
Abstract
Rationale
Diverse preclinical studies suggest the potential therapeutic utility of the modulation of the glutamatergic system in brain via metabotropic glutamate (mGlu) receptors. Lu AF21934, a positive allosteric modulator of the mGlu4 receptor, was previously shown to reverse behavioral phenotypes in animal models thought to mimic positive, negative, and cognitive symptoms of schizophrenia.
Objectives
To begin elucidating the brain circuitry involved in mGlu4 receptor pharmacology and add mechanistic support to Lu AF21934-induced phenotypic responses, the potential involvement of 5-HT1A receptors in these antipsychotic-like effects was explored. The tests used were the following: MK-801-induced hyperactivity and 2,5-dimethoxy-4-iodoamphetamine (DOI)-induced head twitches in mice, for positive symptoms; MK-801-induced disruptions of social interactions for negative symptoms; and novel object recognition and spatial delayed alteration test for cognitive symptoms. The microdialysis studies in which the effect of Lu AF21934 on MK-801-induced dopamine and serotonin release was investigated.
Results
The effects caused by Lu AF2193 were inhibited by administration of the selective 5-HT1A receptor antagonist WAY100635 (0.1 mg/kg). That inhibition was observed across all models used. Moreover, the concomitant administration of sub-effective doses of Lu AF21934 and a sub-effective dose of the selective 5-HT1A receptor agonist tool compound (R)-(+)-8-hydroxy-DPAT hydrobromide (0.01 mg/kg) induced a clear antipsychotic-like effect in all the procedures used. Lu AF21934 (5 mg/kg) also inhibited MK-801-induced increase in dopamine and 5-HT release.
Conclusions
The actions of Lu AF21934 are 5-HT1A receptor-dependent. Activation of the mGlu4 receptor may be a promising mechanism for the development of novel antipsychotic drugs, efficacious toward positive, negative, and cognitive symptoms.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The glutamatergic system represents the basic excitatory neurotransmission mechanism in the brain. Clinically, the administration of N-methyl-D-aspartate (NMDA) antagonists (e.g., MK-801, PCP) leads to the arousal of the full spectrum of schizophrenic symptoms, including positive, negative, and cognitive disturbances (Krystal et al. 2002; Moghaddam and Jackson 2003). In contrast, drugs known as psychotomimetics (dopaminergic and serotonergic agonists) evoke only positive symptoms of schizophrenia. These observations support the involvement of the glutamatergic system in the pathophysiology of psychotic disorders (Bickel and Javitt 2009; Javitt and Zukin 1991; Javitt et al. 2004).
According to the glutamatergic hypothesis, blockade of NMDA receptors expressed on GABAergic neurons leads to the inhibition of GABAergic control over pyramidal neurons in the thalamocortical pathway. In turn, this would lead to abnormal glutamate release and to the overactivation of cortical and subcortical structures (Krystal et al. 2002; Moghaddam and Jackson 2003; Conn et al. 2009). Therefore, the reharmonization of such enhanced release may lead to the normalization of glutamate efflux and to the reversal of the symptoms of schizophrenia (Conn et al. 2009).
GABAergic and glutamatergic metabotropic receptors have been proposed as relevant targets involved in the regulation of glutamate release (Conn and Pin 1997; Pałucha-Poniewiera et al. 2008; Pin and Duvoisin 1995; Schoepp and Marek 2002; Wierońska et al. 2011). Among these, the presynaptic metabotropic glutamate mGlu2/3 and mGlu4 receptors appear optimal targets for potential antipsychotic treatment. All of these are negatively linked to adenylyl cyclase activity, and their activation inhibits glutamate release (Pin and Duvoisin 1995). Moreover, agonist and positive allosteric modulator (PAM) tool compounds such as mGlu2/3 activators (LY354740, LY379268, CIBeS, BINA) (Cartmell et al. 2000; Galici et al. 2005; Fell et al. 2008), and mGlu4 activators (ACPT-I, LSP1-2111, Lu AF21934, and Lu AF32615) were shown to be active in animal models of schizophrenia-like behaviors (Pałucha-Poniewiera et al. 2008; Sławińska et al. 2013). A few mGlu2/3 agonists (most recently pomaglumetad methionil or LY2140023), and an mGlu2 PAM (ADX71149 or JNJ-40411813) have also reached clinical testing, with mixed results (Patil et al. 2007; Kinon et al. 2011; Hopkins 2013).
Herein, we report follow-up investigations on the mechanism of action of Lu AF21934, focusing on the functional interaction between the mGlu4 and 5-hydroxytryptamine (HT)1A receptors.
The 5-HT1A receptors seem to be of special importance in schizophrenia, as the increased 5-HT1A receptor level was observed in the postmortem study of the cerebral cortex and hippocampus of schizophrenic patients (Burnet et al. 1996; Simpson et al. 1996). Moreover, the majority of novel atypical neuroleptics with good antipsychotic efficacy (ziprasidone and lurasidone) are partial agonists of 5-HT1A receptors (Schmidt et al. 2001; Sprouse et al. 1999; Ichikawa et al. 2002; Hagiwara et al. 2008). And finally, the 5-HT1A agonist, 8-OH-DPAT, at the low doses, induced antipsychotic-like effects in animal models (Bubeníková-Valesová et al. 2007; Boulay et al. 2004).
These studies were motivated by earlier work showing that the actions of the mGlu4 receptor agonist LSP1-2111 is 5-HT1A receptor-dependent. This interdependence was shown in a number of preclinical models thought to represent positive, negative, and cognitive symptoms of schizophrenia. We used the antagonist WAY100635 and the agonist (R)-(+)-8-hydroxy-DPAT as 5-HT1A receptor-selective tool compounds, concomitantly administered with Lu AF21934. In addition, Lu AF21934 was also co-administered with lurasidone, a recently approved atypical antipsychotic with nonselective CNS effects, including potent 5-HT1A receptor agonism.
Materials and methods
Animals and housing
Male Albino Swiss (20–25 g) mice were used to assess MK-801-induced hyperlocomotion, and 2,5-dimethoxy-4-iodoamphetamine (DOI)-induced head twitches. Male Wistar rats weighing 250–300 g were used in the social interaction, novel object recognition, and spatial delayed alternation tests. The choice of animal species was based on our preliminary experiments, as well as on literature data, and was dictated by the optimization of the method to obtain credible results and minimization of the costs of experiments. The animals were kept under a 12:12 light-dark cycle at a room temperature of 19–21 °C, with free access to food and water. Each experimental group consisted of eight to ten animals, and the animals were used only once in each test. All the compounds were used in a volume of 10 ml/kg when given to mice and 1 ml/kg when injected to rats. All behavioral measurements were made by an observer blinded to the treatment. All procedures were conducted according to the guidelines of the National Institutes of Health Animal Care and Use Committee and were approved by the Ethics Committee of the Institute of Pharmacology, Polish Academy of Sciences in Krakow and Lundbeck Research USA.
Drugs
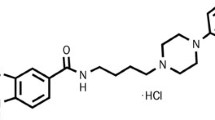
The following drugs were used: Lu AF21934 (mGlu4 receptor PAM) synthesized through Lundbeck Research USA, and characterized using H-1 and C-13 nuclear magnetic resonance spectroscopy, high performance liquid chromatography (HPLC)/mass-spectrometry methods, and X-ray crystallography. The compound was dosed as a suspension in 20 % (2-hydropropyl)-β-cyclodextrin and was administered subcutaneously (s.c.) 60 min before the tests. The administration schedule of Lu AF21934 was planned according to our previous studies on anxiety and psychosis (Sławińska et al. 2013). MK-801 (0.35, Sigma-Aldrich, St. Louis, USA) was dissolved in 0.9 % NaCl, and the doses were selected consistently with our previous work (Pałucha-Poniewiera et al. 2008; Wierońska et al. 2012, 2013a, b) and that of others (Geyer and Ellenbroek 2003; Leite et al. 2008; Satow et al. 2009). WAY100635 and (R)-(+)-8-hydroxy-DPAT (Tocris Bioscience, Bristol, United Kingdom) were dissolved in 0.9 % saline and were administered according to our previous studies and also those made by others (Wierońska et al. 2013a, b; Wedzony et al. 2000). Lurasidone hydrochloride (Ontario Chemicals, 7A-291 Woodlawn Road, Guelph, ON, N1H7L6, Canada) was dissolved in 0.5 % methylcellulose (Horiguchi et al. 2012).
Locomotor activity of habituated mice
The locomotor activity was recorded individually for each animal in OPTO-M3 locomotor activity cages (Columbus Instrument) linked online to a compatible PC. Each cage (13 cm × 23 cm × 15 cm) was surrounded with an array of photocell beams. Interruptions of these photobeams resulted in horizontal activity defined as ambulation scores. Mice were placed separately into activity cages for an acclimatization period of 30 min, and then, they were injected s.c. with Lu AF21934, WAY100635, (R)-(+)-8-hydroxy-DPAT and their combinations (time and doses of administration similar as described below for MK-801-induced hyperactivity). From this point on, the ambulation scores were measured for 60 min.
MK-801-induced hyperactivity
The locomotor activity was recorded for each animal in locomotor activity cages (according to Rorick-Kehn et al. 2007a, b), with small modifications used in our previous studies (Pałucha-Poniewiera et al. 2008; Wierońska et al. 2012, 2013a, b). The mice were placed individually into actometers for an acclimatization period of 30 min; then they were administered Lu AF21934 (60 min before MK-801, s.c), WAY100635 (45 min before MK-801, intraperitoneally (i.p)) and (R)-(+)-8-hydroxy-DPAT (15 min before MK-801, s.c), or vehicle and placed again in the same cages. After the proper time, all of the mice were administered intraperitoneally with MK-801 at a dose of 0.35 mg/kg and once again returned to the same cage. From then on, the ambulation scores were counted for 60 min. All groups were compared with the MK-801 control group. The experiment also included a control group not treated with MK-801.
Head twitch test
The experiment was performed according to Pałucha-Poniewiera et al. (2008) and Wieronska et al. (2012, 2013a, b). In order to habituate mice to the experimental environment, each animal was transferred to a 12 (diameter) × 20 cm (height) glass cage, lined with sawdust, 30 min before the treatment. The head twitches of the mice were induced by DOI (2.5 mg/kg, i.p.). Immediately after the treatment, the number of head twitches was counted during a 20-min session. Lu AF21934, WAY100635 and (R)-(+)-8-hydroxy-DPAT were dosed 60, 45, and 15 min prior to the DOI administration, respectively.
MK-801-induced deficits in social interaction test in rats
Social interaction tests were performed according to the method described by Satow et al. 2009, using a circle made of wood, 90 cm in diameter divided into 10 × 10 cm squares by faint yellow lines. Each social interaction test between two rats was carried out during the light phase of the light/dark cycle. Rats were selected from separate housing cages to make a part for the study. The body weights of the paired rats were matched within 20 g of variance. All rats were placed in an experimental room, and the study was conducted 3.5 h after the subcutaneous administration of MK-801 at a dose of 0.1 mg/kg, s.c. Lu AF21934, WAY100635, and (R)-(+)-8-hydroxy-DPAT were dosed 60, 45, and 15 min prior to the MK-801 administration, respectively.
The test box was wiped clean between each trial. Social interaction between two rats was determined as the total time spent participating in social behavior such as sniffing, genital investigation, chasing, and fighting each other. The total number of social episodes was also measured. In addition, control experiments in animals not receiving MK-801 were also conducted, in order to establish if the drugs had any influence on social behavior when given alone.
Spatial delayed alternation test in rats
The animals, deprived of water overnight, were trained and tested in four wooden T-mazes, which consisted of white and black end-arms (33 × 22 × 25 cm) and a gray starting arm (15 × 20 × 25 cm). The end-arms were equipped with a spout bottle located 9 cm above the floor and containing a 10 % sucrose solution. The three arms were separated from each other by guillotine doors.
Adaptation
On the first 3 days, the animals were allowed freely to explore the whole T-maze for 10 min. On the next 2 days, they were confined to either of the two end-arms and allowed to drink the sucrose solution there for 10 min twice daily.
Training
On the next 2 weeks, the animals received once daily training sessions. Each session consisted of one forced trial (i.e., when one of the end-arm was closed) followed by ten free choice trials. For each free choice trial, the animals were placed in the starting arm, the guillotine doors were raised, and when the rat entered one of the end-arm, the guillotine door was closed, and the rat was allowed to drink the sucrose solution there for 5 s. Then, the rat was gently returned to the starting arm, where it stayed for 10 s (delayed interval). After that time, the guillotine door was raised, and the rat was allowed to enter the end-arms. If the end-arm chosen was opposite to that visited on the previous trial (a correct response), the sucrose solution was provided, and the drinking was allowed for 5 s. If the end-arm chosen was the same as on the previous trial, an incorrect response was scored, and the animal gently returned to the starting arm for 10 s (delayed interval). This training was continued until the animals reach performance criterion, which was defined as at least seven correct responses in ten trials for two consecutive daily sessions.
Testing
The animals were injected with a drug, and the above procedure was repeated; the rat was placed in the starting arm, the guillotine doors raised, and the rat was allowed to enter the end-arms. If it chose the correct end-arm (i.e., opposite to that visited on the previous trial), the sucrose solution was provided; 5 s later, the rat was returned to the starting arm for 10 s (delayed interval). If the end-arm selected was incorrect (i.e., the same as on the previous trial), the rat was returned to the starting arm for 10 s. Such testing sessions were carried out once a week and were preceded by two daily training sessions.
Novel object recognition (NOR)
The method was adapted from Horiguchi et al (2011a, b) and Dere et al. (2007). The animals were trained and tested in a black wooden circular open field (100 cm in diameter, 35 cm high) with the floor divided into 20-cm square sections. The open field was in a dark room illuminated only by a 25 W bulb. On the first day (adaptation), the animals were allowed to explore the open field for 10 min. On the next day (training, T1) the animals were administered with the tested drugs, placed in the apparatus, and allowed to explore two identical objects (cylinder-shaped objects with walls painted white, 7 cm in diameter, 11 cm high) for the time required to complete 15 s of exploration of either object. For the retention trial (T2) conducted 1 h later, one of the objects presented in T1 was replaced with a novel object (a prism-shaped object with walls painted black, 5 cm wide, 14 cm high). The rats were returned to the open field for 5 min, and the duration of exploration (i.e., sitting in close proximity to the objects, sniffing, or touching them) of each object was measured separately by a trained observer. All drugs were administered before the training (T1) session. MK-801 (0.1 mg/kg, s.c) was given 30 min before the session. Lu AF21934, WAY100635, and (R)-(+)-8-hydroxy-DPAT were dosed 60, 45, and 15 min prior to the MK-801 administration, respectively. All injections were given at a volume of 1 ml/kg of the body weight. The treatment groups included eight animals.
In vivo microdialysis
Rats were anesthetized with ketamine (75 mg/kg intramuscular (i.m.)) and xylazine (10 mg/kg i.m.) and placed in a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA, USA). Their skulls will be exposed, and small holes were drilled for the insertion of microdialysis probes into the brain structures using appropriate coordinates (Paxinos and Watson 1982). Vertical microdialysis probes were constructed as described in detail elsewhere (Golembiowska et al. 2012). Following 24 h of the surgery probe, inlets were connected to a syringe pump (BAS, IN, USA) which delivered an artificial CSF composed of [mM]: NaCl 147, KCl 4.0, MgCl2 1.0, CaCl2 2.2; pH = 7.4 at a flow rate of 2.0 μl/min. Baseline samples were collected every 20 min for 2 h after the washout period to obtain a stable extracellular neurotransmitter level. Then, tested drugs were injected, and subsequent fractions of dialysates were collected for 3 h. At the end of the experiment, the rats were sacrificed, and their brains were examined histologically to validate probe placement.
Analytical procedure
DA and 5-HT were analyzed by HPLC with coulochemical detection. Chromatography was performed using an Ultimate 3000 System (Dionex, USA), coulochemical detector Coulochem III (model 5300, ESA, USA) with 5020 guard cell, 5014B microdialysis cell, and Hypersil Gold-C18 analytical column (3 × 100 mm). The mobile phase will be composed of 0.1 M potassium phosphate buffer adjusted to pH = 3.6, 0.5 mM EDTA, 16 mg/l 1-octanesulfonic acid sodium salt, and 2 %. The flow rate during analysis was set at 0.7 ml/min. The applied potential of a guard cell was +600 mV, while those of microdialysis cells were the following: E 1 = −50 mV, E 2 = +300 mV with a sensitivity set at 50 nA/V. The chromatographic data was processed by Chromeleon v. 6.80 (Dionex, USA) software run on a PC computer.
Statistical analysis
The data are presented as the means ± SEM. Statistical analysis of the data was performed using the Graph Pad Prism ver.5 and Statistica 10 package (StatSoft Inc., OK, USA). Two-way ANOVA, followed by Newman-Keuls post hoc comparison test, was used in the interaction studies. The P value of at least P < 0.05 was considered as statistically significant. In vivo microdialysis: All obtained data were given as a percent of basal level assumed as 100 %. The statistical significance was calculated using repeated-measures ANOVA, followed by Tukey’s post hoc test. The results were considered statistically significant when P < 0.05.
Results
The effects of the combined administration of WAY100635 and Lu AF21934 on MK-801-induced hyperactivity in mice
Lu AF21934 administered at a dose of 1 mg/kg, s.c, induced a clear antipsychotic-like effect, reversing MK-801-induced hyperactivity (P < 0.009). WAY100635, administered at a dose of 0.1 mg/kg, i.p., did not have any effect on its own. Co-administration of WAY100635 with Lu AF21934 (1 mg/kg) resulted in the inhibition of Lu AF21934-induced effect in the hyperactivity test. The two-way ANOVA main effects analysis revealed the significant effect of Lu AF21934 × WAY100635 interaction [F(1.29) = 8.02; P < 0.008] (Fig. 1a).

Effects on MK-801-induced hyperactivity. The combined administration of Lu AF21934 (1 mg/kg) with WAY100635 (0.1 mg/kg) (a), and the co-administration of Lu AF21934 (0.1) with 8-OH-DPAT (0.01) (b) are presented. Data are presented as means ± SEM. Doses in milligrams per kilogram are indicated in parentheses. # P < 0.01 versus control, **P < 0.01 versus MK-801-treated group, @P < 0.05 versus Lu AF21934-treated group
The effects of the combined administration of sub-effective doses of (R)-(+)-8-hydroxy-DPAT hydrobromide and Lu AF21934 on MK-801-induced hyperactivity in mice
Lu AF21934 was administered at a dose of 0.1 mg/kg, and (R)-(+)-8-hydroxy-DPAT hydrobromide was given at a dose of 0.01 mg/kg. Neither compound had any effects when dosed separately. Concomitant administration of the sub-effective doses of 5-HT1A receptor agonist and the mGlu4 receptor PAM-induced clear reversal of hyperactivity. Two-way ANOVA main effects analysis revealed the significant effect of (R)-(+)-8-hydroxy-DPAT and Lu AF21934 interaction [F(1.29) = 5.11, P < 0.03]. Post hoc Newman-Keuls analysis revealed the significant effect of (R)-(+)-8-hydroxy-DPAT × Lu AF21934 when compared to the MK-801-treated animals, P < 0.02 (Fig. 1b). The attenuation of the MK-801-induced hyperlocomotion reached 46 %.
The effects of the combined administration of WAY100635 and Lu AF21934 on DOI-induced head twitches in mice
Lu AF21934 administered at a dose of 2 mg/kg significantly inhibited the number of DOI-induced head twitches (P < 0.01). WAY100635, administered at a dose of 0.1 mg/kg did not have any effect on its own. Co-administration of Lu AF21934 and WAY100635 resulted in the inhibition of Lu AF21934-induced effect [F(1.26) = 9.46, P < 0.004]. Post hoc Newman-Keuls analysis revealed a significant Lu AF21934 × WAY100635 interaction when compared to Lu AF21934 treated group, P < 0.01 (Fig. 2a).

Effects on DOI-induced head twitches. The combined administration of Lu AF21934 (5 mg/kg) with WAY100635 (0.1 mg/kg) (a), and the co-administration of Lu AF21934 (0.5 mg/kg) with 8-OH-DPAT (0.01 mg/kg) (b), as well as the administration of lurasidone (0.1 and 0.03 mg/kg) (c), and the effect of the lurasidone + Lu AF21934 combination (d) are presented. Data are presented as means ± SEM. Doses in milligrams per kilogram are indicated in parentheses. *P < 0.05 and **P < 0.01 versus DOI-treated group, @P < 0.05 versus Lu AF21934-treated group
The effect of the combined administration of (R)-(+)-8-hydroxy-DPAT hydrobromide and a sub-effective dose of Lu AF21934 on DOI-induced head twitches in mice
Lu AF21934 was administered at a dose of 1 mg/kg, and (R)-(+)-8-hydroxy-DPAT hydrobromide was given at a dose of 0.01 mg/kg. Neither drug had effects when dosed alone. Concomitant administration of sub-effective doses of 5-HT1A receptor agonist and mGlu4 receptor PAM-induced clear reduction in the number of DOI-induced head twitches. The two-way ANOVA main effects analysis revealed the significant effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction [F(1.26) = 8.25, P < 0.007]. Post hoc Newman-Keuls analysis revealed the significant effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide when compared to the DOI-treated animals, P < 0.0003 (Fig. 2b). The abolishment of the DOI-induced head twitches reached 37 %.
The effect of the combined administration of lurasidone and a sub-effective dose of Lu AF21934 on DOI-induced head twitches in mice
Lurasidone hydrochloride abolished the DOI-induced head twitches at a dose of 0.1 mg/kg, when injected 30 min prior to testing [F(2.19) = 13.45; P < 0.0002]. The dose of 0.03 mg/kg was ineffective. The concomitant administration of ineffective doses of Lu AF21934 (1 mg/kg), and lurasidone (0.03 mg/kg) induced clear antipsychotic-like effect in the DOI-induced head twitch test (Fig. 2c). The two-way ANOVA main effect analysis revealed the significant effect of Lu AF21934 × lurasidone interaction [F(1.27) = 7.31, P < 0.02]. Post hoc Newman-Keuls analysis revealed the significant effect of Lu AF21934 × lurasidone when compared to the DOI-treated animals, P < 0.0002 (Fig. 2d).
The effect of the combined administration of WAY100635 and Lu AF21934 in the social interaction test in rats
Lu AF21934 was given at a dose of 0.5 mg/kg, 60 min before the test. The compound reversed social deficits induced by MK-801 administration (number of social contacts and the total duration of contacts, P < 0.0002). WAY100635 administered at a dose of 0.1 mg/kg had no effect. Co-administration of Lu AF21934 with WAY100635 resulted in the inhibition of Lu AF21934-induced behavioral phenotype in the social interaction test. Two-way ANOVA main effects analysis of the number of episodes revealed the significant effect of Lu AF21934 × WAY100635 interaction [F(1.33) = 8.3, P < 0.006]. Post hoc Newman-Keuls analysis revealed the significant Lu AF21934 × WAY100635 interaction, P < 0.0004, when compared to Lu AF21934-treated group. Two-way ANOVA analysis of the time of interaction also revealed significant effect of Lu AF21934 × WAY100635 [F(1.33) = 6.02; P < 0.01], and post hoc Newman-Keuls analysis revealed the significant effect of Lu AF21934 × WAY100635 interaction, P < 0.004, when compared to Lu AF21934-treated rats (Fig. 3a, b).

Effects on MK-801-induced deficits in social interaction. Number of episodes of social contacts and time of social interactions were measured. Effects of combined administration of Lu AF21934 and WAY100635 in MK-801-treated rats and control experiments (a, b). Effects of combined administration of Lu AF21934 and 8-OH-DPAT in MK-801-treated rats and control experiments (c, d). Data are presented as means ± SEM. Doses in milligrams per kilogram are indicated in parentheses. At least #P < 0.05 versus controls, *P < 0.05 versus MK-801-treated group and @P < 0.05 versus Lu AF21934-treated group
The control experiment with the groups of Lu AF21934, WAY100635, and Lu AF21934 + WAY100635 revealed that Lu AF21934, WAY100635, or Lu AF21934 + WAY100635 interaction had no influence on the number of episodes [F(1.33) = 0.64] (Fig. 3a, b).
The effect of combined administration of (R)-(+)-8-hydroxy-DPAT hydrobromide and a sub-effective dose of Lu AF21934 in the social interaction test in rats
Lu AF21934 was administered at a dose of 0.1 mg/kg, 60 min before the test and did not exhibit any effects. (R)-(+)-8-hydroxy-DPAT hydrobromide was given at a dose of 0.01 mg/kg, 15 min before the test, and also did not have any effects. Simultaneous administration of sub-effective doses of 5-HT1A receptor agonist and mGlu4 receptor PAM-induced clear antipsychotic-like effects measured in two parameters. Two-way ANOVA main effect analysis of the number of episodes revealed the significant effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction [F(1.32) = 7.7; P < 0.009]. Post hoc Newman-Keuls analysis revealed the significant effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide only, P < 0.0001, when compared to all other groups. Two-way ANOVA analysis of the time of interaction revealed significant effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction [F(1.32) = 20.08; P < 0.00008]. Post hoc Newman-Keuls analysis revealed the significant effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction only, P < 0.0001, when compared to all the other groups (Fig. 3c, d).
The control experiment with the groups of Lu AF21934, (R)-(+)-8-hydroxy-DPAT hydrobromide, and Lu AF21934 + (R)-(+)-8-hydroxy-DPAT hydrobromide revealed that neither Lu AF21934, (R)-(+)-8-hydroxy-DPAT hydrobromide, nor Lu AF21934 + (R)-(+)-8-hydroxy-DPAT hydrobromide interaction had any effect on the number of episodes [F(1.33) = 0.76] (Fig. 3c, d).
The effect of combined administration of lurasidone and a sub-effective dose of Lu AF21934 in the social interaction test in rats
Lurasidone hydrochloride was administered at doses of 0.03 and 0.1 mg/kg. The drug administered at a dose of 0.1 mg/kg reversed MK-801-induced deficits only in the time of interaction parameter [F(3.16) = 10.55; P < 0.0005] (Fig. 4a, b). Neither of the doses tested had any influence on the number of social episodes between rats.

Effects on MK-801-induced deficits in social interaction. Dose-dependent effect of lurasidone alone (a, b) and the effect of combined administration of lurasidone and Lu AF21934 (c, d). Data are presented as means ± SEM. Doses in milligrams per kilogram are indicated in parentheses. #P < 0.05 versus controls, **P < 0.05 versus MK-801-treated group versus Lu AF21934-treated group
The concomitant administration of ineffective doses of lurasidone (0.03 mg/kg) and Lu AF21934 (0.1 mg/kg) induced clear antipsychotic-like effect, increasing the duration of interactions between rats disrupted after MK-801 administration [F(1.36) = 4.54; P < 0.03] and number of interactions [F(1.36) = 9.41; P < 0.004] (Fig. 4c, d).
The effect of combined administration of WAY100635 and Lu AF21934 in the novel object recognition test in rats
Lu AF21934 was administered at a dose of 5 mg/kg, 60 min before the test, increasing the recognition index disturbed by MK-801 administration (P < 0.005). WAY100635 was given at a dose of 0.1 mg/kg, 45 min before the test, and did not have any effect. WAY100635, when dosed with Lu AF21934, antagonized Lu AF21934-induced behavioral phenotype in the NOR test. Two-way ANOVA main effects revealed statistical effect of Lu AF21934 × WAY100635 interaction [F(1.28) = 9.17; P < 0.005]. Post hoc Newman-Keuls comparison revealed that WAY100635 antagonized Lu AF21934-induced effect, decreasing recognition index in a statistically significant way (P < 0.005), comparing to Lu AF21934-treated animals (Fig. 5a). The control experiment with the groups of Lu AF21934, WAY100635, and Lu AF21934 + WAY100635 revealed that neither of the combinations interaction had any influence on the recognition index [F(1.28) = 0.92].

Effects on MK-801-induced deficits in NOR. The combined administration of Lu AF21934 (5 mg/kg) with WAY100635 (0.1 mg/kg) (a), and the co-administration of Lu AF21934 (0.5) with 8-OH-DPAT (0.01) (b) are presented. The control experiments with the same group treatments are also included. Data are presented as means ± SEM. Doses in milligrams per kilogram are indicated in parentheses. At least #P <0.05 versus controls, *P < 0.05 versus MK-801-treated group, @P < 0.05 versus Lu AF21934-treated group
The effect of combined administration of (R)-(+)-8-hydroxy-DPAT hydrobromide and a sub-effective dose of Lu AF21934 in the novel object recognition test in rats
Lu AF21934 was given at a dose of 2 mg/kg, 60 min before the test, and (R)-(+)-8-hydroxy-DPAT hydrobromide was given at a dose of 0.01 mg/kg, 15 min before the test. Neither drug had an effect when dosed alone. Concomitant administration of the sub-effective doses of Lu AF21934 and (R)-(+)-8-hydroxy-DPAT hydrobromide induced a clear antipsychotic-like phenotype in the NOR test. Two-way ANOVA main effects revealed statistical effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction [F(1.28) = 7.4; P < 0.01]. Post hoc Newman-Keuls comparison revealed the statistical effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction, P < 0.00001 (Fig. 5b). The control experiment with the groups of Lu AF21934, (R)-(+)-8-hydroxy-DPAT hydrobromide and Lu AF21934 + (R)-(+)-8-hydroxy-DPAT hydrobromide revealed that neither of the combinations interaction had any influence on the recognition index [F(1.28) = 0.65].
The effect of combined administration of lurasidone hydrochloride and a sub-effective dose of Lu AF21934 in the novel object recognition test in rats
Lurasidone was administered at doses of 0.03, 0.1, and 0.5 mg/kg (Fig. 6a). The lowest dose of 0.03 mg/kg was sub-effective, while two higher doses clearly induced the characteristic behavioral phenotype of the NOR paradigm [F(4.43) = 4.89; P < 0.002]. Two-way ANOVA main effects revealed statistical effect of the concomitant administration of sub-effective doses of lurasidone (0.03 mg/kg) and Lu AF21934 (2 mg/kg), by increasing the recognition index that was disrupted by MK-801 administration [F(1.35) = 4.9; P < 0.03]. Post hoc Newman-Keuls comparison revealed the statistical effect of Lu AF21934 × lurasidone interaction, P < 0.001 (Fig. 6b). None of the drug combinations had any effect when administered alone (Fig. 6c).

Effects on MK-801-induced deficits in NOR. The dose-dependent studies of lurasidone (a), and the co-administration of Lu AF21934 (2) with lurasidone (0.03) (b), as well as control experiments (c) are presented. Data are presented as means ± SEM. Doses in milligrams per kilogram are indicated in parentheses. At least #P < 0.05 versus controls, *P < 0.05 versus MK-801-treated group
The effect of combined administration of WAY100635 and Lu AF21934 in the spatial delayed alternation test in rats
Lu AF21934 was given at a dose of 5 mg/kg, 60 min before the test, inducing antipsychotic-like effects in the spatial delayed alteration (SDA) paradigm (P < 0.0007). WAY100635 was given at a dose of 0.1 mg/kg, 45 min before the test, having no own activity. WAY100635, when dosed with Lu AF21934, antagonized Lu AF21934-induced antipsychotic-like effect in the SDA test. Two-way ANOVA main effects revealed statistical effect of Lu AF21934 × WAY100635 interaction [F(1.28) = 4.71; P < 0.03]. Post hoc Newman-Keuls comparison revealed that WAY100635 antagonized Lu AF21934-induced effect, P < 0.0007, comparing to Lu AF21934-treated animals (Fig. 7a). The control experiment with the groups of Lu AF21934, WAY100635, and Lu AF21934 + WAY100635 revealed that neither of the combinations had any effects on the behavior of the animals [F(1.28) = 0.82].

Effects on MK-801-induced deficits in SDA. The combined administration of Lu AF21934 (2 mg/kg) with WAY100635 (0.1 mg/kg) (a), and the co-administration of Lu AF21934 (0.5 mg/kg) with 8-OH-DPAT (0.01 mg/kg) (b) are presented. The control experiments are also included. Data are presented as means ± SEM. Doses in milligrams per kilogram are indicated in parentheses. At least #P < 0.05 versus controls, *P < 0.05 versus MK-801-treated group, @P < 0.05 versus Lu AF21934-treated group
The effect of combined administration of (R)-(+)-8-hydroxy-DPAT hydrobromide and a sub-effective dose of Lu AF21934 in the spatial delayed alteration test in rats
Lu AF21934 was administered at a dose of 2 mg/kg, 60 min before the test, and did not induce any effects on its own. (R)-(+)-8-Hydroxy-DPAT hydrobromide was given at a dose of 0.01 mg/kg, 15 min before the test, and also had no effects. Concomitant administration of sub-effective doses of 5-HT1A receptor agonist and the mGlu4 receptor PAM induced a clear SDA test phenotypic response. Two-way ANOVA main effects revealed statistical effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction [F(1.28) = 13.32; P < 0.001]. Post hoc Newman-Keuls comparison revealed the statistical effect of Lu AF21934 × (R)-(+)-8-hydroxy-DPAT hydrobromide interaction, P < 0.001 (Fig. 7b). The control experiment with the groups of Lu AF21934, (R)-(+)-8-hydroxy-DPAT hydrobromide, and Lu AF21934 + (R)-(+)-8-hydroxy-DPAT hydrobromide revealed that none of the drug combinations had any influence on the rats behavior [F(1.28) = 0.65].
In vivo microdialysis
MK-801 at a dose of 0.3 mg/kg significantly increased DA and 5-HT in the rat frontal cortex reaching maximal effect at 80 and 60 min after administration, respectively (Fig. 8a, b). LU AF 21934 (5 mg/kg) attenuated increase in extracellular DA level induced by MK-801 (Fig. 8a). Repeated measures ANOVA showed significant effect of treatment [F3,16 = 196, P = 0], significant effect of time [F8,128 = 27, P = 0], and interaction between both factors [F24,128 = 15, P = 0].

Effect of Lu AF21934 (5 mg/kg s.c) on extracellular concentration of dopamine (DA) (a) and 5-HT (b) in the rat prefrontal cortex. The figure shows time-course of the DA and 5-HT level between 20 and 180 min of the microdialysis sample collection. Values are presented as the mean ± SEM, n = 12–13 rats. *P < 0.05 and **P < 0.01 show significant differences from basal level
Similarly, LU AF 21934 (5 mg/kg) lowered increased by MK-801 extracellular level of 5-HT (Fig. 8b). Repeated measures ANOVA showed significant effect of treatment [F3,17 = 54, P = 0], significant effect of time [F8,136 = 25, P = 0], and interaction between both factors [F24,136 = 8.2, P = 0].
Basal extracellular levels of DA and 5-HT (in pg/10 μl) in frontal cortex were, respectively, 1.50 ± 0.20 and 0.62 ± 0.06, and there were no significant differences observed between experimental groups.
Discussion
The present studies expand on our recent reports showing that the selective mGlu4 receptor PAMs, Lu AF21934, and Lu AF32615, exhibited antipsychotic-like activity in animal models broadly used with drugs showing clinical efficacy in schizophrenia (Sławińska et al. 2013). Both of these compounds have been extensively characterized, and of special importance to this work, lack any cross-reactivity with 5-HT1A receptors was reported (Bennouar et al. 2013). Herein, we elucidate aspects of the brain circuitry involved in the pharmacology on novel mGlu4 PAMs by exploiting the recently reported functional interactions between mGlu4 and 5-HT1A receptors (Wierońska et al. 2013a, b). We used a number of animal models of schizophrenia, including MK-801-induced hyperactivity, DOI-induced head twitches, social interaction, novel object recognition, and spatial delayed alteration tests. In all of these procedures, the pharmacological cross-talk between mGlu4 and 5-HT1A receptors was verified. Conceptually, these behavioral models may be considered as providing a phenotypic platform reporting on mGlu4 receptor activation in vivo by Lu AF21934.
The mechanism of action of all presently used neuroleptic drugs involves D2 receptor blockade, thought to play a critical role in reversing positive symptoms of psychosis (exaggerations and distortions of normal perception and thinking). However, D2 receptor blockade is not enough to reverse negative and cognitive psychotic symptoms, making typical neuroleptics such as haloperidol not efficient in these aspects. The novel antipsychotic drugs that tend to act on 5-HT1A receptors, such as clozapine, olanzapine, risperidone, or lurasidone, demonstrated some benefits for negative symptoms in the clinic (Innamorati et al. 2013; Stauffer et al. 2012).
We recently reported our investigations on the role of 5-HT1A receptors in the preclinical pharmacological actions of various mGlu receptor ligands (Wierońska et al. 2013a, b). The unique role of these receptors was proposed in preclinical models of cognitive disturbances of schizophrenia for atypical neuroleptics (APD) known to have agonist activity at the 5-HT1A receptor (e.g., risperidon and lurasidone) (Nagai et al. 2009; Snigdha et al. 2011; Horiguchi and Meltzer 2013; Horiguchi et al. 2013). Such an activity is a common attribute with selective agonists at 5-HT1A receptors, such as 8-OH-DPAT or tandospirone, which have shown efficacy in a number of animal models of schizophrenia, e.g., novel object recognition test (Bubeníková-Valesová et al. 2007; Horiguchi and Meltzer 2012; Horiguchi et al. 2012).
In our present investigations, we used a number of animal models thought to represent positive, negative, and cognitive symptoms of schizophrenia. As previously, we used MK-801-induced hyperactivity, DOI-induced head twitches, the social interaction test, novel object recognition test, and spatial delayed alteration task. This wide range of methods may reflect on the main aspects of the pathophysiology of schizophrenia, mirroring the clinical picture of the disease, and allowing for broadly conceived behavioral research. Historically, these standard pharmacological protocols have worked well in our laboratories, they all are well-known and well-established procedures with excellent predictive validity, e.g., positive response is observed predominantly for antipsychotic drugs, and not for other class of psychotropics, such as antidepressants or anxiolytics.
In our studies, a number of well-known, validated chemical probes were used, such as WAY100635 as a selective inhibitor of the 5-HT1A receptor-mediated signaling, and the agonist (R)-(+)-8-hydroxy-DPAT as a stimulator of the signaling (Foster and Goa 1999; Cosi and Koek 2000). We also used lurasidone, an antipsychotic drug recently receiving FDA approval, that has strong affinity toward 5-HT1A receptors (K i = 6.8 nM) and demonstrated partial agonist functionality (Horiguchi and Meltzer 2012; Horiguchi et al. 2012).
Previously, we showed that low doses of 5-HT1A receptor ligands inhibited (WAY100635) or enhanced ((R)-8-hydroxy-DPAT) the pharmacological effects of LSP1-2111, a brain penetrant, mGlu4-preferred orthosteric agonist (Wierońska et al. 2013a, b; Cajina et al. 2014). We used the 5-HT1A receptor antagonist WAY100635 at a dose of 0.1 mg/kg, and the agonist (R)-(+)-8-hydroxy-DPAT at a dose of 0.01 mg/kg. At that dose, WAY100635 inhibited the action of an effective dose of LSP1-2111, while a sub-effective dose of (R)-(+)-8-hydroxy-DPAT given together with a sub-effective dose of LSP1-2111, enhanced its action (Wierońska et al. 2013a, b). Higher doses of WAY100635 (1 mg/kg and above) or (R)-(+)-8-hydroxy-DPAT (0.5 mg/kg and above) induced their own effects, namely, antipsychotic-like behaviors or the so-called serotonergic syndrome, respectively. It is worth to mention that (R)-(+)-8-hydroxy-DPAT given at the dose of 0.025 mg/kg had clear antipsychotic-like action.
In our present studies, the actions of Lu AF21934 were inhibited by WAY100635 in all tests conducted, suggesting that in these models, 5-HT1A receptors play an important mechanistic role in the expression of Lu AF21934-mediated effects, independently of the mechanism of mGlu4 receptor activation.
To further establish the role of the 5-HT1A receptor in the actions mediated by Lu AF21934, a sub-effective dose of (R)-(+)-8-hydroxy-DPAT, together with a sub-effective dose of Lu AF21934, were administered. As seen previously with LSP1-2111, the drugs combination reversed MK-801-induced dysfunctions, decreasing the hyperactivity and increasing social behavior in the social interaction test. The improvement of the recognition index in the NOR paradigm and the reversal of the SDA dysfunction were also evident. The drugs combinations had no any own effects when tested in separated tests.
In this work, we present mechanistic evidence of an interaction between the 5-HT1A and mGlu4 receptors, and we propose a multidirectional action of the novel class of neuroleptics. Marketed antipsychotics are multimodal drugs, targeting several receptors. For example, lurasidone is an antipsychotic recently approved in the USA and Canada for the treatment of schizophrenia and bipolar depression. Like other atypical antipsychotics, such as ziprasidone and clozapine, lurasidone is a high-affinity partial agonist at the 5-HT1A receptor with known efficacy in preclinical studies in some animal models of psychotic disturbances (Schmidt et al. 2001; Sprouse et al. 1999; Horiguchi and Meltzer 2012; Horiguchi et al. 2012). To gain support for our developing hypothesis of mGlu4 receptor/5-HT1A receptor interactions, we tested the novel antipsychotic lurasidone in preclinical models thought to represent the positive, negative and cognitive disturbances (DOI-induced head twitches, social interaction and novel object recognition test, respectively). While lurasidone has been widely discussed in the field for its efficacy in a model of cognitive disturbances (novel object recognition test, Horiguchi et al. 2012), to our best knowledge, no reports exist on its effectiveness in animal models thought to reflect positive and negative symptoms. Therefore, this work for the first time shows the efficacy of lurasidone in DOI-induced head twitches and in the social interaction models. However, in the latter procedure, the drug reversed MK-801-induced deficit in time of interaction and not in the number of episodes. The compound was active in a relatively narrow range of doses. In the combination studies, we used lurasidone at the sub-effective dose of 0.03 mg/kg, together with a sub-effective dose of Lu AF21934, relevant to each particular test. In all of these procedures, we showed that the combination effectively reversed all phenotypes induced by psychotomimetic agents. Interestingly, in the social interaction test, the combination was effective also in reversing deficit in the number of episodes, not influenced by lurasidone alone. All those results suggest a crosstalk between mGlu4 and 5-HT1A receptors, with potential implications for antipsychotic medications.
We are aware that the pharmacological tools used in the present paper are not enough to claim whether 5-HT1A receptors are necessary for mGlu4-based expression of the phenotypic responses explored herein. However, the results constitute strong support for our previous studies with orthosteric, preferential mGlu4 agonists, LSP1-2111, suggesting the possibility of the existence of a pharmacological crosstalk between mGlu4 and 5-HT1A receptors.
Considering the putative mechanism of this interaction, the theory of antipsychotic activity of mGlu ligands raised by Conn et al. (2009) and the data on the distribution of 5-HT1A receptors in the brain will be analyzed. According to Conn’s theory, schizophrenia is a consequence of the dysfunction of NMDA receptors expressed postsynaptically on GABAergic interneurons. The following loss of GABAergic inhibition over glutamatergic thalamocortical projections results in the enhancement of glutamate release. Presynaptically expressed mGlu4 receptors would counteract this increased glutamatergic activity (see: Conn et al. 2009; Sławińska et al. 2013).
On the other hand, increased level of serotonin is also thought to be responsible for the induction of psychotic symptoms. Variety of studies, including our present results, show that MK-801 may induce increase not only in dopamine but also serotonin release that presumably contributes to the observed MK-801-induced disturbations (López-Gil et al. 2009; Wedzony et al. 1993). With the use of in vivo microdialysis studies, we showed that MK-801-induced increase in dopamine and, most importantly, serotonin release in the prefrontal cortex was counteracted by the Lu AF21934 administration, supporting the hypothesis of the interaction between serotonergic system and mGlu4 receptors in MK-801 evoked actions.
The exact mechanism, by which mGlu4 receptors activation contributes to inhibition of that serotonergic outflow from terminals projecting to the cortex, and the role of 5-HT1A receptors in that chain of events remain to be explored. However, the existence of such an interaction may have important practical connotations, as the use of two different compounds exerting therapeutic efficacy in combination creates novel treatment opportunities, which may be more effective and less burdened with the risk of adverse effects than monotherapy.
In summary, we propose that mGlu4 and 5-HT1A receptors play synergistically in the regulation of glutamatergic activity in the CNS and therefore may constitute a dual target for putative antipsychotic treatment.
References
Bennouar KE, Uberti MA, Melon C, Bacolod MD, Jimenez HN, Cajina M, Kerkerian-Le Goff L, Doller D, Gubellini P (2013) Synergy between L-DOPA and a novel positive allosteric modulator of metabotropic glutamate receptor 4: implications for Parkinson's disease treatment and dyskinesia. Neuropharmacology 66:158–169
Bickel S, Javitt DC (2009) Neurophysiological and neurochemical animal models of schizophrenia: focus on glutamate. Behav Brain Res 204(2):352–362
Boulay D, Depoortre R, Louis C, Perrault G, Griebel G, Soubrié P (2004) SSR181507, a putative atypical antipsychotic with dopamine D2 antagonist and 5-HT1A agonist activities: improvement of social interaction deficits induced by phencyclidine in rats. Neuropharmacology 46(8):1121–1129
Bubeníková-Valesová V, Votava M, Pálenícek T, Horácek J (2007) The opposite effect of a low and a high dose of serotonin-1A agonist on behavior induced by MK-801. Neuropharmacology 52(4):1071–1078
Burnet PW, Eastwood SL, Harrison PJ (1996) 5-HT1A and 5-HT2A receptor mRNAs and binding site densities are differentially altered in schizophrenia. Neuropsychopharmacology 15(5):442–455
Cajina M, Nattini M, Song D, Smagin G, Jørgensen EB, Chandrasena G, Bundgaard C, Toft DB, Huang X, Acher F, Doller D (2013) Qualification of LSP1-2111 as a brain penetrant group III metabotropic glutamate receptor orthosteric agonist. ACS Med Chem Lett 5(2):119–123
Cartmell J, Monn JA, Schoepp DD (2000) Attenuation of specific PCP-evoked behaviors by the potent mGlu2/3 receptor agonist, LY379268 and comparison with the atypical antipsychotic, clozapine. Psychopharmacol (Berl) 148(4):423–429
Conn PJ, Pin JP (1997) Pharmacology and functions of metabotropic glutamate receptors. Annu Rev PharmacolToxicol 37:205–237
Conn PJ, Lindsley CW, Jones CK (2009) Activation of metabotropic glutamate receptors as a novel approach for the treatment of schizophrenia. Trends Pharmacol Sci 30(1):25–31
Cosi C, Koek W (2000) The putative <<silent>> 5-HT1A receptor antagonist, WAY 100635, has inverse agonist properties at cloned human 5-HT1A receptors. Eur J Pharmacol 401(1):9–15
Dere E, Huston JP, De Souza Silva MA (2007) The pharmacology, neuroanatomy and neurogenetics of one-trial object recognition in rodents. NeurosciBiobehav Rev 31(5):673–704
Fell MJ, Svensson KA, Johnson BG, Schoepp DD (2008) Evidence for the role of metabotropic glutamate (mGlu)2 not mGlu3 receptors in the preclinical antipsychotic pharmacology of the mGlu2/3 receptor agonist (-)-(1R,4S,5S,6S)-4-amino-2-sulfonylbicyclo[3.1.0]hexane-4,6-dicarboxylic acid (LY404039). J Pharmacol Exp Ther 326(1):209–217
Foster RH, Goa KL (1999) Olanzapine. A pharmacoeconomic review of its use in schizophrenia. Pharmacoeconomics 15(6):611–640
Galici R, Echemendia NG, Rodriguez AL, Conn PJ (2005) A selective allosteric potentiator of metabotropic glutamate (mGlu) 2 receptors has effects similar to an orthosteric mGlu2/3 receptor agonist in mouse models predictive of antipsychotic activity. J Pharmacol Exp Ther 315(3):1181–1187
Geyer MA, Ellenbroek B (2003) Animal behavior models of the mechanisms underlying antipsychotic atypicality. ProgNeuropsychopharmacolBiol. Psychiatry 27(7):1071–1079
Golembiowska K, Kowalska M, Bymaster FP (2012) Effects of the triple reuptake inhibitor amitifadine on extracellular levels of monoamines in rat brain regions and on locomotor activity. Synapse 66(5):435–444
Hagiwara H, Fujita Y, Ishima T, Kunitachi S, Shirayama Y, Iyo M, Hashimoto K (2008) Phencyclidine-induced cognitive deficits in mice are improved by subsequent subchronic administration of the antipsychotic drug perospirone: role of serotonin 5-HT1A receptors. EurNeuropsychopharmacol 18(6):448–454
Hopkins CR (2013) Is there a path forward for mGlu(2) positive allosteric modulators for the treatment of schizophrenia? ACS ChemNeurosci 4(2):211–213
Horiguchi M, Meltzer HY (2012) The role of 5-HT1A receptors in phencyclidine (PCP)-induced novelobjectrecognition (NOR) deficit in rats. Psychopharmacology (Berl) 221(2):205–215
Horiguchi M, Meltzer HY (2013) Blonanserin reverses the phencyclidine (PCP)-induced impairment in novelobjectrecognition (NOR) in rats: role of indirect 5-HT(1A) partial agonism. Behav Brain Res 247:158–164
Horiguchi M, Huang M, Meltzer HY (2011a) The role of 5-hydroxytryptamine 7 receptors in the phencyclidine-induced novelobjectrecognition deficit in rats. J Pharmacol Exp Ther 338(2):605–614
Horiguchi M, Huang M, Meltzer HY (2011b) Interaction of mGlu2/3 agonism with clozapine and lurasidone to restore novelobjectrecognition in subchronic phencyclidine-treated rats. Psychopharmacol (Berl) 217(1):13–24
Horiguchi M, Hannaway KE, Adelekun AE, Jayathilake K, Meltzer HY (2012) Prevention of the phencyclidine-induced impairment in novelobjectrecognition in female rats by co-administration of lurasidone or tandospirone, a 5-HT1A partial agonist. Neuropsychopharmacology 37(10):2175–2183
Horiguchi M, Hannaway KE, Adelekun AE, Huang M, Jayathilake K, Meltzer HY (2013) D(1) receptor agonists reverse the subchronic phencyclidine (PCP)-induced novel object recognition (NOR) deficit in female rats. Behav Brain Res 238:36–43
Ichikawa J, Li Z, Dai J, Meltzer HY (2002) Atypical antipsychotic drugs, quetiapine, iloperidone, and melperone, preferentially increase dopamine and acetylcholine release in rat medial prefrontal cortex: role of 5-HT1A receptor agonism. Brain Res 956(2):349–357
Innamorati M, Baratta S, Di Vittorio C, Lester D, Girardi P, Pompili M, Amore M (2013) Atypicalantipsychotics in the treatment of depressive and psychotic symptoms in patients with chronic schizophrenia: a naturalistic study. Schizophr Res Treatment 2013:423205
Javitt DC, Zukin SR (1991) Recent advances in the phencyclidine model of schizophrenia. Am J Psychiatry 148(10):1301–1308, Review
Javitt DC, Balla A, Burch S, Suckow R, Xie S, Sershen H (2004) Reversal of phencyclidine-induced dopaminergic dysregulation by N-methyl-D-aspartate receptor/glycine-site agonists. Neuropsychopharmacology 29(2):300–307
Kinon BJ, Zhang L, Millen BA, Osuntokun OO, Williams JE, Kollack-Walker S, Jackson K, Kryzhanovskaya L, Jarkova N, HBBI Study Group (2011) A multicenter, inpatient, phase 2, double-blind, placebo-controlled dose-ranging study of LY2140023 monohydrate in patients with DSM-IV schizophrenia. J ClinPsychopharmacol 31(3):349–355
Krystal JH, Anand A, Moghaddam B (2002) Effects of NMDA receptor antagonists: implications for the pathophysiology of schizophrenia. Arch Gen Psychiatry 59(7):663–664
Leite JV, Guimarães FS, Moreira FA (2008) Aripiprazole, an atypical antipsychotic, prevents the motor hyperactivity induced by psychotomimetics and psychostimulants in mice. Eur J Pharmacol 578(2–3):222–227
López-Gil X, Artigas F, Adell A (2009) Role of different monoamine receptors controlling MK-801-induced release of serotonin and glutamate in the medial prefrontal cortex: relevance for antipsychotic action. Int J Neuropsychopharmacol 12(4):487–499
Moghaddam B, Jackson ME (2003) Glutamatergic animal models of schizophrenia. Ann N Y Acad Sci 1003:131–137
Nagai T, Murai R, Matsui K, Kamei H, Noda Y, Furukawa H, Nabeshima T (2009) Aripiprazole ameliorates phencyclidine-induced impairment of recognition memory through dopamine D1 and serotonin 5-HT1A receptors. Psychopharmacology (Berl) 202(1–3):315–328
Pałucha-Poniewiera A, Kłodzińska A, Stachowicz K, Tokarski K, Hess G, Schann S, Frauli M, Neuville P, Pilc A (2008) Peripheral administration of group III mGlu receptor agonist ACPT-I exerts potential antipsychotic effects in rodents. Neuropharmacology 55(4):517–524
Patil ST, Zhang L, Martenyi F, Lowe SL, Jackson KA, Andreev BV, Avedisova AS, Bardenstein LM, Gurovich IY, Morozova MA, Mosolov SN, Neznanov NG, Reznik AM, Smulevich AB, Tochilov VA, Johnson BG, Monn JA, Schoepp DD (2007) Activation of mGlu2/3 receptors as a new approach to treat schizophrenia: a randomized Phase 2 clinical trial. Nat Med 13(9):1102–1107
Paxinos G, Watson C (1982) The rat brain in stereotaxic coordinates
Pin JP, Duvoisin R (1995) The metabotropic glutamate receptors: structure and functions. Neuropharmacology 34(1):1–26
Rorick-Kehn LM, Johnson BG, Knitowski KM, Salhoff CR, Witkin JM, Perry KW, Griffey KI, Tizzano JP, Monn JA, McKinzie DL, Schoepp DD (2007a) In vivo pharmacological characterization of the structurally novel, potent, selective mGlu2/3 receptor agonist LY404039 in animal models of psychiatric disorders. Psychopharmacology (Berl) 193(1):121–136, Epub2007 Mar 24
Rorick-Kehn LM, Johnson BG, Burkey JL, Wright RA, Calligaro DO, Marek GJ, Nisenbaum ES, Catlow JT, Kingston AE, Giera DD, Herin MF, Monn JA, McKinzie DL, Schoepp DD (2007b) Pharmacological and pharmacokinetic properties of a structurally novel, potent, and selective metabotropic glutamate 2/3 receptor agonist: in vitro characterization of agonist (-)-(1R,4S,5S,6S)-4-amino-2-sulfonylbicyclo[3.1.0]-hexane-4,6-dicarboxylic acid (LY404039). J Pharmacol Exp Ther 321(1):308–317
Satow A, Suzuki G, Maehara S, Hikichi H, Murai T, Murai T, Kawagoe-Takaki H, Hata M, Ito S, Ozaki S, Kawamoto H, Ohta H (2009) Unique antipsychotic activities of the selective metabotropic glutamate receptor 1 allosteric antagonist 2-cyclopropyl-5-[1-(2-fluoro-3-pyridinyl)-5-methyl-1H-1,2,3-triazol-4-yl]-2,3-dihydro-1H-isoindol-1-one. J PharmacolExpTher 330(1):179–190
Schmidt AW, Lebel LA, Howard HR Jr, Zorn SH (2001) Ziprasidone: a novel antipsychotic agent with a unique human receptor binding profile. Eur J Pharmacol 425(3):197–201
Schoepp DD, Marek GJ (2002) Preclinical pharmacology of mGlu2/3 receptor agonists: novel agents for schizophrenia? Curr Drug Targets CNS Neurol Disord 1(2):215–225
Simpson MD, Lubman DI, Slater P, Deakin JF (1996) Autoradiography with [3H]8-OH-DPAT reveals increases in 5-HT(1A) receptors in ventral prefrontal cortex in schizophrenia. Biol Psychiatry 39(11):919–928
Sławińska A, Wierońska JM, Stachowicz K, Marciniak M, Lasoń-Tyburkiewicz M, Gruca P, Papp M, Kusek M, Tokarski K, Doller D, Pilc A (2013) The antipsychotic-like effects of positive allosteric modulators of metabotropic glutamate mGlu4 receptors in rodents. Br J Pharmacol 169(8):1824–1839
Snigdha S, Idris N, Grayson B, Shahid M, Neill JC (2011) Asenapine improves phencyclidine-induced object recognition deficits in the rat: evidence for engagement of a dopamine D1 receptor mechanism. Psychopharmacology (Berl) 214(4):843–853
Sprouse JS, Reynolds LS, Braselton JP, Rollema H, Zorn SH (1999) Comparison of the novel antipsychotic ziprasidone with clozapine and olanzapine: inhibition of dorsal raphe cell firing and the role of 5-HT1A receptor activation. Neuropsychopharmacology 21(5):622–631
Stauffer VL, Song G, Kinon BJ, Ascher-Svanum H, Chen L, Feldman PD, Conley RR (2012) Responses to antipsychotic therapy among patients with schizophrenia or schizoaffective disorder and either predominant or prominent negative symptoms. Schizophr Res 134(2–3):195–201
Wedzony K, Klimek V, Gołembiowska K (1993) MK-801 elevates the extracellular concentration of dopamine in the rat prefrontal cortex and increases the density of striatal dopamine D1 receptors. Brain Res 622(1–2):325–329
Wedzony K, Maćkowiak M, Zajaczkowski W, Fijał K, Chocyk A, Czyrak A (2000) WAY100135, an antagonist of 5-HT1A serotonin receptors, attenuates psychotomimetic effects of MK-801. Neuropsychopharmacology 23(5):547–559
Wierońska JM, Kusek M, Tokarski K, Wabno J, Froestl W, Pilc A (2011) The GABA B receptor agonist CGP44532 and the positive modulator GS39783 reverse some behavioural changes related to positive syndromes of psychosis in mice. Br J Pharmacol 163(5):1034–1047
Wierońska JM, Stachowicz K, Acher F, Lech T, Pilc A (2012) Opposing efficacy of group III mGlu receptor activators, LSP1-2111 and AMN082, in animal models of positive symptoms of schizophrenia. Psychopharmacology (Berl) 220(3):481–494
Wierońska JM, Sławińska A, Stachowicz K, Łasoń-Tyburkiewicz M, Gruca P, Papp M, Pilc A (2013a) The reversal of cognitive, but not negative or positive symptoms of schizophrenia, by the mGlu2/3 receptor agonist, LY379268, is 5-HT1A dependent. Behav Brain Res 256:298–304
Wierońska JM, Acher FC, Sławińska A, Gruca P, Lasoń-Tyburkiewicz M, Papp M, Pilc A (2013b) The antipsychotic-like effects of the mGlu group III orthosteric agonist, LSP1-2111, involves 5-HT1A signalling. Psychopharmacol (Berl) 227(4):711–725
Acknowledgments
The study was supported by the Statutory Funds of the Polish Academy of Science and by the National Science Centre, Grant no. 2012/6/06/A/NZ7/00014 (MAESTRO) given to A. Pilc.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
ESM 1
(DOCX 44 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article

Cite this article
Wierońska, J.M., Sławińska, A., Łasoń-Tyburkiewicz, M. et al. The antipsychotic-like effects in rodents of the positive allosteric modulator Lu AF21934 involve 5-HT1A receptor signaling: mechanistic studies. Psychopharmacology 232, 259–273 (2015). https://doi.org/10.1007/s00213-014-3657-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00213-014-3657-4