongre K, Rybak F (2008) How to identify dear enemies: the group signature in the complex song of the skylark Alauda arvensis. J Exp Biol 211:317–326. https://doi.org/10.1242/jeb.013359
Byers BE (2006) Extrapair paternity in chestnut-sided warblers is correlated with consistent vocal performance. Behav Ecol 18:130–136. https://doi.org/10.1093/beheco/arl058
Byers BE, Kroodsma DE (2009) Female mate choice and songbird song repertoires. Anim Behav 77:13–22. https://doi.org/10.1016/j.anbehav.2008.10.003
Caro SP, Sewall KB, Salvante KG, Sockman KW (2010) Female Lincoln’s sparrows modulate their behavior in response to variation in male song quality. Behav Ecol 21:562–569. https://doi.org/10.1093/beheco/arq022
Catchpole CK, Slater PJB (2008) Bird song. Biological themes and variations. Cambridge University Press, Cambridge
Cramer ERA (2012) Vocal deviation and trill consistency do not affect male response to playback in house wrens. Behav Ecol 24:412–420. https://doi.org/10.1093/beheco/ars178
Cramer ERA (2013) Physically challenging song traits, male quality, and reproductive success in house wrens. PLoS ONE 8:e59208. https://doi.org/10.1371/journal.pone.0059208
Cramp S, Perrins CM (1994) Birds of the Western Palearctic. Oxford University Press, Oxford
De Kort SR, Eldermire ER, Valderrama S, Botero CA, Vehrencamp SL (2009) Trill consistency is an age-related assessment signal in banded wrens. Proc R Soc B 276:2315–2321. https://doi.org/10.1098/rspb.2009.0127
DuBois AL, Nowicki S, Searcy WA (2009) Swamp sparrows modulate vocal performance in an aggressive context. Biol Lett 5:163–165. https://doi.org/10.1098/rsbl.2008.0626
DuBois AL, Nowicki S, Searcy WA (2011) Discrimination of vocal performance by male swamp sparrows. Behav Ecol Sociobiol 65:717–726. https://doi.org/10.1007/s00265-010-1073-2
Ferreira AC, Atwell JW, Whittaker DJ, Ketterson ED, Cardoso GC (2016) Communication value of mistakes in dark-eyed junco song. Am Nat 188:289–305. https://doi.org/10.1086/687520
Funghi C, Cardoso GC, Mota PG (2014) Increased syllable rate during aggressive singing in a bird with complex and fast song. J Avian Biol 46:283–288. https://doi.org/10.1111/jav.00480
Griessmann B, Naguib M (2002) Song sharing in neighboring and non-neighboring thrush nightingales (Luscinia luscinia) and its implications for communication. Ethology 108:377–387. https://doi.org/10.1046/j.1439-0310.2002.00781.x
Hothorn T, Bretz F, Westfall P (2008) Simultaneous inference in general parametric models. Biom J 50:346–363. https://doi.org/10.1002/bimj.200810425
Hultsch H, Todt D (1981) Repertoire sharing and song-post distance in nightingales (Luscinia megarhynchos B.). Behav Ecol Sociobiol 8:183–188. https://doi.org/10.1007/bf00299828
Illes AE, Hall ML, Vehrencamp SL (2006) Vocal performance influences male receiver response in the banded wren. Proc R Soc B 273:1907–1912. https://doi.org/10.1098/rspb.2006.3535
Ivanitskii VV, Marova IM, Antipov VA (2016) Sequential organization in the song of thrush nightingale (Luscinia luscinia): clustering and sequential order of the song types. Bioacoustics 26:199–215. https://doi.org/10.1080/09524622.2016.1239132
Jakubowska A, Osiejuk TS (2018) Soft songs in male ortolan buntings are used in an aggressive context but are not an aggressive signal. Ethology 124:549–558. https://doi.org/10.1111/eth.12758
Kipper S, Mundry R, Hultsch H, Todt D (2004) Long-term persistence of song performance rules in nightingales (Luscinia megarhynchos): a longitudinal field study on repertoire size and composition. Behaviour 141:371–390. https://doi.org/10.1163/156853904322981914
Kipper S, Mundry R, Sommer C, Hultsch H, Todt D (2006) Song repertoire size is correlated with body measures and arrival date in common nightingales, Luscinia megarhynchos. Anim Behav 71:211–217. https://doi.org/10.1016/j.anbehav.2005.04.011
Kunc HP, Amrhein V, Naguib M (2005) Seasonal variation in dawn song characteristics in the common nightingale. Anim Behav 70:1265–1271. https://doi.org/10.1016/j.anbehav.2005.02.010
Kunc HP, Amrhein V, Naguib M (2006) Vocal interactions in nightingales, Luscinia megarhynchos: more aggressive males have higher pairing success. Anim Behav 72:25–30. https://doi.org/10.1016/j.anbehav.2005.08.014
Lind H, Dabelsteen T, McGregor PK (1996) Female great tits can identify mates by song. Anim Behav 52:667–671. https://doi.org/10.1006/anbe.1996.0211
Linhart P, Jaška P, Petrusková T, Petrusek A, Fuchs R (2013) Being angry, singing fast? Signalling of aggressive motivation by syllable rate in a songbird with slow song. Behav Processes 100:139–145. https://doi.org/10.1016/j.beproc.2013.06.012
Liu IA, Soha JA, Nowicki S (2018) Song type matching and vocal performance in territorial signalling by male swamp sparrows. Anim Behav 139:117–125. https://doi.org/10.1016/j.anbehav.2018.03.007
Marova IM, Ivlyeva AL, Veprintzeva OD, Ivanitskii VV (2015a) A comparative analysis of song differentiation in thrush nightingale (Luscinia luscinia) and common nightingale (Luscinia megarhynchos) within their ranges. Zool Zh 94:701–710. https://doi.org/10.7868/S0044513415060124
Marova IM, Antipov VA, Ivanitskii VV (2015b) Anomalous weather events and horizontal transfer of the thrush nightingale (Luscinia luscinia) vocal models. Dokl Biol Sci 463:186–189. https://doi.org/10.1134/s001249661504002x
Naguib M, Kipper S (2006) Effects of different levels of song overlapping on singing behaviour in male territorial nightingales (Luscinia megarhynchos). Behav Ecol Sociobiol 59:419–426. https://doi.org/10.1007/s00265-005-0066-z
Naguib M, Mundry R, Hultsch H, Todt D (2002) Responses to playback of whistle songs and normal songs in male nightingales: effects of song category, whistle pitch, and distance. Behav Ecol Sociobiol 52:216–223. https://doi.org/10.1007/s00265-002-0511-1
Naguib M, Schmidt R, Sprau P, Roth T, Flörcke C, Amrhein V (2008) The ecology of vocal signaling: male spacing and communication distance of different song traits in nightingales. Behav Ecol 19:1034–1040. https://doi.org/10.1093/beheco/arn065
Oppel JJ (1869) Ueber den Goldammergesang Zool Garten 10:280–286
Payne RB (1982) Ecological consequences of song matching: Breeding success and intraspecific song mimicry in indigo buntings. Ecology 63:401–411. https://doi.org/10.2307/1938958
Petrusková T, Kinštová A, Pišvejcová I, Mula Laguna J, Cortezón A, Brinke T, Petrusek A (2014) Variation in trill characteristics in tree pipit songs: different trills for different use? Ethology 120:586–597. https://doi.org/10.1111/eth.12231
Pinheiro J, Bates D, DebRoy S, Sarkar D, R Core Team (2019) nlme: linear and nonlinear mixed effects models. R package version 3.1-140. https://CRAN.R-project.org/package=nlme
Podos J (1997) A performance constraint on the evolution of trilled vocalizations in a songbird family (Passeriformes: Emberizidae). Evolution 51:537–551. https://doi.org/10.1111/j.1558-5646.1997.tb02441.x
Podos J, Warren PS (2007) The evolution of geographic variation in bird songs. Adv Stud Behav 37:403–458. https://doi.org/10.1016/S0065-3454(07)37009-5
R Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. http://www.R-project.org
Reif J, Jiran M, Reifová R, Vokurková J, Dolata PT, Petrusek A, Petrusková T (2015) Interspecific territoriality in two songbird species: potential role of song convergence in male aggressive interactions. Anim Behav 104:131–136. https://doi.org/10.1016/j.anbehav.2015.03.016
Rivera-Gutierrez HF, Pinxten R, Eens M (2010) Multiple signals for multiple messages: great tit, Parus major, song signals age and survival. Anim Behav 80:451–459. https://doi.org/10.1016/j.anbehav.2010.06.002
Rivera-Gutierrez HF, Pinxten R, Eens M (2012) Tuning and fading voices in songbirds: age-dependent changes in two acoustic traits across the life span. Anim Behav 83:1279–1283. https://doi.org/10.1016/j.anbehav.2012.03.001
Schmidt R, Kunc HP, Amrhein V, Naguib M (2008) Aggressive responses to broadband trills are related to subsequent pairing success in nightingales. Behav Ecol 19:635–641. https://doi.org/10.1093/beheco/arn021
Searcy WA, Beecher MD (2009) Song as an aggressive signal in songbirds. Anim Behav 78:1281–1292. https://doi.org/10.1016/j.anbehav.2009.08.011
Searcy WA, Anderson RC, Nowicki S (2006) Bird song as a signal of aggressive intent. Behav Ecol Sociobiol 60:234–241. https://doi.org/10.1007/s00265-006-0161-9
Skierczyński M, Czarnecka KM, Osiejuk TS (2007) Neighbour-stranger song discrimination in territorial ortolan bunting Emberiza hortulana males. J Avian Biol 38:415–420. https://doi.org/10.1111/j.0908-8857.2007.04123.x
Sorjonen J (1983) Transmission of the two most characteristic phrases of the song of the thrush nightingale Luscinia luscinia in different environmental conditions. Ornis Scand 14:278–288. https://doi.org/10.2307/3676320
Sorjonen J (1986) Mixed singing and interspecific territoriality – consequences of secondary contact of two ecologically and morphologically similar nightingale species in Europe. Ornis Scand 17:53–67. https://doi.org/10.2307/3676753
Sorjonen J (1987) Temporal and spatial differences in traditions and repertoires in the song of the Thrush Nightingale (Luscinia luscinia). Behaviour 102:196–211. https://doi.org/10.1163/156853986x00126
Souriau A, Kohoutová H, Reif J, Vokurková J, Petrusek A, Reifová R, Petrusková T (2018) Can mixed singing facilitate coexistence of closely related nightingale species? Behav Ecol 29:925–932. https://doi.org/10.1093/beheco/ary053
Sprau P, Roth T, Schmidt R, Amrhein V, Naguib M (2010a) Communication across territory boundaries: distance-dependent responses in nightingales. Behav Ecol 21:1011–1017. https://doi.org/10.1093/beheco/arq097
Sprau P, Schmidt R, Roth T, Amrhein V, Naguib M (2010b) Effects of rapid broadband trills on responses to song overlapping in nightingales. Ethology 116:300–308. https://doi.org/10.1111/j.1439-0310.2010.01743.x
Storchová R, Reif J, Nachman MW (2010) Female heterogamety and speciation: reduced introgression of the Z chromosome between two species of nightingales. Evolution 64:456–471. https://doi.org/10.1111/j.1558-5646.2009.00841.x
Szymkowiak J, Kuczyński L (2017) Song rate as a signal of male aggressiveness during territorial contests in the wood warbler. J Avian Biol 48:275–283. https://doi.org/10.1111/jav.00969
Turčoková L, Pavel V, Chutný B, Petrusek A, Petrusková T (2011) Differential response of males of a subarctic population of Bluethroat Luscinina svecica svecica to playbacks of their own and foreign subspecies. J Ornithol 152:975–982. https://doi.org/10.1007/s10336-011-0680-7
Vehrencamp SL, Yantachka J, Hall ML, de Kort SR (2013) Trill performance components vary with age, season, and motivation in the banded wren. Behav Ecol Sociobiol 67:409–419. https://doi.org/10.1007/s00265-012-1461-x
Vokurková J, Petrusková T, Reifová R, Kozman A, Mořkovský L, Kipper S, Weiss M, Reif J, Dolata PT, Petrusek A (2013) The causes and evolutionary consequences of mixed singing in two hybridizing songbird species (Luscinia spp.). PLoS ONE 8:e60172. https://doi.org/10.1371/journal.pone.0060172
Węgrzyn E, Leniowski K, Osiejuk TS (2010) Whistle duration and consistency reflect philopatry and harem size in great reed warblers. Anim Behav 79:1363–1372. https://doi.org/10.1016/j.anbehav.2010.03.012
Wilson DR, Mennill DJ (2010) Black-capped chickadees, Poecile atricapillus, can use individually distinctive songs to discriminate among conspecifics. Anim Behav 79:1267–1275. https://doi.org/10.1016/j.anbehav.2010.02.028
Zipple M, Nowicki S, Searcy W, Peters S (2019) Full life course analysis of bird song reveals maturation and senescence of highly repeatable song characteristics. Behav Ecol 30:1761–1768. https://doi.org/10.1093/beheco/arz146
Zipple MN, Peters S, Searcy WA, Nowicki S (2020) Sounds of senescence: male swamp sparrows respond less aggressively to the songs of older individuals. Behav Ecol 31:533–539. https://doi.org/10.1093/beheco/arz218
Acknowledgements
We thank Jana Vokurková for her contribution to the early recordings and Jiří Reif for his support in the field. We are grateful to two anonymous referees for their helpful comments and suggestions on the manuscript.
Funding
This study was supported by the Czech Science Foundation project 18-14325S.
Author information
Authors and Affiliations
Contributions
AS, TP and AP contributed to the study conception and design. AS and RR collected the song recordings necessary for the playback. AS prepared the experiment, collected and analysed the data with the input from AP and TP. AS and TP drafted the manuscript, AP and RR contributed to the revisions. All the authors gave their final approval for publication.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
Due to our non-invasive recording approach, no bird had to be captured or handled during our experiments. Playback settings were designed to minimise the territorial disturbance and the tested birds usually resumed their singing routine shortly after.
Additional information
Communicated by T. S. Osiejuk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Souriau, A., Reifová, R., Petrusek, A. et al. No evidence for a role of trills in male response to territorial intrusion in a complex singer, the Thrush Nightingale. J Ornithol 162, 865–878 (2021). https://doi.org/10.1007/s10336-021-01879-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-021-01879-9