
Abstract
Animals often engage in contests over limited resources. The probability of winning a contest is primarily determined by the individual's fighting ability relative to its opponent and the perceived value of the disputed resource. Individuals of the hermit crab Calcinus californiensis frequently fight over gastropod shells. We conducted a free-choice experiment to assess the factors that determine individuals’ choice of opponent, contest initiation, and contest resolution. We placed hermit crabs occupying two shell species into a large container and monitored agonistic interactions between crabs. We assessed the asymmetries in fighting ability based on the differences in body mass between the opponents. The shell species and fit (i.e., shell size relative to crab body size) were used as measures of the objective and subjective resource value, respectively. Motivation influenced contest initiation; the crabs occupying too-tight shells were more likely to initiate a contest than the ones in looser-fitting shells. In most cases, the attackers fought for a shell with a looser fit, even if that meant losing a tighter-fitting shell of the preferred shell species. The fighting success for attackers was positively associated with the number of bouts of rapping and the shell size improvement. However, success was negatively correlated with body size asymmetry; attackers that chose opponents larger than themselves were more likely to evict their opponent than the attackers that chose opponents that were smaller. Experimental designs that allow animals to select their own opponents, rather than assigning specific opponents, can strongly contribute to knowledge of agonistic interactions.
Significance statement
Fighting dynamics are commonly assessed by estimating the influence of the resource-holding potential and resource value in pair-matched opponents. Here, however, we examined the influence of asymmetries in resource-holding potential and the objective and subjective value of gastropod shells on the choice of a contender and contest resolution through a free-choice opponent experiment in hermit crabs. The contest initiation was driven by the motivation to obtain a better gastropod shell species or one with a better size fit—factors which are known to improve individual fitness. Furthermore, fighting success was associated with an individual's persistence in displaying aggressive behaviors and its motivation to obtain a better resource. However, contrary to the predictions of game theoretical models, fighting success was higher when attackers chose opponents larger than themselves. Our results highlight the relevance of assessing fighting under more natural conditions by allowing animals to select their opponents.
Similar content being viewed by others

Introduction
Fighting can be highly costly in terms of risk of injury (Neat et al. 1998a), energy expenditure (Hack 1997), accumulation of metabolites (Briffa and Sneddon 2010; Ellington 1983), and decreased time available for other important activities (e.g., reproduction and foraging; Briffa and Sneddon 2010). Opponents commonly differ in fighting ability, motivation, and previous experience (which in turn can influence fighting ability and motivation); these asymmetries often determine the likelihood of success in fight encounters (Bradbury and Vehrencamp 2011). The differences in fighting ability (also known as resource holding potential; RHP) between opponents may result from asymmetries of conspicuous traits, such as body and weapon size, where opponents with larger body size and weapons are more likely to win a contest (Emlen 2014; Parker 1974; Vieira and Peixoto 2013), although inconspicuous RHP components (e.g., muscular strength and metabolic competence) and aggressiveness can also influence fight outcomes (Briffa and Sneddon 2007).
The value that individuals place on a disputed resource is also an important aspect in determining fighting dynamics and outcomes. The resource value (RV) is determined by the potential benefits that the contested resource may bring to the owner (Hurd 2006). Differences in motivation between opponents arise from the differential value that individuals place on the contested resource, which is determined by the individual’s requirements (Maynard and Parker 1976; Nosil 2002). Resource value determines an individual´s motivation, and it depends on the quality of the contested resource (objective resource value) or on the value that each contestant places on the resource (subjective resource value; Gherardi 2006). High motivation drives animals to pay high costs for fighting to obtain a required resource (Enquist and Leimar 1987) and can increase the probability that the highly motivated individual will win the fight (Brown et al. 2007; Dissanayake et al. 2009). As the motivational gap narrows between opponents, the role of RHP in shaping fighting dynamics becomes more pronounced. Conversely, when the RHP disparity between opponents decreases, the influence of motivation is expected to intensify (Humphries et al. 2006). Therefore, researchers often experiment with contest dynamics by pairing animals with specific RHP characteristics and allocating resources to opponents based on their perceived value to better understand the impact of RHP traits and motivational factors on contest initiation, escalation, and outcomes (Dowds and Elwood 1983; Mowles and Briffa 2012). When researchers pair opponents based on specific characteristics during typical paired staged encounters, conspicuous key elements influencing animals' decisions at different levels of the agonistic encounter may be overlooked. The animals may fight opponents with which they would not naturally compete. These staged encounter setups are valuable in that they allow the experimenter to generate specific predictions about contest dynamics and fight outcomes under controlled conditions; however, the overall view of the fighting dynamic in naturally matched individuals (free-choice) is rarely explored (for an exception, see O'Connor et al. 2015).
Hermit crabs serve as excellent biological models due to their unique dependence on gastropod shells, which are discrete units with easily identifiable attributes, including their absolute value (shell species) and subjective value (shell size relative to hermit crab body size). These attributes can be manipulated to test specific biological hypotheses. Hermit crabs are known to engage in contests for shells, primarily in intertidal environments where suitable shells are limited (Kellogg 1976). They generally cannot kill and remove dead gastropods from shells, so they rely on the availability of unoccupied shells (Hazlett 1981; for an exception see Alcaraz et al. 2020). During a fight, the attacker uses its legs and antennae to inspect the outside of a shell that interests it, and which is currently occupied by another crab (known as “shell exploring”). The attacker then aligns the aperture of its own shell with that of the opponent, which remains retracted into its shell, and inserts both chelae to grab its rival. Subsequently, the attacker repeatedly strikes its shell against that of the opponent (bouts of rapping). At the end of the encounter, the attacking crab either evicts its opponent and moves to a new, more suitable shell or loses the contest by retreating (Gherardi 2006). In contrast to other aggressive encounters where the winner claims the contested resource and the loser gets nothing, in hermit crabs, the winner acquires the better quality shell species or size that better fits its body, while the defeated crab ends up with the remaining shell, which is generally suboptimal. However, some hermit crab species can also engage in negotiations, in which case both opponents end up with a better shell than the one they originally occupied (Hazlett 1978).
Hermit crabs recognize and choose shell sizes and species that maximize one or several components of individual fitness, such as growth and reproduction (Osorno et al. 2005; Wada et al. 1997), protection against different predators (Alcaraz and Arce 2017), and the challenges of coping with wave action (Alcaraz et al., 2020). Accordingly, hermit crabs assign different absolute value to different shell species or types (e.g., with or without epibionts) and different subjective value based on the shell’s size. The absolute value that hermit crab species place on specific shell species and types can be identified by their preference, which may differ between sexes and ontogenetic stages (e.g., Briffa and Dallaway 2007; Straughan and Gosselin 2014). Meanwhile, the subjective resource value placed on the shells is strongly linked to the shell’s size relative to the crab’s body size (i.e., fit). The Shell Adequacy Index (SAI) can be used to estimate the subjective value that individuals place on the resource; this index is calculated as the ratio between the size of the shell occupied and the preferred shell size (estimated under controlled conditions; Asakura 1995). Hermit crabs place the greatest subjective resource value on the shell of the preferred size (SAI = 1), and the value decreases when the shell fit is tighter (SAI < 1) or looser (SAI > 1) relative to their body size. Therefore, fighting initiation, escalation, and outcome are influenced by a) the quality of the shell occupied, b) the shell quality of the potential opponent, and c) the potential gain if the attacker obtains the shell of the opponent (as proposed by Dowds and Elwood 1983).
The hermit crab Calcinus californiensis Bouvier, 1898 has shown a clear preference for certain shell species (Arce and Alcaraz 2012), most notably Stramonita biserialis Blainville, 1832, a species that it occupies frequently in the wild (Arce and Alcaraz 2011). By contrast, Nerita scabricosta Lamarck, 1822 is a gastropod shell that is non-preferred in laboratory tests but still commonly occupied in the field (Arce and Alcaraz 2011, 2012). The shells of S. biserialis are conical and relatively heavy with thick walls, whereas the shells of N. scabricosta are globose, relatively light, and thin-walled (Chávez-Solís and Alcaraz 2015). The shell preference of C. californiensis matches the degree of protection provided by each shell species against predators; crabs occupying shells of S. biserialis exhibit a higher survival in the presence of the shell peeler Arenaeus mexicanus than crabs occupying N. scabricosta shells (Arce and Alcaraz 2013). Calcinus californiensis also shows preference for shell size depending on their body size, but this may vary depending on the type of predator (shell breaking or peeler; Alcaraz and Arce 2017).
In the present study, we have investigated decision-making by the hermit crab C. californiensis during contests, from the choice of opponent to the contest outcome, in a free-choice scenario. We used the body mass asymmetry between the two opponents as the measure of RHP and the shell fit and species as estimators of the subjective and objective resource values, respectively. We hypothesized that if RHP is important during the choice of an opponent, the crabs would be more likely to initiate, escalate, and subsequently win more fights against individuals whose body mass is smaller than their own. Additionally, if motivation plays a role in the initiation, escalation, and resolution of fights, the attackers occupying lower-quality shells (suboptimal fit and/or non-preferred shell species) would have a higher probability of initiating a contest and be more likely to escalate and win fights than crabs possessing a better-quality resource.
Materials and methods
Hermit crab capture
The study was conducted during six consecutive days on the Pacific rocky shore of Troncones, Guerrero, Mexico. Each day, we captured 25 hermit crabs (C. californiensis) occupying shells of the preferred gastropod species S. biserialis and 25 occupying shells of the non-preferred species, N. scabricosta (Arce and Alcaraz 2011) without apparent damage or epibionts (i.e., a total of 300 hermit crabs). The hermit crabs were captured by hand along a line parallel to the coast during ebb tides in wave-protected sites.
Behavioral observations
Three observers recorded behaviors. The observer with most experience in scoring fighting behavior (more than five years) was responsible for training the other observers before the onset of observations. Inter-observer reliability (IR) was tested by asking each observer to score and record specific fighting behaviors in four pairs of hermit crabs and was calculated as (Number of agreements/ Total number of observations) × 100 (Kaufman and Rosenthal 2009). The tests did not start until IR was at least 90% in contests that escalated to shell exchange.
We attempted to evaluate the potential factors influencing a) likelihood to initiate an agonistic encounter (i.e., act as an “attacker”), b) the choice of opponent, c) escalation to shell rapping, and d) the contest resolution (winning the shell or giving up). We considered “shell exploration” (when the hermit crab used its legs and antennae to inspect the shell occupied by a congener), to mark the beginning of the agonistic interaction because it was the first evident sign of interest in the resource. In addition, shell exploration allowed us to identify which of the individuals that engaged in a contest was the attacker and which was the defender (i.e., which had initiated the contest). We considered escalation to shell rapping when an attacker aligned the aperture of its own shell with that of the opponent, inserting both chelae to grab its rival, and repeatedly struck its shell against that of the opponent. We recorded the number of bouts of rapping. The end of the contest was defined when the attacker either evicted its opponent or gave up (i.e., moved away from the defender) without obtaining the defender's shell. We also noted the contest duration from the recordings as an indicator of fighting assessment (Arnott and Elwood 2009). Only the attacker's behaviors were considered for statistical analyses.
General procedure
During each of the six trials (one per day), the hermit crabs (25 occupying N. scabricosta and 25 S. biserialis shells) were placed immediately after capture into individual opaque cylinders (0.05 L) to prevent physical interaction among crabs. Subsequently, the hermit crabs were transported to a laboratory where the cylinders were carefully submerged within a large experimental arena (80 × 50 × 30 cm) filled with natural seawater (34 PSU; 27 ± 1 °C). The water was constantly aerated. The bottom of the arena was covered with clean sand that had been previously washed. After one hour, the cylinders were removed to release the hermit crabs into the arena together.
The three observers simultaneously observed the hermit crabs, looking for the initiation of agonistic interactions between pairs (shell exploration). Once an agonistic interaction between two crabs had been identified, it was assigned to an observer who recorded the behaviors that occurred until the fight outcome was reached (contest resolution: eviction or non-eviction). After that, both crabs were carefully removed from the arena and placed in previously numbered glass flasks (0.05 L); the role played by the individual during the contest was recorded according to the behavior displayed (attacker or defender). At the end of each trial, which lasted a maximum of 3 h, all the hermit crabs were removed from the arena and placed in individual containers to be measured. The contests were videotaped using a Panasonic DMC-GH4 video camera. We then recorded from the videos the individuals that escalated to shell rapping, the number of bouts of rapping, contest duration, and contest outcome, using BORIS software (Friard and Gamba 2016).
Resource holding potential
At the end of the trials, each crab, whether or not it had participated in agonistic interactions, was evicted from its shell by gently heating the shell apex; this technique induces short-term effects and does not compromise the physical or behavioral integrity of individuals (Burciaga and Alcaraz 2023). Each crab was weighed on a balance plate (M; OHAUS, ± 0.01 g). For each pair of crabs that had engaged in a contest, the body size asymmetry was calculated as MAsy = ((M attacker—M defender) / ((M attacker + M defender)/2) 100, such that positive values of this index indicate that the attacker was heavier than the defender, while negative values indicate that the attacker was lighter than the defender (O’Connor et al. 2015). All hermit crabs were sexed. Females participating as attackers or defenders were excluded from the analyses because males and females can place different objective and subjective values on the shell species and fit (Briffa and Dallaway 2007; Suárez-Rodríguez et al., 2019), which would be difficult to analyze.
Resource value
The gastropod shells were weighed and total shell width and length were measured (± 0.1 mm). The shell fit for each hermit crab was quantified using the Shell Adequacy Index (SAI), calculated as the ratio between the mass of the shell occupied by the crabs in the wild and the mass of the shells preferred by the crabs under experimental conditions (Asakura 1995). An SAI = 1 indicates a shell that is of adequate size for the crab, SAI < 1 indicates that the crab occupies a shell smaller than the preferred size (tight fit), and SAI > 1 indicates that the crab occupies a larger shell than the preferred size (loose fit). The preferred size for N. scabricosta and S. biserialis shells (SAI = 1) was calculated for each individual using the equations that describe the shell mass preferred by C. californiensis as a function of hermit crab body mass (Arce and Alcaraz 2011). We considered SAI an indicator of resource value that could potentially influence contest initiation, escalation and contest resolution. Thus, we tested the potential influence of the SAI on:
-
1)
the likelihood to initiate a contest as a function of the shell fit of each shell species occupied by the attacker (N. scabricosta and S. biserialis; SAINer and SAIStr, respectively);
-
2)
the choice of an opponent by the attacker as a function of its shell fit (SAIAtt) and the SAI of the shell the attacker would attain if they evicted the defender (SAICon; SAI calculated using the defender’s shell and the attacker’s body mass); and
-
3)
the likelihood that the attacker would attain an eviction as a function of its motivation, potentially defined by: a) the fit of its initial shell (SAIAtt), b) the defender’s motivation to keep its shell (expressed as the potential improvement of the defender’s fit in the event of a shell exchange; SAIDef), and c) the potential improvement in shell fit if the attacker evicts the defender (shell adequacy improvement; SAIImp), estimated as: SAIImp = (|SAIAtt—1|—|SAIcon—1|). The magnitude of this index is the same regardless of the direction of the improvement; in other words, the degree of potential improvement does not distinguish between an attacker going from a too-tight shell (SAIAtt < 1) to a suitably sized shell or a too-loose shell (SAIAtt > 1) to a suitably sized shell.
Statistics
Comparison of the six trials
We compared the sex ratio of all crabs present in each trial, as well as the body mass and SAI of all of the males among the six trials to test for similarity within the samples. The number of males and females in the six trials were compared by a heterogeneity Chi-square test for goodness of fit (Zar 2010). The body mass and shell fit of the hermit crabs occupying N. scabricosta and S. biserialis shells (SAINer and SAIStr) were compared by two-way ANOVAs containing the shell species and trial as factors, and with body mass and SAI as the response variables. Since there were no significant differences among the six trials in the number of males and females, SAI, or male body mass, all data were pooled and analyzed as a single sample.
Factors determining the probability of initiating a contest
We tested the influence of fighting ability (RHP) and resource value (RV) on the probability that an individual would initiate a contest (i.e., become an attacker) using a GLM with a logit-link function and binomial distribution. We used each male’s body mass as an estimator of RHP and shell species (N. scabricosta or S. biserialis) and fit (SAINer and SAIStr, respectively) as predictor variables. The global model included the interaction of shell species and shell fit. Individuals that did not initiate a fight were scored as 0, and individuals that initiated a fight were scored as 1.
Choice of opponent
To assess whether the RHP influences the choice of the opponent, we compared the body mass of the attackers and defenders using a Wilcoxon matched pairs signed ranks test. The influence of objective RV on the choice of an opponent in terms of shell species was tested by comparing the number of fights initiated by hermit crabs occupying N. scabricosta (non-preferred species) and S. biserialis shells (preferred species; Arce and Alcaraz 2012), and by comparing the number of contests initiated by males occupying N. scabricosta and S. biserialis shells to obtain a shell of N. scabricosta or S. biserialis from the defenders using two different goodness of fit Chi-square tests.
The influence of subjective RV on the choice of opponent in terms of shell fit was estimated by comparing the shell fit of the attackers (SAIAtt) originally occupying N. scabricosta or S. biserialis shells and the shell fit that the attacker could attain from the defenders occupying shells of N. scabricosta or S. biserialis shells (SAICon) using Wilcoxon matched pairs signed ranks tests.
Escalation and contest resolution
The influence of resource holding potential (RHP) and resource value (RV) on escalation to shell rapping and contest resolution were tested using separate GLMs with a logit-link function and binomial distribution. For both models, we used as predictive variables the body size asymmetry between the opponents, the shell species of the attacker and the defender (N. scabricosta or S. biserialis), and the SAIAtt, SAIDef, and SAIImp. We also included the number of bouts of rapping as a predictive variable in the model of contest resolution. In the escalation to shell rapping model, individuals that did not escalate to a higher level of aggression were scored as 0, and individuals that escalated were scored as 1. In the contest resolution model, attackers that did not evict their opponents were scored as 0, and attackers that attained the eviction were scored as 1. The escalation model included the body size asymmetry by SAIImp interaction term, while the contest resolution model included two interaction terms: body size asymmetry by SAIImp and body size asymmetry by number of bouts of rapping.
For all of the final models, we calculated the explained deviance as a measure of fit (Zuur et al. 2009). We then performed a model selection using the ‘drop1’ function (‘stats’ R package). The assumptions necessary for binomial logistic regression (independence of variables, absence of multicollinearity, linearity of independent variables and log odds) were verified by examining residual plots (“performance” R package; Lüdecke et al. 2021). All statistical analyses were performed in R, version 4.2.0 (R Core Team 2022).
Results
Comparison of the six trials
Of the 300 hermit crabs used in the study, seventy-one were excluded from the analyses because they were females, and nine of them had participated in contests with males. Consequently, these contests were also excluded from the study. The number of male:male contests was 54. The number of males and females was not significantly different among the six trials (3:1; Chi-square test: χ25 = 8.77, P = 0.10). The body mass of the males did not differ across the six trials, regardless of the shell species occupied (Two-way ANOVA, trial by shell species interaction term: F5,272 = 0.78, P = 0.57). The body mass of the males did not differ between the shell species occupied (F1,272 = 0.08, P = 0.78). The shell adequacy index of the males did not differ across the six trials, regardless of the shell species occupied (Two-way ANOVA, trial by shell species interaction term: F5,272 = 1.90, P = 0.10). However, the SAI of hermit crabs occupying S. biserialis was lower than the ones occupying N. scabricosta (SAIStr = 0.58 ± 0.021, SAINer = 0.88 ± 0.023; Two-way ANOVA, shell species main effect: F1,272 = 119.81, P < 0.001). Since there was no significant variation of body size, sex proportion, and SAI among trials, the six sets of data were pooled for all subsequent analyses.
Factors determining the probability to initiate a contest
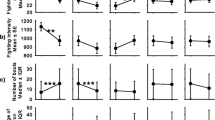
The selected model assessing the influence of the resource holding potential (RHP) and the resource value (RV) on initiating shell fighting explained 52% of the variation. Attackers showed a tendency to be more likely to initiate a contest when the opponent was smaller than themselves; however, this effect was not significant (body size asymmetry; GLM: χ2 1,229 = 3.14, P = 0.08). The interaction between the shell species occupied (N. scabricosta or S. biserialis) and the fit of those shells (SAIStr and SAINer) influenced the probability of initiating a contest (GLM: χ2 1,229 = 7.28, P = 0.007). Individuals occupying tighter shells showed a higher probability to engage in shell fighting than individuals in shells with a loose fit in shells of either species, but attackers occupying S. biserialis required a tighter shell than those in N. scabricosta shells to start a fight (Fig. 1).

Effect of the interaction of shell species and Shell Adequacy Index (SAIStr and SAINer) on the probability to initiate a fight as attackers. Males initially occupying Stramonita biserialis and Nerita scabricosta shells are represented in black and blue, respectively
Choice of opponent
The body mass of the attacker and defenders did not differ (Wilcoxon; W = 819.5, P = 0.62). The body mass asymmetry within fighting pairs ranged from attackers being 25% smaller than defenders to attackers being 20% larger than the defenders, while approximately 80% of the attackers chose an opponent within 10% of their body size (smaller or larger, Fig. 2).

Relative difference in the body mass of attackers and defenders (body mass assymmetry). Positive values indicate contests where the attackers are heavier than defenders, and negative values indicate contests where the attackers are lighter than defenders
In terms of the influence of the objective RV on the choice of an opponent, the number of contests initiated by hermit crabs occupying N. scabricosta shells (non-preferred species; n = 34) did not differ from those initiated by crabs in S. biserialis (preferred species; n = 20; χ21 = 0.78, P = 0.30). However, more crabs initiated agonistic encounters against opponents occupying N. scabricosta (n = 42) than against those in S. biserialis shells (n = 12; χ21 = 9.51, P < 0.01; Fig. 3a), independently of the attacker’s initial shell species (χ21 = 5.75, P = 0.30).

a Shell species occupied by attackers that initiated a contest and the defenders they chose. “NN”, attackers and defenders occupying shells of N. scabricosta; "NS", attackers in N. scabricosta and defenders in S. biserialis; “SN”, attackers in S. biserialis and defenders in N. scabricosta; and “SS”, attackers and defenders in S. biserialis. b Shell fit of the attacker (SAIAtt) and potential fit if the attacker obtained the contested shell from the defender (SAICon). Mean values ± SE. * Significant difference (P < 0.05)
The attackers occupying an N. scabricosta shell that fought against defenders in an N. scabricosta shell chose opponents occupying a shell that offered a better size fit than the attacker’s initial shell (Wilcoxon: W = 87, P = 0.01). The attackers occupying an N. scabricosta with defenders in an S. biserialis shell chose opponents whose shell fit was less adequate than the fit of their own shell (Wilcoxon: W = 34, P = 0.02; Fig. 3b). The attackers in S. biserialis with opponents in N. scabricosta chose opponents with a shell that promised a better fit than that of their current shell (Wilcoxon: W = 2, P < 0.01), and the attackers in S. biserialis chose opponents occupying S. biserialis whose shells were neither better nor worse than that of their own shell (Wilcoxon: W = 10, P = 0.58; Fig. 3b).
Escalation and Contest Resolution
The model selection assessing the influence of RV and RHP on escalation to bouts of rapping is shown in Table 1. The selected model explained 39% of the deviance. The escalation to shell rapping was not explained by shell species, SAIAtt, SAIDef, or SAIImp (Table 1). The body size asymmetry influenced the escalation to bouts of rapping. The probability of escalation to shell rapping increased as the body size asymmetry was increasingly positive (i.e., the attacker had higher body mass relative to the defender; Fig. 4).

Effect of body size asymmetry on the probability of escalation to shell rapping. Some data are displaced from their binary positions for visual clarity (random jitter). Body size asymmetry equal to zero indicates that the attacker and defender were of similar body size
The model selection assessing the influence of RV, RHP, contest duration, and the number of bouts of rapping on contest resolution is shown in Table 2. The selected model explained 71% of the deviance. Contest resolution was not influenced by shell species, SAIAtt, SAIDef, or contest duration (Table 2). The success in the fight was determined by SAIImp, body size asymmetry, and the number of bouts of rapping. The attackers that achieved the eviction improved their shell fit (SAIImp = 0.16 ± 0.11; Fig. 5a). Contrary to expectation, attackers that were smaller than their opponents showed a greater probability of winning a contest than did the attackers that were larger than their opponents (Fig. 5b). Additionally, the attackers that performed a greater number of bouts of rapping had a large chance of winning a contest (Fig. 5c).

Probability of winning a contest as a function of a) the Shell Adequacy Improvement (SAIImp), b the body mass asymmetry between contestants, and c) the number of bouts of rapping. Some data are displaced from their binary positions for visual clarity (random jitter). Body size asymmetry equal to zero indicates that the attacker and defender were of similar body size
Discussion
The species and size of the shells hermit crabs occupy are important for their fitness (Reese 1969); however, hermit crabs are commonly found occupying suboptimal shells. We predicted that the hermit crabs occupying shells with an inadequate fit or of the non-preferred shell species (N. scabricosta) would initiate more agonistic encounters than those occupying shells of a more adequate size or the preferred species (S. biserialis). In accordance with these predictions, individuals occupying tighter shells showed a higher probability of engaging in fighting than crabs occupying shells with a looser fit (see Fig. 1). However, the hermit crabs occupying the preferred and the non-preferred shell species showed the same probability of initiating a fight. Further, crabs occupying shells of N. scabricosta and S. biserialis chose with higher frequency opponents occupying the non-preferred shell (N. scabricosta) rather than the preferred shell (S. biserialis). The behavioral preference for specific resources (e.g. food, habitat, mate, and gastropod shells in hermit crabs) is commonly an adaptive response that results in maximizing one or more fitness components (e.g. Lemire and Himmelman 1996; Martin 1998). Therefore, it is expected that individuals initiate and escalate in a contest to obtain the preferred shell species and size. Although S. biserialis gives more protection against predators than other shell species commonly found at Troncones, including N. scabricosta (Arce and Alcaraz 2013), shell fit also influences the fitness of hermit crabs, since a tight shell will decrease the growth rate (Bertness 1981), and will hinder the full retreat of the hermit crab inside, thereby increasing its vulnerability to predators, regardless of the shell species occupied (Arce and Alcaraz 2013). Calcinus californiensis is found in the wild occupying tighter shells of S. biserialis than of N. scabricosta (SAI = 0.58 and 0.87, respectively). Therefore, motivation to improve shell fit can explain why the hermit crabs occupying the preferred shell S. biserialis engaged in agonistic encounters, as well as why these crabs so frequently chose opponents occupying the non-preferred shell species from which they can obtain a better fit. It is particularly interesting that the crabs that were occupying the preferred shell species and chose opponents in the non-preferred shell were the ones that had the tightest shells, showing that these crabs were willing to obtain a larger shell at the cost of losing the advantages granted by the preferred species. On the other hand, some hermit crabs in N. scabricosta chose opponents in the preferred shell species even when this implied a fight for a shell that would fit them less well. Alternatively, most of the attackers (80%) choosing opponents occupying Nerita shells could also be attributed to the internal configuration of nerites (reduced columella), which reduces the owner’s ability to resist eviction (Burciaga and Alcaraz 2023); therefore, by choosing defenders in Nerita shells, the attackers decrease their fighting costs and increase their likelihood of obtaining a new shell.
In several groups of animals most of the fighting is commonly initiated by the larger individuals, including hermit crabs (e.g., 93%; Pagurus bernhardus; Dowds and Elwood 1983). However, in hermit crabs under pair-matched experimental conditions, researchers often induce a larger opponent to attack by providing it with a shell that is too tight, while the smaller opponent occupies a shell of adequate fit. In contrast, in our free choice experiment, 50% of the attackers chose a larger opponent than themselves, being that the body size of the attackers varied from 25% smaller to 20% larger than the defenders.
In Pagurus bernhardus, most of the contests were initiated by the larger individual (Dowds and Elwood 1983), whose decision to fight was not influenced by the shell species occupied by the smaller crab of the pair. However, interestingly, the relatively small proportion of conflicts that were initiated by the smaller opponent (11%) were all initiated by crabs occupying the non-preferred shell (Gibbula spp.; Dowds and Elwood (1983). Similarly, a high proportion of small opponents of C. californiensis (40%), which previously occupied non-preferred shells (too heavy), initiated fights against larger individuals occupying the preferred shell (S. biserialis; Alcaraz and Jofre 2017). Likely, in natural encounters, a higher proportion of fights than expected by theory can be initiated by smaller contestants. Few studies assess the fighting dynamics of hermit crabs occupying different shell species, and the opportunity to obtain the preferred shell could likely be a strong stimulus for small individuals to fight against larger opponents.
In this study, the body size asymmetry between opponents was important in determining escalation and fighting outcome. We found that body size asymmetry had contrasting effects on the escalation and resolution of contests. Attackers that were larger than their opponents were more likely to escalate to bouts of rapping, but less likely to win the contest. According to Dowds and Elwood (1983), smaller opponents initiate contests against larger ones, possibly due to an inaccurate assessment of RHP. However, in the case of C. californiensis, attackers that investigated a smaller opponent and its shell were likely to discover that the smaller defender’s shell was too small and decide to withdraw without rapping. It is conceivable that animals gather information throughout the progression of the contest and make a complete RHP assessment after inspecting the interior of the shell before engaging in bouts of rapping.
In contrast to several studies where the smaller contestant has a lower probability to win a fight (including hermit crabs; e.g., Hazlett 1970), we found that an attacker smaller than the defender was more likely to win the contest than was an attacker that was larger than the defender (see Fig. 5b). This is a surprising result since, in several animals, even small differences in size between individuals bias the outcome in favor of the larger contestant, such as in cichlid fishes (Neat et al.1998b), three-spined sticklebacks (Rowland 1989), fiddler crabs (Jennions and Backwell 1996), and crayfish (Pavey and Fielder 1996). In the crayfish Procambarus clarkii, a difference of body size and mass as small as 3% is enough to favor the success of the larger contestant in 80% of the encounters (Ueno and Nagayama 2012). There are several possible reasons why some individuals defeat larger opponents in a contest. An increase in aggression and fighting success of relatively smaller individuals can be expected when the difference in body size between opponents is small, and the subjective resource value is higher for the individual at an apparent disadvantage (Morrell et al. 2005; Tinnesand et al. 2013). In those cases, a high motivation to fight can overcome small differences in individual RHP (Barnard and Brown 1984). For instance, in the hermit crab Pagurus longicarpus, asymmetries in motivation allow smaller contestants to defeat a larger opponent and occupy a better shell (Gherardi 2006), and in C. californiensis, individuals that occupied broken shells in the wild with lower muscular strength won contests against stronger defenders with higher RHP (Alcaraz and Jofre 2017). In the present study, the subjective RV regarding the potential improvement in shell fit (SAIImp) explained the overall fighting success of the attackers; that is, the attackers that would potentially get a shell of better fit (with higher motivation) were the ones that had a higher probability of winning, regardless of whether they were larger or smaller than their opponents. However, the subjective RV did not explain why smaller attackers defeated larger defenders, since the interaction of these variables was not significant.
The success of the contestants that engage in aggressive interactions against larger opponents could also be explained by negotiation (Doake and Elwood 2011). However, contrary to other studies in which both opponents gain in RV (Hazlett 1983, 1996), in this study the defenders that were evicted were left with a shell that had a poorer fit than their original shell. This was especially true for defenders that were larger than the attacker, as their SAI decreased more on average (ΔSAI Def = -0.26 ± 0.09) than the improvement in shell fit gained by their smaller attackers (SAIImp = 0.16 ± 0.11). A potential attacker that investigates a smaller defender will probably discover that its shell will be too tight a fit; thus, it would be expected to give up without rapping. On the contrary, a hermit crab that investigates the shell of a larger defender would likely find that it has a relatively large shell, and thus, it would attempt to take it. In other words, in contrast to other biological groups, hermit crabs are unlikely to gain much benefit from fighting against smaller opponents, since even if they could easily win the resource, the resource has low subjective value.
The fighting success of the attacker with body size disadvantage could also be attributed to an inconspicuous asymmetry in RHP. Despite our awareness of body size and armor as key factors determining the RHP, understanding of the contribution of physical and physiological factors to the resolution of animal conflicts is limited (e.g. Humphries et al. 2006; O’Connor et al. 2015). Shell rapping is energetically demanding (Briffa and Elwood 2002, 2005) and is a strong predictor of fighting success (Briffa and Elwood 2000). The success of the smaller attackers could be associated with their ability to persist in a contest performing bouts of rapping against a larger defender. However, although fighting success was correlated with body size asymmetry and with the number bouts, these variables did not interact, i.e. the smaller attackers that evicted their opponents were not the ones that displayed more bouts of rapping. Therefore, the number of bouts was a good predictor of contest success, but it did not explain the success of the attackers at a body size disadvantage.
We expected that the longer the attacker spent attempting to evict the defender, the higher the likelihood that it would be successful, since energy expenditure thresholds could be determinant for the decision to persist in a contest in C. californiensis, as in other hermit crab species (e.g., P. bernhardus; Briffa and Elwood 2002). However, although there is a positive trend toward increased probability of winning with increasing contest duration (winners: \(\overline{{\text{X}} }\) = 1684 s ± 403.43 s.e.; losers: \(\overline{{\text{X}} }\) = 322 s ± 56.67 s.e.), this trend was not statistically significant (X2 = 2.81, P = 0.10). This trend could also reflect a loss of motivation on the part of the attackers, as they realize that the opponent's shell is not suitable for them. Therefore, they decide to withdraw from the fight early.
More accurate predictors of fighting success, such as the number of raps per bout and the force with which shells are rapped (Briffa and Elwood 2000), should be explored to explain the success of the smaller contestants. Additionally, the influence of strength and persistence-related individual traits are not clearly understood yet (Vieira and Peixoto 2013); inconspicuous asymmetries associated with a functional performance (e.g. metabolic competence and muscular strength) could be favoring the fighting success of some crabs despite a body size disadvantage. Inconspicuous asymmetries in strength-related traits could play an important role in shell fighting, as suggested by Vieira and Peixoto (2013), for encounters depending on physical contact. Some animals can advertise the RHP of the opponents in terms of strength, stamina, energy reserves and/or metabolic costs accrued early in the conflict, which might inform them about their chances to win a conflict with high reliability (e.g. Marden and Rollins 1994; Mowles and Briffa 2012). In cichlids, the eventual winners and losers behave differently before beginning a fight (Maan et al. 2001), while in hermit crabs the frequency of cheliped flicks during prefight assessment strongly predict the fighting outcome (Mowles and Briffa 2012). Calcinus californiensis might be gathering information about its chances to win by assessing inconspicuous asymmetries during early stages of the interaction since all the attackers smaller than the defenders that escalated to bouts of rapping evicted their opponents. The attackers seem to escalate to shell rapping against larger defenders only when they have a high probability to win. The traits that allow weight-disadvantaged individuals to initiate and win a contest remain to be evaluated.
Scenarios where animals freely choose their opponents do not allow rigorous control of variables, and hence make interpretation more difficult; however, this approach permits a more integrative understanding of individuals’ decision making. Therefore, despite the considerable understanding of the rules governing animal conflicts, the reliability of the sequence of individuals’ decisions in most natural conditions and its actual consequences in the outcome of the fight require an assessment (Humphries et al. 2006; O’Connor et al. 2015). Our findings highlight the complex interaction between objective and subjective resource value in initiating a fight and that smaller opponents can initiate and win more contests than expected based on results in controlled dyad experiments. Our study shows that agonistic encounters consist of an inseparable sequence of events that require evaluation as a whole.
Data availability
The datasets generated during the current study are available in the FigShare repository, https://doi.org/10.6084/m9.figshare.22972988.
References
Alcaraz G, Arce E (2017) Predator discrimination in the hermit crab Calcinus californiensis: tight for shell breakers, loose for shell peelers. Oikos 126:1–9. https://doi.org/10.1111/oik.03742
Alcaraz G, Jofre GI (2017) Aggressiveness compensates for low muscular and metabolic disadvantages in shell fighting: an outcome of the individual´s past. Behav Ecol Sociobiol 71:1–11. https://doi.org/10.1007/s00265-017-2311-7
Alcaraz G, Toledo B, Burciaga LM (2020) The energetic costs of living in the surf and impacts on zonation of shells occupied by hermit crabs. J Exp Biol 223:jeb222703. https://doi.org/10.1242/jeb.222703
Arce E, Alcaraz G (2011) Shell use by the hermit crab Calcinus californiensis at different levels of the intertidal zone. Sci Mar 75:121–128. https://doi.org/10.3989/scimar.2011.75n1121
Arce E, Alcaraz G (2012) Shell preference in a hermit crab: comparison between a matrix of paired comparisons and a multiple-alternative experiment. Mar Biol 159:853–862. https://doi.org/10.1007/s00227-011-1861-x
Arce E, Alcaraz G (2013) Plasticity of shell preference and its antipredatory advantages in the hermit crab Calcinus californiensis. Can J Zool 91:321–327. https://doi.org/10.1139/cjz-2012-0310
Arnott G, Elwood RW (2009) Assessment of fighting ability in animal contests. Anim Behav 77:991–1004. https://doi.org/10.1016/j.anbehav.2009.02.010
Asakura A (1995) Sexual differences in life history and resource utilization by the hermit crab. Ecology 76:2295–2313
Barnard CJ, Brown CAJ (1984) A payoff asymmetry in resident-resident disputes between shrews. Anim Behav 32:302–304. https://doi.org/10.1016/S0003-3472(84)80357-7
Bertness MD (1981) The influence of the shell-type on hermit crab growth rate and clutch size (Decapoda, Anomura). Crustaceana 40:197–205
Bradbury JW, Vehrencamp SL (2011) Principles of animal communication. Sinauer Associates Inc, Massachusetts
Briffa M, Dallaway D (2007) Inter-sexual contests in the hermit crab Pagurus bernhardus: females fight harder but males win more encounters. Behav Ecol Sociobiol 61:1781–1787. https://doi.org/10.1007/s00265-007-0411-5
Briffa M, Elwood RW (2000) Cumulative or sequential assessment during hermit crab shell fights: effects of oxygen on decision rules. Proc R Soc Lon B 267:2445–2452. https://doi.org/10.1098/rspb.2000.1304
Briffa M, Elwood RW (2002) Power of shell-rapping signals influences physiological costs and subsequent decisions during hermit crab fights. Proc R Soc Lon B 269:2331–2336. https://doi.org/10.1098/rspb.2002.2158
Briffa M, Elwood RW (2005) Rapid change in energy status in fighting animals: causes and effects of strategic decisions. Anim Behav 70:119–124. https://doi.org/10.1016/j.anbehav.2004.10.013
Briffa M, Sneddon LU (2007) Physiological constraints on contest behaviour. Funct Ecol 21:627–637. https://doi.org/10.1111/j.1365-2435.2006.01188.x
Briffa M, Sneddon LU (2010) Contest behavior. In: Fox CW, Westneat DF (eds) Evolutionary Behavioral Ecology. Oxford University Press, Oxford, pp 246–265
Brown WD, Chimenti AJ, Siebert JR (2007) The payoff of fighting in house crickets: motivational asymmetry increases male aggression and mating success. Ethology 113:457–465. https://doi.org/10.1111/j.1439-0310.2007.01357.x
Burciaga LM, Alcaraz G (2023) Metabolic and behavioral effects of hermit crab shell removal techniques: is heating less invasive than cracking? Anim Welf 32(e24):1–11. https://doi.org/10.1017/awf.2023.17
Chávez-Solís C, Alcaraz G (2015) Hiding time of the hermit crab Calcinus californiensis: observations from different shell types. Can J Zool 93:559–565. https://doi.org/10.1139/cjz-2014-0312
Dissanayake A, Galloway TS, Jones MB (2009) Physiological condition and intraspecific agonistic behaviour in Carcinus maenas (Crustacea: Decapoda). J Exp Mar Biol Ecol 375:57–63. https://doi.org/10.1016/j.jembe.2009.05.007
Doake S, Elwood RW (2011) How resource quality differentially affects motivation and ability to fight in hermit crabs. Proc R Soc Lon B 278:567–573. https://doi.org/10.1098/rspb.2010.1418
Dowds BM, Elwood RW (1983) Shell wars: assessment strategies and the timing of decisions in hermit crab shell fights. Behaviour 85:1–24
Ellington WR (1983) The recovery from anaerobic metabolism in invertebrates. J Exp Zool 228:431–444. https://doi.org/10.1002/jez.1402280305
Emlen DJ (2014) Animal weapons the evolution of battle. Henry Holt and Company, New York, pp 53–70
Enquist M, Leimar O (1987) Evolution of fighting behaviour: the effect of variation in resource value. J Theor Biology 127:187–205. https://doi.org/10.1016/S0022-5193(87)80130-3
Friard O, Gamba M (2016) BORIS: a free, versatile open-source event-logging software for video/audio coding and live observations. Methods Ecol Evol 7:1325–1330. https://doi.org/10.1111/2041-210x.12584
Gherardi F (2006) Fighting behavior in hermit crabs: the combined effect of resource-holding potential and resource value in Pagurus longicarpus. Behav Ecol Sociobiol 59:500–510. https://doi.org/10.1007/s00265-005-0074-z
Hack MA (1997) Assessment strategies in the contests of male crickets, Acheta domesticus (L.). Anim Behav 53:733–747. https://doi.org/10.1006/anbe.1996.0310
Hazlett BA (1970) The effect of shell size and weight on the agonistic behavior of a hermit crab. Z Tierpsychol 27:369–374. https://doi.org/10.1111/j.1439-0310.1970.tb01878.x
Hazlett BA (1978) Shell exchanges in hermit crabs: aggression, negotiation, or both? Anim Behav 26:1278–1279. https://doi.org/10.1016/0003-3472(78)90123-9
Hazlett BA (1981) The behavioral ecology of hermit crabs. Annu Rev Ecol Evol Syst 12:1–22. https://doi.org/10.1146/annurev.es.12.110181.000245
Hazlett BA (1983) Interspecific negotiations: mutual gain in exchanges of a limiting resource. Anim Behav 31:160–163
Hazlett BA (1996) Assessments during shell exchanges by the hermit crab Clibanarius vittatus: The complete negotiator. Anim Behav 51:567–573. https://doi.org/10.1006/anbe.1996.0060
Humphries EL, Hebblethwaite AJ, Batchelor TP, Hardy ICW (2006) The importance of valuing resources: host weight and contender age as determinants of parasitoid wasp contest outcomes. Anim Behav 72:891–898. https://doi.org/10.1016/j.anbehav.2006.02.015
Hurd PL (2006) Resource holding potential, subjective resource value, and game theoretical models of aggressiveness signalling. J Theor Biol 241:639–648. https://doi.org/10.1016/j.jtbi.2006.01.001
Jennions MD, Backwell PRY (1996) Residency and size affect fight duration and outcome in the fiddler crab Uca annulipes. Biol J Linn Soc 57:293–306. https://doi.org/10.1111/j.1095-8312.1996.tb01851.x
Kaufman AB, Rosenthal R (2009) Can you believe my eyes? The importance of interobserver reliability statistics in observations of animal behaviour. Anim Behav 78:1487–1491. https://doi.org/10.1016/j.anbehav.2009.09.014
Kellogg CW (1976) Gastropod shells: a potentially limiting resource for hermit crabs. J Exp Mar Biol Ecol 22:101–111. https://doi.org/10.1016/0022-0981(76)90112-X
Lemire M, Himmelman JH (1996) Relation of food preference to fitness for the green sea urchin, Strongylocentrotus droebachiensis. Mar Biol 127:73–78. https://doi.org/10.1007/BF00993646
Lüdecke D, Ben-Shachar MS, Patil I, Waggoner P, Makowski D (2021) Performance: An R package for assessment, comparison and testing of statistical models. J Open Source Softw 6:3139. https://doi.org/10.21105/joss.03139
Maan ME, Groothuis TGG, Wittenberg J (2001) Escalated fighting despite predictors of conflict outcome: solving the paradox in a South American cichlid fish. Anim Behav 62:623–634. https://doi.org/10.1006/anbe.2001.1819
Marden JH, Rollins RA (1994) Assessment of energy reserves by damselflies engaged in aerial contests for mating territories. Anim Behav 48:1023–1030. https://doi.org/10.1006/anbe.1994.1335
Martin TE (1998) Are microhabitat preferences of coexisting species under selection and adaptive? Ecology 79:656–670. https://doi.org/10.1890/0012-9658(1998)079[0656:AMPOCS]2.0.CO;2
Maynard S, Parker GA (1976) The logic of asymmetric contests. Anim Behav 24:159–175. https://doi.org/10.1016/S0003-3472(76)80110-8
Morrell LJ, Backwell PRY, Metcalfe NB (2005) Fighting in fiddler crabs Uca mjoebergi: what determines duration? Anim Behav 70:653–662. https://doi.org/10.1016/j.anbehav.2004.11.014
Mowles SL, Briffa M (2012) Forewarned is forearmed: early signals of RHP predict opponent fatigue in hermit crab shell fights. Behav Ecol 23:1324–1329. https://doi.org/10.1093/beheco/ars124
Neat FC, Huntingford FA, Beveridge MM (1998) Fighting and assessment in male cichlid fish: the effects of asymmetries in gonadal state and body size. Anim Behav 55:883–891
Neat FC, Taylor AC, Huntingford FA (1998) Proximate costs of fighting in male cichlid fish: the role of injuries and energy metabolism. Anim Behav 55:875–882. https://doi.org/10.1006/anbe.1997.0668
Nosil P (2002) Food fights in house crickets, Acheta domesticus, and the effects of body size and hunger level. Can J Zool 80:409–417. https://doi.org/10.1139/Z02-018
O’Connor CM, Reddon AR, Ligocki IY, Hellmann JK, Garvy KA, Marsh-Rollo SE, Balshine S (2015) Motivation but not body size influences territorial contest dynamics in a wild cichlid fish. Anim Behav 107:19–29. https://doi.org/10.1016/j.anbehav.2015.06.001
Osorno JL, Contreras-Garduño J, Macías-Garcia C (2005) Long-term costs of using heavy shells in terrestrial hermit crabs (Coenobita compressus) and the limits of shell preference: An experimental study. J Zool 266:377–383. https://doi.org/10.1017/S0952836905007028
Parker GA (1974) Assessment strategy and the evolution of fighting behaviour. J Theor Biol 47:223–243. https://doi.org/10.1016/0022-5193(74)90111-8
Pavey CR, Fielder DR (1996) The influence of size differntial on agonistic behaviour in the freshwater crayfish, Cherax cuspidatus (Decapoda: Parastacidae). J Zool 238:445. https://doi.org/10.1111/j.1469-7998.1996.tb05405.x
Reese ES (1969) Behavioral adaptations of intertidal hermit crabs. Amer Zool 9:343–355
R Core Team (2022) R: a language and environment for statistical computing. R Foundation for Statistical Computing Vienna, Austria. https://www.R-project.org/
Rowland WJ (1989) The effects of body size, aggression and nuptial coloration on competition for territories in male threespine sticklebacks, Gasterosteus aculeatus. Anim Behav 37:282–289. https://doi.org/10.1016/0003-3472(89)90117-6
Straughan NA, Gosselin LA (2014) Ontogenetic changes in shell preferences and resource partitioning by the hermit crabs Pagurus hirsutiusculus and P.granosimanus. J Exp Mar Biol Ecol 451:1–8. https://doi.org/10.1016/j.jembe.2013.10.028
Tinnesand HV, Jojola S, Zedrosser A, Rosell F (2013) The smell of desperadoes? Beavers distinguish between dominant and subordinate intruders. Behav Ecol Sociobiol 67:895–904. https://doi.org/10.1007/s00265-013-1512-y
Ueno R, Nagayama T (2012) Interlocking of chelae is a key factor for dominance hierarchy formation in crayfish. J Exp Biol 215:2841–2848. https://doi.org/10.1242/jeb.072520
Vieira MC, Peixoto PE (2013) Winners and losers: a meta-analysis of functional determinants of fighting ability in arthropod contests. Funct Ecol 27:305–313. https://doi.org/10.1111/1365-2435.12051
Wada S, Ohmori H, Goshima S, Nakao S (1997) Shell-size preference of hermit crabs depends on their growth rate. Anim Behav 54:1–8. https://doi.org/10.1006/anbe.1996.0319
Zar JH (2010) Biostatistical analysis. Pearson Education, New Jersey
Zuur AF, Ieno EN, Walker NJ, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Acknowledgements
This study was supported by Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT, IN-218321). We thank Dr. Mark Briffa and Dr. Robert Elwood for their valuable suggestions on this manuscript. We also thank Dr. Karla Kruesi and Dr. Olivia de los Santos for their technical assistance, and Dr. Lynna Kiere for editing the English.
Author information
Authors and Affiliations
Contributions
Conceptualization: Luis M. Burciaga and Guillermina Alcaraz; Methodology: Luis M. Burciaga and Guillermina Alcaraz; Formal analysis and investigation: Luis M. Burciaga and Guillermina Alcaraz; Writing—original draft preparation: Luis M. Burciaga; Writing—review and editing: Luis M. Burciaga and Guillermina Alcaraz; Funding acquisition: Guillermina Alcaraz; Resources: Guillermina Alcaraz; Supervision: Guillermina Alcaraz.
Corresponding author
Ethics declarations
Ethics approval
This study protocol was approved by the Commission of Ethics and Scientific Responsibility, Faculty of Sciences, UNAM (CEARC/Bioética/0307202).
Competing interests
The authors have no competing interests to declare that are relevant to the content of this article.
Additional information
Communicated by T. Breithaupt.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Burciaga, L.M., Alcaraz, G. Choosing a bigger opponent: the path to victory and greater gains for the small contestant. Behav Ecol Sociobiol 78, 36 (2024). https://doi.org/10.1007/s00265-024-03447-6
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00265-024-03447-6