
Abstract
Through their rooting profiles and water demands, plants affect the distribution of water in the soil profile. Simultaneously, soil water content controls plant development and interactions within and between plant communities. These plant-soil water feedbacks might vary across plant communities with different rooting depths and species composition. In semiarid environments, understanding these differences will be essential to predict how ecosystems will respond to drought, which may become more frequent and severe with climate change. In this study, we tested how plant-soil water feedbacks responded to drought in two contrasting ecosystem types—grassland and shrubland—in the coastal foothills of southern California. During years 5–8 of an ongoing precipitation manipulation experiment, we measured changes in plant communities and soil moisture up to 2 m depth. We observed different water use patterns in grassland and shrubland communities with distinct plant functional types and water use strategies. Drought treatment did not affect perennial, deep-rooted shrubs because they could access deep soil water pools. However, mid-rooted shrubs were sensitive to drought and experienced decreased productivity and die-off. As a result, water content actually increased with drought at soil depths from 50–150 cm. In grassland, biomass production by annual species, including annual grasses and forbs, declined with drought, resulting in lower water uptake from the surface soil layer. An opportunistic “live fast, die young“ life strategy allowed these species to recover quickly once water availability increased. Our results show how drought interacts with plant community composition to affect the soil water balance of semiarid ecosystems, information that could be integrated into global scale models.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Highlights
-
The study elucidates drought effects on the ecohydrology of semiarid ecosystems.
-
Drought leads to a negative plant-soil water feedback in grassland communities.
-
In shrubland communities, mid-rooted shrubs are most impacted by drought.
Introduction
Drought is a global threat (Hanson and Weltzin 2000; West and others 2019; Vicente-Serrano and others 2020) that is projected to increase in severity and frequency at global and local scales as a consequence of climate change and anthropogenic activities (Pokhrel and others 2021). Drought can impact terrestrial water storage, including soil water stocks (Marks and others 1993; Bhattacharya 2021; Wossenyeleh and others 2022). At the same time, the effects of drought on water resource availability depend on vegetation dynamics and responses to climate change (Beaulieu and others 2016) via plant-soil water feedbacks (Yang and others 2009; Asbjornsen and others 2011; Wang and others 2019), in which plant communities control water distribution in soil and, in turn, soil water content constrains plant establishment and growth. Because of the complexity of these feedbacks, drought effects on the structure and function of terrestrial ecosystems are uncertain (Ostle and others 2009; Bhattacharya 2021) even when considering vegetation in global climate models (Ostle and others 2009; Silber and others 2017).
Soil water content reflects the balance between precipitation, irrigation, infiltration, net lateral flow, deep discharge, evaporation, and transpiration (Rodriguez-Iturbe and others 1999; Rodriguez-Iturbe 2000; Rodriguez-Iturbe and others 2001). Changes in water input due to drought events directly influence soil moisture dynamics (Porporato and others 2001; Wang and others 2006; Hickel and Zhang 2006; Hajek and Knapp 2022), with significant consequences for plant physiology (Porporato and others 2001; Wang and others 2006), density, and biomass. Plants have evolved different strategies to cope with water stress (Lambers and others 2008; Malíková and others 2016; Arbona and others 2017), forming a fast-slow plant economy spectrum among species (Carvajal and others 2019) to optimize scarce water resources. According to Ryel and others (2008), plant species access water from two different soil water pools: (i) a near-surface water pool called the “growth pool” that is accessed by all plant species, but exploited more efficiently by opportunistic “live fast, die young” annual grasses and forbs (ruderal species), which possess shallow root systems and are characterized by rapid growth and water exploitation (Burnett and others 2016); and (ii) a deeper water pool called the “maintenance pool” which is mainly exploited by deep-rooted and perennial plant species (Schenk and Jackson 2002). Thus, plants within an ecosystem may respond to drought differently. This is important because vegetation cover is a crucial component of the water cycle (Asbjornsen and others 2011), controlling soil water over space and time (Wang and others 2019). Understanding community shifts in plant-soil water interactions in response to global change plays an important role in land management, conservation, and restoration (Asbjornsen and others 2011).
Arid and semiarid ecosystems cover about 50 % of the world’s surface (Ryel and others 2008). The foothill regions of southern California are a semiarid environment that encompasses a stable mosaic of coastal sage scrub (CSS) and annual grassland ecosystems (Minnich 1983; Bowler 2000). This area is considered a hotspot of biodiversity and endemism (Myers and others 2000; Kimball and others 2014). CSS is a diverse community composed of subshrubs, woody drought-deciduous shrubs, and evergreen shrubs (Kimball and others 2014). Southern California’s annual grasslands are dominated by invasive Eurasian grass and forb species, with native grasses and forbs making up the minority of the community (Bowler 2000). These diverse ecosystems are ideal for testing how different coexisting plant communities respond to global change (drought) and how the plant-soil water feedback differs from one plant community to the other.
In this study, we investigated plant-soil water interactions and soil water dynamics of the two dominant plant communities (shrubland and grassland) by looking at vegetation changes and soil moisture patterns to a depth of 2 m during years 5–8 of the ongoing Loma Ridge Global Change Experiment established in 2007. This experiment includes a long-term precipitation manipulation and aims to understand the resilience of Southern California’s ecosystems to changing environmental conditions like drought, fire, and nitrogen deposition, with a particular focus on aboveground biomass development, ecological succession (Potts and others 2012; Kimball and others 2014; Khalili and others 2016) and soil microbial activity (Allison and others 2013; Finks and others 2021). Acknowledging the importance of vegetation effects on soil water content, the specific questions addressed in this work are (i) how do changes in water input affect shrubland and grassland communities in Southern California? (ii) how do shrubs and grasses affect water content over time and along the soil profile? and (iii) how do shrubs and grasses amplify or dampen the effect of altered water input on soil water content? We hypothesized that due to plant-soil water feedbacks as a consequence of different rooting depths and plant functional types, there should be distinct soil water profiles in grassland versus shrubland. For example, under drought, the combined effect of low precipitation and efficient water exploitation by grassland species should be reflected in lower soil water content throughout the soil profile. Conversely, due to their deep root systems, shrubs should be more resistant to drought and thus, soil water content throughout the soil profile should be more stable regardless of water treatment. However, prolonged drought might reduce water recharge to deeper soil layers to a point where it could potentially affect perennial shrubs more than opportunistic, annual plant species.
Materials and Methods
Study Site
The Loma Ridge Global Change Experiment hosts a mosaic of annual grassland, dominated by invasive Eurasian grasses, adjacent to CSS, a drought-deciduous shrubland with some evergreen chaparral species (Allison and others 2013; Khalili and others 2016). It is located on a gentle, northwest-facing slope near the crest of Loma Ridge in the foothills of the Santa Ana Mountains in Orange County, California (Lulow and Young 2011; Kimball and others 2014; Khalili and others 2016; Kimball and others 2016). The site is located on the unceded ancestral homelands of the Acjachemen and Tongva Indigenous peoples. The border between these two plant communities is well-defined and has been relatively stable over the last 80 years (Potts and others 2012).
The site has a Mediterranean climate with monthly average temperatures ranging from 9.7 to 24.1 \(^\circ \)C, and minimum temperatures rarely dropping below 0\(^\circ \)C. Mean annual precipitation between 2007 and 2020 was 276 mm, and mostly occurs between November and April (Kimball and others 2014; Khalili and others 2016; Kimball and others 2016).
Experimental Design: Precipitation Manipulation Experiment
The precipitation manipulation of the experiment, as described in previous studies (Potts and others 2012; Allison and others 2013; Kimball and others 2014; Khalili and others 2016; Kimball and others 2016), started in 2007 and consisted of 24 plots within 8 blocks in CSS (each 18.3 \(\times \) 12.2 m) and 24 plots within 8 blocks in grassland (each 9.1 \(\times \) 6.1 m) communities. In both grassland and shrubland, one plot in each block was randomly selected to receive 40% additional water input (“added water” treatment), another eight were designated to receive 40% less water input (“drought” treatment), and the remaining eight were used as control plots (“ambient water” treatment). Each plot was subdivided into six subplots, 2 \(\times \) 2 m in the grassland and 4 \(\times \) 4 m in the shrubland (Figure S1). The uphill and downhill subplots of each plot were subject to artifacts due to wind and lateral water flow. Therefore, data were only collected from the middle subplots, either from the right or left side (Figure S1).
The drought treatment was achieved by constructing frames with translucent, retractable polyethylene roofs. These roofs remained open for roughly 95% of the year to reduce potential impacts on temperature and humidity (Kimball and others 2014). Air temperature and relative humidity sensors installed in representative plots showed no differences among the different treatments. Roofs were closed during larger storms to reduce precipitation frequency and amount in the drought treatment plots. Collected rainwater was stored in polyethylene tanks and applied to the added water treatment plots using a drip irrigation system. The volume of water added was controlled using Hersey positive-displacement water meters.
Over the study period (2011–2015), “drought” plots received 51% less water than ambient water plots, but added water plots received only 33% more than ambient water plots because storms occasionally overwhelmed the experiment’s water storage capacity (Kimball and others 2014). Based on the analysis of six nearby precipitation datasets, the water addition and restriction targets were selected to reflect the regional precipitation variability in southern California. This avoids the simulation of flood or drought events that are outside the study area’s observable historic range.
Soil Measurements
Soil Texture
Following the hydrometer method (Gee and Bauder 1986), soil texture was measured for each plot from 22 soil samples taken at depths 0, 15, 30, 45, 100, and 200 cm under constant temperature.
Soil Water Content
Soil volumetric water content was measured at the surface and down to 2 m along the soil profile. Three different devices were used, which allowed cross-validation of the measurements. Automated TDR sensors (Campbell CS616 30 cm Water Content Reflectometers) measured volumetric water content in the top 30 cm of soil every 30 min year-round. Two sensors were installed in each of the middle subplots and each of the lower subplots, totaling eight sensors for the entire plot.
A Campbell Hydrosense handheld TDR sensor was used to manually measure volumetric water content in the top 20 cm of soil at eight locations per plot with permanently installed TDR waveguides, four in the middle subplots and four in the lower subplots. (see Figure S1). These measurements were taken every two to three weeks during the wet (winter) season and every four to six weeks during the dry (summer) season.
A CPN Hydroprobe 503DR neutron probe was used to measure volumetric soil water content every 25 cm down to 2 m every two to three weeks during the wet season and four to six weeks during the dry season. Further information regarding data management and units transformation are found in the supplementary section “Additional data processing for soil water content”.
Estimation of Soil Water Content Metrics
We interpolated in time and space the measured volumetric water content along the soil profile using the Kriging method. To perform Kriging, we first transformed the data using the R package “sp” (Pebesma and Bivand 2005; Bivand and others 2013) and fitted the semivariogram using the function “variogram” with a Gaussian model in the package “gstat” (Pebesma and Wesseling 1998; Pebesma 2004; Gräler and others 2016) (Figure S3). Kriging was obtained with the function “krige” also included in the package “gstat”. Using the interpolated volumetric water content, together with our soil texture measurements, we estimated matric potential using empirical equations from Saxton and others (1986) that relate soil volumetric water content and soil texture:
where \(\theta \) is the soil volumetric water content and A and B:
where %C is the percent clay content in soil, and %S is the percent sand content in soil.
Additionally, we calculated the total extractable water (TEW) in mm as the difference between the ambient soil water content and water content at the wilting point, as proposed by Vicca and others (2012):
where sw is the soil volumetric water content, \(sw_{\text{wp}}\) is soil volumetric water content at the wilting point, and d is the thickness of the soil layer in mm.
The full analysis can be found in the following repository (https://doi.org/10.5281/zenodo.8242236).
Plant Measurements
Biomass of Grassland Species
Biomass in the grassland was measured by harvesting all annual herbaceous (including grass and forb species) that grow within 14 cm \(\times \) 50 cm quadrat placed on the ground at four different locations within the permanent monitoring subplot of each plot (Figure S1). These locations were changed annually to avoid resampling bias. The sampling was performed in late spring. Relative cover of grasses, forbs, litter, and bare soil was assessed and recorded within each quadrat prior to harvest. Live biomass and litter within the quadrats were harvested, separated, dried at 65\(^\circ \)C for 48 h, weighed, and scaled to the appropriate units (\(\hbox {g m}^{-2}\)).
Shrubland Community Composition
Shrubland community composition was measured annually. Each permanent subplot was divided into a grid of 0.5 m by 0.5 m squares, and all species present in each grid cell were recorded, with each cell accounting for approximately 1.6% of the total grid area. Measurements were conducted in late spring. Cover values were calculated based on the total ground cover by adding all recorded species, bare ground, and litter and rescaling to 100%.
Statistical Analysis
Statistical analyses were carried out for soil texture and plant variables (grassland biomass and shrubland community composition). Assumptions of normality of residuals and homogeneity of variances were visually verified using “qq-plots”-like plots from the “edress” package (Goode and others 2021) in R version 4.0.4 (2021-02-15) for all variables. In most cases, the collected data slightly violated the homoscedasticity and normality of residual assumptions even after data transformation (logarithmic, exponential, and Box-Cox transformations were tested). We proceeded with the statistical analysis using the untransformed data due to the robustness of the statistical method to minor deviations from these assumptions (Knief and Forstmeier 2021).
For soil texture, we used a linear mixed model for each soil fraction (clay, silt, and sand) using the function “lmer” from the package “lme4” (Bates and others 2015). As fixed effects, we used plant community (grassland and shrubland) and soil depth, and we used the block from the original experimental design as a random effect. Pairwise comparisons were carried out using the “glht” function from the “multcomp” package (Hothorn and others 2008). Similarly, we used a linear mixed model for biomass of grassland species, keeping water treatment (added, drought, and ambient) and year of sampling (2012–2015) as fixed effects and block as a random effect. Pairwise comparisons were also carried out using the “glht” function from the “multcomp” package (Hothorn and others 2008).
To test the effects of water treatment on shrubland community composition, we performed a PERMANOVA analysis using the seven most dominant species in the community that make up more than 75% of the total cover in the shrubland community from 2011 to 2015. The PERMANOVA analysis was performed using the “vegan” package (Oksanen and others 2022) and the function “adonis2” with a Bray–Curtis dissimilarity distance (Finks and others 2021) calculated with the “vegdist” function. Convergence of the “vegdist” was achieved, and the stress parameter obtained was 0.239. Additionally, a linear mixed model was performed on the cover of the six most dominant species, bare ground and litter. The linear mixed model included block as a random factor and water treatment and year as the fixed effects with log-transformed cover as the dependent variable.
To test differences in soil water content among treatments and plant communities, we used a multiple regression model, considering total extractable water as the dependent variable, and plant community type, soil depth, water treatment, and year and month of sampling as the independent variables.
Results
Water Input Across Treatments in Loma Ridge
During the first 10 years of the Loma Ridge Global Change experiment, the site observed varying precipitation patterns (Figure 1). In 2011, prior to the observation period of this study, a particularly wet year occurred and was followed by a drought between 2012 and 2015, with a severe drought event in 2014.
Effect of Water Input on Vegetation Patterns in Shrubland and Grassland
We compared changes in aboveground biomass in the grassland community, and vegetation cover and community composition in the shrubland community, under three water treatments (added, drought, and ambient) to identify potential plant-soil water feedbacks in response to altered precipitation regimes. We used metrics that are appropriate and tractable for each vegetation type. From a water balance perspective, grassland species composition is not very informative because all the grassland species are withdrawing water from the top 50 cm, and therefore, biomass is the most relevant metric for plant water relations in grassland. In shrubland, however, different species have different water use strategies; therefore, tracking shrubland community composition is meaningful.
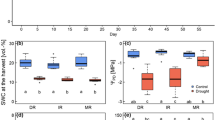
Grassland biomass under the drought treatment (mean value over the study period: 216.7 ± 168.5 \(\hbox {g m}^{-2}\)) was lower than for the ambient treatment (mean value over the study period: 361.8 ± 176.3 \(\hbox {g m}^{-2}\)), excepting years 2011, 2014 and 2015 (Figure 2). Water addition led to increased grassland biomass (mean value over the study period: 467.0 ± 151.7 \(\hbox {g m}^{-2}\)) compared to the drought treatment, but was only statistically higher than the ambient treatment in years 2012 and 2013 (Figure 2). In general, low biomass production was apparent from 2012 to 2014, when a severe drought in California exacerbated the severity of the drought treatment, reducing biomass to almost zero.
We observed potential shifts in shrubland community composition as a consequence of drought (Table 1). However, drought responses in the shrubland were species-specific (Figures 3, S6). For example, the grass species B. madritensis and E. condensatus were not significantly affected by the drought treatment throughout the study period, whereas the mid-rooted shrub S. mellifera declined (Figures 3a, S6, post hoc glht, \(p = 0.009\)). Cover of the relatively shallow-rooted shrub A. glaber was lower under the drought treatment than under the other treatments (Figure S6, \(p < 0.001\)), and its cover strongly declined regardless of water treatment after 2012 (Figures 3a, S6, \(p < 0.001\) for year effect). Cover of the mid-rooted shrub species A. californica and the deep-rooted species M. laurina were mostly unaffected by the drought treatment (Figures 3a, S6). Only bare soil (\(p = 0.003\)) and litter (\(p < 0.001\)) consistently increased under the drought treatment (Figures 3f, S6).
Effect of Vegetation Type on Soil Water Content
In our precipitation manipulation experiment, we observed distinct soil water patterns in grassland and shrubland communities. At Loma Ridge, soil texture was remarkably consistent with depth and between the grassland and shrubland communities (predominantly sandy loam throughout the soil profile, see Figure S4). Due to similar soil properties and close proximity of shrubland and grassland communities, we expect surface runoff, groundwater recharge, and evaporation to have similar impacts on the observed soil water patterns. Therefore, we assumed that matric potential differences are mainly driven by vegetation effects (Figure 4). Community differences in soil water content were more pronounced in the deep soil layers, and seasonal patterns were only visible at shallow soil depths (Figure 4a, b). To a depth of approximately 50 cm, the soil was slightly drier in grassland compared to shrubland, with grassland soils containing 10 mm less water than shrubland soils (Figure 4c, Table S1). Soil matric potential (pF) patterns in shrubland were consistent across the entire sampling depth of 2 m. For example, shrubland layers below 75 cm depth hold 75 mm less water than the corresponding grassland layers, suggesting water uptake from deeper soil layers by shrubs (Figure 4c, Table S1). Additionally, deep grassland soils were mostly saturated, except for soil layers between 150 to 200 cm depth, potentially due to low infiltration amounts from precipitation to this depth (Figure 1).
Coupled Effect of Water Treatment and Vegetation on Soil Water Content
We tested how plant communities influence soil water patterns along the soil profile under different water treatments. The observed soil water dynamics across water treatments were a combined result of the varying plant responses to water input via plant-soil water feedbacks. Without the influence of vegetation, we would expect seasonal soil water content variation at the surface and diminishing seasonal signal with increasing soil depth. Thus, we would expect greater water content in the surface soils of the added treatment (green color, Figure 5a). Conversely, we would expect drier soils (purple color), at the surface when the drought treatment is applied during the rainy season (Figure 5d). Any deviations from these null expectations should be due to plant-soil water feedbacks affecting soil water infiltration, uptake, or evapotranspiration.
In both plant communities, added water treatment effects followed our expectations based on our null hypothesis, especially near the soil surface (Figure 5b, c). Shrubland soils responded more strongly to the added water treatment in which deeper soil layers were wetter, potentially as a consequence of water percolation.
Stronger effects were observed under the drought treatment (Figure 5e, f). For grassland, at shallow depths (0 cm to 50 cm), soils were slightly drier under drought compared to ambient (soils under drought contained about 10 mm less water than under the ambient treatment) (Figure 5f, Table S1). This is the zone of most active water uptake in grassland (Coupland and Johnson 1965; Sala and others 1989; Feng and others 2021), largely because these grass species have root systems that do not extend below 50 cm. Soil water content slightly increased between 50 and 125 cm and decreased again below 125 cm under the drought treatment in grassland. In 2014 and 2015, this pattern changed. Between 75 cm and 125 cm, there was little to no drought treatment effect (Figure 5f). This zone was consistently at or near saturation year-round because it is below the main rooting depth. However, treatment effects were weaker in 2014–2015 (Figure 1), likely because water input for the drought treatment was only reduced by 10% (target reduction value was 40%) compared to the ambient treatment.
Soil water content in shrubland soils was affected by the drought treatment (Figure 5e). Interestingly, under the drought treatment, shrubland soils held about 40 mm more water at depths between 50 cm and 150 cm than under the ambient treatment during the entire observation period (Figure 5e and Table S1)
Discussion
In our study, we evaluated how drought affects plant-soil water feedbacks of two plant communities (shrubland and grassland) in Southern California. The plants in these communities have distinct rooting depths and growth strategies, resulting in different responses to drought stress (Figure 6). The grassland is relatively uniform, consisting of plant species with similar root architectures that efficiently exploit soil water from the top 50 cm. These species are severely impacted by drought but quickly recover when drought ends due to their opportunistic “live fast, die young” life history strategy. The response of shrubland species to drought is more diverse, consistent with observed shifts in community composition. For example, mid-rooted species declined under drought, but shallow- and deep-rooted species remained unaffected. The different responses of grassland and shrubland to drought are reflected in unique water distributions across the soil profile. For example, ambient grassland soils below 50 cm are close to saturation but drought-induced reductions in plant cover may favor surface runoff and water evaporation, leading to reductions in soil water at that depth. In shrubland, drought increases soil moisture at a depth of 75 to 150 cm, potentially due to the loss of mid-rooted species that would exploit soil water from those depths. However, greater water exploitation with drought is observed at shallow depths and at deep soil layers below 150 cm in shrubland.
Contrasting Soil Water Profiles as a Consequence of Different Plant Life Strategies
Specific plant communities can develop characteristic patterns of soil water due to the interplay of root uptake and soil physical properties like soil texture (Fernandez-Illescas and others 2001; Lehmann and others 2018). Soil texture, for example, is an important soil property for water evaporation (Lehmann and others 2018) and runoff (Fernandez-Illescas and others 2001), and thus indirectly influences plant water stress (Fernandez-Illescas and others 2001; Rodriguez-Iturbe and others 2001). Soil texture at Loma Ridge was remarkably consistent with depth and also between the grassland and shrubland communities (Figure S4). Therefore, the distinct soil water usage patterns observed at Loma Ridge are largely driven by the different water use strategies of the grassland versus shrubland plants.
Plants use different strategies to cope with adverse conditions (Lambers and others 2008; Reichstein and others 2014; Malíková and others 2016; Carvajal and others 2019), forming a so-called fast-slow plant economy spectrum among species (Carvajal and others 2019). Strategies can include changes in aboveground or belowground morphology and architecture (Lee and Lauenroth 1994; Parolari and others 2015; Arbona and others 2017; Seleiman and others 2021; Kirschner and others 2021), special plant traits (early maturity, rolling leaves, high stomatal conductance, etc.) (Franks 2011; Seleiman and others 2021), or changes in plant metabolism (Arbona and others 2017; Seleiman and others 2021) to optimize scarce resource use. Annual shallow-rooted grasses and forbs, especially those in more arid environments, are typically characterized by a “live fast, die young” life strategy (Burnett and others 2016) in which germination, rapid growth and flowering occur following the first rain of the wet season and senescence takes place after soil water is depleted (Clary and others 2004). Shrubs and other woody species are perennials that rely on deep root systems to withdraw water more slowly than shallowly rooted annuals but for a longer time of the year (Schenk and Jackson 2002).
The water use dynamics observed in Loma Ridge are in line with the two soil water pools described by (Ryel and others 2008). The “growth pool” is heavily exploited and contributes to the growth of both grass and shrub species regardless of year and precipitation treatment. The “maintenance pool” is mainly exploited by mid- and deep-rooted, perennial shrub species. The active water withdrawal zone in grassland ends at a depth of approximately 50 cm, indicated by the zone of stable soil water content that occurs below 50 cm, showing no seasonal influence (Figure 4b). These results are in line with previous studies showing that water uptake in grasslands occurs predominantly from shallow soil layers (Coupland and Johnson 1965; Sala and others 1989; Feng and others 2021).
In contrast to grasses, shrubs exploit soil water throughout the entire soil profile, although with less intensity in deeper soil layers (Figure 4a), which is in line with previous studies (Canadell and others 1996; Schenk and Jackson 2002; Sala and others 1989; Lee and Lauenroth 1994). CSS at Loma Ridge is composed of shrub species with three main root morphologies (Davis and Mooney 1986; Jacobsen and Pratt 2018): (i) shallow-rooted species (A. glaber), (ii) mid-rooted species (S. mellifera), and (iii) deep-rooted species (M. laurina), all of which are often associated with chaparral communities (Paddock and others 2013; Rundel 2018). Although we recorded water uptake to a depth of 2 m, some shrub species’ roots can reach substantially deeper than 2 m (Hellmers and others 1955; Canadell and others 1996). Therefore, water uptake occurs along the soil profile, with particularly high intensity at shallow depths because fine roots of shallow-rooted species are more active (Rodríguez and others 2007) (Figure 4a).
Plant-Soil Water Feedbacks in Loma Ridge Under Drought
We hypothesized that the combined effect of drought and plant life strategies might lead to plant-soil water feedbacks potentially favoring one community over the other. Understanding these feedbacks is particularly important for plant communities in semiarid environments such as California, where the severity and frequency of droughts are projected to increase (Griffin and Anchukaitis 2004; Yoon and others 2015).
Based on our conceptual null hypothesis, which excludes the influence of vegetation on soil water content, we expected that seasonal patterns of soil moisture would be limited to the upper soil layer and disappear with depth (Figure 5a, d). Under the influence of vegetation, our observations show distinct plant-soil water feedbacks throughout the soil profile (Figure 5b, c, e, f). For example, grassland biomass was consistently higher under the added water treatment than the other treatments (Figure 2). Added water promoted more herbaceous growth and increased moisture withdrawal from the shallow soil layer (Seyfried and Wilcox 2006; Lauenroth and Adler 2008) which quantitatively offset the moisture addition, resulting in similar levels of near-surface water content compared to the other treatments (Figure 5c). We also observed that reduced precipitation led to reduced biomass growth. Between 2012 and 2014, a marked drought period correlated with strongly reduced productivity under the drought treatment (Figure 2). The loss of plant cover increases evaporation and favors superficial runoff (Liu and others 2020) as a consequence of crust formation due to the disturbance of soil aggregates by rain droplets (Carlesso and others 2011; de Almeida and others 2018). The substantial reduction in biomass was therefore reflected by lower soil water content, a lower maximum depth of infiltration and a lack of recharge especially below 150 cm in the drought treatment (Figure 5f). Compared to ambient and added treatment, water that infiltrated into the soil led to greater increases in total extractable soil water, likely because biomass loss reduced transpiration (Figure S5). In 2014 and 2015, soil water content was similar or greater in the drought treatment compared to the ambient treatment below 75 cm (Figure 5f). Because a severe drought in the area was affecting all treatments, the absolute reduction in precipitation was lower in 2014, which may have weakened the drought treatment effect on soil water. The drought treatment effect remained weak in 2015 because precipitation was only reduced by 10% in that year.
Unlike in the grassland, we did not observe short-term plant-soil feedbacks in the shrubland. Instead, the abundance and dominance of shrubs and herbaceous species exhibited diverse responses to water treatments over multiple years, showing a weak coupling between plant species and recent water inputs. One major observation was the consistently wetter zone at depths between 50 cm and 150 cm under the drought treatment (Figure 5e). This observation may be explained by a loss of mid-rooted shrub species that take up more water from these depths. Shallow- and mid-rooted shrub species tend to be more sensitive to prolonged and severe drought (Paddock and others 2013; Venturas and others 2016; Jacobsen and Pratt 2018), despite the fact that drought tolerance does not necessarily correlate with root depth (Davis and Mooney 1986).
However, S. mellifera was the only mid-rooted shrub affected by the drought treatment at Loma Ridge (Figures 3a, S6). Cover of the shallow-rooted shrub A. glaber was also lower in the drought treatment, consistent with results from 2009–2012 at the same site (Kimball and others 2014). A. glaber cover declined sharply between 2013 to 2015 in all water treatments, which is likely due to ecological succession following the Santiago wildfire in 2007, perhaps compounded by severe drought that impacted the region during this period (Figure 1). The initial cover of A. glaber in the drought treatment was lower than the other treatments in 2009, suggesting that the drought treatment might have affected germination rates of A. glaber after the Santiago fire. Another mid-rooted shrub, A. californica, was mostly unaffected by drought (Figures 3a, S6), suggesting that other phenological and physiological differences between species also contribute to their tolerance to drought stress. For example, A. californica is both drought-deciduous and has small, feathery leaves that resist stomatal water loss, whereas S. mellifera has larger, evergreen leaves. Additionally, A. californica is a resprouting species (Frazer and Davis 1988) that is therefore more resilient to disturbance compared to the obligate seeder species like A. glaber (Keeley and others 2006). Deep-rooted species such as M. laurina were generally unaffected by drought (Figure 3 A and Figure S6). These species can access very deep water and as adult shrubs are typically not affected by the transient abundance of annual grasses and forbs.
Moreover, grass species such as B. madritensis and E. condensatus were unaffected by the drought treatment over the course of the study period (Figures 3a, S6) in the shrubland community. This observation is in line with Seyfried and Wilcox (2006), who stated that herbaceous species with shallow root systems typically invade empty niches left by lost woody species. At Loma Ridge, these two species could be taking over the space left by S. mellifera under drought. Moreover, these grass species could be profiting from deep surface water stocks through hydraulic lift from deep-rooted species (Caldwell and others 1998). Thus, the plant-soil water feedbacks could be changing toward greater competition for water from the surface soil layers, deep surface water use through hydraulic lift and a prominent lack of withdrawal from middle depths by S. mellifera.
Based on our observations, prolonged drought events might significantly affect the water maintenance pool (See Section 6.1), which is critical for the survival of shrubs, thus posing a long-term risk to this plant community. Early signs of this trend are noticeable at Loma Ridge, where slower-growing shrub species are displaced by annual grasses exhibiting a live fast, die young water use strategy. Thus, drought plays a role in the long-term community composition, as indicated by the PERMANOVA analysis (Table 1). However, caution is advised in interpreting this shift in community composition. The current patterns of shrub community composition may be the result of a combined effect of fire and water treatment (Figure 3) due to a wildfire that occurred in the area in 2007 Potts and others (2012). Nevertheless, the plant-soil water feedbacks observed in this study can be leveraged to complement global climate models to better assess the effects of drought on the ecohydrology of semiarid terrestrial ecosystems.
Our long-term experiment at Loma Ridge revealed important plant-soil feedbacks (Figure 6). By comparing the effects of altered precipitation regimes on two coexisting plant communities, we observed different drought responses depending on the communities’ distinct plant life strategies. The plant-soil water feedbacks found between shrubs and grasses might help maintain the current vegetation mosaic at Loma Ridge by impeding the encroachment of grass species into well-established patches of shrubs and vice versa (Eppinga and others 2018). Shallow-rooted grasses seem to prevent the invasion of shrubs due to the rapid exploitation of water, leading to soil layers near the surface (Eliason and Allen 1997) becoming too dry for new shrubs to establish. On the other hand, established deep-rooted shrubs can either outcompete or at least tolerate new grasses (Kimball and others 2014) because they can access deeper water stocks. Even within the same plant community, plant responses to drought differ. For example, shrub species that are obligate seeders, like S. mellifera, tend to have a disadvantage in reestablishment after a strong environmental disturbance (Frazer and Davis 1988; Hodgkinson 1998; Jacobsen and Pratt 2018), due to i) higher water stress and ii) direct competition of shrub seedlings with faster-growing annual grasses. Moreover, these mid-rooted shrub species might be more susceptible to drought as they cannot compete with shallow rooted water exploiters (grasses), nor access deep water pools. In order to design management practices for the conservation of these ecosystems, it is important that future studies focus on the combined effects of major disturbances, e.g., long-term droughts, wildfires, and pollution, and their impacts on plant-soil water feedbacks.

Annual water input (mm) under the different treatments from water years 2008 to 2017. Each water year includes the precipitation events from October of the preceding year to September of that water year. The vertical red lines indicate the start and end years of the study period used in our analysis. The horizontal line indicates the mean annual precipitation over the last 40 years calculated using grid data from the Center for Hydrometeorology and Remote Sensing (CHRS) at the University of California, Irvine (UCI) (Sorooshian 2021).

Herbaceous aboveground biomass of grassland (\({\text{g m}}^{-2}\)) by treatment and year determined by the annual harvest of a subset of each experimental plot. Error bars represent mean ± standard error. Letters indicate significant statistical difference among water treatments and sampling year following a mixed model and pairwise comparisons (see section “Materials and methods”).

Distribution of dominant shrubland species per water treatment from 2011 to 2015 (a–c). Shades of blue represent shrubs, shades of green represent grasses and shades of red represent forbs. Distribution of soil cover types: bare ground, litter, and vegetation per water treatment from 2011 to 2015 (d–f). The shrubland species shown were the most abundant species that accounted for at least 75% of the ground cover (litter and bare ground included).

Spatiotemporal variation in matric potential (pF) in shrubland (a) and grassland (b) under the ambient water treatment. Black lines indicate the matric potential at the wilting point and red lines indicate the matric potential at field capacity. c Total extractable water (mm/cm) differences between shrubland and grassland (a, b).

Spatiotemporal differences in total extractable water (mm/cm) in shrubland and grassland under the added water and drought treatments. a, d A conceptual null hypothesis of total extractable water differences without the influence of vegetation. Color patterns in the null hypothesis are intended to show general trends and values were not derived from real data. Additional panels show total extractable water differences between the added and ambient (b, c), and the drought and ambient water treatments (e, f) for shrubland (b, e) and grassland (c, f).

Conceptual model of plant-soil water interactions in shrubland and grassland in Loma Ridge under drought including the growth and maintenance pools model described by Ryel and others (2008). In shrubland, all species mainly use water from the first 50 cm so that under drought, soil water content declines even if shrub biomass is lower. Mid-rooted species are the most impacted by drought, and biomass loss is reflected in increased soil water content at depths between 50 and 100 cm. Finally, deep-rooted shrub species are mostly unaffected by drought. Grassland species, a uniform community with similar root architectures among species, mainly take up water from the top 50 cm. Under drought, low water input reduces biomass growth and water uptake. However, grassland plants and evaporation continue to remove water from the surface soil. The loss of grassland might also reduce deep water recharge due to higher surface runoff and evaporation (“created with Biorender.com”).
Data availability
The data that support the findings of this study are openly available in the repository: https://doi.org/10.5281/zenodo.8242236.
References
Allison S, Lu Y, Weihe C, Goulden M, Martiny A, Treseder K, Martiny J. 2013. Microbial abundance and composition influence litter decomposition response to environmental change. Ecology 94(3):714–725. https://doi.org/10.1890/12-1243.1.
Arbona V, Manzi M, Zandalinas SI, Vives-Peris V, Pérez-Clemente RM, Gómez-Cadenas A. 2017. Physiological, Metabolic, and Molecular Responses of Plants to Abiotic. Stress. https://doi.org/10.1007/978-3-319-42183-4_1.
Asbjornsen H, Goldsmith GR, Alvarado-Barrientos MS, Rebel K, Van Osch FP, Rietkerk M, Chen J, Gotsch S, Tobon C, Geissert DR, Gomez-Tagle A, Vache K, Dawson TE. 2011. Ecohydrological advances and applications in plant-water relations research: a review. Journal of Plant Ecology 4(1–2):3–22. https://doi.org/10.1093/jpe/rtr005.
Beaulieu E, Lucas Y, Viville D, Chabaux F, Ackerer P, Goddéris Y, Pierret M-C. 2016. Hydrological and vegetation response to climate change in a forested mountainous catchment. Modeling Earth Systems and Environment 2(4):1–15. https://doi.org/10.1007/s40808-016-0244-1.
Bhattacharya, A. (2021). Effect of Soil Water Deficits on Plant–Water Relationship: A Review. inSoil Water Deficit and Physiological Issues in Plants (pages 1–98). Springer Singapore. https://doi.org/10.1007/978-981-33-6276-5_1
Bivand R, Pebesma E, Gómez-Rubio V. 2013. Classes for Spatial Data in R. https://doi.org/10.1007/978-1-4614-7618-4_2.
Bowler PA. 2000. Ecological Restoration of Coastal Sage Scrub and Its Potential Role in Habitat Conservation Plans. Environmental Management 26:S85–S96. https://doi.org/10.1007/s002670010064.
Burnett AC, Rogers A, Rees M, Osborne CP. 2016. Carbon source-sink limitations differ between two species with contrasting growth strategies. Plant, Cell Environment 39:2460–2472. https://doi.org/10.1111/pce.12801.
Caldwell MM, Dawson TE, Richards JH. 1998. Hydraulic lift: consequences of water efflux from the roots of plants. Oecologia 113(2):151–161. https://doi.org/10.1007/s004420050363.
Canadell J, Jackson RB, Ehleringer JB, Mooney HA, Sala OE, Schulze E-D. 1996. Maximum rooting depth of vegetation types at the global scale. Oecologia 108:583–595. https://doi.org/10.1007/BF00329030.
Carlesso R, Spohr R, Eltz F, Flores C. 2011. Runoff estimation in southern Brazil based on Smith’s modified model and the Curve Number method. Agricultural Water Management 98(6):1020–1026. https://doi.org/10.1016/j.agwat.2011.01.012.
Carvajal DE, Loayza AP, Rios RS, Delpiano CA, Squeo FA. 2019. A hyper-arid environment shapes an inverse pattern of the fast-slow plant economics spectrum for above-, but not below-ground resource acquisition strategies. Journal of Ecology 107:1079–1092. https://doi.org/10.1111/1365-2745.13092.
Clary J, Save R, Biel C, Herralde F. 2004. Water relations in competitive interactions of Mediterranean grasses and shrubs. Annals of Applied Biology 144:149–155. https://doi.org/10.1111/j.1744-7348.2004.tb00328.x.
Coupland RT, Johnson RE. 1965. Rooting Characteristics of Native Grassland Species in Saskatchewan. The Journal of Ecology 53:475. https://doi.org/10.2307/2257990.
Davis SD, Mooney HA. 1986. Tissue water relations of four co-occurring chaparral shrubs. Oecologia 70:527–535. https://doi.org/10.1007/BF00379899.
de Almeida WS, Panachuki E, de Oliveira PTS, da Silva Menezes R, Sobrinho TA, de Carvalho DF. 2018. Effect of soil tillage and vegetal cover on soil water infiltration. Soil and Tillage Research 175:130–138. https://doi.org/10.1016/j.still.2017.07.009.
Eliason SA, Allen EB. 1997. Exotic Grass Competition in Suppressing Native Shrubland Re-establishment. Restoration Ecology 5:245–255. https://doi.org/10.1046/j.1526-100X.1997.09729.x.
Eppinga MB, Baudena M, Johnson DJ, Jiang J, Mack KML, Strand AE, Bever JD. 2018. Frequency-dependent feedback constrains plant community coexistence. Nature Ecology Evolution 2:1403–1407. https://doi.org/10.1038/s41559-018-0622-3.
Feng H, Zhou J, Zhou A, Bai G, Li Z, Chen H, Su D, Han X. 2021. Grassland ecological restoration based on the relationship between vegetation and its below-ground habitat analysis in steppe coal mine area. Science of The Total Environment 778:146221. https://doi.org/10.1016/j.scitotenv.2021.146221.
Fernandez-Illescas CP, Porporato A, Laio F, Rodriguez-Iturbe I. 2001. The ecohydrological role of soil texture in a waterlimited ecosystem. Water Resources Research 37:2863–2872. https://doi.org/10.1029/2000WR000121.
Finks S, Weihe C, Kimball S, Allison S, Martiny A, Treseder K, Martiny J. 2021. Microbial community response to a decade of simulated global changes depends on the plant community. Elementa: Science of the Anthropocene 9(1):00124. https://doi.org/10.1525/elementa.2021.00124.
Franks SJ. 2011. Plasticity and evolution in drought avoidance and escape in the annual plant Brassica rapa. New Phytologist 190:249–257. https://doi.org/10.1111/j.1469-8137.2010.03603.x.
Frazer JM, Davis SD. 1988. Differential survival of chaparral seedlings during the first summer drought after wildfire. Oecologia 76:215–221. https://doi.org/10.1007/BF00379955.
Gee, G. W., & Bauder, J. W. (1986). Particle-size Analysis in Methods of Soil Analysis (A. Klute, editor). https://doi.org/10.2136/sssabookser5.1.2ed.c15
Goode, K., McClernon, K., Zhao, J., Zhang, Y., & Huo, Y. (2021). goodekat/redres: Residuals and Diagnostic Plots for Mixed Models [MIT + file LICENSE].
Gräler, B., Pebesma, E., & Heuvelink, G. (2016). Spatio-Temporal Interpolation using gstat. The R Journal, 8, 204. https://doi.org/10.32614/RJ-2016-014
Griffin D, Anchukaitis KJ. 2004. How unusual is the 2012–2014 California drought? Geophysical Research Letters 41:9017–9023. https://doi.org/10.1002/2014GL062433.
Hajek O, Knapp A. 2022. Shifting seasonal patterns of water availability: ecosystem responses to an unappreciated dimension of climate change. New Phytologist 233(1):119–125. https://doi.org/10.1111/nph.17728.
Hanson PJ, Weltzin JF. 2000. Drought disturbance from climate change: response of United States forests. Science of The Total Environment 262(3):205–220. https://doi.org/10.1016/S0048-9697(00)00523-4.
Hellmers H, Horton JS, Juhren G, O’Keefe J. 1955. Root Systems of Some Chaparral Plants in Southern California. Ecology 36:667–678. https://doi.org/10.2307/1931305.
Hickel K, Zhang L. 2006. Estimating the impact of rainfall seasonality on mean annual water balance using a top-down approach. Journal of Hydrology 331:409–424. https://doi.org/10.1016/j.jhydrol.2006.05.028.
Hodgkinson KC. 1998. Sprouting success of shrubs after fire: heightdependent relationships for different strategies. Oecologia 115:64–72. https://doi.org/10.1007/s004420050492.
Hothorn T, Bretz F, Westfall P. 2008. Simultaneous Inference in General Parametric Models. Biometrical Journal 50(3):346–363.
Jacobsen AL, Pratt RB. 2018. Extensive drought-associated plant mortality as an agent of type-conversion in chaparral shrublands. New Phytologist 219:498–504. https://doi.org/10.1111/nph.15186.
Keeley JE, Fotheringham CJ, Baer-Keeley M. 2006. Demographic patterns of postfire regeneration in Mediterranean-climate shrublands of California. Ecological Monographs 76(2):235–255. https://doi.org/10.1890/0012-9615(2006)076[0235:DPOPRI]2.0.CO;2.
Khalili B, Ogunseitan O, Goulden M, Allison S. 2016. Interactive effects of precipitation manipulation and nitrogen addition on soil properties in California grassland and shrubland. Applied Soil Ecology 107:144–153. https://doi.org/10.1016/j.apsoil.2016.05.018.
Kimball S, Funk J, Spasojevic M, Suding K, Parker S, Goulden M. 2016. Can functional traits predict plant community response to global change? Ecosphere 7(12):e01602. https://doi.org/10.1002/ecs2.1602.
Kimball S, Goulden M, Suding K, Parker S. 2014. Altered water and nitrogen input shifts succession in a southern California coastal sage community. Ecological Applications 24(6):1390–1404. https://doi.org/10.1890/13-1313.1.
Kirschner GK, Xiao TT, Blilou I. 2021. Rooting in the Desert: A Developmental Overview on Desert Plants. Genes 12:709. https://doi.org/10.3390/genes12050709.
Knief U, Forstmeier W. 2021. Violating the normality assumption may be the lesser of two evils. Behavior Research Methods 53(6):2576–2590. https://doi.org/10.3758/s13428-021-01587-5.
Lambers, H., Chapin, F. S., & Pons, T. L. (2008). Plant Physiological Ecology (2 edition). Springer New York. https://doi.org/10.1007/978-0-387-78341-3
Lauenroth & Adler, P. 2008. Demography of perennial grassland plants: survival, life expectancy and life span. Journal of Ecology 96(1023):1032. https://doi.org/10.1111/j.1365-2745.2008.01415.x.
Lee CA, Lauenroth WK. 1994. Spatial Distributions of Grass and Shrub Root Systems in the Shortgrass Steppe. American Midland Naturalist 132:117. https://doi.org/10.2307/2426206.
Lehmann P, Merlin O, Gentine P, Or D. 2018. Soil Texture Effects on Surface Resistance to Bare-Soil Evaporation. Geophysical Research Letters 45. https://doi.org/10.1029/2018GL078803.
Liu Y, Guo L, Huang Z, López-Vicente M, Wu G-L. 2020. Root morphological characteristics and soil water infiltration capacity in semi-arid artificial grassland soils. Agricultural Water Management 235:106153. https://doi.org/10.1016/j.agwat.2020.106153.
Lulow ME, Young TP. 2011. Is there still native diversity in California grasslands. Fremontia 39(2):6–11.
Malíková L, Latzel V, Smilauer P, Klimeaova J. 2016. Local adaptation of annual weed populations to habitats differing in disturbance regime. Evolutionary Ecology 30(861):876. https://doi.org/10.1007/s10682-016-9845-4.
Marks D, King G, Dolph J. 1993. Implications of climate change for the water balance of the Columbia River Basin, USA. Climate Research 2(203):213. https://doi.org/10.3354/cr002203.
Minnich RA. 1983. Fire Mosaics in Southern California and Northern Baja California. Science 219(1287):1294. https://doi.org/10.1126/science.219.4590.1287.
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J. 2000. Biodiversity hotspots for conservation priorities. Nature 403(6772):853–858. https://doi.org/10.1038/35002501.
Oksanen, J., Simpson, G. L., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’Hara, R. B., Solymos, P., Stevens, M. H. H., Szoecs, E., Wagner, H., Barbour, M., Bedward, M., Bolker, B., Borcard, D., Carvalho, G., Chirico, M., Caceres, M. D., Durand, S., . . . Weedon, J. (2022). vegan: Community Ecology Package. https://CRAN.Rproject.org/package=vegan
Ostle NJ, Smith P, Fisher R, Ian Woodward F, Fisher JB, Smith JU, Galbraith D, Levy P, Meir P, McNamara NP, Bardgett RD. 2009. Integrating plant-soil interactions into global carbon cycle models. Journal of Ecology 97(5):851–863. https://doi.org/10.1111/j.1365-2745.2009.01547.x.
Paddock W, Davis S, Pratt B, Jacobsen A, Tobin M, López-Portillo J, Ewers F. 2013. Factors Determining Mortality of Adult Chaparral Shrubs in an Extreme Drought Year in California. Aliso 31:49–57. https://doi.org/10.5642/aliso.20133101.08.
Parolari AJ, Goulden M, Bras R. 2015. Ecohydrological controls on grass and shrub above-ground net primary productivity in a seasonally dry climate. Ecohydrology 8:1572–1583. https://doi.org/10.1002/eco.1605.
Pebesma E. 2004. Multivariable geostatistics in S: the gstat package. Computers Geosciences 30:683–691. https://doi.org/10.1016/j.cageo.2004.03.012.
Pebesma, E., & Bivand, R. (2005). Classes and methods for spatial data in R. R News, 5, 9–13. https://CRAN.R-project.org/doc/Rnews/
Pebesma E, Wesseling C. 1998. Gstat: a program for geostatistical modelling, prediction and simulation. Computers Geosciences 24:17–31. https://doi.org/10.1016/S0098-3004(97)00082-4.
Pokhrel Y, Felfelani F, Satoh Y, Boulange J, Burek P, Gädeke A, Gerten D, Gosling SN, Grillakis M, Gudmundsson L, Hanasaki N, Kim H, Koutroulis A, Liu J, Papadimitriou L, Schewe J, Müller Schmied H, Stacke T, Telteu C-E, Wada Y. 2021. Global terrestrial water storage and drought severity under climate change. Nature Climate Change 11(3):226–233. https://doi.org/10.1038/s41558-020-00972-w.
Porporato A, Laio F, Ridolfi L, Rodriguez-Iturbe I. 2001. Plants in water-controlled ecosystems: active role in hydrologic processes and response to water stress. Advances in Water Resources 24:725–744. https://doi.org/10.1016/S0309-1708(01)00006-9.
Potts D, Suding K, Winston G, Rocha A, Goulden M. 2012. Ecological effects of experimental drought and prescribed fire in a southern California coastal grassland. Journal of Arid Environments 81:59–66. https://doi.org/10.1016/j.jaridenv.2012.01.007.
Reichstein M, Richardson AD, Migliavacca M, Carvalhais N. 2014. Plant-Environment Interactions Across Multiple Scales. https://doi.org/10.1007/978-1-4614-7501-922.
Rodríguez MV, Bertiller MB, Bisigato A. 2007. Are fine roots of both shrubs and perennial grasses able to occupy the upper soil layer? A case study in the arid Patagonian Monte with non-seasonal precipitation. Plant and Soil 300:281–288. https://doi.org/10.1007/s11104-007-9415-1.
Rodriguez-Iturbe I. 2000. Ecohydrology: A hydrologic perspective of climate-soil-vegetation dynamies. Water Resources Research 36:3–9. https://doi.org/10.1029/1999WR900210.
Rodriguez-Iturbe I, Porporato A, Laio F, Ridolfi L. 2001. Plants in water-controlled ecosystems: active role in hydrologic processes and response to water stress. Advances in Water Resources 24:695–705. https://doi.org/10.1016/S0309-1708(01)00004-5.
Rodriguez-Iturbe I, Porporato A, Ridolfi L, Isham V, Coxi DR. 1999. Probabilistic modelling of water balance at a point: the role of climate, soil and vegetation. Proc. R. Soc. Lond. A. 455:3789–3805. https://doi.org/10.1098/rspa.1999.0477.
Rundel PW. 2018. California Chaparral and Its Global. Significance. https://doi.org/10.1007/978-3-319-68303-4_1.
Ryel, R. J., Ivans, C. Y., Peek, M. S., & Leffler, A. J. (2008). Functional Differences in Soil Water Pools: a New Perspective on Plant Water Use in Water- Limited Ecosystems. inU. Lüttge, W. Beyschlag & J. Murata (editors), Progress in Botany (pages 397–422). Springer Berlin Heidelberg. https://doi.org/10.1007/978-3-540-72954-9_16
Sala OE, Golluscio RA, Lauenroth WK, Soriano A. 1989. Resource partitioning between shrubs and grasses in the Patagonian steppe. Oecologia 81:501–505. https://doi.org/10.1007/BF00378959.
Saxton KE, Rawls WJ, Romberger JS, Papendick RI. 1986. Estimating Generalized Soil-water Characteristics from Texture. Soil Science Society of America Journal 50(4):1031–1036. https://doi.org/10.2136/sssaj1986.03615995005000040039x.
Schenk HJ, Jackson RB. 2002. The global biogeography of roots. Ecological Monographs 72:311–328. https://doi.org/10.1890/0012-9615(2002)072[0311:TGBOR]2.0.CO;2.
Seleiman MF, Al-Suhaibani N, Ali N, Akmal M, Alotaibi M, Refay Y, Dindaroglu T, Abdul- Wajid HH, Battaglia ML. 2021. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 10:259. https://doi.org/10.3390/plants10020259.
Seyfried MS, Wilcox BP. 2006. Soil water storage and rooting depth: key factors controlling recharge on rangelands. Hydrological Processes 20:3261–3275. https://doi.org/10.1002/hyp.6331.
Silber, M., Bonetti, S., Gandhi, P., Gowda, K., Iams, S., & Porporato, A. M. (2017). The Role of Different Plant Soil-Water Feedbacks in Models of Dryland Vegetation Patterns. 2017, H21M–04. https://ui.adsabs.harvard.edu/abs/2017AGUFM.H21M..04S
Sorooshian, S. (2021). PERSIANN-CCS-CDR [Type: dataset]. https://doi.org/10.11572/P24W2F
Venturas MD, MacKinnon ED, Dario HL, Jacobsen AL, Pratt RB, Davis SD. 2016. Chaparral Shrub Hydraulic Traits, Size, and Life History Types Relate to Species Mortality during California’s Historic Drought of 2014. PLOS ONE 11:e0159145. https://doi.org/10.1371/journal.pone.0159145.
Vicca S, Gilgen AK, Camino Serrano M, Dreesen FE, Dukes JS, Estiarte M, Gray SB, Guidolotti G, Hoeppner SS, Leakey ADB, Ogaya R, Ort DR, Ostrogovic MZ, Rambal S, Sardans J, Schmitt M, Siebers M, van der Linden L, van Straaten O, Granier A. 2012. Urgent need for a common metric to make precipitation manipulation experiments comparable. New Phytologist 195(3):518–522. https://doi.org/10.1111/j.1469-8137.2012.04224.x.
Vicente-Serrano SM, Quiring SM, Peña-Gallardo M, Yuan S, Domínguez-Castro F. 2020. A review of environmental droughts: Increased risk under global warming? Earth-Science Reviews 201:102953. https://doi.org/10.1016/j.earscirev.2019.102953.
Wang C, Fu B, Zhang L, Xu Z. 2019. Soil moisture-plant interactions: an ecohydrological review. Journal of Soils and Sediments 19(1):1–9. https://doi.org/10.1007/s11368-018-2167-0.
Wang, E., Zhang, L., Cresswell, H., & Hickel, K. (2006). Comparison of Top- Down and Bottom-Up Models for Simulation of Water Balance as affected by Seasonality, Vegetation Type and Spatial Land Use.
West H, Quinn N, Horswell M. 2019. Remote sensing for drought monitoring impact assessment: Progress, past challenges and future opportunities. Remote Sensing of Environment 232:111291. https://doi.org/10.1016/j.rse.2019.111291.
Wossenyeleh BK, Kasa AS, Verbeiren B, Huysmans M. 2022. Drought propagation in the hydrological cycle in a semiarid region: a case study in the Bilate catchment. Ethiopia. Hydrogeology Journal 30(3):751–766. https://doi.org/10.1007/s10040-022-02459-8.
Yang, D., Shao, W., Yeh, P. J.-F., Yang, H., Kanae, S., & Oki, T. (2009). Impact of vegetation coverage on regional water balance in the nonhumid regions of China: Impact of vegetation on water. Water Resources Research, 45(7). https://doi.org/10.1029/2008WR006948
Yoon J-H, Wang S-YS, Gillies RR, Kravitz B, Hipps L, Rasch PJ. 2015. Increasing water cycle extremes in California and in relation to ENSO cycle under global warming. Nature Communications 6:8657. https://doi.org/10.1038/ncomms9657.
Acknowledgements
Funding for this work was provided by the US National Science Foundation (DEB-2016482) and the US Department of Energy, Office of Science, BER (SC-0020382). This work is based on and expands the second chapter of the Ph.D. thesis from Dr. Scot Parker, coauthor of this paper, and can be found at the following link https://escholarship.org/content/qt5xx4599n/qt5xx4599n.pdf?t=q8rw3i. All graphs, figures, and tables are originally produced.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
None declared.
Additional information
Author Contribution: SP, SDA, and MG conceived the study. SP and NMF gathered the data. LCR analyzed the data and interpreted the results. LCR produced the paper figures and created a repository to facilitate data sharing. SP contributed to the interpretation of the data. SDA and MG provided the funding for the project. LCR wrote the first draft of the papers, and all authors contributed to the final manuscript. LCR and SP contributed equally to this work.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Chavez Rodriguez, L., Parker, S., Fiore, N.M. et al. Impact of Drought on Ecohydrology of Southern California Grassland and Shrubland. Ecosystems 27, 106–121 (2024). https://doi.org/10.1007/s10021-023-00876-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-023-00876-8