
Abstract
The gut microbiota, housing trillions of microorganisms within the gastrointestinal tract, has emerged as a critical regulator of host health and homeostasis. Through complex metabolic interactions, these microorganisms produce a diverse range of metabolites that substantially impact various physiological processes within the host. This review aims to delve into the intricate relationships of gut microbiota-derived metabolites and their influence on the host homeostasis. We will explore how these metabolites affect crucial aspects of host physiology, including metabolism, mucosal integrity, and communication among gut tissues. Moreover, we will spotlight the potential therapeutic applications of targeting these metabolites to restore and sustain host equilibrium. Understanding the intricate interplay between gut microbiota and their metabolites is crucial for developing innovative strategies to promote wellbeing and improve outcomes of chronic diseases.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Gut microbiota–host immune interaction with metabolites
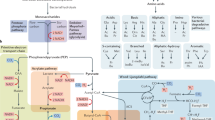
The human body hosts a wide array of commensal microorganisms, including bacteria, fungi, viruses, archaea, and protozoa [1,2,3]. Among this consortium, bacteria take precedence over other eukaryotic symbionts and have been extensively studied for their interactions with the host. The dominant bacterial phyla, Bacteroidetes and Firmicutes, alongside less abundant phyla such as Proteobacteria, Actinobacteria, and Verrucomicrobia, collectively constitute over 90% of microbiota in the gut. [4]. Initially recognized for their role in extracting energy from food, the metabolic contributions of gut microbiota have expanded to include fat storage and the regulation of glucose metabolism [5, 6]. Over the past two decades, advances in sequencing technologies have further revealed the association of gut microbiota with host homeostasis [7, 8]. The homeostatic gut microbiota fortifies the epithelial barrier against microbial invasion and promotes tolerance to innocuous luminal components, including the microbes themselves [9, 10]. However, disruptions in the gut microbiota consortium through broad-range antibiotic treatments render the host more vulnerable to inflammation [11, 12]. Research on germ-free mice has shown that the absence of gut microbiota impairs gut barrier function and renders animals susceptible to microbial invasions [13, 14]. As the essential role of gut microbiota in host immune homeostasis has been realized, the importance of understanding the mechanisms by which the host communicates with the gut microbiota has become pertinent. Recent advancements in multi-omics techniques have provided examples of host–microbiota interaction via diverse microbial products, including metabolites. Gut microbiota-derived metabolites, predominantly short-chain fatty acids (SCFAs), intermediate bile acid (BA) metabolites, amino acid-derived metabolites, and membrane-associated lipids, possess characteristics enabling communication with the immune system of their host (Table 1). These metabolites exert their biological activity through specific recognition by cognate receptors, primarily mediated by members of G-protein-coupled receptors (GPRs) [42]. GPRs exhibit distinct expression across various cell types, and the response to the same metabolite can vary based on specific cell roles [43]. Consequently, the response to gut microbiota-derived metabolites shows vast combinatorial diversity, posing substantial challenges in understanding their effects. Metabolites exhibit both paracrine and endocrine effects, enabling them to influence both local and systemic levels [44]. Systemically, metabolites can traverse the intestinal epithelium through active or passive transportation and directly impact organ function or contribute to diseases in distant tissues. In the gut–liver axis, BA processed by the gut microbiota can be transported back to the liver via the portal vein or disseminated to other body parts [45]. Gut microbiota-derived metabolites can also be carried to various tissues directly through the bloodstream, thereby influencing organ function and overall host physiology. Moreover, microbiota can generate metabolites that influence other microbiota members or pathogens, thereby modifying the overall composition or behavior of certain members. These changes can ultimately affect the host at the local and systemic level. This comprehensive review integrates up-to-date knowledge on the pivotal role of gut-microbiota-derived metabolites, emphasizing the need for further investigations to elucidate the complex mechanisms underlying their interaction with the host immune system.
Gut microbiota-derived metabolites
The diversity of gut microbiota-derived metabolites is vast, and their synthesis is intricately interconnected. Specific metabolites, such as SCFAs, lactate, succinate, amino acid-derived compounds, and BA, are pivotal to preserving homeostasis, despite belonging to distinct metabolic classes. In this review, we elucidate the primary types of gut microbiota-derived metabolites, focusing on those of particular relevance to host homeostasis.
Short-chain fatty acids
SCFAs, also known as volatile fatty acids, are a category of fatty acid species containing fewer than six carbons that represent the most extensively studied metabolites in host–microbiota interactions. The biosynthetic pathways of SCFAs are closely integrated and share gut microbiota-derived monosaccharides as instigators. Each SCFA can be generated by multiple synthetic pathways provided by distinct microbiota sets. The biosynthetic pathway of SCFAs has been comprehensively reviewed previously [46]. While gut microbiota-derived SCFAs with one or five carbons, such as formate and valerate, are rarely recognized for their role in health and disease [47, 48], SCFAs produced by the microbiota, such as acetate (a two-carbon SCFA), propionate (a three-carbon SCFA), and butyrate (a four-carbon SCFA), are recognized for their involvement in various immunological and pathophysiological functions. These gut microbiota-derived SCFAs are absorbed into the bloodstream and have widespread effects on various cell types [49]. These effects occur through the interaction of SCFAs with specific receptors located on intestinal epithelial and immune cells. SCFAs are primarily produced in the colon due to the fermentation of indigestible dietary fiber, mainly by two anaerobic bacterial phyla: Bacteroidetes and Firmicutes [50]. Bacteroidetes, such as Bacteroides spp. play a significant role in the production of acetate and propionate [51]. Meanwhile, Firmicutes, exemplified by Faecalibacterium prausnitzii, are primarily responsible for generating butyrate [52].
Lactate and succinate
Central carbon metabolism not only produces SCFAs as primary end-products, but also plays an important role in the production of crucial intermediate metabolites, including lactate and succinate. Lactate is generated in the gut through various pathways, including the fermentation of dietary fibers by gut microbiota and lactate production by epithelial cells [53, 54]. The gut microbiota can produce lactate through the fermentation of diverse substrates, such as glucose, fructose, and complex carbohydrates [55, 56]. Representative lactic acid-producing bacteria (LAB) include probiotics such as Lactobacillus, Bifidobacterium, Lactococcus, Leuconostoc, and Weissella [57]. Two types of LAB exist depending on the fermentation method. Homofermentative bacteria exclusively generate lactic acid as their main byproduct during glucose fermentation. In contrast, heterofermentative bacteria synthesize a substantial quantity of other metabolites, such as ethanol, acetic acid, propionic acid, acetaldehyde, or diacetyl, alongside lactate [58]. Lactate also significantly impacts the composition and diversity of the gut microbiota as it serves as a substrate for microbial growth and metabolism in the gut microbiota. Succinate functions as an intermediate metabolite in the production of propionate and is found in large quantities in intestinal luminal contents. Its levels significantly fluctuate under dysbiotic conditions, such as antibiotic treatment or inflammation, indicating a close association between succinate metabolism and the gut microbiota [59]. Although little is known about the microbial source of succinate, studies suggest that Prevotella is associated with its production in the human intestine [60]. Another study revealed that monocolonization of germ-free mice with Bacteroides acidifaciens increased intestinal succinate concentration compared with colonization with other gut microbiota, such as the Clostridia consortium [61].
Bacterial membrane and dietary lipids
In addition to well-characterized smaller lipids, such as SCFAs and their intermediates, a significant proportion of larger lipid metabolites is produced by the gut microbiota. Some of these lipids are synthesized de novo and presented in the microbial membrane to influence host immunity. Most bacterial membrane lipids comprise fatty acid chains, also known as acyl chains, attached to distinct backbones. These acyl chains differ in length and saturation, giving rise to a variety of lipid species. The most recognized classes of membrane-bound lipids found in bacterial membranes include saccharolipids, phospholipids, sphingolipids, sulfonolipids, and cardiolipins [62,63,64,65,66]. The gut microbiota also plays a significant role in transforming, purifying, and digesting dietary fats, and the resulting products can impact both local and systemic homeostasis [67]. For instance, the Bacteroidetes phylum transforms dietary sphingolipid and cholesterol using their enzymes [68,69,70], while Bifidobacterium expresses enzymes that break down complex sphingolipids [69]. Lactobacillus and Bifidobacterium convert linoleic acid to conjugated linoleic acid (CLA) using their enzymes [71], and dietary polyunsaturated fatty acids (e.g., \(\omega\)-6 and -3 fatty acids) can be modified by gut microbiota-derived enzymes, causing changes in the gut microbiota composition [72,73,74]. Clostridium butyricum induces \(\omega\)-3 fatty acid production in the gut [75], while saturated fatty acids, such as oleic and palmitic acid, are produced by Bacteroides thetaiotaomicron, Lactobacillus johnsonii, and Lactobacillus paracasei, inhibiting viruses and fungi [33, 76].
Amino acids-derived metabolites
Due to the substantial presence of gut microbiota in the intestine, their integral metabolism of amino acids results in the production of a wide range of related metabolites, including ammonia, amines, nitrogen compounds, phenols, and precursors to SCFAs [77]. Byproducts originating from aromatic amino acids such as tryptophan, tyrosine, and phenylalanine give rise to a diverse array of phenolic and indolic compounds through a series of alterations [78]. Tryptophan, an aromatic amino acid, undergoes metabolism by gut microbiota, leading to the formation of its derivatives, such as indole, indolepropionic acid (IPA), indoleacrylic acid (IAA), 3-methylindole (Skatole), indolealdehyde (IAld), indoleethanol (IE), indolelactic acid (ILA), and tryptamine [79]. Whereas bacteria belonging to Actinobacteria, Firmicutes, Bacteroidetes, and Proteobacteria produce a variety of tryptophan metabolites with different genera, yielding distinct types of these metabolites [79, 80]; for example, Clostridium sporogenes and Clostridium botulinum produce IPA [81, 82]. Meanwhile, broader gram-positive and gram-negative gut microbiota, such as Enterococcus faecalis and Escherichia coli, produce indole, in addition to Firmicutes [83, 84]. Additionally, l-carnitine, an amino acid abundant in red meat, is converted to trimethylamine (TMA) by the gut microbiota [85]. TMA is delivered to the liver, where it is then converted to trimethylamine N-oxide (TMAO), accelerating atherosclerosis in mouse models.
Bile acids
BAs, synthesized in the liver, are released into the small intestine to facilitate the digestion and absorption of dietary fats. The liver produces primary BAs by converting cholesterol through the enzyme cholesterol 7α-hydroxylase (CYP7a1) [86]. The two main BAs, chenodeoxycholic acid (CDCA) and cholic acid (CA), conjugate with glycine or taurine to form glycol-conjugated or tauro-conjugated BAs. Conjugated primary BAs are vital in the digestion and absorption of lipids and vitamins [87]. Approximately 95% of primary BAs are actively reabsorbed from the ileum and undergo recycling within the liver, a process known as enterohepatic circulation [88]. Primary conjugated BAs can also undergo further modification by the gut microbiota. Initially deconjugated, they are subsequently transformed into various secondary BAs, such as deoxycholic acid (DCA), lithocholic acid (LCA), and ursodeoxycholic acid (UDCA). These secondary BAs are then passively reabsorbed into the circulating BA pool or eliminated through feces. Meanwhile, a portion of both primary and secondary BAs enters systemic circulation, exerting biological functions in distal parts of the body [89]. To generate secondary BAs, gut microbiota must express bile salt hydrolases (BSHs), necessary for deconjugation, and multiple transforming enzymes [90, 91]. Bacteria belonging to Firmicutes, Proteobacteria, Fusobacterium, Bacteroidetes, and Actinobacteria express these enzymes; however, the types of secondary BAs they produce vary according to the diversity of BSH-encoding genes and specific dehydrogenase they express [91, 92].
Receptors for gut microbiota-derived metabolites
G protein-coupled receptors
GPRs constitute a class of cell membrane receptors that play a fundamental role in cell signaling [93]. They are involved in various physiological processes, including the regulation of neurotransmission, hormonal signaling, immune responses, and the perception of taste and odor. Abundantly present throughout the body, GPRs are also found in the gastrointestinal tract. Endeavors to identify the functions of orphan GPRs have revealed interactions between several selective GPRs with various metabolites [42]. These metabolites, acting as signaling molecules, include substances such as SCFAs and BAs, some of which are derived from the gut microbiota, and play a role in triggering classical signal transduction processes. GPRs specific to gut microbiota-derived metabolites include GPR40, GPR41, GPR43, GPR81, GPR91, GPR109A, and GPR120, which are classified into four families according to their alpha subunit (Gi, Gs, G12/13, and Gq) [94,95,96]. Most GPRs recognizing metabolites derived from the gut microbiota have Gi or Gq-coupled receptors. For instance, GPR41, GPR81, and GPR109A comprise Gαi receptors, while GPR40, GPR43, and GPR91 have Gαi/Gαq, and GPR120 has Gq. The Gi family modulates adenylyl cyclase, and Gq stimulates phospholipase C [42]. Specific GPR ligands exhibit bias by selectively activating particular pathways downstream of a GPR. These biased ligands target only a subset of these pathways, leading to a more focused and specific cellular response [97]. For instance, GPR40, proposed as a potential target for diabetes treatment, functions as a Gq-coupled receptor responsive to long-chain fatty acids (LCFA). When triggered by specific agonists, GPR40 can initiate signaling not only through Gq and IP3 but also via Gs and cAMP, setting it apart from natural LCFA ligands [98].
Pattern recognition receptors
Membrane lipids distinguish themselves among microbial metabolites by virtue of their exclusive presence in microbes, rendering them potent pathogen-associated molecular patterns (PAMPs). They serve as distinctive microbial signatures that can be readily recognized by the host's pattern recognition receptors (PRRs), such as Toll-like receptors (TLRs). A notable example is lipopolysaccharide (LPS), a prominent member of the saccharolipid class produced by a broad spectrum of gram-negative bacteria [99]. The recognition of LPS by TLR4 initiates a cascade of reactions that can result in overt inflammation. However, this inflammation can, at times, be beneficial in preserving homeostasis because weak TLR4 stimuli can induce tolerogenic immune cell activation, thereby restoring disrupted intestinal immune balance [100]. Similarly, phospholipids produced by A. muciniphila can stimulate TLR1/2 to induce homeostatic immunity [29]. There is a possibility that the continuous interaction between coexisting gut bacteria and their inherent PAMPs has evolved towards maintaining immunity rather than over-activating it. Hence, further research into the immunoregulatory function of PRRs holds substantial promise for our understanding and the development of strategies to maintain immune balance in the future.
Aryl hydrocarbon receptor
The host experiences a diverse array of effects when tryptophan-derived metabolites engage with the aryl hydrocarbon receptor (AhR), a nuclear receptor [101]. Tryptophan derivatives, such as indole propionic acid, indole-3-aldehyde, indole lactic acid, indole pyruvate, and indole acetic acid, act as microbiota-derived AhR ligands [79]. In the absence of a ligand, AhR exists in the cytosol as a protein complex comprising HSP90 (90 KDa heat shock protein), AhR-interacting protein (AIP), cochaperone p23, and protein kinase c-Src [102]. After ligand binding, a conformational change induced in AhR leads to the dissociation of the complex and nuclear translocation. In the nucleus, a series of downstream gene expressions occur. The activation of the AhR signaling pathway holds promise in alleviating inflammation-related disorders, such as ulcerative colitis [103], and regulation of gut motility by modulating enteric neurons, potentially bolstering mucosal barrier function [36, 37, 104, 105].
Farnesoid X receptor and Takeda G protein-coupled receptor 5
The Farnesoid X Receptor (FXR) is a nuclear receptor protein that is critical in the regulation of various metabolic processes throughout the body [106]. Primarily located in the liver, intestine, and other tissues involved in BA metabolism and lipid homeostasis, FXR acts as a transcription factor, controlling the expression of specific genes by binding to their regulatory regions [107]. Its primary function is to regulate the synthesis of BAs, which, in turn, reduces the expression of CYP7a1 by triggering hepatic FXR [108]. Similarly, in the small intestine, activated FXR hinders BA synthesis by releasing fibroblast growth factor (FGF) 15/19 [109]. FGF15/19 then moves to the liver, where it activates FGF receptor 4 on hepatocytes and causes suppression of CYP7a1 expression. FXR also has a broader role in regulating glucose and lipid metabolism, including influencing the expression of lipid and glucose homeostasis-related genes in the liver and adipose tissue [110]. Takeda G protein-coupled receptor 5 (TGR5) is found on cell surfaces and activated by BAs [111]. It is present in different body tissues such as the gallbladder, intestine, and brown adipose tissue. When BAs bind to TGR5, they initiate internal signaling pathways that control several bodily functions, including energy processing, glucose level maintenance, and immune system responses [112,113,114]. BAs act as signaling molecules by activating both TGR5 and FXR receptors, regulating various metabolic processes, and maintaining BA homeostasis. The interaction between these receptors and BAs is crucial for preserving metabolic balance and regulating diverse physiological functions throughout the body.
Impact of gut microbiota-derived metabolites on host homeostasis
Modulation of mucosal immune cell functions
Microbiota-produced metabolites act as signaling molecules, engaging with host cells, especially immune cells, at mucosal surfaces [115]. These metabolites directly impact immune cell behavior, influencing factors such as cell activation, differentiation, migration, and cytokine production. By interacting with receptors on immune cells, microbiota-derived metabolites enhance or suppress immune responses depending on the specific metabolite and context. This finely tuned modulation is essential for maintaining balance in the immune system. SCFAs wield significant influence on immune cells in the gut mucosa, which affects their activation and regulation, particularly the differentiation and activation of regulatory T (Treg) cells [15, 116]. Dietary fiber and bacterial SCFAs also heighten the activity of tolerogenic dendritic cells and expression of retinaldehyde dehydrogenase, leading to increased Treg differentiation [117, 118]. Propionates suppress IL-17 and IL-22-producing intestinal γδ T cells through a histone deacetylase-dependent mechanism [17]. Butyrate, derived from gut microbiota, alleviates arthritis by boosting the production of IL-10-producing regulatory B cells. It achieves this by increasing the concentration of the tryptophan-derived metabolite 5-hydroxyindole-3-acetic acid, which activates the AhR [18]. Butyrate also enhances the memory potential of CD8+ T cells [19]. SCFAs also modulate dendritic cell function by promoting their maturation, enhancing antigen presentation, and influencing cytokine production [119, 120]. Microbiota-produced succinates contribute to the expansion of tuft cells and the suppression of ileal gut inflammation [27]. These microbial succinates also activate tuft cells and initiate a type 2 innate immune response in the small intestine [28]. Microbiota-derived BAs influence colonic Treg homeostasis [121, 122]. For instance, BA isoDCA restricts the activity of FXR in dendritic cells, conferring an anti-inflammatory phenotype and enhancing the induction of Foxp3 in Treg cells [38]. 3-OxoLCA impedes the development of Th17 cells by interacting directly with the crucial transcription factor RORγt, while isoalloLCA encourages the formation of Treg cells [39]. Gut microbiota-derived lipid metabolites also have immunomodulatory effects. Metabolites such as 9, 10-dihydroxy-12Z-octadecenoic acid, and all-trans retinoic acid from specific-pathogen-free mice induce Treg activity [30]. Moreover, conjugated linoleic acid (CLA) derived from gut microbiota promotes the induction of a CD4+ intraepithelial lymphocyte population expressing CD8αα in the small intestine [34]. Bacteroides fragilis sphingolipids regulate colonic iNKT cell homeostasis and shield against colitis challenges [123]. Additionally, cell surface polysaccharides derived from Bifidobacterium bifidum stimulate Treg cells, effectively suppressing experimental colitis [31]. Overall, metabolites derived from the gut microbiota collectively play a crucial role in modulating the function and equilibrium of mucosal immune cells.
Modulation of intestinal barrier function
The intestinal barrier comprises three protective layers that prevent the entry of bacteria from the gut lumen [124]. These layers include the luminal mucus layer, a continuous sheet of gut epithelial cells, and an inner layer responsible for forming the mucosal immune system. Metabolites derived from the microbiota help maintain the integrity of the mucosal barrier [125]. They aid in enhancing mucus production and tightening epithelial cell junctions, preventing the entry of harmful substances. Some metabolites also support tissue repair and regeneration, crucial for the quick recovery of mucosal surfaces after damage or inflammation [126]. By providing both physical and immunological defense mechanisms, this barrier is critical in defending against luminal microorganisms, viruses, food antigens, and environmental toxins. The gut epithelial barrier comprises a single layer of intestinal epithelial cells (IECs) that renew every 3–5 days. These cells originate from intestinal stem cells (ISCs), expressing leucine-rich repeat-containing G-protein-coupled receptor 5 (Lgr5) on their surface (Fig. 1). These ISCs give rise to ISC daughters or transit-amplifying (TA) cells [127]. TA cells then eventually undergo terminal differentiation and develop into absorptive or secretory cell lineages, including enterocyte, Paneth, goblet, enteroendocrine, and tuft cells. Our group has documented the interaction between microbiota-derived metabolites and the development of IECs. Lactates, produced by probiotics such as Lactobacillus and Bifidobacterium boost the proliferation of Lgr5+ ISCs through the Wnt/β-catenin pathway in Paneth cells and intestinal stromal cells [24] (Fig. 1). Moreover, lactates influence ISC-mediated epithelial development in a GPR81-dependent manner. A. muciniphila is recognized as a prominent symbiont and next-generation probiotic due to its health-promoting effects [128]. Interestingly, during the process of mucin degradation A. muciniphila also paradoxically enhances mucin production [16]. Moreover, it stimulates the regeneration of Lgr5+ ISCs and facilitates the differentiation of secretory lineage cells, including Paneth and goblet cells, within the gut. This process involves the utilization of metabolites such as acetate and propionate through GPR41/43 signaling. Additionally, BAs, specifically LCA, activate IEC regeneration via the BAs-TGR5 axis [40]. IECs establish connections through junctional proteins, including tight junction proteins, adherens junction proteins, gap junction proteins, and desmosomes [129]. The relationship between microbiota-derived metabolites and tight junctions is complex and bidirectional. These metabolites impact the expression and function of proteins associated with tight junctions, and assist in fortifying the assembly and integrity of these junctions, contributing to a more robust gut barrier. Various gut microbiota-derived metabolites, such as butyrate, quercetin, indole 3-propionic acid, tryptamine, and L-homoserine, can trigger an increase in the production of these junctional proteins [20, 130, 131]. Conversely, imbalances in the microbiota and their metabolites can affect tight junction function. Dysbiosis has been associated with changes in tight junction integrity and increased intestinal permeability [132], allowing harmful substances to translocate into the bloodstream, leading to inflammation and the development of metabolic diseases. Research suggests that conditions such as leaky gut syndrome, disruption of gut barrier function, and endotoxemia originating from the gut microbiota may contribute to the development of obesity and type 2 diabetes [133, 134]. A Western-style diet has been shown to increase gut permeability and reduce the expression of tight junction proteins, such as zonula occludens-1 and occludin, in the intestinal epithelial cells of mice [135]. In obese mice induced either genetically or by a high-fat diet, disruption of the gut barrier increases permeability, resulting in the release of LPS into the portal blood circulation [136, 137]. Multiple studies indicate that LPS plays a crucial role in initiating obesity and developing type 2 diabetes, with elevated LPS levels observed in obese and diabetic patients, suggesting its role as a marker for decreased intestinal barrier function [138]. Furthermore, mice subjected to a high-fat diet demonstrated that dysfunction in the intestinal barrier leads to increased disorders related to glucose dysmetabolism and liver steatosis [139].

The process of maintaining the balance in the renewal and differentiation of intestinal epithelial cells. Lgr5+ stem cells reside adjacent to Paneth cells at the base of the crypt. These Lgr5+ stem cells constantly generate rapidly proliferating TA cells, filling the crypt. Subsequently, TA cells undergo differentiation into various functional cell types present within the villi, including enterocytes, tuft cells, goblet cells, and enteroendocrine cells. Lactic acid-producing bacteria, such as Bifidobacterium and Lactobacillus, contribute to the renewal of intestinal epithelial cells. The lactate produced by these symbiotic organisms is recognized by the GPR81 on Paneth and stromal cells, facilitating regeneration through the Wnt3/β-catenin pathway. The mucin-degrading bacterium A. muciniphila promotes intestinal stem cell-mediated epithelial regeneration using metabolites such as SCFA within the Wnt3/β-catenin pathway. Gut microbiota-derived tryptophan derivatives, along with SCFA, stimulate the expression of junctional proteins (such as occludin, claudins, and zonula occludens), thereby enhancing the gut barrier functions
Gut–bone marrow interaction
The bone marrow, located within bone cavities, contains pluripotent hematopoietic stem cells (HSCs) that differentiate into lymphoid progenitor or myeloid progenitor cells, which play a crucial role in the immune response, oxygen transport, and blood clotting. The bone marrow is therefore a central component of the circulatory and immune systems. The "gut–bone marrow axis" implies that there is a dynamic interplay between the gut and the bone marrow extending beyond their individual functions (Fig. 2). Recent research has revealed that substances produced by the gut microbiota, such as metabolites and signaling molecules, can impact the activity and function of bone marrow cells [25, 140]. Lactate produced by probiotics enters the bloodstream and reaches the bone marrow, where it stimulates the expression of stem cell factors necessary for the proliferation of HSCs in a GPR81-dependent manner [25]. These findings underscore a crucial link between microbiota-produced lactate and the expansion of HSCs, thereby providing continuous support for hematopoiesis and erythropoiesis. Moreover, increased myeloid cell differentiation from HSCs, prompted by microbiota-derived LPS stimulation, leads to higher and more sustained levels of IL-1α and IL-1β in the bone marrow [140]. Conversely, the bone marrow can reciprocally influence the gut by producing immune cells and factors that regulate gut inflammation [141, 142]. Bone marrow transplantation induces marked changes in microbiota composition and impacts the severity of intestinal graft-versus-host disease [141]. Transferring bone marrow from mice colonized with Clostridium scindens protects naive mice against colitis induced by Entamoeba histolytica [142]. Signals derived from commensals influence basophil hematopoiesis and susceptibility to allergic inflammation [143]. Peptidoglycan from the gut microbiota regulates the steady-state lifespan of neutrophils and monocytes, contributing to myeloid homeostasis [144]. Additionally, microbiota-derived compounds maintain steady-state granulopoiesis through MyD88 signaling [145]. Butyrates derived from the gut microbiota alter hematopoiesis by increasing patrolling Ly6c− monocytes in the context of influenza virus infection [21]. The microbiota regulates bone marrow function via butyrates and controls HSC self-renewal in an iron-dependent manner [22].

Cross-talk between gut microbiota-derived metabolites and various organs. Microbial byproducts, such as lactate, traverse through the bloodstream to reach the bone marrow, triggering stromal cells to release SCF and Cxcl2 influenced by GPR81 signaling. This interaction impacts bone marrow functions. Butyrate, generated by the gut microbiota, reinforces the gut barrier, fortifies the host against pathogen infiltration, and alleviates the effects of pancreatitis. Certain bacteria producing BSH elevate levels of deconjugated and secondary BAs within the colon. These BAs stimulate the production of Wnt2b in colonic mesenchymal stem cells through FXR activation, promoting increased cell growth in colonic crypts. Palmitic acid derived from Lactobacillus exhibits the ability to enhance the production of type-I IFN from pDC and monocytes. This increase in fatty acids contributes to the host's defense against influenza infection through the GPR40/120 mechanism
Gut–lung interaction
Recent findings have illuminated the intricate communication network between the gut microbiota and the respiratory system, often referred to as the gut–lung axis. This multifaceted framework encompasses the gut microbiota, immune responses, inflammatory signaling, and dietary influences to link the gut and the lungs [146]. The dynamic interplay between these distant anatomical sites underscores the potential influence of metabolites originating from the gut microbiota on immune responses in the lungs [147]. Gut signals from Proteobacteria direct the migration of group 2 innate lymphoid cells from the gut to the lung via the gut-lung axis [148]. During an infectious state, gut microbiota-derived metabolites are crucial in protecting the host. Palmitic acid, derived from Lactobacillus paracasei, increases the production of type-I IFN from plasmacytoid DC and monocyte [33], which helps protect the host against influenza infection through a mechanism relying on GPR40/120. Metabolites derived from Clostridium orbiscindens in the human gut, such as desaminotyrosine, protect against respiratory virus infection by activating type-I IFN in macrophage [149], while glycolipids associated with Bacteroides fragilis confer resistance to vesicular stomatitis virus and influenza virus by type-I IFN producing colonic dendritic cells [32]. Additionally, the presence of SCFAs produced through bacteria-induced inulin fermentation increases the numbers of patrolling monocytes and enhances the effectiveness of CD8+ T cells, thereby mitigating the lung damage associated with influenza virus infection [21]. Furthermore, SCFAs help defend against allergic inflammation in the lungs by promoting the development of dendritic cell precursors through hematopoiesis [150]. Supplementation of CLA produced by gut microbiota, such as Bifidobacterium and Lactobacillus, reduces inflammation in individuals with allergies by reducing TNF-α and IFN-γ production from peripheral blood mononuclear cells [35].
Gut–liver interaction
The gut–liver interaction is a multifaceted and dynamic relationship characterized by intricate biochemical, immunological, and endocrine communication. The balance and synergy between the gut and the liver play a pivotal role in BA metabolism, digestion, immune regulation, hormonal signaling, and overall health [151]. The portal vein receives most of the blood from the small and large intestine, carrying nutrients absorbed from the digestive tract and metabolites derived from the microbiota [152]. For example, gut microbiota-derived D-lactate arriving at the liver via the portal vein promotes the killing of circulating pathogens by Kupffer cells [26]. BAs modulate the composition of gut microbiota by utilizing their detergent properties to disrupt bacterial membranes [153], and induce stress, including DNA damage and protein misfolding [154]. BAs also activate FXR in IEC, promoting the production of antimicrobial molecules and inhibiting bacteria overgrowth [155]. Tauro-cholic acid and β-tauro-muricholic acid accelerate postnatal microbiota maturation in newborn mice [156]. Dietary signals influence BA-mediated gut microbiota composition and homeostasis, with a high-fat diet shown to increase the levels of primary BAs (e.g., chenodeoxycholic acid) and bacteria-producing BSHs [41]. These bacteria upregulate secondary BAs, which induce the production of Wnt2b through mesenchymal stem cells in the colon in a BA–FXR-dependent manner. Furthermore, BSH-producing bacteria and retinol modulate BA production in the gut in an intestinal liver kinase B1-dependent manner [157].
Gut–pancreas interaction
The pancreas is connected to the gastrointestinal tract through the pancreatic duct system, creating an inherent association with the gut microbiota. In its healthy state, the pancreas does not directly interact with the gut microbiota and is traditionally not believed to have its own microbiome. Nevertheless, gut bacteria can migrate into the pancreas and potentially affect the pancreatic environment, even in individuals with a healthy pancreas. Dysbiosis has been detected in pancreatic disorders and could contribute to the development of various pancreatic conditions such as acute and chronic pancreatitis, and pancreatic cancer [158, 159]. Gut microbiota can also be directly transported through the pancreatic ductal system [160], a process facilitated by dysbiosis through the disruption of mucosal immune function and increase in gut permeability [161]. A recent study also validates the concept of gut microbiota-produced butyrate enhancing the integrity of the gut barrier, protecting the host from pathogen invasion and alleviating pancreatitis [23]. Patients with pancreatitis who received probiotics such as Lactobacillus plantarum showed improved gut permeability and clinical outcomes compared with the control group [162]. Additionally, administrating Lactobacillus plantarum and oat fiber to these patients effectively reduced pancreatic sepsis [163]. In pancreatic cancer, the gut microbiota modulates the tumor microbiome landscape and influences clinical outcomes [164]. Gut microbiota therefore plays a crucial role in pancreatic health and disease.
Therapeutic implications and future perspective
The therapeutic landscape is actively exploring the potential of gut microbiota-derived metabolites across various clinical domains. Probiotics, encompassing distinct bacterial strains, are currently used to modulate the gut microbiota and stimulate the production of beneficial metabolites. Concurrently, dietary interventions, prebiotics, and postbiotics are emerging as promising therapeutic strategies to target these metabolites for therapeutic purposes. As this field matures, we could expect to see: (1) personalized therapies: tailoring treatments to the unique gut microbiota composition of an individual to optimize the production of beneficial metabolites; (2) advanced metabolomics: ongoing advancements in metabolomics technologies will furnish a more comprehensive understanding of the diverse spectrum of metabolites originating from the gut microbiota; (3) microbiome engineering: pioneering approaches to engineering the gut microbiota, driving production of specific metabolites for therapeutic utility. The intricate interplay between gut microbiota-derived metabolites and host homeostasis represents a flourishing area brimming with therapeutic promise. Harnessing these metabolites stands to unlock novel interventions across a broad spectrum of health conditions. The future holds the promise of innovative therapies leveraging the extraordinary capabilities of the gut microbiota in shaping host health and patient outcomes.
Conclusion
This comprehensive review aims to consolidate current knowledge regarding the role of gut microbiota-derived metabolites in shaping host homeostasis. By elucidating the mechanisms of action and the impact on various physiological processes, this paper provides a foundation for future research and therapeutic interventions targeting the gut microbiota and its metabolites. Understanding the intricate relationship between gut microbiota and host physiology is essential for advancing our knowledge of health and disease and has the potential to revolutionize medical interventions.
Data availability
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
References
Gordon JI (2012) Honor thy gut symbionts redux. Science 336:1251–3. https://doi.org/10.1126/science.1224686
Lynch SV, Pedersen O (2016) The human intestinal microbiome in health and disease. N Engl J Med 375:2369–2379. https://doi.org/10.1056/NEJMra1600266
Fan Y, Pedersen O (2021) Gut microbiota in human metabolic health and disease. Nat Rev Microbiol 19:55–71. https://doi.org/10.1038/s41579-020-0433-9
Rinninella E, Raoul P, Cintoni M, Franceschi F, Miggiano GAD et al (2019) What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 7(1):14. https://doi.org/10.3390/microorganisms7010014
De Vadder F, Kovatcheva-Datchary P, Zitoun C, Duchampt A, Backhed F, Mithieux G (2016) Microbiota-produced succinate improves glucose homeostasis via intestinal gluconeogenesis. Cell Metab 24:151–157. https://doi.org/10.1016/j.cmet.2016.06.013
Tremaroli V, Backhed F (2012) Functional interactions between the gut microbiota and host metabolism. Nature 489:242–249. https://doi.org/10.1038/nature11552
Ansaldo E, Farley TK, Belkaid Y (2021) Control of Immunity by the Microbiota. Annu Rev Immunol 39:449–479. https://doi.org/10.1146/annurev-immunol-093019-112348
Skelly AN, Sato Y, Kearney S, Honda K (2019) Mining the microbiota for microbial and metabolite-based immunotherapies. Nat Rev Immunol 19:305–23. https://doi.org/10.1038/s41577-019-0144-5
Ashida H, Ogawa M, Kim M, Mimuro H, Sasakawa C (2011) Bacteria and host interactions in the gut epithelial barrier. Nat Chem Biol 8:36–45. https://doi.org/10.1038/nchembio.741
Stefka AT, Feehley T, Tripathi P, Qiu J, McCoy K et al (2014) Commensal bacteria protect against food allergen sensitization. Proc Natl Acad Sci USA 111:13145–13150. https://doi.org/10.1073/pnas.1412008111
Integrative HMP Research Network Consortium (2019) The integrative human microbiome project. Nature 569:641–648. https://doi.org/10.1038/s41586-019-1238-8
Lloyd-Price J, Arze C, Ananthakrishnan AN, Schirmer M, Avila-Pacheco J et al (2019) Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 569:655–662. https://doi.org/10.1038/s41586-019-1237-9
Kamada N, Kim YG, Sham HP, Vallance BA, Puente JL et al (2012) Regulated virulence controls the ability of a pathogen to compete with the gut microbiota. Science 336:1325–1329. https://doi.org/10.1126/science.1222195
Hudault S, Guignot J, Servin AL (2001) Escherichia coli strains colonising the gastrointestinal tract protect germfree mice against Salmonella typhimurium infection. Gut 49:47–55. https://doi.org/10.1136/gut.49.1.47
Smith PM, Howitt MR, Panikov N, Michaud M, Gallini CA et al (2013) The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 341:569–573. https://doi.org/10.1126/science.1241165
Kim S, Shin YC, Kim TY, Kim Y, Lee YS et al (2021) Mucin degrader Akkermansia muciniphila accelerates intestinal stem cell-mediated epithelial development. Gut Microbes 13:1–20. https://doi.org/10.1080/19490976.2021.1892441
Dupraz L, Magniez A, Rolhion N, Richard ML, Da Costa G et al (2021) Gut microbiota-derived short-chain fatty acids regulate IL-17 production by mouse and human intestinal γδ T cells. Cell Rep 36:109332. https://doi.org/10.1016/j.celrep.2021.109332
Rosser EC, Piper CJM, Matei DE, Blair PA, Rendeiro AF et al (2020) Microbiota-derived metabolites suppress arthritis by amplifying aryl-hydrocarbon receptor activation in regulatory B cells. Cell Metab 31:837–51. https://doi.org/10.1016/j.cmet.2020.03.003
Bachem A, Makhlouf C, Binger KJ, de Souza DP, Tull D et al (2019) Microbiota-derived short-chain fatty acids promote the memory potential of antigen-activated CD8+ T Cells. Immunity 51:285–297. https://doi.org/10.1016/j.immuni.2019.06.002
Wang RX, Lee JS, Campbell EL, Colgan SP (2020) Microbiota-derived butyrate dynamically regulates intestinal homeostasis through regulation of actin-associated protein synaptopodin. Proc Natl Acad Sci USA 117:11648–11657. https://doi.org/10.1073/pnas.1917597117
Trompette A, Gollwitzer ES, Pattaroni C, Lopez-Mejia IC, Riva E et al (2018) Dietary fiber confers protection against flu by shaping Ly6c- patrolling monocyte hematopoiesis and CD8+ T cell metabolism. Immunity 48:992–1005. https://doi.org/10.1016/j.immuni.2018.04.022
Zhang D, Gao X, Li H, Borger DK, Wei Q et al (2022) The microbiota regulates hematopoietic stem cell fate decisions by controlling iron availability in bone marrow. Cell Stem Cell 29:232–247. https://doi.org/10.1016/j.stem.2021.12.009
van den Berg FF, van Dalen D, Hyoju SK, van Santvoort HC, Besselink MG et al (2021) Western-type diet influences mortality from necrotising pancreatitis and demonstrates a central role for butyrate. Gut 70:915–927. https://doi.org/10.1136/gutjnl-2019-320430
Lee YS, Kim TY, Kim Y, Lee SH, Kim S et al (2018) Microbiota-derived lactate accelerates intestinal stem-cell-mediated epithelial development. Cell Host Microbe 24:833–846. https://doi.org/10.1016/j.chom.2018.11.002
Lee YS, Kim TY, Kim Y, Kim S, Lee SH et al (2021) Microbiota-derived lactate promotes hematopoiesis and erythropoiesis by inducing stem cell factor production from leptin receptor+ niche cells. Exp Mol Med 53:1319–1331. https://doi.org/10.1038/s12276-021-00667-y
McDonald B, Zucoloto AZ, Yu IL, Burkhard R, Brown K et al (2020) Programing of an intravascular immune firewall by the gut microbiota protects against pathogen dissemination during infection. Cell Host Microbe 28:660–668. https://doi.org/10.1016/j.chom.2020.07.014
Banerjee A, Herring CA, Chen B, Kim H, Simmons AJ et al (2020) Succinate produced by intestinal microbes promotes specification of tuft cells to suppress ileal inflammation. Gastroenterology 159:2101–2115. https://doi.org/10.1053/j.gastro.2020.08.029
Nadjsombati MS, McGinty JW, Lyons-Cohen MR, Jaffe JB, DiPeso L et al (2018) Detection of succinate by intestinal tuft cells triggers a type 2 innate immune circuit. Immunity 49:33–41. https://doi.org/10.1016/j.immuni.2018.06.016
Bae M, Cassilly CD, Liu X, Park SM, Tusi BK et al (2022) Akkermansia muciniphila phospholipid induces homeostatic immune responses. Nature 608:168–173. https://doi.org/10.1038/s41586-022-04985-7
Shiratori H, Oguchi H, Isobe Y, Han KH, Sen A et al (2023) Gut microbiota-derived lipid metabolites facilitate regulatory T cell differentiation. Sci Rep 13:8903. https://doi.org/10.1038/s41598-023-35097-5
Verma R, Lee C, Jeun EJ, Yi J, Kim KS et al (2018) Cell surface polysaccharides of Bifidobacterium bifidum induce the generation of Foxp3+ regulatory T cells. Sci Immunol 3:eaat6975. https://doi.org/10.1126/sciimmunol.aat6975
Stefan KL, Kim MV, Iwasaki A, Kasper DL (2020) Commensal microbiota modulation of natural resistance to virus infection. Cell 183:1312–24. https://doi.org/10.1016/j.cell.2020.10.047
Kim S, Lee S, Kim TY, Lee SH, Seo SU, Kweon MN (2023) Newly isolated Lactobacillus paracasei strain modulates lung immunity and improves the capacity to cope with influenza virus infection. Microbiome 11:260. https://doi.org/10.1186/s40168-023-01687-8
Song X, Zhang H, Zhang Y, Goh B, Bao B et al (2023) Gut microbial fatty acid isomerization modulates intraepithelial T cells. Nature 619:837–843. https://doi.org/10.1038/s41586-023-06265-4
Turpeinen AM, Ylonen N, von Willebrand E, Basu S, Aro A (2008) Immunological and metabolic effects of cis-9, trans-11-conjugated linoleic acid in subjects with birch pollen allergy. Br J Nutr 100:112–119. https://doi.org/10.1017/S0007114507886326
Scott SA, Fu J, Chang PV (2020) Microbial tryptophan metabolites regulate gut barrier function via the aryl hydrocarbon receptor. Proc Natl Acad Sci USA 117:19376–19387. https://doi.org/10.1073/pnas.2000047117
Zelante T, Iannitti RG, Cunha C, De Luca A, Giovannini G et al (2013) Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 39:372–385. https://doi.org/10.1016/j.immuni.2013.08.003
Campbell C, McKenney PT, Konstantinovsky D, Isaeva OI, Schizas M et al (2020) Bacterial metabolism of bile acids promotes generation of peripheral regulatory T cells. Nature 581:475–479. https://doi.org/10.1038/s41586-020-2193-0
Hang S, Paik D, Yao L, Kim E, Trinath J et al (2019) Bile acid metabolites control Th17 and Treg cell differentiation. Nature 576:143–148. https://doi.org/10.1038/s41586-019-1785-z
Sorrentino G, Perino A, Yildiz E, El Alam G, Bou Sleiman M et al (2020) Bile acids signal via TGR5 to activate intestinal stem cells and epithelial regeneration. Gastroenterology 159:956–968. https://doi.org/10.1053/j.gastro.2020.05.067
Kim TY, Kim S, Kim Y, Lee YS, Lee S et al (2022) A high-fat diet activates the BAs-FXR axis and triggers cancer-associated fibroblast properties in the colon. Cell Mol Gastroenterol Hepatol 13:1141–1159. https://doi.org/10.1016/j.jcmgh.2021.12.015
Husted AS, Trauelsen M, Rudenko O, Hjorth SA, Schwartz TW (2017) GPCR-mediated signaling of metabolites. Cell Metab 25:777–796. https://doi.org/10.1016/j.cmet.2017.03.008
Kim CH (2021) Control of lymphocyte functions by gut microbiota-derived short-chain fatty acids. Cell Mol Immunol 18:1161–1171. https://doi.org/10.1038/s41423-020-00625-0
Liu J, Tan Y, Cheng H, Zhang D, Feng W, Peng C (2022) Functions of gut microbiota metabolites, current status and future perspectives. Aging Dis 13:1106–26. https://doi.org/10.14336/AD.2022.0104
Zheng Z, Wang B (2021) The gut-liver axis in health and disease: The role of gut microbiota-derived signals in liver injury and regeneration. Front Immunol 12:775526. https://doi.org/10.3389/fimmu.2021.775526
Koh A, De Vadder F, Kovatcheva-Datchary P, Backhed F (2016) From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 165:1332–1345. https://doi.org/10.1016/j.cell.2016.05.041
Ternes D, Tsenkova M, Pozdeev VI, Meyers M, Koncina E et al (2022) The gut microbial metabolite formate exacerbates colorectal cancer progression. Nat Metab 4:458–475. https://doi.org/10.1038/s42255-022-00558-0
Kuan AHY, Tey HL (2021) Topical glycopyrrolate followed by microneedling: a novel treatment option for eccrine hidrocystomas. J Dermatolog Treat 32:997–998. https://doi.org/10.1080/09546634.2020.1716929
Wang YQ, Wang HJ, Howard AG, Meyer KA, Tsilimigras MCB et al (2020) Circulating short-chain fatty acids are positively associated with adiposity measures in chinese adults. Nutrients 12:2127. https://doi.org/10.3390/nu12072127
Louis P, Flint HJ (2017) Formation of propionate and butyrate by the human colonic microbiota. Environ Microbiol 19:29–41. https://doi.org/10.1111/1462-2920.13589
El Hage R, Hernandez-Sanabria E, Calatayud Arroyo M, Props R, Van de Wiele T (2019) Propionate-producing onsortium restores antibiotic-induced dysbiosis in a dynamic in vitro model of the human intestinal microbial ecosystem. Front Microbiol 10:1206. https://doi.org/10.3389/fmicb.2019.01206
Reichardt N, Duncan SH, Young P, Belenguer A, McWilliam Leitch C et al (2014) Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J 8:1323–1335. https://doi.org/10.1038/ismej.2014.14
Ghio C, Soukup JM, Dailey LA, Ghio AJ, Schreinemachers DM et al (2022) Lactate production can function to increase human epithelial cell iron concentration. Cell Mol Bioeng 15:571–585. https://doi.org/10.1007/s12195-022-00741-z
Stolberg L, Rolfe R, Gitlin N, Merritt J, Mann L Jr et al (1982) D-Lactic acidosis due to abnormal gut flora: diagnosis and treatment of two cases. N Engl J Med 306:1344–1348. https://doi.org/10.1056/NEJM198206033062207
Chai LJ, Shen MN, Sun J, Deng YJ, Lu ZM et al (2020) Deciphering the d-/l-lactate-producing microbiota and manipulating their accumulation during solid-state fermentation of cereal vinegar. Food Microbiol 92:103559. https://doi.org/10.1016/j.fm.2020.103559
Belenguer A, Holtrop G, Duncan SH, Anderson SE, Calder AG et al (2011) Rates of production and utilization of lactate by microbial communities from the human colon. FEMS Microbiol Ecol 77:107–119. https://doi.org/10.1111/j.1574-6941.2011.01086.x
Abedi E, Hashemi SMB (2020) Lactic acid production - producing microorganisms and substrates sources-state of art. Heliyon 6:e04974. https://doi.org/10.1016/j.heliyon.2020.e04974
Muller T (1990) Comparison of methods for differentiation between homofermentative and heterofermentative lactic-acid bacteria. Zentralblatt Fur Mikrobiologie 145:363–366. https://doi.org/10.1016/S0232-4393(11)80064-8
Connors J, Dawe N, Van Limbergen J (2018) The role of succinate in the regulation of intestinal inflammation. Nutrients 11:25. https://doi.org/10.3390/nu11010025
Kovatcheva-Datchary P, Nilsson A, Akrami R, Lee YS, De Vadder F et al (2015) Dietary fiber-induced improvement in glucose metabolism is associated with increased abundance of Prevotella. Cell Metab 22:971–982. https://doi.org/10.1016/j.cmet.2015.10.001
Kim YG, Sakamoto K, Seo SU, Pickard JM, Gillilland MG 3rd et al (2017) Neonatal acquisition of Clostridia species protects against colonization by bacterial pathogens. Science 356:315–319. https://doi.org/10.1126/science.aag2029
Balasubramanian K, Maeda A, Lee JS, Mohammadyani D, Dar HH et al (2015) Dichotomous roles for externalized cardiolipin in extracellular signaling: Promotion of phagocytosis and attenuation of innate immunity. Sci Signal 22:8. https://doi.org/10.1126/scisignal.aaa6179
Heaver SL, Le HH, Tang P, Basle A, Mirretta Barone C et al (2022) Characterization of inositol lipid metabolism in gut-associated Bacteroidetes. Nat Microbiol 7:986–1000. https://doi.org/10.1038/s41564-022-01152-6
Radka CD, Miller DJ, Frank MW, Rock CO (2022) Biochemical characterization of the first step in sulfonolipid biosynthesis in Alistipes finegoldii. J Biol Chem 298:102195. https://doi.org/10.1016/j.jbc.2022.102195
Schlame M (2008) Cardiolipin synthesis for the assembly of bacterial and mitochondrial membranes. J Lipid Res 49:1607–1620. https://doi.org/10.1194/jlr.R700018-JLR200
Walker A, Pfitzner B, Harir M, Schaubeck M, Calasan J et al (2017) Sulfonolipids as novel metabolite markers of Alistipes and Odoribacter affected by high-fat diets. Sci Rep 7:11047. https://doi.org/10.1038/s41598-017-10369-z
Chadaideh KS, Carmody RN (2021) Host-microbial interactions in the metabolism of different dietary fats. Cell Metab 33:857–872. https://doi.org/10.1016/j.cmet.2021.04.011
Yao L, D’Agostino GD, Park J, Hang S, Adhikari AA et al (2022) A biosynthetic pathway for the selective sulfonation of steroidal metabolites by human gut bacteria. Nat Microbiol 7:1404–1418. https://doi.org/10.1038/s41564-022-01176-y
Lee MT, Le HH, Johnson EL (2021) Dietary sphinganine is selectively assimilated by members of the mammalian gut microbiome. J Lipid Res 62:100034. https://doi.org/10.1194/jlr.RA120000950
Le HH, Lee MT, Besler KR, Comrie JMC, Johnson EL (2022) Characterization of interactions of dietary cholesterol with the murine and human gut microbiome. Nat Microbiol 7:1390–1403. https://doi.org/10.1038/s41564-022-01195-9
Miyamoto J, Igarashi M, Watanabe K, Karaki SI, Mukouyama H et al (2019) Gut microbiota confers host resistance to obesity by metabolizing dietary polyunsaturated fatty acids. Nat Commun 10:4007. https://doi.org/10.1038/s41467-019-11978-0
Hennessy AA, Barrett E, Paul Ross R, Fitzgerald GF, Devery R, Stanton C (2012) The production of conjugated alpha-linolenic, gamma-linolenic and stearidonic acids by strains of bifidobacteria and propionibacteria. Lipids 47:313–327. https://doi.org/10.1007/s11745-011-3636-z
Druart C, Bindels LB, Schmaltz R, Neyrinck AM, Cani PD et al (2015) Ability of the gut microbiota to produce PUFA-derived bacterial metabolites: Proof of concept in germ-free versus conventionalized mice. Mol Nutr Food Res 59:1603–1613. https://doi.org/10.1002/mnfr.201500014
Costantini L, Molinari R, Farinon B, Merendino N (2017) Impact of omega-3 fatty acids on the gut microbiota. Int J Mol Sci 18:2645. https://doi.org/10.3390/ijms18122645
Hagihara M, Yamashita M, Ariyoshi T, Eguchi S, Minemura A et al (2022) Clostridium butyricum-induced ω-3 fatty acid 18-HEPE elicits anti-influenza virus pneumonia effects through interferon-λ upregulation. Cell Rep 41:111755. https://doi.org/10.1016/j.celrep.2022.111755
Charlet R, Le Danvic C, Sendid B, Nagnan-Le Meillour P, Jawhara S (2022) Oleic acid and palmitic acid from Bacteroides thetaiotaomicron and Lactobacillus johnsonii exhibit anti-inflammatory and antifungal properties. Microorganisms 10:1803. https://doi.org/10.3390/microorganisms10091803
Neis EP, Dejong CH, Rensen SS (2015) The role of microbial amino acid metabolism in host metabolism. Nutrients 7:2930–2946. https://doi.org/10.3390/nu7042930
Smith EA, Macfarlane GT (1996) Enumeration of human colonic bacteria producing phenolic and indolic compounds: effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J Appl Bacteriol 81:288–302. https://doi.org/10.1111/j.1365-2672.1996.tb04331.x
Roager HM, Licht TR (2018) Microbial tryptophan catabolites in health and disease. Nat Commun 9:3294. https://doi.org/10.1038/s41467-018-05470-4
Kaur H, Bose C, Mande SS (2019) Tryptophan metabolism by gut microbiome and gut-brain-axis: An in silico analysis. Front Neurosci 13:1365. https://doi.org/10.3389/fnins.2019.01365
Dodd D, Spitzer MH, Van Treuren W, Merrill BD, Hryckowian AJ et al (2017) A gut bacterial pathway metabolizes aromatic amino acids into nine circulating metabolites. Nature 551:648–652. https://doi.org/10.1038/nature24661
Elsden SR, Hilton MG, Waller JM (1976) The end products of the metabolism of aromatic amino acids by Clostridia. Arch Microbiol 107:283–288. https://doi.org/10.1007/BF00425340
Smith T (1897) A modification of the method for determining the production of indol by bacteria. J Exp Med 2:543–547. https://doi.org/10.1084/jem.2.5.543
Lee JH, Lee J (2010) Indole as an intercellular signal in microbial communities. FEMS Microbiol Rev 34:426–444. https://doi.org/10.1111/j.1574-6976.2009.00204.x
Koeth RA, Wang Z, Levison BS, Buffa JA, Org E et al (2013) Intestinal microbiota metabolism of L-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med 19:576–85. https://doi.org/10.1038/nm.3145
Chiang JY (2009) Bile acids: regulation of synthesis. J Lipid Res 50:1955–66. https://doi.org/10.1194/jlr.R900010-JLR200
de Aguiar Vallim TQ, Tarling EJ, Edwards PA (2013) Pleiotropic roles of bile acids in metabolism. Cell Metab 17:657–669. https://doi.org/10.1016/j.cmet.2013.03.013
Cai JS, Chen JH (2014) The mechanism of enterohepatic circulation in the formation of gallstone disease. J Membr Biol 247:1067–1082. https://doi.org/10.1007/s00232-014-9715-3
Dawson PA, Karpen SJ (2015) Intestinal transport and metabolism of bile acids. J Lipid Res 56:1085–1099. https://doi.org/10.1194/jlr.R054114
Jones BV, Begley M, Hill C, Gahan CG, Marchesi JR (2008) Functional and comparative metagenomic analysis of bile salt hydrolase activity in the human gut microbiome. Proc Natl Acad Sci USA 105:13580–13585. https://doi.org/10.1073/pnas.0804437105
Lucas LN, Barrett K, Kerby RL, Zhang Q, Cattaneo LE et al (2021) Dominant bacterial phyla from the human gut show widespread ability to transform and conjugate bile acids. mSystems 6:e0080521. https://doi.org/10.1128/mSystems.00805-21
Foley MH, O’Flaherty S, Allen G, Rivera AJ, Stewart AK et al (2021) Lactobacillus bile salt hydrolase substrate specificity governs bacterial fitness and host colonization. Proc Natl Acad Sci USA 118:e2017709118. https://doi.org/10.1073/pnas.2017709118
Weis WI, Kobilka BK (2018) The molecular basis of G protein-coupled receptor activation. Annu Rev Biochem 87:897–919. https://doi.org/10.1146/annurev-biochem-060614-033910
Kamato D, Thach L, Bernard R, Chan V, Zheng W et al (2015) Structure, function, pharmacology, and therapeutic potential of the G Protein, Gα/q,11. Front Cardiovasc Med 2:14. https://doi.org/10.3389/fcvm.2015.00014
Neves SR, Ram PT, Iyengar R (2002) G protein pathways. Science 296:1636–9. https://doi.org/10.1126/science.1071550
Pfleger J, Gresham K, Koch WJ (2019) G protein-coupled receptor kinases as therapeutic targets in the heart. Nat Rev Cardiol 16:612–622. https://doi.org/10.1038/s41569-019-0220-3
Wingler L, Lefkowitz R (2020) Conformational basis of G protein-coupled receptor signaling versatility. Trends Cell Biol 30(9):736–747. https://doi.org/10.1016/j.tcb.2020.06.002
Hauge M, Vestmar M, Husted A, Ekberg J, Wright M et al (2014) GPR40 (FFAR1)-Combined Gs and Gq signaling in vitro is associated with robust incretin secretagogue action ex vivo and in vivo. Mol Metab 4(1):3–14. https://doi.org/10.1016/j.molmet.2014.10.002
Poltorak A, He X, Smirnova I, Liu MY, Van Huffel C et al (1998) Defective LPS signaling in C3H/HeJ and C57BL/10ScCr mice: mutations in Tlr4 gene. Science 282:2085–2088. https://doi.org/10.1126/science.282.5396.2085
Steimle A, Michaelis L, Di Lorenzo F, Kliem T, Munzner T et al (2019) Weak agonistic LPS restores intestinal immune homeostasis. Mol Ther 27:1974–1991. https://doi.org/10.1016/j.ymthe.2019.07.007
Heath-Pagliuso S, Rogers WJ, Tullis K, Seidel SD, Cenijn PH et al (1998) Activation of the Ah receptor by tryptophan and tryptophan metabolites. Biochemistry 37:11508–11515. https://doi.org/10.1021/bi980087p
Tsuji N, Fukuda K, Nagata Y, Okada H, Haga A et al (2014) The activation mechanism of the aryl hydrocarbon receptor (AhR) by molecular chaperone HSP90. FEBS Open Bio 4:796–803. https://doi.org/10.1016/j.fob.2014.09.003
Lv Q, Wang K, Qiao S, Yang L, Xin Y et al (2018) Norisoboldine, a natural AhR agonist, promotes Treg differentiation and attenuates colitis via targeting glycolysis and subsequent NAD+/SIRT1/SUV39H1/H3K9me3 signaling pathway. Cell Death Dis 9:258. https://doi.org/10.1038/s41419-018-0297-3
Obata Y, Castano A, Boeing S, Bon-Frauches AC, Fung C et al (2020) Neuronal programming by microbiota regulates intestinal physiology. Nature 578:284–289. https://doi.org/10.1038/s41586-020-1975-8
Wang X, Huang S, Zhang M, Su Y, Pan Z et al (2023) Gegen Qinlian decoction activates AhR/IL-22 to repair intestinal barrier by modulating gut microbiota-related tryptophan metabolism in ulcerative colitis mice. J Ethnopharmacol 302:115919. https://doi.org/10.1016/j.jep.2022.115919
Sun L, Cai J, Gonzalez FJ (2021) The role of farnesoid X receptor in metabolic diseases, and gastrointestinal and liver cancer. Nat Rev Gastroenterol Hepatol 18:335–47. https://doi.org/10.1038/s41575-020-00404-2
Gardes C, Chaput E, Staempfli A, Blum D, Richter H, Benson GM (2013) Differential regulation of bile acid and cholesterol metabolism by the farnesoid X receptor in Ldlr-/- mice versus hamsters. J Lipid Res 54:1283–1299. https://doi.org/10.1194/jlr.M033423
Goodwin B, Jones SA, Price RR, Watson MA, McKee DD et al (2000) A regulatory cascade of the nuclear receptors FXR, SHP-1, and LRH-1 represses bile acid biosynthesis. Mol Cell 6:517–526. https://doi.org/10.1016/s1097-2765(00)00051-4
Inagaki T, Choi M, Moschetta A, Peng L, Cummins CL et al (2005) Fibroblast growth factor 15 functions as an enterohepatic signal to regulate bile acid homeostasis. Cell Metab 2:217–225. https://doi.org/10.1016/j.cmet.2005.09.001
Schoeler M, Caesar R (2019) Dietary lipids, gut microbiota and lipid metabolism. Rev Endocr Metab Disord 20:461–472. https://doi.org/10.1007/s11154-019-09512-0
Kawamata Y, Fujii R, Hosoya M, Harada M, Yoshida H et al (2003) A G protein-coupled receptor responsive to bile acids. J Biol Chem 278:9435–9440. https://doi.org/10.1074/jbc.M209706200
Castellanos-Jankiewicz A, Guzman-Quevedo O, Fenelon VS, Zizzari P, Quarta C et al (2021) Hypothalamic bile acid-TGR5 signaling protects from obesity. Cell Metab 33:1483–1492. https://doi.org/10.1016/j.cmet.2021.04.009
Thomas C, Gioiello A, Noriega L, Strehle A, Oury J et al (2009) TGR5-mediated bile acid sensing controls glucose homeostasis. Cell Metab 10:167–77. https://doi.org/10.1016/j.cmet.2009.08.001
Jia W, Xie G, Jia W (2018) Bile acid-microbiota crosstalk in gastrointestinal inflammation and carcinogenesis. Nat Rev Gastroenterol Hepatol 15:111–128. https://doi.org/10.1038/nrgastro.2017.119
Spivak I, Fluhr L, Elinav E (2022) Local and systemic effects of microbiome-derived metabolites. EMBO Rep 23:e55664. https://doi.org/10.15252/embr.202255664
Furusawa Y, Obata Y, Fukuda S, Endo TA, Nakato G et al (2013) Commensal microbe-derived butyrate induces the differentiation of colonic regulatory T cells. Nature 504:446–50. https://doi.org/10.1038/nature12721
Tan J, McKenzie C, Vuillermin PJ, Goverse G, Vinuesa CG et al (2016) Dietary fiber and bacterial SCFA enhance oral tolerance and protect against food allergy through diverse cellular pathways. Cell Rep 15:2809–24. https://doi.org/10.1016/j.celrep.2016.05.047
Goverse G, Molenaar R, Macia L, Tan J, Erkelens MN et al (2017) Diet-derived short chain fatty acids stimulate intestinal epithelial cells to induce mucosal tolerogenic dendritic cells. J Immunol 198:2172–2181. https://doi.org/10.4049/jimmunol.1600165
Nastasi C, Fredholm S, Willerslev-Olsen A, Hansen M, Bonefeld CM et al (2017) Butyrate and propionate inhibit antigen-specific CD8+ T cell activation by suppressing IL-12 production by antigen-presenting cells. Sci Rep 7:14516. https://doi.org/10.1038/s41598-017-15099-w
Xiu W, Chen Q, Wang Z, Wang J, Zhou Z (2020) Microbiota-derived short chain fatty acid promotion of Amphiregulin expression by dendritic cells is regulated by GPR43 and Blimp-1. Biochem Biophys Res Commun 533:282–288. https://doi.org/10.1016/j.bbrc.2020.09.027
Cai J, Sun L, Gonzalez FJ (2022) Gut microbiota-derived bile acids in intestinal immunity, inflammation, and tumorigenesis. Cell Host Microbe 30:289–300. https://doi.org/10.1016/j.chom.2022.02.004
Song X, Sun X, Oh SF, Wu M, Zhang Y et al (2020) Microbial bile acid metabolites modulate gut RORγ+ regulatory T cell homeostasis. Nature 577:410–415. https://doi.org/10.1038/s41586-019-1865-0
An D, Oh SF, Olszak T, Neves JF, Avci FY et al (2014) Sphingolipids from a symbiotic microbe regulate homeostasis of host intestinal natural killer T cells. Cell 156:123–133. https://doi.org/10.1016/j.cell.2013.11.042
Turner JR (2009) Intestinal mucosal barrier function in health and disease. Nat Rev Immunol 9:799–809. https://doi.org/10.1038/nri2653
Ghosh S, Whitley CS, Haribabu B, Jala VR (2021) Regulation of intestinal barrier function by microbial metabolites. Cell Mol Gastroenterol Hepatol 11:1463–82. https://doi.org/10.1016/j.jcmgh.2021.02.007
Kamada N, Seo SU, Chen GY, Nunez G (2013) Role of the gut microbiota in immunity and inflammatory disease. Nat Rev Immunol 13:321–335. https://doi.org/10.1038/nri3430
Clevers H (2013) The intestinal crypt, a prototype stem cell compartment. Cell 154:274–284. https://doi.org/10.1016/j.cell.2013.07.004
Zhang T, Li Q, Cheng L, Buch H, Zhang F (2019) Akkermansia muciniphila is a promising probiotic. Microb Biotechnol 12:1109–1125. https://doi.org/10.1111/1751-7915.13410
Groschwitz KR, Hogan SP (2009) Intestinal barrier function: molecular regulation and disease pathogenesis. J Allergy Clin Immunol 124:3–20. https://doi.org/10.1016/j.jaci.2009.05.038
Venkatesh M, Mukherjee S, Wang H, Li H, Sun K et al (2014) Symbiotic bacterial metabolites regulate gastrointestinal barrier function via the xenobiotic sensor PXR and Toll-like receptor 4. Immunity 41:296–310. https://doi.org/10.1016/j.immuni.2014.06.014
Grosheva I, Zheng D, Levy M, Polansky O, Lichtenstein A et al (2020) High-throughput screen identifies host and microbiota regulators of intestinal barrier function. Gastroenterology 159:1807–1823. https://doi.org/10.1053/j.gastro.2020.07.003
Leclercq S, Matamoros S, Cani PD, Neyrinck AM, Jamar F et al (2014) Intestinal permeability, gut-bacterial dysbiosis, and behavioral markers of alcohol-dependence severity. Proc Natl Acad Sci USA 111:E4485–E4493. https://doi.org/10.1073/pnas.1415174111
Cani P, Amar J, Iglesias M, Poggi M, Knauf C et al (2007) Metabolic endotoxemia initiates obesity and insulin resistance. Diabetes 56(7):1761–1772. https://doi.org/10.2337/db06-1491
Kort S, Keszthelyi D, Masclee A (2011) Leaky gut and diabetes mellitus: what is the link? Obes Rev 12(6):449–458. https://doi.org/10.1111/j.1467-789X.2010.00845.x
Cani P, Possemiers S, Wiele T, Guiot Y, Everard A et al (2009) Change in gut microbiota control inflammation in obese mice through a mechanism invoving GLP-2-driven improvement of gut permeability. Gut 58(8):1097–1103. https://doi.org/10.1136/gut.2008.165886
Brun P, Castagliuolo I, Leo V, Buda A, Pinzani M et al (2007) Increased intestinal permeability in obese mice: new evidence in the pathogenesis of nonalchoholic steatohepatis. Am J Physiol Gastrointest Liver Physiol 292(2):G518–G525. https://doi.org/10.1152/ajpgi.00024.2006
Massier L, Bluher M, Kovacs P, Chakaroun R (2021) Imparied intestinal barrier and tissue bacteria: pathomechanisms for metabolic diseases. Front Endocrinol 12:616506. https://doi.org/10.3389/fendo.2021.616506
Shen L, Ao L, Xu H, Shi J, You D et al (2019) Poor shor-term glycemic control in patients with type 2 diabetes imparis the intestinal mucosal barrier: a prospective, single-center, observational study. BMC Endocr Disord 19(1):29. https://doi.org/10.1186/s12902-019-0354-7
Natividad J, Lamas B, Pham H, Michel M, Rainteau D et al (2018) Bilophila wadsworthia aggravates high fat diet induced metabolic dysfunction in mice. Nat Commun 9(1):2802. https://doi.org/10.1038/s41467-018-05249-7
Kovtonyuk LV, Caiado F, Garcia-Martin S, Manz EM, Helbling P et al (2022) IL-1 mediates microbiome-induced inflammaging of hematopoietic stem cells in mice. Blood 139:44–58. https://doi.org/10.1182/blood.2021011570
Jenq RR, Ubeda C, Taur Y, Menezes CC, Khanin R et al (2012) Regulation of intestinal inflammation by microbiota following allogeneic bone marrow transplantation. J Exp Med 209:903–911. https://doi.org/10.1084/jem.20112408
Burgess SL, Leslie JL, Uddin J, Oakland DN, Gilchrist C et al (2020) Gut microbiome communication with bone marrow regulates susceptibility to amebiasis. J Clin Invest 130:4019–4024. https://doi.org/10.1172/JCI133605
Hill DA, Siracusa MC, Abt MC, Kim BS, Kobuley D et al (2012) Commensal bacteria-derived signals regulate basophil hematopoiesis and allergic inflammation. Nat Med 18:538–46. https://doi.org/10.1038/nm.2657
Hergott CB, Roche AM, Tamashiro E, Clarke TB, Bailey AG et al (2016) Peptidoglycan from the gut microbiota governs the lifespan of circulating phagocytes at homeostasis. Blood 127:2460–2471. https://doi.org/10.1182/blood-2015-10-675173
Balmer ML, Schurch CM, Saito Y, Geuking MB, Li H et al (2014) Microbiota-derived compounds drive steady-state granulopoiesis via MyD88/TICAM signaling. J Immunol 193:5273–5283. https://doi.org/10.4049/jimmunol.1400762
Enaud R, Prevel R, Ciarlo E, Beaufils F, Wieers G et al (2020) The gut-lung axis in health and respiratory diseases: A place for inter-organ and inter-kingdom crosstalks. Front Cell Infect Microbiol 10:9. https://doi.org/10.3389/fcimb.2020.00009
Dang AT, Marsland BJ (2019) Microbes, metabolites, and the gut-lung axis. Mucosal Immunol 12:843–850. https://doi.org/10.1038/s41385-019-0160-6
Pu Q, Lin P, Gao P, Wang Z, Guo K et al (2021) Gut microbiota regulate gut-lung axis inflammatory responses by mediating ILC2 compartmental migration. J Immunol 207:257–267. https://doi.org/10.4049/jimmunol.2001304
Steed AL, Christophi GP, Kaiko GE, Sun L, Goodwin VM et al (2017) The microbial metabolite desaminotyrosine protects from influenza through type I interferon. Science 357:498–502. https://doi.org/10.1126/science.aam5336
Trompette A, Gollwitzer ES, Yadava K, Sichelstiel AK, Sprenger N et al (2014) Gut microbiota metabolism of dietary fiber influences allergic airway disease and hematopoiesis. Nat Med 20:159–66. https://doi.org/10.1038/nm.3444
Albillos A, de Gottardi A, Rescigno M (2020) The gut-liver axis in liver disease: Pathophysiological basis for therapy. J Hepatol 72:558–77. https://doi.org/10.1016/j.jhep.2019.10.003
Carneiro C, Brito J, Bilreiro C, Barros M, Bahia C et al (2019) All about portal vein: a pictorial display to anatomy, variants and physiopathology. Insights Imaging 10:38. https://doi.org/10.1186/s13244-019-0716-8
Sannasiddappa TH, Lund PA, Clarke SR (2017) In vitro antibacterial activity of unconjugated and conjugated bile salts on staphylococcus aureus. Front Microbiol 8:1581. https://doi.org/10.3389/fmicb.2017.01581
Kandell RL, Bernstein C (1991) Bile salt/acid induction of DNA damage in bacterial and mammalian cells: implications for colon cancer. Nutr Cancer 16:227–38. https://doi.org/10.1080/01635589109514161
Inagaki T, Moschetta A, Lee YK, Peng L, Zhao G et al (2006) Regulation of antibacterial defense in the small intestine by the nuclear bile acid receptor. Proc Natl Acad Sci USA 103:3920–3925. https://doi.org/10.1073/pnas.0509592103
van Best N, Rolle-Kampczyk U, Schaap FG, Basic M, Olde Damink SWM et al (2020) Bile acids drive the newborn’s gut microbiota maturation. Nat Commun 11:3692. https://doi.org/10.1038/s41467-020-17183-8
Kim Y, Lee S, Kim S, Kim TY, Lee SH et al (2022) LKB1 in intestinal epithelial cells regulates bile acid metabolism by modulating FGF15/19 production. Cell Mol Gastroenterol Hepatol 13:1121–1139. https://doi.org/10.1016/j.jcmgh.2021.12.017
Han MM, Zhu XY, Peng YF, Lin H, Liu DC, Li L (2019) The alterations of gut microbiota in mice with chronic pancreatitis. Ann Transl Med 7:464. https://doi.org/10.21037/atm.2019.08.18
Kartal E, Schmidt TSB, Molina-Montes E, Rodriguez-Perales S, Wirbel J et al (2022) A faecal microbiota signature with high specificity for pancreatic cancer. Gut 71:1359–1372. https://doi.org/10.1136/gutjnl-2021-324755
Pushalkar S, Hundeyin M, Daley D, Zambirinis CP, Kurz E et al (2018) The pancreatic cancer microbiome promotes oncogenesis by induction of innate and adaptive immune suppression. Cancer Discov 8:403–416. https://doi.org/10.1158/2159-8290.CD-17-1134
Gomez-Hurtado I, Santacruz A, Peiro G, Zapater P, Gutierrez A et al (2011) Gut microbiota dysbiosis is associated with inflammation and bacterial translocation in mice with CCl4-induced fibrosis. PLoS One 6:e23037. https://doi.org/10.1371/journal.pone.0023037
Qin HL, Zheng JJ, Tong DN, Chen WX, Fan XB et al (2008) Effect of Lactobacillus plantarum enteral feeding on the gut permeability and septic complications in the patients with acute pancreatitis. Eur J Clin Nutr 62:923–30. https://doi.org/10.1038/sj.ejcn.1602792
Olah A, Belagyi T, Issekutz A, Gamal ME, Bengmark S (2002) Randomized clinical trial of specific lactobacillus and fibre supplement to early enteral nutrition in patients with acute pancreatitis. Br J Surg 89:1103–1107. https://doi.org/10.1046/j.1365-2168.2002.02189.x
Riquelme E, Zhang Y, Zhang L, Montiel M, Zoltan M et al (2019) Tumor microbiome diversity and composition influence pancreatic cancer outcomes. Cell 178:795–806. https://doi.org/10.1016/j.cell.2019.07.008
Funding
This research was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2022R1I1A1A01053022, 2021R1A6A1A03040260).
Author information
Authors and Affiliations
Contributions
Seungil Kim, Sang-Uk Seo, and Mi-Na Kweon wrote the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Kim, S., Seo, SU. & Kweon, MN. Gut microbiota-derived metabolites tune host homeostasis fate. Semin Immunopathol 46, 2 (2024). https://doi.org/10.1007/s00281-024-01012-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00281-024-01012-x