
Abstract
Purpose
To investigate the effects of 60 min daily, short-term (STHA) and medium-term (MTHA) isothermic heat acclimation (HA) on the physiological and perceptual responses to exercise heat stress.
Methods
Sixteen, ultra-endurance runners (female = 3) visited the laboratory on 13 occasions. A 45 min sub-maximal (40% Wmax) cycling heat stress test (HST) was completed in the heat (40 °C, 50% relative humidity) on the first (HSTPRE), seventh (HSTSTHA) and thirteenth (HSTMTHA) visit. Participants completed 5 consecutive days of a 60 min isothermic HA protocol (target Tre 38.5 °C) between HSTPRE and HSTSTHA and 5 more between HSTSTHA and HSTMTHA. Heart rate (HR), rectal (Tre), skin (Tsk) and mean body temperature (Tbody), perceived exertion (RPE), thermal comfort (TC) and sensation (TS) were recorded every 5 min. During HSTs, cortisol was measured pre and post and expired air was collected at 15, 30 and 45 min.
Results
At rest, Tre and Tbody were lower in HSTSTHA and HSTMTHA compared to HSTPRE, but resting HR was not different between trials. Mean exercising Tre, Tsk, Tbody, and HR were lower in both HSTSTHA and HSTMTHA compared to HSTPRE. There were no differences between HSTSTHA and HSTMTHA. Perceptual measurements were lowered by HA and further reduced during HSTMTHA.
Conclusion
A 60 min a day isothermic STHA was successful at reducing physiological and perceptual strain experienced when exercising in the heat; however, MTHA offered a more complete adaptation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Exercising for a prolonged duration in a thermally stressful environment places the body under greater physiological and perceptual strain than when exercising in temperate conditions (Galloway and Maughan 1997; Tucker et al. 2004). The greater strain often results in reduced aerobic exercise performance (Ely et al. 2008) and may even result in serious heat illness, such as endotoxemia, heat exhaustion and heat stroke (Wendt et al. 2007). Heat acclimation (HA) has been proposed as one of the most effective interventions to be incorporated into an athlete’s training programme to reduce physiological strain and improve exercise performance in hot environmental conditions (Tyler et al. 2016). Currently, the optimal HA protocol is still unknown despite a large body of research manipulating the intensity and duration of exercise, the frequency of HA exposures, and the type of HA used (Tyler et al. 2016).
Repeated exposure to thermal stress can induce beneficial adaptations that include a reduced body temperature, increases in sweating sensitivity and rate, improved cardiovascular stability, lower perceptual strain, and improved exercise economy (Tyler et al. 2016). Up to 80% of adaptations occur in the first 4–7 days of exposure (short-term heat acclimation (STHA)); however, the magnitude of adaptation appears to be greater when medium-term (MTHA; 7–14 days) and long-term (LTHA; > 15 days) HA protocols are used (Tyler et al. 2016) and not all adaptations occur over the same time course. For example, heart rate adaptations typically occur well before improvements in performance and the sudomotor responses are observed (Periard et al. 2016; Tyler et al. 2016). For heat adaptations to occur, heat stress must induce physiological strain of a magnitude above an adaptation threshold (Taylor 2014) and the magnitude of the adaptation appears dependent on the extent and frequency of the thermal strain and impulse provided. It has been proposed that the threshold for adaptation may be the attainment and maintenance of a core temperature of ~ 38.5 °C because at this temperature, sudomotor and vasomotor thermoeffector responses are challenged and heat shock proteins are expressed (Fox et al. 1964; Gibson et al. 2015a, b). It may be difficult to reach and maintain such an internal temperature using traditional constant work HA protocols, but a controlled isothermic HA protocol overcomes this issue by ensuring that the target core temperature is reached through exercise and then maintained using passive (e.g. resting) and active (e.g. exercise) heat stress.
Another potential practical benefit of isothermic HA protocols is that thermal adaptations may be achieved with shorter exercise durations and lower exercise intensities than fixed-intensity HA protocols (Gibson et al. 2015a) and, therefore, they may be appropriate during the tapering phase in training (Tyler et al. 2016). Heat adaptations are lost at a rate of approximately ~ 2.5% per day when individuals are not exposed to heat and so HA should be undertaken as close to competition as possible to minimise de-acclimation (Daanen et al. 2018); however, an overly exerting HA protocol may compromise subsequent exercise performance and health due to over-activation of the hypothalamic–pituitary–thyroid axis (Reeve et al. 2019) (resulting in increased cortisol concentrations) and/or increased permeability of the gut (leading to the translocation of endotoxins such as lipopolysaccharide (LPS) in to the blood stream) (Lim et al. 2009). Within the current literature, an isothermic HA approach has been investigated either with an absolute increase in core temperature (Magalhaes et al. 2010) or the attainment of a set thermal strain (Garrett et al. 2012; Gibson et al. 2015a). A potential limitation to using a set increase in core body temperature (e.g. + 1 °C) is that as adaptation occurs and resting core temperature lowers, individuals may not be reaching a sufficient thermal strain to elicit HA adaptations. The attainment of a set thermal strain (e.g. 38.5 °C) ensures that as adaptation occurs an adaptation stimulus continues to be provided (Taylor 2014; Tyler et al. 2016). Recent isothermic HA literature has shown that a daily 90 min isothermic HA protocol offers an adequate stimulus for heat adaptation (Garrett et al. 2012; Gibson et al. 2015a), but such a duration may be problematic to integrate in to an athlete’s preparation. Shorter (30–60 min) constant work approaches can induce beneficial heat adaptations when exercise is maintained throughout to induce the strain (Houmard et al. 1990) and so it seems prudent to suggest that maintaining the strain for a similar duration using a less-intense exercise intensity, isothermic HA regimen would also be effective and desirable to tapering athletes. It is currently unknown whether such an approach provides enough time to induce physiological and perceptual adaptations as the time spent above the thermal impulse will be considerably reduced and the total time exposed to the heat is ~ 33.3% less than in previous isothermic HA protocols (Garrett et al. 2012, 2014; Gibson et al. 2015a).
The primary aim of this study, therefore, was to investigate whether a 60 min daily isothermic (core temperature of 38.5 °C) HA regimen would reduce the physiological and perceptual strain experienced when exercising in the heat. The secondary aim was to investigate if there was a time-course effect on the physiological and perceptual adaptations and whether MTHA was more effective than STHA. We hypothesised that the isothermic HA protocol would provide a sufficient thermal impulse to induce positive changes in physiological and perceptual measurements and that these adaptations would be more complete following MTHA than STHA.
Methods
Sixteen, non-heat acclimated, endurance runners (females = 3) participated. The mean (± SD) age, body mass, stature, body fat percentage and maximal work rate (Wmax) were 36.1 ± 9.1 years, 74.2 ± 9.4 kg, 176.1 ± 5.8 cm, 10.7 ± 4.9% and 302 ± 76 W, respectively. Before participation, all participants completed a health screening questionnaire and provided their fully informed, written consent to participate. The study was approved by the University of Roehampton’s ethical committee (LSC 18/228) and all procedures and protocols adhered to the guidelines of the World Medical Association (Declaration of Helsinki). Data were collected between March and April (Mean outside temperature ~ 6 °C) in the United Kingdom to avoid heat acclimatisation.
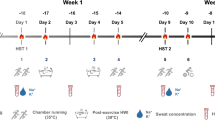
Experimental design (Fig. 1)

Experimental Schematic. Salivary cortisol (C) and venous blood (B) samples were collected pre- and post-HST on the first (HSTPRE), seventh (HSTSTHA) and thirteenth (HSTMTHA) visit. Between HSTPRE and HSTSTHA and again between HSTSTHA and HSTMTHA, participants completed 5 consecutive days of 60 min isothermic heat acclimation during which time rectal temperature (Tre) was elevated to, and maintained at ~ 38.5 °C
Participants undertook one preliminary visit (for the assessment of maximal power output) and 13 experimental visits. Participants performed a 45 min sub-maximal (40% Wmax) heat stress test (HST) on the first (HSTPRE), seventh (HSTSTHA) and thirteenth (HSTMTHA) experimental visit. Between HSTPRE and HSTSTHA and again between HSTSTHA and HSTMTHA, participants completed 5 consecutive days of isothermic heat acclimation (HA) (60 min each visit) during which time rectal temperature (Tre) was elevated to, and maintained at, ~ 38.5 °C. The environmental conditions were 40 °C and 50% relative humidity (rh) with no convective cooling for all sessions. Participants were instructed to avoid caffeine, alcohol, and strenuous exercise 24 h before all HSTs. HSTs and HA sessions were performed at the same time of day for each participant throughout the study to avoid the effects of circadian rhythm. Food intake was recorded for the 24 h prior to HSTPRE and participants were instructed to replicate this before HSTSTHA and HSTMTHA.
Preliminary testing
Stature (Harpenden Stadiometer, Holtain Ltd, UK) and body mass (Seca, Birmingham, UK) were recorded before Wmax was determined in ambient laboratory conditions (21 ± 1 °C and 55 ± 4% rh) using an incremental exercise test to volitional exhaustion (Kuipers et al. 1985), on a cycle ergometer (Monark 847E, Vansbro, Sweden). During this test, participants initially cycled at 100 watts (W) for 5 min, thereafter, work was increased by 50 W every 2.5 min until heart rate (HR) reached 160 b min−1, once reached, work was increased by 25 W every 2.5 min until exhaustion. The maximum work rate was calculated using the equation of Kuipers et al. (1985): Wmax = Wcom + ((t/150) × ΔW) [Wcom is the last work rate completed; t is the duration (in seconds) of the final, uncompleted, stage; ΔW is the final load increment (typically 21 W)]. Percentage of body fat (%) was measured using whole body air displacement plethysmography method (BodPod, Cosmed, Italy).
Heat stress tests (HSTs)
Upon arrival, a mid-flow urine sample was provided to measure urine specific gravity (USG), using a hand-held pen refractometer (Atago, pen refractometer, PEN-Urine S.G, Tokyo, Japan). All participants reported to the laboratory euhydrated (USG < 1.020). Following this, participants self-recorded nude body mass (BM) and self-inserted a rectal thermistor (REC-U-VL3-0, Grant Instruments (Cambridge) Ltd., UK) ~ 10 cm past the anal sphincter before affixing a HR monitor (Polar Electro Ltd., Kempele, Finland) to their upper torso. The rectal thermistor was connected to a portable data logger (Squirrel 2020 Series, Grant Instruments, (Cambridge) Ltd., UK). Skin temperature (Tsk) was recorded continuously using wireless Thermochron iButton skin temperature data loggers (DS1922L, Thermochron iButton, USA). iButtons were attached using transparent adhesive dressing (Tegaderm, 3M Health Care, St Paul, MN) and waterproof tape (Transpore, 3M Health Care, St Paul, MN) to the sternal notch, forearm, thigh and calf muscle on the right side of the body. Mean-weighted Tsk was calculated (Ramanathan 1964) and mean body temperature (Tbody) was estimated (Stolwijk and Hardy 1966). Due to methodological issues, Tsk, and as a result Tbody, data were collected from only seven participants.
Participants entered the controlled environment (40 °C, 50% rh) and rested for 2 min before baseline measurements of HR, Tre, thermal sensation (TS (Young et al. 1987)), and thermal comfort (TC (Gagge et al. 1967)) were recorded and rated. Once baseline measurements were taken ,participants cycled at 40% of their Wmax for 45 min, during which HR, Tre, ratings of perceived exertion (RPE (Borg 1982)), TS and TC were measured every 5 min. One min expired air samples were collected at 14 min, 29 min and 44 min using the Douglas bag method and subsequently analysed (1400 series, Servomex, East Sussex, UK; Harvard Dry Gas Meter, Harvard Ltd., Kent, UK). To prevent further, uncontrolled per-cooling, participants drank warm (~ 37 °C) water ad libitum. The water was stored in the environmental chamber and the volume consumed was recorded. Once final measurements were recorded, participants exited the controlled environment and self-recorded a final nude BM measurement after they had towel dried. Sweat losses were determined from trial changes in BM, subtracting the weight of urine produced and adding fluid consumed (ml) during the trial.
Heat acclimation (HA)
Participants initially repeated the same procedures as undertaken in HSTPRE. After USG and BM were measured and a rectal thermistor was self-inserted, iButtons were placed on the same four sites and a HR strap was fitted before entering the controlled environment. Once baseline Tre, HR, TS, and TC measurements were recorded after 2 min rest, participants were instructed to reach a target Tre of ~ 38.5 °C as quickly as possible and self-selected their workload and cadence accordingly. Once the target Tre had been attained, the distance cycled, and the time taken to reach the target Tre were recorded, as was the time spent at or above it. Participants then sat for the remainder of the 60 min session unless Tre fell to 38.55 °C, at which point participants resumed cycling to increase Tre. During the 60 min, HR, Tre, RPE, TS, and TC were recorded every 5 min and on completion of the session, nude BM was recorded post-session to estimate sweat losses. Participants drank warm water ad libitum to prevent uncontrolled per-cooling (~ 37 °C). The water was stored in the environmental chamber and the volume consumed was recorded. Due to methodological issues, Tsk, and as a result Tbody, data were collected from only six participants. The peak intra-session strain was calculated as the peak Tre minus the starting Tre during each HA session. Peak cumulative strain was then calculated as the total strain for the 5 (STHA) and 10 day (MTHA) HA regimens.
Salivary cortisol sample collection and analyses
Saliva samples were collected from each participant immediately upon awakening on 2 of the 5 days before HSTPRE (B1 and B2) to establish normal basal concentrations of cortisol and then again immediately before and after each HST. Saliva was collected by each participant chewing an absorbent swab (Salivette Cortisol, Code Blue, Sarstedt, Leicester, UK) then inserting it into a Salivette tube. All samples were centrifuged at 1000g for 2 min with the resulting saliva sample transferred into 2 ml Eppendorf tubes and stored in a freezer at − 80 °C until analysis. Salivary cortisol levels were determined with a high-sensitivity (0.007 µg dL−1) salivary cortisol enzyme-linked immunosorbent assay (Salumetrics, State College, PA, USA) as per the manufacturer’s instructions.
Plasma lipopolysaccharide sample collection and analyses
Venous blood was drawn from nine participants, pre- and post-HST, using a butterfly cannula that drained directly into a sterile EDTA tube before being centrifuged at 3000 rpm for 10 min at 4 °C. Plasma was extracted using pyrogen-free pipette tips into pyrogen-free microtubes (Eppendorf, Hamburg, Germany) before being frozen at − 80 °C. Plasma concentrations of LPS were analysed using a high-sensitivity (0.04 EU ml−1) chromogenic limulus amoebocyte lysate end-point assay kit (Hycult Biotechnology b.v., Uden, Netherlands). Plasma samples were thawed and brought to room temperature before being diluted by 1000 times with endotoxin-free water. Fifty microliters of each sample were then transferred into the wells of pyrogen-free microplate in duplicates, followed by 50 µL of bacterial endotoxin (LAL) reagent. Optical density of the reaction was read with a microplate reader (Thermo Scientific Multiskan EX) at a wavelength of 405 nm.
Statistical analyses
Data were analysed using SPSS (version 26, SPSS Inc.). One-way and two-way repeated measures ANOVAs were performed to determine differences between time points and trials in HSTPRE, HSTSTHA, and HSTMTHA, and in the first, middle, and final HA session (HA1, HA5 and HA10). Where the assumption of sphericity had been violated, the degrees of freedom were corrected using the Greenhouse–Geisser estimate. Where significant outcomes were present, post hoc tests with Bonferroni corrections were performed. The alpha level was P ≤ 0.05. Cohen’s d effect sizes were calculated for post-preload data and interpreted as follows: small effect: d = 0.2 to < 0.5; medium effect: 0.5 to < 0.8; large effect: d ≥ 0.8 (Cohen 1988). Data are presented as mean ± SD.
Results
Heat acclimation
There was a main effect for trial of distance (P = 0.025) but not duration (P = 0.108) cycled before the attainment of the target Tre (38.5 °C). The distance was similar between HA1 (18 ± 4 km) and HA5 (21 ± 4 km; P = 0.240), but greater in HA10 (23 ± 5 km) compared to HA1 (P = 0.018) and HA5 (P = 0.026). It took longer to reach the target Tre in HA10 (41.6 ± 6.4 min) than in HA1 (35.7 ± 6.1 min), but this was not statistically significant. The duration was also similar between HA1 and HA5 (36.9 ± 6.8 min) and between HA5 and HA10. Despite differences in the distance cycled, the thermal impulse to 38.5 °C was not different between HA1 (0.032 ± 0.006 °C min−1), HA5 (0.033 ± 0.006 °C min−1), and HA10 (0.036 ± 0.007 °C min−1) (Main effect for trial: P = 0.200). The peak cumulative thermal strain and impulse provided by the STHA and MTHA interventions were 9.85 ± 1.35 °C and 0.033 ± 0.0042 °C min−1 and 20.52 ± 2.26 °C and 0.034 ± 0.004 °C min−1, respectively. There were no changes in the classic physiological or perceptual markers of heat adaptation measured before or during HA1, HA5, and HA10 (Table 1).
Heat stress test–physiological data (Table 2)
There was a main effect for trial for resting Tre (P < 0.001) and Tbody (P < 0.001) with both lower in HSTSTHA (P < 0.001, d = 1.2; P < 0.006, d = 1.6) and HSTMTHA (both P < 0.001, d = 1.3 and 1.6, respectively) than HSTPRE, whereas no difference in resting Tsk was measured between HSTs (P = 0.243, d = 0.3–0.7). Resting HR was similar between HSTPRE and both HSTSTHA (P = 0.244, d = 0.6) and HSTMTHA (P = 0.113, d = 0.7), despite a significant main effect (P = 0.042). There were no differences in resting Tre, Tbody, or HR between HSTSTHA and HSTMTHA (all P > 0.99, d < 0.2).
The thermoregulatory and cardiovascular strain experienced was lower during (main effects: P < 0.007) and at the end (main effects: P < 0.005) of the HSTs performed following HA (Figs. 2 and 3). Mean Tre, Tsk and Tbody were higher during HSTPRE compared to HSTSTHA (P = 0.002, d = 1.1; P = 0.026, d = 1.3; P = 0.007, d = 1.5, respectively) and HSTMTHA (P < 0.001, d = 1.4; P = 0.034, d = 1.3; P = 0.005, d = 1.5, respectively), but were similar in HSTSTHA and HSTMTHA (P = 0.223, d = 0.6; P > 0.99, d = 0.1; P = 0.692, d = 0.5, respectively). The rise in Tre over the exercise bout was similar in all three HSTs (P = 0.292). Tre was lower at all time-points in HSTMTHA compared to HSTPRE (P < 0.05) and for the first 40 min in HSTSTHA compared to HSTPRE (all P < 0.05). Tre was similar in HSTSTHA and HSTMTHA at each time point (all P > 0.05) (Fig. 2). Mean HR was higher in HSTPRE than HSTSTHA (P = 0.004, d = 0.8) and HSTMTHA (P = 0.004, d = 1.1) but was similar between HSTSTHA and HSTMTHA (P = 0.074, d = 0.6). HR was higher in HSTSTHA than HSTMTHA at 10 (P = 0.034), 15 (P = 0.018), and 20 min (P = 0.035) during the HST (Fig. 3).

Rectal temperature (Tre) on each time point during HSTPRE, HSTSTHA, and HSTMTHA. There were main effects of trial (P < 0.001) and time (P < 0.001) for Tre, *Significant (P < 0.05) difference between HSTPRE and both HSTSTHA and HSTMTHA. Data mean ± SD

Heart rate during HSTPRE, HSTSTHA, and HSTMTHA. There were main effects of trial (P < 0.001) and time (P < 0.001) for HR. †Significant (P < 0.05) difference between HSTPRE and HSTSTHA. *Significant (P < 0.05) difference between HSTPRE and HSTMTHA. #Significant (P < 0.05) difference between HSTMTHA and HSTSTHA. Data mean ± SD
After 45 min of exercise, Tre was not different between HSTPRE compared to HSTSTHA (P = 0.081, d = 0.7) but was lower in HSTMTHA compared to HSTPRE (P < 0.001, d = 1.1), there were no differences between HSTSTHA and HSTMTHA (P = 0.488, d = 0.4). Tsk and Tbody was lower in both HSTSTHA (P = 0.044, d = 1.2; P = 0.048, d = 1.3) and HSTMTHA (P = 0.018, d = 1.0; P = 0.008, d = 1.6) compared to HSTPRE but no differences were seen between HSTSTHA and HSTMTHA (all P > 0.99, d = 0.41, d = 0.3). Final HR was higher in HSTPRE than HSTSTHA (P = 0.012, d = 0.6) and HSTMTHA (P = 0.003, d = 1.0) but no differences were observed between HSTSTHA and HSTMTHA (P = 0.065, d = 0.7). Sweat rate was similar in HSTPRE and HSTSTHA (P > 0.99), but higher during HSTMTHA than HSTPRE (P < 0.001, d = 1.0) and HSTSTHA (P < 0.001, d = 0.8). Fluid consumption was similar in all trials (main effect: P = 0.827) and so the percentage dehydration differed (main effect: P < 0.001), being greater in HSTMTHA compared to both HSTSTHA (P < 0.001, d = 1.1) and HSTMTHA (P < 0.001, d = 0.8). V̇O2 and respiratory exchange ratio (RER) were similar between trials (main effect trial: P = 0.094; P = 0.089) and did not change over time (P = 0.515, P = 0.116).
Baseline cortisol concentrations and the concentrations prior to all HSTs were similar among the trials (P = 0.11). The coefficient of variation between B1 and B2 was 15 ± 6%. The within trial increase was different between trials (P < 0.001) being greater in HSTPRE (0.53 ± 0.40 µg dL−1; + 339 ± 284%) compared to HSTSTHA (0.08 ± 0.29 µg dL−1, P < 0.001, d = 1.1; + 90 ± 183%) and HSTMTHA (0.04 ± 0.35 µg dL−1, P = 0.003, d = 1.1; + 93 ± 181%). There were no differences in the within trial change in cortisol between HSTSTHA and HSTMTHA (P > 0.99, d = 0.2). There was a large degree of variation in the percentage change within trials in HSTPRE ( – 37 to + 730%), HSTSTHA ( – 62 to 550%), and HSTMTHA ( – 63 to 580%); however, the number of participants who had an increase in cortisol concentration in their post-sample compared to their pre-sample was higher in HSTPRE (10/12) than HSTSTHA (6/12) and HSTMTHA (6/12).
Plasma LPS levels were similar between trials before (HSTPRE: 1.46 ± 0.57 EU ml−1; HSTSTHA: 1.49 ± 0.54 EU ml−1; HSTMTHA: 1.52 ± 1.36 EU ml−1, P = 0.926) and after 45 min of exercise (HSTPRE: 2.00 ± 1.65 EU ml−1; HSTSTHA: 1.74 ± 0.85 EU ml−1; HSTMTHA: 1.72 ± 0.82 EU ml−1, P = 0.869). Within HSTs, the mean change was similar (P = 0.420)—0.54 ± 1.17 EU ml−1, 0.25 ± 1.01 EU ml−1 and 0.05 ± 0.38 EU ml−1, for HSTPRE, HSTSTHA and HSTMTHA, respectively. Similar to the cortisol response, there was a large degree of variation in the percentage change within trials in HSTPRE (+ 50 – 218%), HSTSTHA (+ 49 – 213%), and HSTMTHA (+ 29 – 402%), but unlike cortisol, LPS concentrations increased in all participants in all trials.
Heat stress test–perceptual measurements
There were main effects of trial and time for TS, TC, and RPE (all P < 0.002), with all increasing progressively throughout the HST (all P < 0.001), there was an interaction effect for TC and RPE (all P < 0.006) but not for TS (P = 0.248). Data are reported in Table 2. Resting TC was unaffected by HA (P = 0.487) but resting TS was different between trials (P < 0.001) being lower in HSTSTHA (P = 0.014) and HSTMTHA (P = 0.002). All the reduction had occurred within 5 days of HA with no differences between HSTSTHA and HSTMTHA (P > 0.99). During exercise, mean TS, TC, and RPE were different between trials (P < 0.001). TS and TC were both higher in HSTPRE than HSTSTHA (P < 0.001, P = 0.037) and HSTMTHA (P < 0.002, P < 0.001). Both were further reduced from HSTSTHA to HSTMTHA (TS: P = 0.031; TC: P = 0.030) (Fig. 4). Mean RPE was not different between HSTPRE and HSTSTHA (P = 0.456), but was lower in HSTMTHA compared to HSTPRE (P = 0.006) and HSTSTHA (P = 0.015). At the end of exercise, TS, TC, and RPE were all lower after HSTSTHA (P < 0.001, P = 0.039, P = 0.037) and HSTMTHA (all P < 0.001) compared to HSTPRE. Thermal sensations were rated lower at the end of HSTMTHA than HSTSTHA (P = 0.046; d = 0.56) but neither TC (P = 0.083; d = 0.52) nor RPE (P = 0.120; d = 0.13) were rated differently between HSTSTHA and HSTMTHA.

a Thermal sensation and b thermal comfort were recorded before exercise and every 5 min intervals during HSTPRE, HSTSTHA, and HSTMTHA. †Significant (P < 0.05) difference between HSTPRE and HSTSTHA. *Significant (P < 0.05) difference between HSTPRE and HSTMTHA. #Significant (P < 0.05) difference between HSTMTHA and HSTSTHA. Data mean ± SD
Discussion
The present study investigated whether a daily 60 min isothermic HA protocol provided a sufficient thermal impulse to induce the physiological and perceptual adaptations and whether there was a time-course response when comparing STHA and MTHA. The main findings of the present study are (1) an isothermic STHA protocol provides a sufficient cumulative thermal strain (9.85 ± 1.35 °C) to effectively lower physiological and perceptual strain and (2) MTHA induces further beneficial heat adaptations but only to sweat losses, final thermoregulatory strain, and perceptions of exertion, thermal strain and comfort, despite providing double the cumulative thermal strain (20.52 ± 2.26 °C).
Physiological adaptations
The isothermic HA protocol was successful at lowering resting Tre ( − 0.38 ± 0.26 °C) and HR (8 ± 16 b.min−1) after 5 days of heat exposure. A further 5 days of heat acclimation (MTHA) did not elicit any further resting adaptations in Tre ( − 0.02 ± 0.31 °C from STHA) or HR (0 ± 9 b.min−1 from STHA). Both these responses are in accordance with previous findings that reported that these adaptations occurred at a rapid rate and were not further enhanced after longer exposure periods (Tyler et al. 2016). A lower resting Tre is an important indicator of a successful HA protocol, because it can delay the attainment of high-core body temperatures often reported to limit exercise capacity in the heat (Gonzalez-Alonso et al. 1999; Tucker et al. 2004). Our observed reductions are greater than mean changes reported in a recent meta-analysis (STHA: − 0.17 ± 0.12 °C; MTHA: − 0.17 ± 0.1 °C) (Tyler et al. 2016) and those reported in previous 5 day isothermic-controlled studies (Garrett et al. 2012, 2014; Neal et al. 2016). The more substantial reductions in physiological markers of heat acclimation may be due to the lower training status of our participants compared to those presented in the literature, where comparably smaller reductions in resting Tre were observed in more highly trained (mean peak power output of 375 ± 31 W) individuals (Garrett et al. 2012, 2014; Neal et al. 2016). This is likely because highly trained individuals have already developed some thermal adaptations from their long-term training history (e.g. a greater evaporative heat loss capacity and a decrease in resting core temperature) (Cheung and McLellan. 1998) which would limit the potential for a HA protocol to induce further adaptations.
The reduction in resting Tre coupled with similar (HSTPRE: 1.85 ± 0.57 °C, HSTSTHA: 1.84 ± 0.65 °C) and lower (1.66 ± 0.50 °C) delta changes in Tre after 5 and 10 days of HA, respectively, resulted in reduced thermal strain throughout HSTSTHA and HSTMTHA compared to HSTPRE. HA has been shown to reduce the oxygen cost of exercise in the heat (Lorenzo et al. 2010); however, this is not always reported when using cycling as the mode of exercise, where the utilisation of the upper body muscles are minimal. This might explain why V̇O2 and RER were not altered following either STHA or MTHA in the present study. Due to the lack of change in efficiency, it seems reasonable to assume that the reduced thermal strain was due to an increase in heat loss mechanisms, as sweat rate increased after 10 days of HA, facilitating a greater heat loss through evaporative cooling. Our data support previous findings, that found sudomotor responses took longer to occur than other adaptations (Tyler et al. 2016). While local methods to access sweat rates were not used in the current study, whole body sweat rates were increased following 10 (MTHA) but not 5 (STHA) days of heat exposure. Previous results have found that HA increases sweat rate, as a result of an earlier onset of sweating at a lower core temperature and a more pronounced sudomotor thermosensitivity (Buono et al. 2018).
It is well known that cardiovascular strain can limit prolonged exercise performed under heat stress (Périard et al. 2011) and a reduced cardiovascular strain is a classic marker of an effective HA regimen (Tyler et al. 2016). In the present study, resting HR was unaffected by HA; however, the mean exercising and end of exercise HR was reduced following STHA. There was no additional benefit of a longer exposure period (MTHA) on resting and end of exercise HR; however, longer exposure time lowered HR response at certain time points during exercise. Adaptations in HR occur rapidly and are often complete within 7 days (Périard et al. 2015; Tyler et al. 2016) and data from the present study support this. With physiological measurements including stroke volume, skin blood flow and plasma volume not measured in the current study, identifying the factor that influenced this response can only be speculated. It has been previously suggested that the improved cardiovascular stability from HA is achieved through an increase in plasma volume, better maintained fluid balance, and enhanced sweating and skin blood flow responses (Périard et al. 2015, 2016; Tyler et al. 2016).
Cortisol is often used as a marker of physical and psychological strain and, as observed elsewhere (e.g. Silva et al. (2019)), cortisol concentrations increased following an initial bout of exercise in the heat (HSTPRE). Following HA, we observed an attenuated increase—data which are in contrast to Costello et al. (2018) and Garrett et al. (2009) but in agreement with Watkins et al. (2008) who reported reductions in the session increase in cortisol after 7 days of HA. Costello et al. (2018) did not report statistical reductions in cortisol following HA but noted that there was a “trend” for the increase to be lower post-HA and so it appears that the cortisol response to exercise in the heat may be sensitive to heat adaptation. While cortisol may be a potential marker of heat adaptation, due to the variation within and between investigations, it is advisable to use the more established variables (e.g. resting core body temperature and heart rate) at present. In contrast to the cortisol response, neither STHA nor MTHA altered the LPS response to the HST. Similar observations have been reported previously by Guy et al. (2016) who also observed no effect of HA on the inflammatory response to exercise. Guy et al. (2016) postulated that their use of non-consecutive heat exposures may have provided an inadequate strain to trigger a systemic inflammatory response, but we used a more intensive HA regimen and saw comparable LPS responses. These limited data suggest that MTHA may offer some protection against endotoxemia in healthy individuals through the reduction of physiological strain and that the MTHA protocols investigated to date do not trigger an endotoxic response themselves.
Perceptual adaptations
Participants felt more thermally comfortable and reported lower thermal sensations after 5 days of HA (STHA). An additional 5 days of HA (MTHA) had an additional beneficial effect on final thermal sensation. Both Tre and Tsk are key drivers of thermal perceptions, but in the present study, neither Tre nor Tsk continued to decline with longer exposure and so these observations do not explain why these perceptual responses continued to be improved over the HA regimen. An increase in perceived exertion (RPE) and thermal perceptions (TS, TC) have been reported to play a role in downregulating self-paced time trials (TT) when performed under heat stress to reduce the rate of heat storage well before hyperthermia is present (Tucker et al. 2004). Although not measured, it seems reasonable to suggest that a lower perceived exertion and improved perceptions of thermal strain and comfort would enable participants to select a higher exercise intensity and improve subsequent performance. Our data suggest that the 5 day isothermic STHA regimen provided a sufficient thermal stimulus to improve perceptions of strain, but MTHA (10 days) offered further benefit and so is the preferred approach.
Limitations and practical recommendations
We cannot exclude the possibility that there was a training effect that may have occurred during HA as there was not a passive control group; however, in previous studies that included a control group, there was no reported training benefit in performance outcomes (Lorenzo et al. 2010). Additionally, the intensity and duration of exercise used in the present study was substantially lower than the participants were used to as all participants were about to take part in the Marathon des Sables, a 250 km foot-race across the Sahara desert.
Identifying an effective HA protocol that reduces the risk of overexerting an athlete so close to competition, while optimising thermal adaptations, is of current focus while athletes prepare for upcoming sporting events, including the Olympic Games in Tokyo, 2020. We did not measure whether the isothermic STHA and MTHA regimens improved subsequent exercise performance or reduced heat illness risk, but we speculate that progressive improvements would have been observed as a result of the reductions in physiological and perceptual strain as has been reported previously (Lorenzo et al. 2010). We suggest using an isothermal HA regimen during the taper phase of an athlete’s schedule and highlight that although 5 days is sufficient to induce meaningful beneficial adaptations to heat, 10 days is more effective and so should be used when possible.
Conclusion
A 5 day 60 min isothermic HA regimen provides a sufficient thermal stimulus to elicit beneficial adaptations to reduce physiological and perceptual strain during subsequent exercise in the heat, despite providing a lower cumulative thermal strain than commonly observed in the HA literature. Most of the beneficial adaptations occurred within the STHA time-frame; however, an additional 5 days of HA (MTHA) induced further thermoregulatory, sudomotor, and perceptual adaptations and so isothermic MTHA is preferred over isothermic STHA when possible.
Abbreviations
- HA:
-
Heat acclimation
- HR:
-
Heart rate
- HST:
-
Heat stress test
- LPS:
-
Lipopolysaccharide
- MTHA:
-
Medium-term heat acclimation
- RPE:
-
Rating of perceived exertion
- STHA:
-
Short-term heat acclimation
- Tbody :
-
Mean body temperature
- TC:
-
Thermal comfort
- T re :
-
Rectal temperature
- TS:
-
Thermal sensation
- T sk :
-
Skin temperature
- W :
-
Watts
References
Borg GA (1982) Psychophysical bases of perceived exertion. Med Sci Sports Exerc 14:377–381
Buono MJ, Kolding M, Leslie E et al (2018) Heat acclimation causes a linear decrease in sweat sodium ion concentration. J Therm Biol 71:237–240
Cheung SS, McLellan TM (1998) Heat acclimation, aerobic fitness, and hydration effects on tolerance during uncompensable heat stress. J Appl Physiol (1985) 84:1731–1739. https://doi.org/10.1152/jappl.1998.84.5.1731
Cohen J (1988) Statistical power analysis for the behavioral sciences. Lawrence Erlbaum Associates, Hillsdale
Costello JT, Rendell RA, Furber M et al (2018) Effects of acute or chronic heat exposure, exercise and dehydration on plasma cortisol, IL-6 and CRP levels in trained males. Cytokine 110:277–283
Daanen HAM, Racinais S, Periard JD (2018) Heat acclimation decay and re-induction: a systematic review and meta-analysis. Sports Med 48:409–430. https://doi.org/10.1007/s40279-017-0808-x
Ely MR, Martin DE, Cheuvront SN, Montain SJ (2008) Effect of ambient temperature on marathon pacing is dependent on runner ability. Med Sci Sports Exerc 40:1675–1680. https://doi.org/10.1249/MSS.0b013e3181788da9
Fox RH, Goldsmith R, Hampton IF, Lewis HE (1964) The nature of the increase in sweating capacity produced by heat acclimatization. J Physiol 171:368–376. https://doi.org/10.1113/jphysiol.1964.sp007382
Gagge AP, Stolwijk JA, Hardy JD (1967) Comfort and thermal sensations and associated physiological responses at various ambient temperatures. Environ Res 1:1–20
Galloway SD, Maughan RJ (1997) Effects of ambient temperature on the capacity to perform prolonged cycle exercise in man. Med Sci Sports Exerc 29:1240–1249
Garrett AT, Goosens NG, Rehrer NJ, Patterson MJ, Cotter JD (2009) Induction and decay of short-term heat acclimation. Eur J Appl Physiol 107:659–670. https://doi.org/10.1007/s00421-009-1182-7
Garrett AT, Creasy R, Rehrer NJ, Patterson MJ, Cotter JD (2012) Effectiveness of short-term heat acclimation for highly trained athletes. Eur J Appl Physiol 112:1827–1837. https://doi.org/10.1007/s00421-011-2153-3
Garrett AT, Goosens NG, Rehrer NJ et al (2014) Short-term heat acclimation is effective and may be enhanced rather than impaired by dehydration. Am J Hum Biol 26:311–320. https://doi.org/10.1002/ajhb.22509
Gibson OR, Turner G, Tuttle JA, Taylor L, Watt PW, Maxwell NS (2015a) Heat acclimation attenuates physiological strain and the HSP72, but not HSP90α, mRNA response to acute normobaric hypoxia. J Appl Physiol (1985) 119:889–899. https://doi.org/10.1152/japplphysiol.00332.2015
Gibson OR, Mee JA, Tuttle JA, Taylor L, Watt PW, Maxwell NS (2015b) Isothermic and fixed intensity heat acclimation methods induce similar heat adaptation following short and long-term timescales. J Therm Biol 49–50:55–65. https://doi.org/10.1016/j.jtherbio.2015.02.005
Gonzalez-Alonso J, Teller C, Andersen SL, Jensen FB, Hyldig T, Nielsen B (1999) Influence of body temperature on the development of fatigue during prolonged exercise in the heat. J Appl Physiol (1985) 86:1032–1039. https://doi.org/10.1152/jappl.1999.86.3.1032
Guy JH, Pyne DB, Deakin GB, Miller CM, Edwards AM (2016) Acclimation training improves endurance cycling performance in the heat without inducing endotoxemia. Front Physiol 7:318. https://doi.org/10.3389/fphys.2016.00318
Houmard JA, Costill DL, Davis JA, Mitchell JB, Pascoe DD, Robergs RA (1990) The influence of exercise intensity on heat acclimation in trained subjects. Med Sci Sports Exerc 22:615–620
Kuipers H, Verstappen FT, Keizer HA, Geurten P, van Kranenburg G (1985) Variability of aerobic performance in the laboratory and its physiologic correlates. Int J Sports Med 6:197–201. https://doi.org/10.1055/s-2008-1025839
Lim CL, Pyne D, Horn P et al (2009) The effects of increased endurance training load on biomarkers of heat intolerance during intense exercise in the heat. Appl Physiol Nutr Metab 34:616–624. https://doi.org/10.1139/H09-021
Lorenzo S, Halliwill JR, Sawka MN, Minson CT (2010) Heat acclimation improves exercise performance. J Appl Physiol (1985) 109:1140–1147. https://doi.org/10.1152/japplphysiol.00495.2010
Magalhaes FC, Passos RL, Fonseca MA et al (2010) Thermoregulatory efficiency is increased after heat acclimation in tropical natives. J Physiol Anthropol 29:1–12
Neal RA, Massey HC, Tipton MJ, Young JS, Corbett J (2016) Effect of permissive dehydration on induction and decay of heat acclimation, and temperate exercise performance. Front Physiol 7:564. https://doi.org/10.3389/fphys.2016.00564
Periard JD, Cramer MN, Chapman PG, Caillaud C, Thompson MW (2011) Cardiovascular strain impairs prolonged self-paced exercise in the heat. Exp Physiol 96:134–144. https://doi.org/10.1113/expphysiol.2010.054213
Periard JD, Racinais S, Sawka MN (2015) Adaptations and mechanisms of human heat acclimation: applications for competitive athletes and sports. Scand J Med Sci Sports 25(Suppl 1):20–38. https://doi.org/10.1111/sms.12408
Periard JD, Travers GJS, Racinais S, Sawka MN (2016) Cardiovascular adaptations supporting human exercise-heat acclimation. Auton Neurosci 196:52–62
Ramanathan NL (1964) A new weighting system for mean surface temperature of the human body. J Appl Physiol 19:531–533. https://doi.org/10.1152/jappl.1964.19.3.531
Reeve T, Gordon R, Laursen PB, Lee JKW, Tyler CJ (2019) Impairment of cycling capacity in the heat in well-trained endurance athletes after high-intensity short-term heat acclimation. Int J Sports Physiol Perform. https://doi.org/10.1123/ijspp.2018-0537
Silva RPM, Barros CLM, Mendes TT et al (2019) The influence of a hot environment on physiological stress responses in exercise until exhaustion. PLoS ONE 14:e0209510. https://doi.org/10.1371/journal.pone.0209510
Stolwijk JA, Hardy JD (1966) Partitional calorimetric studies of responses of man to thermal transients. J Appl Physiol 21:967–977. https://doi.org/10.1152/jappl.1966.21.3.967
Taylor NA (2014) Human heat adaptation. Compr Physiol 4:325–365. https://doi.org/10.1002/cphy.c130022
Tucker R, Rauch L, Harley YX, Noakes TD (2004) Impaired exercise performance in the heat is associated with an anticipatory reduction in skeletal muscle recruitment. Pflugers Arch 448:422–430. https://doi.org/10.1007/s00424-004-1267-4
Tyler CJ, Reeve T, Hodges GJ, Cheung SS (2016) The effects of heat adaptation on physiology, perception and exercise performance in the heat: a meta-analysis. Sports Med 46:1699–1724. https://doi.org/10.1007/s40279-016-0538-5
Watkins AM, Cheek DJ, Harvey AE, Blair KE, Mitchell JB (2008) Heat acclimation and HSP-72 expression in exercising humans. Int J Sports Med 29:269–276. https://doi.org/10.1055/s-2007-965331
Wendt D, van Loon LJ, Lichtenbelt WD (2007) Thermoregulation during exercise in the heat: strategies for maintaining health and performance. Sports Med 37:669–682
Young AJ, Sawka MN, Epstein Y, Decristofano B, Pandolf KB (1987) Cooling different body surfaces during upper and lower body exercise. J Appl Physiol (1985) 63:1218–1223. https://doi.org/10.1152/jappl.1987.63.3.1218
Acknowledgements
We express our gratitude to the participants for their efforts throughout the study.
Author information
Authors and Affiliations
Contributions
JNM, SJT, RWAM, and CJT conceived and designed research. JNM, FMB, FC, MRN, RJFH Gordon, TCR, and CJT conducted experiments. JNM, TCR, and CJT analysed the blood samples. JNM, SJT, and CJT analysed data. JNM. SJT, and CJT wrote the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Communicated by Narihiko Kondo.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Moss, J.N., Bayne, F.M., Castelli, F. et al. Short-term isothermic heat acclimation elicits beneficial adaptations but medium-term elicits a more complete adaptation. Eur J Appl Physiol 120, 243–254 (2020). https://doi.org/10.1007/s00421-019-04269-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-019-04269-5