
Abstract
Marine turtles encounter different threats during various life-history stages. Therefore, understanding their movements and spatial distribution is crucial for effectively managing these long-lived migratory organisms. This study combines satellite telemetry data with long-term capture-mark-recapture data derived from flipper tag studies to determine distribution patterns of endangered loggerhead turtles (Caretta caretta) during post-nesting migrations from different eastern Australian nesting sites. Individuals from the K’gari-Fraser Island and Great Barrier Reef island rookeries typically migrated northward, whereas individuals from mainland rookeries migrated equally northward and southward. Despite this difference in foraging distribution, loggerheads from the different rookeries did not differ substantially in their migration duration or distance travelled. The foraging distribution identified from successful satellite tag deployments represented 50% of the foraging distribution identified from a large long-term flipper tag recovery database. However, these satellite telemetry results have identified new migration and foraging habitats not previously recognised for loggerhead turtles nesting in eastern Australia. Additionally, they support the conclusion from a past study using flipper tag recovery data that the mainland nesting turtles migrate to different foraging grounds than the turtles nesting on Great Barrier Reef islands. Collectively, the two data sources provide valuable data on the migration route, habitat distribution and ecological range for a threatened genetic stock of loggerhead turtles.
Similar content being viewed by others


Introduction
Migration between resident foraging and breeding sites is a key component in the life history of many marine species. As such, understanding behaviour of animals during these phases is a necessary basis for quantifying their vulnerability to threats. For air-breathing marine species that navigate through oceanic waters, and often remain underwater for extended periods of time, studying their movements and behaviours during their migration and at foraging sites presents additional challenges not encountered with terrestrial species. While these challenges were surmounted through the application of tagging and capture-mark-recapture (CMR) systems (e.g. using natural markers with photo identification, fin clipping, tagging [bird bands, turtle flipper tags, PIT tags]), more recently applied techniques such as telemetry methods (e.g. acoustic, GPS satellite), remote sensing, and analysis techniques for tracking now provide valuable insight towards behaviour and ecological range of migrating marine species (Dingle 2014; Hussey et al. 2015).
The post-nesting migration of marine turtles was first studied using flipper tags in CMR studies and has been useful in determining information such as the distribution or demographic parameters of migrating adults (Bustard and Limpus, 1971; Schroeder et al. 2003). Although this method can determine migration end points and the extent of species distribution, satellite-based telemetry provides detailed information on the movements and behaviours of turtles such as the route travelled, stops made along that route, diving behaviour, and water temperature, among other types of information (Schroeder et al. 2003; Godley et al. 2008). Titanium and Inconel flipper tags have enabled the tracking of movements and habitat use with individual marine turtles across decades (Limpus and Limpus 2003) while current satellite telemetry technology is limited to tracking turtles over short durations, often less than a year, as in the current study. Over time, advances in satellite tag technology, such as the advent of Fastloc-Global Positioning System (GPS) that has allowed for more accurate locations and can receive signals in just milliseconds, have facilitated the tracking of air-breathing marine organisms that may only surface briefly (Dujon et al. 2014; Thomson et al. 2017). Collectively, the information derived from either CMR or telemetry studies has been used for analysis of fine-scale movements and behaviours of marine turtles during migration, and identification of foraging sites (Limpus and Limpus 2001; Troëng et al. 2005; Shimada et al. 2020). This information provides an important basis for the management of threats to migrating species and improving understanding of ecological range as a basis for establishing or refining boundaries for regional management units or other population designations (e.g. Wallace et al. 2010; Hays et al. 2014; 2020).
As with other sea turtle species, it was traditionally believed that post-nesting loggerheads (C. caretta) took a relatively direct path from their nesting to foraging areas, and this indeed remains the case for many individual migrations (Schroeder et al. 2003). However, satellite telemetry studies have demonstrated that this is not always the case, with variation in post-nesting migration behaviour among species and individuals within the same species. Some turtles make a stopover en route to their primary foraging ground, presumably for feeding to acquire additional energy to continue their journey or for resting (Dujon et al. 2017; Evans et al. 2019; Shimada et al. 2020), while others bypass potential foraging sites during their migration (Shimada et al. 2020). Individuals vary in their total post-nesting migration distance depending on where the individuals forage, with some making short-distance migrations and others that use the same nesting site undertaking long-distance migrations (Limpus et al. 1992; Schroeder et al. 2003). Female loggerhead turtles from eastern Australian nesting sites have been recorded foraging domestically in Northern Territory, Queensland, and New South Wales, and internationally in Indonesia, Papua New Guinea, Solomon Islands, and New Caledonia (Bustard and Limpus 1971; Limpus et al. 1992; Limpus 2008). Although there is a wide distribution of foraging areas for this population, up to 2600 km from the rookeries, most individuals undertake shorter breeding migrations of less than 1000 km (Limpus 2008).
With the expansion of Australian trawl fisheries in the 1960 s to 1980 s, a marked decline was observed from the mid-1970 s in the population of annually breeding loggerhead females at all index monitored beaches in Queensland up until the regulated mandatory use of turtle exclusion devices (TEDs) in bottom trawl fisheries (Limpus 2008). Since the 2001 regulation, the use of TEDs in the trawl fisheries of northern Australia and eastern Queensland resulted in the reduced mortality of marine turtles in these fisheries and associated increased abundance of breeding loggerhead females at the index beaches in Queensland (Limpus et al. 2013). Consequently, there has been a substantial increase in loggerhead breeding numbers at the mainland beaches of south-east Queensland, with current nesting numbers resembling those of the early 1980 s. In contrast, there has been only a trivial level of recovery in loggerhead breeding numbers at the island beaches of the southern Great Barrier Reef (GBR; Wreck Island, Heron Island, Lady Musgrave Island) during the same period (Limpus et al. 2013). A past study based on flipper tag recoveries of individuals from eastern Australian rookeries identified significant differences in the distances travelled by individuals from mainland beaches versus those breeding on the GBR islands, with the latter travelling almost twice as far (Limpus et al. 1992). The prominent difference in recovery of the nesting aggregations on the mainland compared to those on the GBR island beaches indicates that the TED-related reduction in trawl bycatch mortality of loggerhead turtles has not been uniformly effective for all the loggerhead turtle nesting populations of eastern Australia,. This difference in recovery coupled with the significant differences in distances travelled implies different degrees of interaction with trawl fisheries or other threats for the mainland versus GBR island individuals.
Knowing the extent of distribution and the degree to which foraging areas are within protected estate is important for conservation initiatives such as establishing management units (Wallace et al. 2010; Lohe and Possardt 2021). Because flipper tag-based projects may take decades to receive sufficient tag recoveries, alternative techniques such as satellite tracking, population-genetics, and stable isotopes are increasingly being used to understand the distribution of turtles from nesting areas to foraging sites (Godley et al. 2008). Satellite tracking of nesting turtles as they migrate to foraging areas is particularly common, because access to the turtles and attachment of the tags is relatively straightforward (Coyne and Godley 2005). However, because satellite tags are expensive, monitoring projects tend to satellite tag fewer individuals than they flipper tag (Godley et al. 2008). A key knowledge gap in sea turtle CMR or monitoring projects is knowing when tagging and sampling is enough to obtain a robust understanding of population trends, dispersal and distribution (Shimada et al. 2020). Therefore, we use a combination of flipper tag recovery data and satellite telemetry to examine the post-nesting migration patterns of loggerheads from five rookeries of the two regions to (1) quantify the differences between loggerhead turtle post-nesting migrations from individual rookeries and between the mainland and GBR island rookeries, (2) compare the foraging distribution identified by the use of satellite telemetry with that identified from flipper tag recoveries (CMR studies) to determine if results from satellite telemetry are representative of the known foraging range of eastern Australian loggerhead turtles, (3) quantify the proportion of suitable habitat that is utilised by the tracked or tagged loggerhead turtles that breed in eastern Australia, and (4) determine whether satellite telemetry can be used alone to determine the distribution of a nesting population.
Methods
Loggerhead migration data
This study analysed satellite telemetry data collected within the Queensland Department of Environment and Science (QDES) Turtle Conservation Project (QTCP) and examines data from 50 satellite-tracked turtles from 5 rookery areas within Queensland, Australia: Wreck and Heron Islands, Woongarra Coast including Mon Repos, K’gari-Fraser Island including Point Vernon within Hervey Bay, and the Sunshine Coast (see Fig. 1). These data span 26 years of telemetry studies, 1994–2020. All satellite-tracked loggerheads were selected from among nesting turtles at the rookeries, while excluding turtles that had been previously recorded at a foraging area (i.e., known foraging areas). All turtles were caught after they had laid a clutch of eggs, and the satellite telemetry tags were attached to carapaces using epoxy following the well-established technique used by Limpus and Limpus (2001) and subsequent researchers.

Nesting locations of southwest Pacific stock of loggerhead turtles within Australia
Flipper tag recovery data (Limpus 1985, 1992) from loggerhead turtles nesting in Queensland were extracted from the QTCP database and span 1968–2016. The recapture data spans 52 years (1968 to 2016) of study on loggerhead turtles in Queensland. In brief, at numerous beaches in Queensland, each nesting loggerhead turtle encountered in each year has been tagged with one or more flipper tags (monel tags prior to 1981; titanium tags since 1981; Limpus 1992; Fig. 1). Approaching 100% of nesting turtles have been tagged annually since 1964 at Heron Island and since 1968 at Woongarra Coast. At least 70% of the annual loggerhead turtle nesting population has been tagged at Wreck Island since 1976, and at Wreck Rock beaches since 1978. At K’gari-Fraser Island, Swain Reefs, and the Sunshine Coast the tagging effort has been intermittent from year to year (Limpus 2008).
This study focused on the Capricorn-Bunker Group, the Woongarra Coast, and K’gari-Fraser Island rookeries where both flipper-tagging and satellite telemetry data are available. The Capricorn-Bunker Group of islands is in the southern GBR and encompasses 14 coral cays where nesting loggerhead turtles have been flipper-tagged (including Wreck and Heron Islands). Annual QTCP tagging data from these islands that are near each other show that a proportion of the nesting loggerhead turtles interchange among the islands within a single breeding season and between breeding seasons. Similar interchange between adjacent beaches by nesting loggerhead turtles is common on the Woongarra coast. The long sand beaches of K’gari-Fraser Island are treated as a single rookery site. Excluded from analysis were recoveries from turtle’s flipper-tagged in the Swain Reefs (n = 4), Wreck Rock/Agnes Water (n = 52), Moreton Island (n = 2), and northern NSW (n = 1) rookeries, because we have no satellite tracks from these locations. The analysed subset of 235 flipper tag recoveries included 80% of all flipper tag recovery data and was chosen to represent the rookeries for which there was satellite telemetry data.
Turtles tracked by satellite telemetry
The satellite tags used were manufactured by Wildlife Computers (USA), Sirtrack (New Zealand), or Telonics (USA). Of the 50 tags, 10 were non-GPS platform transmitting terminal (PTT) tags and 40 were Fastloc-GPS linked PTT tags (Table S1). Fastloc-GPS tags are able to obtain more accurate locations as compared to the traditional Argos-based PTT tags (i.e., within tens of metres vs. hundreds of metres).
The raw data were obtained from CLS-Argos and decoded by the tag manufacturer’s software or online data portal. For the 10 PTT tags, we only included data points corresponding to Argos location classes 1, 2, or 3 and for the Fastloc-GPS tags we only included data points derived from 4 or more GPS satellites. All data points for each of the 50 satellite-tracked individuals were then screened using the R package SDLfilter, which removed duplicate locations, positions on land, and biologically unrealistic locations based on individual-specific calculations of travel speed (> 5 km/h) and residual error (> 30) (Shimada et al. 2012, 2016). We then plotted the data in ArcGIS and distinguished migration locations from inter-nesting and foraging locations using daily averages of traveling speeds and turning angle for each turtle (Barr et al. 2021; Shimada et al. 2021a; Patrício et al. 2022). In general, foraging was classified with daily average travel speeds < 1 km/h, inner angles between three consecutive locations of < 100° and daily movement not orientated in a uniform direction. Because these behavioural modes were clearly distinguishable, advanced analytical methods were not required to differentiate between the three modes. The inter-nesting area was identified as an aggregation of points before direct travel. The start of migration for each individual was chosen as the location at which a female began direct travel from her inter-nesting area towards her foraging ground. The last day of migration was determined as the first location after directed swimming ceased and location points began to aggregate, indicating arrival at a foraging area (Christiansen et al. 2017; Shimada et al. 2020). The foraging area was determined as the aggregation of locations after migration for which there was no further direct travel. Minimum travel distance (i.e., sum of distances between all points during migration) and duration was calculated using the determined beginning and ending migration points for each female.
Of the 50 loggerhead turtles deployed with satellite tags, eight did not complete migration, i.e., the satellite tags stopped transmitting before those individuals reached their respective foraging areas. Thus, there were 42 individuals tracked by satellite telemetry available for detailed analyses of migratory paths and foraging area locations.
Flipper tag data and dispersal of recoveries
The flipper tag dataset (n = 235) contained recaptures both from opportunistic events, such as fisheries bycatch, indigenous hunting and stranding events, and targeted captures through research efforts at index study sites such as Moreton Bay and selected coral reefs of the southern and northern GBR. Stranding of marked animals indicates that turtles would rarely strand more than 20 km from their initial capture location (unpublished data Qld Government). As such, there was uneven effort applied for identification of distribution and abundance of flipper-tagged turtles throughout their foraging range. To account for this bias of effort expended at different recapture sites, and to map the cumulative dispersal of distribution range, an overlay of 0.25° by 0.25° grid cells (approximately 25 km by 25 km) was created, and in each grid cell, only the first record of flipper tag recoveries from each rookery were used. Where there were two or more records from the same rookery within one grid cell, the individual with the earliest capture date was retained and the later recoveries and associated capture date(s) removed from the analysis. For example, a grid cell located within the Capricorn Reefs contains nine flipper tag recoveries, with two from the Capricorn-Bunker rookeries (green), five from the Woongarra Coast rookeries (pink), and two from the K’gari-Fraser Island rookery (purple) (Fig. 2a). After removing all non-unique flipper tag recoveries, the grid cell was left with the first record within the cell, point A, and the first records from other individual rookeries, the B points, which reflect the first record for the K’gari-Fraser and Capricorn-Bunker rookeries (Fig. 2b). The extent of the grid was determined by the extent of recapture points at major meridians (136° and 167°) and parallels (− 5° and − 38°). This resulted in 109 unique flipper tag recoveries (i.e., no 2 records in a grid cell were from the same rookery) being used for analysis.

Example of determination of cell selection for flipper tag recoveries and satellite-tagged migration endpoints using reefs of the southern Great Barrier Reef (GBR) with 0.25° × 0.25° grid cells overlaid. The points shown are a all foraging-site locations of flipper tag recoveries (unique and non-unique) and b only unique foraging locations of flipper tag recoveries with point A indicating the first flipper tag recovery within the cell and the B points indicating the first flipper tag recoveries of other individual rookeries. Points are coloured according to the rookery origin of each flipper-tagged individual (green = Capricorn-Bunker Group rookeries, pink = Woongarra Coast rookeries, purple = K’gari-Fraser Island rookery), and the grey outlines represent the reef areas
Comparison of satellite-derived locations and flipper tag recoveries
To compare the ranges of flipper tag recoveries and satellite tags, flipper and satellite tag rookery locations and foraging ground locations were plotted within the same 0.25° by 0.25° grid layer. Only flipper tag recoveries from the same rookeries as satellite tags were plotted (Capricorn-Bunker Groups, Woongarra Coast, and Point Vernon in Hervey Bay; there were no individuals from the Sunshine Coast rookery within the flipper tag recovery data). Similar to the method used for plotting flipper tag recoveries, only unique satellite-tracked individuals (n = 39) were plotted (i.e., not all 42—only the first record of a satellite-tagged turtle from each rookery foraging within a grid cell). Foraging ground locations for satellite-tagged individuals were chosen by determining the mean centre of all foraging point locations (i.e., centroid) (Shimada et al. 2020).
Suitable habitat and distribution
Areas of suitable habitat were identified as the entire Great Barrier Reef World Heritage Area (GBRWHA), and any area within 25 km of the coast for the Northern Territory (NT), New South Wales (NSW), Indonesia, Papua New Guinea (PNG), New Caledonia, and the Solomon Islands. The entire GBRWHA was chosen because loggerhead turtles are known to occur along the entire coast and reef areas, and forage within the lagoon areas between the coast and reef (Limpus 2008). A 25-km buffer was chosen around the rest of the land areas across the range as home ranges for loggerhead turtles can extend out to 25 km from the centroid foraging home range location (Shimada et al. 2020). Depth was not used, as for our study population, there is no detailed information on the preferred or maximum depth that loggerheads forage at, and therefore the use of depth would be too subjective. To determine the amount of usable habitat used, the full 0.25° by 0.25° grid (Fig. 3a) was clipped to only include those areas of suitable habitat (Fig. 3b). Proportions were then calculated by determining the number of cells that contained a) satellite tag locations, b) flipper tag locations, and c) both satellite and flipper tag locations, divided by the total amount of suitable habitat cells. Locations that were outside of the 25 km buffer or GBRWHA (n = 4) were assigned as one whole cell.

a Extract of the full 0.25° by 0.25° grid used to determine b extent of usable habitat for loggerhead turtles from eastern Australia rookeries. Circles represent flipper tag recovery foraging locations colour-coded by rookery origin: green = Capricorn-Bunker Group rookeries, pink = Woongarra Coast rookeries, purple = K’gari-Fraser Island rookery. Squares represent satellite tag-derived foraging locations and are similarly colour-coded by rookery origin: green = Wreck/Heron Island rookeries, pink = Woongarra Coast rookery, purple = K’gari-Fraser Island rookery, brown = Sunshine Coast rookery. Stars represent the rookery locations where turtles were originally flipper- or satellite-tagged. Dashed line denotes the boundary of the Great Barrier Reef World Heritage Area (GBRWHA)
To determine how many satellite tags would need to be deployed to represent the spatial extent of the population, based in this case on the distribution of unique flipper tag recoveries, the cumulative number of (unique and non-unique) flipper tag recoveries (n = 235) and satellite tags (n = 42) over time were plotted in comparison to the number of cumulative new (i.e., unique) grid cells visited. The number of satellite tags needed to represent 25, 50, and 75% of the spatial range of flipper tag recoveries, was then calculated.
Statistical analyses
R software version 4.0.3 (R Core Team 2020) was used for all statistical analyses. All data were checked for normality (Shapiro–Wilk test) and homogeneity of variance (Levene test, car package; Fox and Weisberg 2019) before conducting statistical tests. To compare distance travelled and duration of migration between the K’gari-Fraser Island, mainland, and GBR island (i.e., from islands of the Capricorn-Bunker Group) nesters, one-way ANOVAs were completed. Data were log transformed to meet the assumption of normality. A Pearson Chi-square test was used to analyse the difference in overall direction travelled between the Woongarra Coast and GBR island nesters. Due to small sample sizes for the other mainland nesters (Point Vernon, Hervey Bay: n = 1, Sunshine Coast: n = 3) and K’gari-Fraser Island nesters (n = 4), these individuals were not included in the Chi-square analysis.
Results
Turtles tracked by satellite telemetry
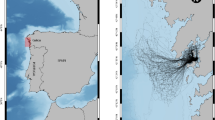
Of the 50 individuals tracked, there were eight incomplete migrations: three turtles tracked from Wreck Island (Capricorn-Bunker Group), three tracked from K’gari-Fraser Island, one tracked from Point Vernon (Hervey Bay), and one tracked from Woongarra Coast (Table S1). These eight individuals travelled for an average of 17 d (range 2–54 d) and 700 km (range 50–1600 km) before tag transmission stopped. For all individuals except the three from K’gari-Fraser Island, transmission stopped when the individual had moved past the continental shelf and was swimming in deep oceanic waters in the Coral Sea. The reason(s) for the loss of transmitted location data is not known. In each case, the transmitted diagnostic data indicates that battery life was very near full (> 80%), and the wet–dry sensors that turn transmissions on and off when turtles surface to breathe were functioning well. Of the 42 tracked individuals that completed a migration, 16 tags were deployed on turtles at Wreck and Heron Island Capricorn–Bunker Group), 18 tags were deployed on turtles at Woongarra Coast, one tag was deployed on a turtle at Point Vernon, Hervey Bay, four tags were deployed on turtles at K’gari-Fraser Island, and three tags were deployed on turtles at the Sunshine Coast (Table 1, Fig. 4).

Migration paths of 42 individuals from K’gari-Fraser Island (n = 4), mainland (n = 22; including Woongarra Coast, Point Vernon in Hervey Bay, and Sunshine Coast), and Capricorn-Bunker rookeries (n = 16). Square symbols denote rookery locations (red = K’gari-Fraser Island, black = mainland, blue = Capricorn-Bunker groups). Dashed line denotes the boundary of the GBRWHA. Migration data ranged from 1994 to 2020
Foraging areas
Females were tracked at their respective foraging areas for a median of 81.5 d (range 8–570 d) after completing migration. The foraging areas of those 42 satellite-tagged individuals were within Australia (Northern Territory, Queensland, New South Wales) and Papua New Guinea (Fig. 4). Two individuals from the K’gari-Fraser Island rookery were the only satellite-tagged individuals here who had foraging grounds in international waters in the Louisiade Archipelago of southeastern PNG. The other 40 individuals had domestic foraging areas within Queensland waters (n = 31), followed by NSW (n = 7) and NT (n = 2). Nearly half of those foraging areas were within the GBRWHA boundary (n = 18) and 12 others were in protected waters elsewhere (Crocodile Islands Marine Indigenous Protected Area, NT—currently a consultation project: n = 1, Anindilyakwa Indigenous Protected Area, NT: n = 1, Great Sandy Marine Park, Qld: n = 5, Moreton Bay Marine Park, Qld: n = 4, and Cape Byron Marine Park, NSW: n = 1; Table 1). The foraging area of most individuals (n = 30) was within coastal waters, with only 11 at a coral reef foraging area and one in estuarine waters.
The individual who ended in an estuarine foraging area, PTT 67092 from Point Vernon in Hervey Bay, originally laid her first recorded clutch for the 2018–2019 breeding season at Mon Repos. She then changed nesting beaches 64 km to the south to lay eggs at Point Vernon, Hervey Bay, on 20 December 2018 where she was fitted with her satellite tag. She then migrated north from Hervey Bay to lay a third recorded clutch for the season at Woodgate Beach on 2 January 2019, 40 km south of Mon Repos and 24 km north of Point Vernon. There was no evidence of her coming ashore for another nesting after leaving Woodgate (i.e., no location positions on or close to land afterwards). After laying this third clutch, she then migrated north along the coast to the Fitzroy River in central Queensland, ending in the Fitzroy River estuary, 50 km upstream from the mouth of the estuary (Fig. S1). This estuarine foraging area falls within the GBRWHA boundary.
Migration travel details
Migrating exclusively along the coast (i.e., remaining within 60 km of the coastline, n = 19) was the most common travel path, followed by migration paths including both coastal and offshore components (n = 17), and just six individuals migrating exclusively offshore (Table 1). The majority of individuals migrated north of their rookery (n = 25), followed by those who migrated south (n = 15), west (n = 1), or east (n = 1). The mean (± SD) distance travelled was 1198 ± 1156 km with an average duration of 30 ± 28 d and travel rate of 41 ± 10 km d−1. Most individuals had direct migrations, though 11 made stopovers (i.e., stop in one location for > 24 h before continuing migration). The longest stopover occurred in a female from the Sunshine Coast, PTT 48861, who first started migrating south, then turned around and went north into Moreton Bay (25 km south from where she began migration) where she remained for 40 d before she continued with her northward migration (Fig. S2). However, we are not certain whether she laid a clutch of eggs during this time.
GBR island and coastal rookeries comparisons
GBR Island nesters (n = 16) and K’gari-Fraser Island nesters (n = 4) on average migrated approximately twice as far as mainland nesters (n = 22), though this difference was not statistically significant (ANOVA: F2,39 = 2.651, P = 0.083) because of the high variance in all groups. The migration duration of GBR island and K’gari-Fraser Island nesters was on average slightly or twice as long as for mainland nesters, respectively, though this difference was also not statistically significant (ANOVA: F2,39 = 1.205, P = 0.311). Individuals from GBR island rookeries migrated north six times more often than south, with only one individual migrating west and none migrating east (Table 1). Individuals from the K’gari-Fraser Island rookery migrated north three times more often than south. A slim majority of individuals from the mainland rookeries migrated southward (n = 12) compared to northward (n = 9) or eastward (n = 1; note that migrating west was not possible for mainland nesters), though this difference was not statistically significant. There was a significant difference in direction of migration (northward or southward) between the Woongarra Coast and GBR island rookeries (Chi-square test: X2 = 10.279, P = 0.013), indicating that the direction of travel does depend on the rookery from which an individual originates, with GBR island nesters showing a marked tendency to migrate northward. The direction of migration for the four other mainland nesters (Point Vernon: n = 1, Sunshine Coast: n = 3) aligned closely with individuals from the Woongarra Coast rookery (n = 18), whereas the direction of migration for the K’gari-Fraser Island individuals (n = 4) aligned closely with the GBR island nesters.
Flipper and satellite tag comparisons
Of the 235 flipper tag recoveries reported between 1968 and 2016, 109 occurred in unique cells. Thirty-five recoveries in 17 unique cells were international and the last international recovery occurred in 2008. Endpoints of satellite (unique 0.25° cells: n = 39) and flipper tags (unique 0.25° cells; n = 109) showed close overlap and a few additional satellite-derived foraging areas not already represented by the locations of flipper tag recoveries (Fig. 5). These newly identified foraging grounds were widely scattered in the waters of the Arafura Sea off the Northern Territory, the Gulf of Carpentaria, Torres Strait, the GBR, and the waters around K’gari-Fraser Island. The satellite tracking did not expand the known range of the stock to the north, east, or south, but did indicate a slight westward expansion of the range in the Northern Territory.

Locations of rookeries (stars) and derived foraging locations (circles) of unique satellite-tagged individuals (n = 39, blue) and unique flipper tag recoveries (n = 109, red). Rookeries are located in both Queensland and New South Wales. Foraging locations were found in Australia (Northern Territory, Queensland and within the GBRWHA–dashed line, and New South Wales), Indonesia, Papua New Guinea, the Solomon Islands, and New Caledonia
There were numerous foraging areas identified via flipper tag recoveries that were not represented among foraging sites identified via satellite telemetry (Fig. 5, red circles). The additional foraging sites from flipper tag recoveries included: international foraging areas in Indonesia (n = 2 turtles), the Solomon Islands (n = 1 turtle) and New Caledonia (n = 3 turtles); waters off Weipa in eastern Gulf of Carpentaria; northern GBR between Cairns and Lizard Island; and southern GBR Reefs: Capricorn-Bunker Group, Swain Reefs. From the extent of habitat we define as suitable (n = 1,381 complete cells), only 2.8% was utilised by only satellite-tagged individuals, 6.7% was utilised by only flipper-tagged individuals, and 0.9% was utilised by both (overlapping) satellite telemetry and flipper-tagged individuals.
When comparing the cumulative spread of satellite-derived locations within the known distribution range based on flipper tag recoveries: to represent 25% of the range of flipper tag recoveries required ~ 45 flipper tag recoveries and ~ 21 satellite tag deployments; to represent 50% of the known range required ~ 80 flipper tag recoveries and ~ 41 satellite tag deployments; to reach 75% of the known range required ~ 135 flipper tag recoveries (Fig. 6). Many more than the 42 satellite tag deployments analysed here are needed to determine the rate of increase and estimate how many tags would be required to reach 75% of the known range based on the flipper tag recoveries.

Comparison of the number of cumulative new cells visited by cumulative number of individuals recorded over time. Flipper tag recoveries (n = 235, red) span from 1968 to 2016. Satellite-tags (n = 42, blue) collected location information from 1994 to 2020. Dashed lines indicate the number of satellite tags needed to represent 25, 50, and 75% of the spatial range of flipper tag recoveries
Discussion
In this study, we combined a long-term flipper tag recovery dataset with satellite tracking of post-breeding adult females to examine the post-nesting migration patterns and foraging area destinations of loggerheads from five rookeries. With the extensive flipper tagging and satellite tracking data set from nesting and foraging loggerhead turtles in eastern Australia, there are now hundreds of data points that link the nesting beaches to foraging areas within approximately 2,600 km of the beaches (Limpus et al. 2013). In addition, many of these records demonstrate high rates of fidelity of the individual turtles to both their home foraging area and distant nesting beach (Limpus and Limpus 2003; Shimada et al. 2020). We found that both flipper tag (our study and Limpus 1992) and satellite tag data show a similar dispersal pattern and indicate that loggerhead turtles nesting on the mainland coast of eastern Australia migrate to different foraging areas than those nesting on the islands of the southern GBR. However, a wider dispersal range from the nesting beaches was detected by flipper tag recoveries than satellite tracks because none of the satellite-tracked turtles migrated across the Coral Sea to New Caledonia or north-eastern Papua New Guinea—where flipper-tagged turtles have been recorded.
Advances in technology have led to an increase in the number of global projects using satellite telemetry to understand migration (Godley et al. 2008; Hays et al. 2019). Given this increased use of telemetry, there is a growing need to understand sample sizes required to develop robust findings (Sequeira et al. 2019; Shimada et al. 2021b). Hence, we combined the results of our satellite telemetry with data from long-term flipper tag recoveries (1970 s to the present) to assess the dispersal and number of satellite-tracked turtles that would be needed to approximate the known foraging range and dispersal of the Southwest Pacific Loggerhead stock, as already determined by flipper tag recoveries. Overall, the endpoints of satellite tracked and flipper tag recovered individuals indicate that from this stock of loggerhead turtles, 21 and 41 satellite tags would be required to represent 25 and 50% of the foraging areas previously identified by flipper tag recoveries. However, both techniques only provide a low cover of all possible (1,381 unique 0.25° cells) foraging habitats for the species in the region. Using satellite telemetry to identify ecological range is also challenging unless large sample sizes can be obtained. In our study, while satellite telemetry expanded the foraging range to the west, it underestimated the range to the north (north-eastern PNG and Solomon Islands), east (New Calendonia) and south (central New South Wales). At present, it has not been resolved whether the recently reduced frequency of recording loggerhead foraging at overseas sites is a consequence of our sample size of tracked individuals or an indication of reduced foraging populations remaining within these areas (UNEP and CMS 2014; Limpus and Casale 2015). This has implications for regional threatened species initiatives such as establishing Regional Management Units (Wallace et al. 2010) or Distinct Population segments (Lohe and Possardt 2021).
Of the 42 satellite-tracked individuals, most (n = 30) migrated to foraging areas within protected waters, with the remaining (n = 12) migrating to unprotected areas in PNG, Gulf of Carpentaria, Torres Strait, and northern NSW. Given the absence of Marine Parks in Torres Strait and, until recently, in the Gulf of Carpentaria, with more of the satellite-tracked turtles from GBR island rookeries and three of the four tracked from K’gari-Fraser Island migrating to the north, it is possible that their foraging areas have not benefited from the more rigorous habitat protection that occurs within Marine Parks (which encompass 97% of the east coast of Queensland between Cape York and the New South Wales Border; Queensland Government 2018). Additionally, being within a Marine Protected Area or Marine Park during breeding seasons does not necessarily exempt loggerheads from encountering harm during migration or at a foraging site, as the degree of protection can differ depending on the specific zoning an individual travels through or resides within.
Satellite telemetry data provide insights into turtle behaviour and habitat use that cannot be learned from reported flipper tag returns. When comparing differences between satellite-tagged turtles from the mainland, Fraser Island, and the GBR islands, individuals from the three types of rookeries did not exhibit statistically significant differences in their migration travel distance or duration, travelling across an average of 1200 km and over 30 d. Although not statistically significant, the magnitude of the differences in distance travelled is consistent with the findings of Limpus et al. (1992), which indicated that the distance travelled by flipper-tagged GBR island nesters is twice that of mainland nesters. In that study, the difference was significant, though there was a greater sample size (mainland nesters: n = 81 mainland nesters and GBR island nesters: n = 37) than the present study (mainland nesters: n = 22, GBR island nesters: n = 16, K’gari-Fraser Island nesters: n = 4). The smaller sample size here likely precludes any robust statistical analyses. Nevertheless, tracked loggerheads significantly differed in their direction of travel, with individuals from the GBR island rookeries predominantly traveling northwards and those from the mainland rookeries more evenly distributed to the north and south, and occasionally to the east of their respective nesting locations. Although Fraser Island individuals were not included in the statistical directional analysis, their direction of travel aligned more closely with turtles from the GBR island rookeries, with three of the four turtles undertaking a northward migration. Migration routes from the nesting areas most commonly occurred close to the coast and migration routes that tracked entirely through the outer reef areas of the GBR away from the coast or off the continental shelf were the least common (n = 6). Sixteen of the 19 individuals that migrated exclusively along the coast were mainland nesters.
As found in similar studies with other stocks of loggerhead turtles, the majority (n = 31) of loggerhead turtles in this Queensland-based study did not make a stopover during migration (Dujon et al. 2017; Evans et al. 2019). Migration routes were typically direct, though en route stops did occur and may be attributed to resting (Evans et al. 2019; Mettler et al. 2019) or opportunistic foraging to re-establish nutrient and energy levels before reaching the resident foraging area (Dujon et al. 2017; Shimada et al. 2020). However, the reason(s) for an en route stop(s) for these migrating loggerheads warrants further investigation, which could be enabled by using combinations of satellite tracking tags coupled with multi-sensor data loggers or critter-cams to record swimming depth, foraging activity, etc. Most females tracked in this study that made an en route stop (n = 11) and spent 1–9 d at their stopover locations. The exception, one individual from the Sunshine Coast rookery (PTT 48861), made a stopover of 40 d to the south of her nesting site before migrating northward. The possibility that this female made one or more additional nesting attempts after she laid the last recorded clutch is unlikely, because there were no location positions on, or close to (within the error radius of locations), land during the “re-nesting” intervals when a clutch could be laid (between 9 and 23 d after she laid a clutch of eggs; Limpus 1985). Although the stopover was in the opposite direction of this female’s foraging area, it is not uncommon for migrating marine turtles to travel off-course (i.e., not directly towards their previously established neritic foraging area) before reorienting and continuing to their intended site (Hays et al. 2020). Additionally, females have occasionally been observed changing their nest site either between nesting seasons or within a single season, including a different individual from this rookery in 2015 (Hofmeister et al. 2019). It is, therefore, possible that this female, PTT 48861, was familiar with the stopover site in Moreton Bay from a previous nesting season as a potential transitory post-nesting foraging area.
This satellite telemetry study of breeding female loggerhead turtles in eastern Australia has provided new insights regarding their behaviour and movements. One female, PTT 196472, migrated to a foraging area beyond the foraging range identified via the extensive flipper tag recovery data set: after nesting at Heron Island, she travelled 2,550 km to occupy a foraging area 200 km beyond the Gulf of Carpentaria off the north coast of Arnhem Land in the Arafura Sea. While loggerhead turtles have been reported sporadically to occur in estuarine habitats in south Queensland (Burrum and Burnett Rivers, QTCP database), one tracked individual, PTT 67092, displayed persistent foraging behaviour in estuarine habitats. After nesting in western Hervey Bay, she travelled 350 km north to forage along ~ 50 km of estuarine habitat from the mouth of the Fitzroy River upstream to the city of Rockhampton across an entire year in 2019. Though this behaviour is less common for loggerhead turtles, estuarine foraging areas have been recorded for eastern Pacific hawksbill turtles (Gaos et al. 2012), loggerheads in Greece and the western Atlantic (Rees et al. 2013; McNeill et al. 2020), and speculated based on digestive tract contents for loggerheads in Uruguay (Carranza et al. 2011). However, use of the river upstream of the estuaries in those different regions has not been confirmed.
With the use of flipper tag recoveries alone, there may be a potential bias in the endpoints (foraging area) identified by mark-recapture research efforts (i.e., more effort placed into those areas for which individuals were found, or likely to be found). Although we attempted to account for this by only including unique flipper tag recoveries, this type of bias is nevertheless present. The use of satellite tag deployments would be free of this bias but comes at a cost. Furthermore, because we used a previously collected dataset on flipper tag recoveries, the temporal match of our data on flipper tag recoveries and our satellite tag deployments are not equal. With nearly half of our satellite deployments from 2018 to 2020, it is not possible to determine whether the year of tagging influences the distribution of foraging sites. A more balanced approach examining long-term patterns in foraging area distribution using a variety of techniques for different nesting cohorts of turtles would certainly be a useful future research direction, which could have important management implications.
Analysis of flipper-tagging data to understand foraging area distribution requires tagging a large number of turtles across years (i.e., in our study 10,000 s), as there is often a low recovery rate of tagged turtles (Limpus 1992; Witzell 1998; Balazs 1999). The budget for tagging projects therefore needs to include the price of tags (e.g. $2,200 USD per 1,000 titanium tags) as well as all related costs, such as salaries, travel expenses and living expenses at site, and data analysis and write-up afterwards. Plus, it should consider the costs associated with complementary projects aimed at recapturing or recovering tagged individuals. In a long-term project, these costs can be significant (Balazs 1999). Therefore, in some regions of the world, research and monitoring agencies are adding value to flipper-tagging projects through the co-inclusion of additional projects aimed at understanding foraging area distribution through stable isotope analysis, molecular ecology, and satellite telemetry (e.g. Jensen et al. 2016; 2020; Pearson et al. 2019; Haywood et al. 2019; Coffee et al. 2020). Given there are high costs associated with long-term flipper-tagging projects and deployment of large numbers of GPS satellite tags (Godley et al. 2008; Thums et al. 2018; Sequeira et al. 2019) understanding costs, benefits and bias associated with each method is important for defining the broad-scale foraging distribution of this loggerhead turtle stock.
Our study highlights the value of combining data from both flipper tag recoveries and satellite tag deployments to assess the distribution and behaviours of migrating loggerhead turtles from eastern Australian rookeries. The information gained from both datasets will be useful in determining ecological range for the species or genetic stock and can be used to concentrate collaborative management efforts, both internationally and within Australia (Miller et al. 2020), and aid in species-based conservation initiatives (UNEP and CMS 2014).
Data availability
Tracking data are available via the QTCP database on request.
Code availability
Not applicable.
References
Balazs GH (1999) Factors to consider in the tagging of sea turtles. In: Eckert KL, Bjorndal KA, Abreu-Grobois FA, Donnelly M (eds) Research and management techniques for the conservation of sea turtles, vol 4. IUCN/SSC Marine Turtle Specialist Group Publication, Blanchard, pp 101–109
Barr CE, Hamann M, Shimada T, Bell I, Limpus CJ, Ferguson J (2021) Post-nesting movements and feeding ground distribution by the hawksbill turtle (Eretmochelys imbricata) from rookeries in the torres strait. Wildl Res 48(7):598–608
Bustard HR, Limpus CJ (1971) Loggerhead turtle movements. Br. J Herpetol 4:228–230
Carranza A, Estrades A, Scarabino F, Segura A (2011) Loggerhead turtles Caretta caretta (Linnaeus) preying on the invading gastropod Rapana venosa (Valenciennes) in the Río de la Plata Estuary. Mar Ecol 32:142–147. https://doi.org/10.1111/j.1439-0485.2010.00424.x
Christiansen F, Esteban N, Mortimer JA, Dujon AM, Hays GC (2017) Diel and seasonal patterns in activity and home range size of green turtles on their foraging grounds revealed by extended Fastloc-GPS tracking. Mar Biol 164:10. https://doi.org/10.1007/s00227-016-3048-y
Coffee OI, Booth DT, Thia JA, Limpus CJ (2020) When isotopes fail: importance of satellite telemetry and multi-site validation when estimating the foraging grounds of migratory species. Mar Ecol Prog Ser 633:197–206
Coyne MS, Godley BJ (2005) Satellite tracking and analysis tool (STAT): an integrated system for archiving, analyzing and mapping animal tracking data. Mar Ecol Prog Ser 301:1–7. https://doi.org/10.3354/meps301001
Dingle H (2014) Migration: the biology of life on the move. Oxford Univ Press. https://doi.org/10.1093/acprof:oso/9780199640386.001.0001
Dujon AM, Lindstrom RT, Hays GC, Backwell P (2014) The accuracy of Fastloc-GPS locations and implications for animal tracking. Methods Ecol Evol 5:1162–1169. https://doi.org/10.1111/2041-210x.12286
Dujon AM, Schofield G, Lester RE, Esteban N, Hays GC (2017) Fastloc-GPS reveals daytime departure and arrival during long-distance migration and the use of different resting strategies in sea turtles. Mar Biol 164:187. https://doi.org/10.1007/s00227-017-3216-8
Evans DR, Carthy RR, Ceriani SA (2019) Migration routes, foraging behavior, and site fidelity of loggerhead sea turtles (Caretta caretta) satellite tracked from a globally important rookery. Mar Biol 166:134. https://doi.org/10.1007/s00227-019-3583-4
Fox J, Weisberg S (2019) An R companion to applied regression, Third edition. Sage, Thousand Oaks CA. https://socialsciences.mcmaster.ca/jfox/Books/Companion/. Accessed Sept 2020
Gaos AR, Lewison RL, Yanez IL, Wallace BP, Liles MJ, Nichols WJ, Baquero A, Hasbún CR, Vasquez M, Urteaga J, Seminoff JA (2012) Shifting the life-history paradigm: discovery of novel habitat use by hawksbill turtles. Biol Lett 8:54–56. https://doi.org/10.1098/rsbl.2011.0603
Godley B, Blumenthal JM, Broderick AC, Coyne MS, Godfrey MH, Hawkes LA, Witt MJ (2008) Satellite tracking of sea turtles: where have we been and where do we go next? Endanger Species Res 4:3–22. https://doi.org/10.3354/esr00060
Hays GC, Mortimer JA, Ierodiaconou D, Esteban N (2014) Use of long-distance migration patterns of an endangered species to inform conservation planning for the world’s largest marine protected area. Conserv Biol 28:1636–1644. https://doi.org/10.1111/cobi.12325
Hays GC, Bailey H, Bograd SJ, Bowen WD, Campagna C, Carmichael RH, Sequeira AM (2019) Translating marine animal tracking data into conservation policy and management. Trends Ecol Evol 34:459–473. https://doi.org/10.1016/j.tree.2019.01.009
Hays GC, Cerritelli G, Esteban N, Rattray A, Luschi P (2020) Open ocean reorientation and challenges of island finding by sea turtles during long-distance migration. Curr Biol 30:3236-3242.e3. https://doi.org/10.1016/j.cub.2020.05.086
Haywood JC, Fuller WJ, Godley BJ, Margaritoulis D, Shutler JD, Snape RT, Widdicombe S, Zdinden JA, Broderick AC (2019) Spatial ecology of loggerhead turtles: Insights from stable isotope markers and satellite telemetry. Divers Distrib 26(3):368–381
Hofmeister K, Twaddle H, O’Connor J, Limpus CJ (2019) Bribie island turtle trackers, turtlecare sunshine coast volunteers and coolum and North Shore coast care volunteers marine turtle nesting populations: sunshine coast region 2005–2016. Caloundra: Sunshine Coast Council 44pp.
Hussey NE, Kessel ST, Aarestrup K, Cooke SJ, Cowley PD, Fisk AT, Harcourt RG, Holland KN, Iverson SJ, Kocik JF, Mills Flemming JE, Whoriskey FG (2015) Aquatic animal telemetry: a panoramic window into the underwater world. Science 348:1255642. https://doi.org/10.1126/science.1255642
Jensen MP, Bell I, Limpus CJ, Hamann M, Ambar S, Whap T, David C, FitzSimmons NN (2016) Spatial and temporal genetic variation among size classes of green turtles (Chelonia mydas) provides information on oceanic dispersal and population dynamics. Mar Ecol Prog Ser 543:241–256
Jensen MP, Dalleau M, Gaspar P, Lalire M, Jean C, Ciccione S, Mortimer J, Quillard M, Taquet C, Wamukota A, Leroux G, Bourjea J (2020) Seascape genetics and the spatial ecology of juvenile green turtles. Genes 11(3):278
Limpus CJ (1985) A study of the loggerhead turtle, Caretta caretta in eastern Australia. Department of zoology. The University of Queensland, PhD Thesis, Queensland, Australia
Limpus CJ (1992) Estimation of tag loss in marine turtle research. Wildl Res 19:457–469. https://doi.org/10.1071/WR9920457
Limpus CJ (2008) Loggerhead turtle, Caretta caretta. A biological review of Australian marine turtle species. Queensland Environmental Protection Agency, Brisbane, Queensland
Limpus C, Casale P (2015) Caretta caretta (South Pacific subpopulation). The IUCN Red List of Threatened Species. https://doi.org/10.2305/IUCN.UK.2015-4.RLTS.T84156809A84156890.en
Limpus CJ, Limpus DJ (2001) The loggerhead turtle, Caretta caretta in queensland: breeding migrations and fidelity to a warm temperate feeding area. Chelonian Conserv Biol 4:142–153
Limpus CJ, Limpus DJ (2003) Biology of the loggerhead turtle in western South Pacific ocean foraging areas. In: Bolten AB, Witherington BE (eds) Loggerhead Sea Turtles. Smithsonian Institution, Washington, D.C., pp 93–113
Limpus C, Miller J, Paramenter C, Reimer D, McLachlan N, Webb R (1992) Migration of green (Chelonia mydas) and loggerhead (Caretta caretta) turtles to and from eastern Australian rookeries. Wildl Res 19:347. https://doi.org/10.1071/WR9920347
Limpus CJ, Parmenter CJ, Chaloupka, M (2013) Monitoring of coastal sea turtles: gap analysis 1. Loggerhead turtles, Caretta caretta, in the port curtis and port alma region. Report produced for the ecosystem research and monitoring program advisory panel as part of gladstone ports corporation’s ecosystem research and monitoring program.
Lohe A, Possardt E (2021) Loggerhead sea turtle (Caretta caretta) North Indian Ocean DPS, Southwest Indian Ocean DPS, Southeast Indo-Pacific Ocean DPS, South Pacific Ocean DPS, South Atlantic Ocean DPS, Northeast Atlantic Ocean DPS, and Mediterranean Sea DPS 5-Year Review: Summary and Evaluation.
McNeill JB, Avens L, Hall AG, Fujisaki I, Iverson AR (2020) Foraging and overwintering behavior of loggerhead sea turtles Caretta caretta in the western North Atlantic. Mar Ecol Prog Ser 641:209–225. https://doi.org/10.3354/meps13296
Mettler E, Clyde-Brockway CE, Honarvar S, Paladino FV (2019) Migratory corridor linking Atlantic green turtle, Chelonia mydas, nesting site on Bioko Island, Equatorial Guinea to Ghanaian foraging grounds. PLoS ONE 14:e0213231. https://doi.org/10.1371/journal.pone.0213231
Miller RL, Marsh H, Benham C, Hamann M (2020) Stakeholder engagement in the governance of marine migratory species: barriers and building blocks. Endanger Species Res 43:1–19. https://doi.org/10.3354/esr01049
Patrício AR, Beal M, Barbosa C, Diouck D, Godley BJ, Madeira FM, Regalla A, Traoré MS, Senhoury C, Sidina E, Catry P (2022) Green turtles highlight connectivity across a regional marine protected area network in West Africa. Front Mar Sci 9:812144. https://doi.org/10.3389/fmars.2022.812144
Pearson RM, van de Merwe JP, Gagan MK, Limpus CJ, Connolly RM (2019) Distinguishing between sea turtle foraging areas using stable isotopes from commensal barnacle shells. Sci Rep 9(1):1–11
Queensland Government (2018) Marine turtle conservation strategy. Conservation and Biodiversity Operations Branch, Department of Environment and Science, Queensland Government, Queensland, Brisbane
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rees AF, Margaritoulis D, Newman R, Riggall TE, Tsaros P, Zbinden JA, Godley BJ (2013) Ecology of loggerhead marine turtles Caretta caretta in a neritic foraging habitat: movements, sex ratios and growth rates. Mar Biol 160:519–529. https://doi.org/10.1007/s00227-012-2107-2
Schroeder BA, Foley AM, Bagley DA (2003) Nesting patterns, reproductive migrations, and adult foraging areas of loggerhead turtles. In: Bolten AB, Witherington BE (eds) Loggerhead Sea Turtles. Smithsonian Institution, Washington, D.C., pp 93–113
Sequeira AMM, Heupel MR, Lea M-A, Eguíluz VM, Duarte CM, Meekan MG, Thums M, Calich HJ, Carmichael RH, Costa DP, Ferreira LC, Fernandéz-Gracia J, Harcourt R, Harrison A-L, Jonsen I, McMahon CR, Sims DW, Wilson RP, Hays GC (2019) The importance of sample size in marine megafauna tagging studies. Ecol Appl 29(6):e01947. https://doi.org/10.1002/eap.1947
Shimada T, Jones R, Limpus C, Hamann M (2012) Improving data retention and home range estimates by data-driven screening. Mar Ecol Prog Ser 457:171–180. https://doi.org/10.3354/meps09747
Shimada T, Jones R, Limpus C, Hamann M (2016) Time-restricted orientation of green turtles. J Exp Mar Biol Ecol 484:31–38
Shimada T, Limpus CJ, Hamann M, Bell I, Esteban N, Groom R, Hays GC (2020) Fidelity to foraging sites after long migrations. J Anim Ecol 89:1008–1016. https://doi.org/10.1111/1365-2656.13157
Shimada T, Duarte CM, Al-Suwailem AM, Tanabe LK, Meekan MG (2021a) Satellite tracking reveals nesting patterns, site fidelity, and potential impacts of warming on major green turtle rookeries in the Red Sea. Front Mar Sci 8:633814. https://doi.org/10.3389/fmars.2021.633814
Shimada T, Thums M, Hamann M, Limpus CJ, Hays GC, FitzSimmons NN, Meekan MG (2021b) Optimising sample sizes for animal distribution analysis using tracking data. Methods Ecol Evol 12:288–297. https://doi.org/10.1111/2041-210X.13506
Thomson JA, Börger L, Christianen MJA, Esteban N, Laloë JO, Hays GC (2017) Implications of location accuracy and data volume for home range estimation and fine-scale movement analysis: comparing Argos and Fastloc-GPS tracking data. Mar Biol 164:204. https://doi.org/10.1007/s00227-017-3225-7
Thums M, Fernández-Gracia J, Sequeira AMM, Eguíluz VM, Duarte CM, Meekan MG (2018) How big data fast tracked human mobility research and the lessons for animal movement ecology. Front Mar Sci 5:21. https://doi.org/10.3389/fmars.2018.00021
Troëng S, Evans DR, Harrison E, Lagueux CJ (2005) Migration of green turtles Chelonia mydas from Tortuguero, Costa Rica. Mar Biol 148:435–447. https://doi.org/10.1007/s00227-005-0076-4
United Nations Environment Programme (UNEP), Convention on Migratory Species (CMS) (2014) Single species action plan for the loggerhead turtle (Caretta caretta) in the South Pacific Ocean
Wallace BP, DiMatteo AD, Hurley BJ, Finkbeiner EM, Bolten AB, Chaloupka MY et al (2010) Regional management units for marine turtles: a novel framework for prioritizing conservation and research across multiple scales. PLoS ONE 5(12):e15465
Witzell WN (1998) Guest editorial: messages in bottles. Mar Turt Newsl 80:3–5
Acknowledgements
Staff, volunteers and collaborating partners of the Queensland Department of Environment and Science Aquatic Threatened Species Program collected, managed and provided the data used in this study. We thank the volunteers and staff of Queensland Turtle Conservation Project for tagging and collecting the data for the Wreck Island, Heron Island and Woongarra Coast loggerheads. The Great Barrier Reef Marine Park Authority provided the funding support for the Wreck and Heron Island tracking. The telemetry studies on the Woongarra coast have been funded in part by the Guinness Water of Life Project, Bundaberg Brewed Drinks, Rush Clothing manufacturers, Mon Repos Conservation Park Tourist Visitor Donations, and JCU PhD studies. The telemetry studies on the Sunshine Coast were funded by Sunshine Coast Council. We thank the anonymous reviewers whose inputs have greatly improved the manuscript. This assistance is gratefully acknowledged.
Funding
Open Access funding enabled and organized by CAUL and its Member Institutions. This study was funded by The Queensland Department of Environment and Science, the Great Barrier Reef Marine Park Authority, Guinness Water of Life Project, Bundaberg Brewed Drinks, Rush Clothing manufacturers, Mon Repos Conservation Park Tourist Visitor Donations, and James Cook University.
Author information
Authors and Affiliations
Contributions
MP, MH, TS and CJL: conceived and designed the study. Data collection was led by CJL, EW, AS and KH. MP: analysed the data and wrote the manuscript with contribution from MH, CJL, EW, TS, and KH.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this study.
Ethical approval
The project was completed under Animal Welfare permits held by the Queensland Department of Environment and Science. The project was completed under research permits held by the Queensland Department of Environment and Science.
Additional information
Responsible Editor: P. Casale.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Perez, M.A., Limpus, C.J., Hofmeister, K. et al. Satellite tagging and flipper tag recoveries reveal migration patterns and foraging distribution of loggerhead sea turtles (Caretta caretta) from eastern Australia. Mar Biol 169, 80 (2022). https://doi.org/10.1007/s00227-022-04061-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-022-04061-8