
Abstract
Alpine landscapes are notable regarding their sensitivity to environmental change. Surface waters are especially sensitive as many are influenced by glacial meltwaters that are diminishing with the retreat of glaciers worldwide. The Macun Lakes region, Swiss National Park, is an alpine cirque landscape housing a number of lakes interconnected by streams. The area has been non-glaciated for decades, although rock glaciers are still present in the south basin. Surface waters, 10 stream sites in the present study, have been monitored in mid-summer since the year 2001 for physico-chemistry, periphyton and macroinvertebrates (including 74 species of chironomids). Water physico-chemistry revealed that the two main basins in the catchment differed, reflecting the inputs of rock glacier waters in the south basin. However, trends suggest that waters are becoming more similar as rock glacier inputs diminish over time along with a general decrease in nitrogen levels in all waters. Periphyton biomass showed some spatial differences among sites and a general decrease after 2010. Macroinvertebrate assemblages, including chironomids, clearly differed among basins and spatially along the stream network in each basin. Notably, no significant temporal trend was observed in the long-term data for macroinvertebrates, including chironomids, at the monitored sites. The results suggest that lotic macroinvertebrates may be buffered by the interconnectedness of streams and lakes in the landscape, which mitigates major response patterns of running waters to environmental change.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Alpine landscapes and associated waters are being rapidly transformed as climate-related warming continues, reflecting ongoing glacial recession globally (Milner et al. 2017). The reduction and loss of glaciers project a substantial decrease in the influence of glacial waters on alpine fluvial networks (Milner et al. 2009; Wilkes et al. 2023). Alpine waters are inhabited by a unique diversity of organisms adapted to the relatively insular landscape of high elevations with distinct environmental conditions that limit the kinds and numbers of biota (Jacobsen et al. 2017; Füreder et al. 2000). With landscape transformation, alpine environments become accessible to lower elevation flora and fauna, as currently documented across the globe (Cauvy-Fraunie et al. 2016; Brown et al. 2017). The increased invasibility of alpine waters, which may in part be related to novel environmental conditions that increase habitat suitability for new taxa, increasing the potential for biotic interactions such as predator–prey relations or competition that cause reductions in and loss of alpine species. Long-term monitoring allows for the documentation and understanding of shifts in the distribution and abundance of organisms over time and in relation to changes in the environment.
Freshwater macroinvertebrates are commonly used as bio-indicators of environmental conditions of surface waters. They are considered sensitive to alterations in physico-chemistry associated with natural disturbances and human-caused perturbations to freshwaters, in particular landscape (wildfire, climate change) and land use (urban, agriculture)-related changes (Allan 2004). In this regard, alpine landscapes have been subject to intense and rapid transformations in response to climate-related environmental change (IPCC 2023), with observed alterations in the distribution and abundance of flora and fauna (Brown et al. 2007; Jacobsen et al. 2014). Current models further stress the reduction and potential loss of alpine aquatic macroinvertebrates, especially cold-water specialists, as these landscapes and their surface waters continue to change with ongoing glacial recession, which has important implications for the availability of potential refugia (Wilkes et al. 2023). The present study documents spatial and temporal changes in the distribution and abundance of lotic macroinvertebrates in a high alpine cirque landscape over the past 21 years. We also examine whether the juxtaposition of lakes and streams in the landscape provide an environmental buffering effect for macroinvertebrates to ongoing landscape transformation.
The aquatic dipterans Chironomidae are ubiquitous biota of alpine waters (Lencioni and Rossaro 2005; Füreder et al. 2006; Lods-Crozet et al. 2012; Montagna et al. 2016). There are numerous species in the Chironomidae, particularly in the subfamily Diamesinae, which are considered cold water specialists and are mostly endemic to the cold waters of alpine aquatic habitats (Robinson et al. 2007; Lods-Crozet et al. 2012; Lencioni et al. 2015). Most alpine chironomids are plurivoltine and thus can colonize and inhabit the intermittent streams common in alpine fluvial networks (Chanut et al. 2022). The Orthocladiinae are another common subfamily that play important roles in alpine freshwaters (Füreder et al. 2006), often replacing the Diamesinae in alpine waters less harsh than those flowing directly from glaciers, so-called proglacial or kyral streams (Finn et al. 2010; Robinson et al. 2016). Because the Macun region encompasses a diversity of water types (a composite of lakes, ponds, streams and springs) influenced differentially by glacial inputs (see Robinson et al. 2022), the potential for a diverse assemblage of chironomids is expected. In fact, some 46 species of chironomids were documented in our earlier studies on the Macun catchment (Lods-Crozet et al. 2012), and the current study adds 28 species to that list (see Supplementary Table 1). Due to this high diversity of taxa and associated biotic traits, the chironomids are excellent organisms, i.e. sentinels, for the study of long-term changes in aquatic habitats of alpine landscapes (e.g., Lencioni et al. 2012).
The primary goal of this paper is to present the long-term patterns (over a period of 21 years) in macroinvertebrate assemblages associated with fluvial surface waters of the Macun Lakes region. We predicted that assemblages would differ spatially among sites in relation to differences in water physico-chemistry. Our previous studies have demonstrated that the physico-chemistry of surface waters differs spatially in the catchment in relation to basin-scale differences associated with inputs from rock glacier waters (see site description below; Robinson et al. 2022). We further predicted a spatial change in macroinvertebrates associated with, first, the merging of basin waters at the outlet site (site 9, see Fig. 1) of the catchment and, second, water inputs of a large rock glacier that influenced waters of the lowermost monitoring site (Zeznina, site 10). Previous analyses have shown a change in surface water physico-chemistry, with surface waters prior to 2010 differing significantly from waters post-2010, and with surface waters showing a general homogenization over the 21-year study period (see Robinson et al. 2022). We thus expected macroinvertebrate assemblages to reflect this temporal change in surface water physico-chemistry over the study period, as was shown for diatoms collected at the same sites over the same study period (see Peszek et al. 2022). A highlight of the current study is the identification of morphospecies of the Chironomidae for most years (see Methods). Chironomids are a dominant group of macroinvertebrates in alpine surface waters, not only in number of individuals but also in number of species (Lods-Crozet et al. 2012). Thus, another objective of this paper is to summarize the results for the chironomids in relation to the predictions generated for macroinvertebrates in general (e.g., do the chironomid data add information that is not evident in the general macroinvertebrate dataset?).

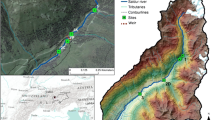
Map of the Macun Lakes region in the Swiss National Park with study sites labeled. Sites 1–4 are located in the north basin and sites 5–8 are in the south basin of the catchment. Note the large rock glacier located between sites 9 (outlet stream) and 10 that strongly influenced the physico-chemistry of the stream at site 10
Site description
The Macun Lakes (46°44’N, 10°08’E) is an alpine cirque [> 2600 m above sea level (a.s.l.)] in the canton of Graubunden, Switzerland (Fig. 1). The 3.6-km2 region has been a designated area for long-term monitoring of alpine surface waters in the Swiss National Park since its annexation in 2000. Minimal livestock grazing (20–30 cows in summer) occurred in the area prior to it becoming part of the national park. Peaks around the catchment have elevations between 2800 and 3000 m a.s.l., and study site elevations range between 2610–2650 m a.s.l. Precipitation is around 850 mm year–1, and air temperature ranges from > 20 °C in summer to < −25 °C in winter (Buffalora climate station ~ 14 km southeast of Macun). Local geology is slow-weathering crystalline (orthogneiss, meta-granitoid) rock. The study area is above the tree line, thus the terrestrial vegetation comprises typical alpine grasses and low-lying herbs, with areas of bare rock.
The Macun drainage comprises a north and south basin (Fig. 1), including 36 small lakes and temporary ponds. Four large lakes (ca 0.12 km2 each, < 10 m deep) are interconnected by streams, each < 500 m in length (Robinson and Oertli 2009). The outlet stream (Zeznina) of the catchment flows into the Inn River near Lavin, Switzerland. Catchment water sources include precipitation and remnant rock glaciers in the south basin [there have been no active glaciers in the catchment since the early 1900s (https://map.geo.admin.ch/]. The water source in each basin causes differences in water physico-chemistry; water temperatures are lower in the south than in the north basin (Robinson et al. 2022). Inputs from a large, active rock glacier [Macun1 (Barsch 1969; Fehr and Reich 2015; Derungs and Tischhauser 2017)] influence surface waters in Zeznina stream, where site 10 is located in this study (see Fig. 1). Stream channels in each basin have low gradients and contain mostly stable cobble substrate. Fish (Salmo trutta fario, Salvenius namaycush, Phoxinus phoxinus) are present in the large lakes, which were last stocked in 1993 (P. Rey, personal communication).
Methods
Field monitoring was conducted each year from 2001 in mid-summer, between 27 July and 1 August. Various kinds of samples were collected from the 10 long-term monitoring sites used for the overall study (Fig. 1). As part of the present study, water samples were collected in 0.5-L pre-rinsed plastic bottles and sent to the Aua laboratory at Eawag for the analysis of nitrogen- and phosphorus-containing constituents, dissolved organic carbon, particulate organic carbon, total inorganic carbon, pH, and silicate, following methods in Tockner et al. (1997) and Robinson and Matthaei (2007). Further, spot measures of electrical conductivity and temperature (WTW LF 323), and turbidity (Cosmos; Züllig, Switzerland), were recorded directly in the field using portable meters.
Periphyton was collected at each site every year (except 2006 and 2014) by scraping the surface of five haphazardly selected stones. Stones were scraped with a metal brush and subsamples (10–15 ml) of the slurry filtered through Whatman GF/F glass fiber filters. The filters were sent to the laboratory and frozen (-25 °C) until analysis. For analysis, each filter was dried at 60 °C, weighed, combusted at 550 °C for 4 h, and then re-weighed for estimates of ash-free dry mass.
Benthic macroinvertebrates were sampled semi-quantitatively using a timed (5 min) kick-net approach. Along a 30-m reach at each site, benthic substrata were disturbed and the loosened material, including macroinvertebrates, was collected in the net (250 µm mesh). All major habitat types (pools, runs, riffles) were sampled during the 5-min sampling period. The material collected was preserved with 70% ethanol for later processing in the laboratory. Species collections of chironomids were supplemented by using Malaise traps and drift nets (2013, 2014) near three lakes (Grond, Mezza Gluna, Immez) for 24-h drift samples [taken in the last 2 years at the two inlets of Lai Immez and the outlet stream of the large rock glacier, Macun1 (Barsch 1969)], located ca. 50 m below site 9] and various samples from selected lakes and ponds (see Lods-Crozet et al. 2012). These latter collections, except the lake samples, were only used to record and search for rare and new species living in particular environments in the catchment and not for long-term monitoring or analysis. In the laboratory, all macroinvertebrates were handpicked from each sample, identified to the lowest possible taxonomic unit, and counted (after Tachet et al. 2010). Notably, Chironomidae were identified to morphospecies for most years (excluding 2003, 2004, 2005, 2008, 2009 and 2013), based on the examination of slide mounts of larvae, pupae or male adults (Lehmann 1972; Wiederholm 1983, 1986, 1989; Sæther 1990, 1995; Schmid 1993; Langton 1991; Ekrem 2004; Stur and Ekrem 2006; Ilyashuk et al. 2010; Ferrington and Sæther 2011; Stur and Spies 2011; Langton et al. 2013; Rossaro et al. 2017).
Data analysis
The physico-chemistry data were summarized using principal components analysis (PCA) on log(x + 1)-transformed data (except for pH). As a detailed assessment has been presented in Robinson et al. (2022), only a general summary is given in this paper to present potential relationships between habitats and macroinvertebrates. Periphyton biomass among sites and dates was tested for differences using two-way ANOVA on log(x + 1)-transformed data followed by a Tukey’s post hoc test when differences were significant (Zar 1984). To better illustrate spatial patterns over time, the periphyton data are summarized by basin (north and south), Immez outlet and Zeznina over the study period. As all the periphyton data have been summarized in Peszek et al. (2022), data are presented in this paper to simply show food resources available to macroinvertebrates for primary consumption. Relations between periphyton biomass and macroinvertebrate abundance and taxa richness were tested using Pearson correlation (Zar 1984).
Macroinvertebrate assemblages were assessed using non-metric dimensional scaling analysis (NMDS) to examine differences among sites and over time. Data were log(x + 1) transformed prior to analysis. The chironomid data were expressed as relative abundances and arcsine square root transformed prior to running the NMDS. Permutational multivariate ANOVA (PERMANOVA) was used following each NMDS to test for differences among groups evident in the NMDS results. The NMDS analyses were conducted in R using the vegan package (R Core Team 2021). Macroinvertebrate data are also summarized with respect to taxa richness, abundance and Simpson’s evenness index, with the chironomids lumped as one taxon. The chironomids were lumped as one taxon in the full dataset because species-level identifications were not completed for some years, as noted above. Species-level chironomid data (encompassing the full 20-year period but excluding the missing dates) were also summarized using these metrics. Based on the NMDS groupings, the metrics above were tested for differences among basins (north, south), Immez outlet (site 9) and Zeznina (site 10) using ANOVA on log(x + 1)-transformed values followed by a Tukey’s post hoc test when significant (Zar 1984). Macroinvertebrate abundance and taxa richness were also summarized across time, but as no clear trends emerged, the data are shown in the Supplementary file.
Results
The PCA of the physico-chemistry of Macun surface waters clearly separated north basin sites from south basin sites, with intermediate scores for Immez outlet (site 9) and Zeznina (site 10) in the PCA scatterplot (Fig. 2). Axes 1 and 2 of the PCA explained a cumulative 52.4% of the variation among site groups. PCA-1 was best explained by values of dissolved nitrogen, dissolved organic carbon and total inorganic carbon, while PCA-2 was best explained by values of particulate phosphorus, total phosphorus and particulate organic carbon. PCA-1 clearly separated years 2001–2010 from years 2011–2020, indicating major changes in the measures explaining PCA-1. Notable was the strong decrease in nitrogen levels in surface waters over the study period (see Robinson et al. 2022). PCA-2 distinguished basin-level differences in phosphorus and particulate organic carbon, with higher values in the north basin than south basin. Also evident in the scatterplot is a general increase in the similarity of surface waters after 2010. Lastly, although associated with axis-3 of the PCA (explaining 13.5% of the variation), average temperature was 3 °C lower and average specific conductance was higher (10.3 versus 6.5 S/cm) in the south compared to the north basin (average turbidity was low among sites at 2–8 nephelometric turbidity units).

Scatterplot of a principal components analysis (PCA) of the physico-chemistry of the Macun streams over the 21 years of monitoring [graph modified from Robinson et al. (2022)]. Note the general homogenization of surface waters after 2010. Shown are means ± SE of site scores among basins and time periods, grouped after the analysis
Periphyton biomass was summarized by basin (north, south) and outlet sites 9 and 10 over the study period (Fig. 3). In general, periphyton biomass on stones was similar between north and south basin sites, with values at Immez outlet (site 9) and Zeznina (site 10) being less than those at the basin sites (F = 31.3, p < 0.001). Temporally, a decrease in biomass was noted from 2002 to 2004 followed by a substantial increase (2005–2009) and then a clear decrease after 2010 at all sites (F = 42.3, p < 0.001). Correlations (r-values) in periphyton biomass among groups ranged from a low of 0.42–0.59 (Zeznina versus other groups) to 0.82 (north versus south basin). Basin sites (north and south) usually had values that were over 10 g/m2 prior to 2010 but less than 10 g/m2 after 2010. Immez outlet showed a peak in biomass from 2005 to 2007 in concert with higher values at basin sites, but values were less than 10 g/m2 in most years. Zeznina had the lowest values of most of the study years (Tukey’s test, p < 0.05), being typically less than 5 g/m2 (Fig. 3).

Temporal changes in periphyton biomass (as ash-free dry mass) seen when sites are combined within basins over the monitoring period. Data represent means ± SE. Note the general decline in biomass after 2010. The graph has been modified from Pezsek et al. (2022)
The NMDS analysis of macroinvertebrates revealed significantly different assemblages among the grouped sites (north basin, south basin, Immez outlet, Zeznina) (Fig. 4; PERMANOVA, p < 0.05). North basin sites were situated on the right side of the plot along axis-1, whereas Immez outlet and Zeznina were located on the far left of the plot. South basin sites were intermediate and more central along axis-1 of the NMDS. Along NMDS axis-2, Zeznina was located near the top of the plot, whereas the other sites were situated lower on the plot (Fig. 4). The NMDS showed that Zeznina had higher abundances of Drusus melanchaetus, Dictyogenus sp., Rhithrogena loyolaea and Baetis alpinus than the other sites, whereas simuliids were most abundant at north basin sites, and stoneflies (Leuctra spp., Protonemura sp., Perlodes sp.), oligochaetes, Crenobia alpina and chironomids dominated south basin sites. Lastly, the NMDS results indicated no major temporal changes in assemblage structure over the study period at any of the sites.

Scatterplot of a non-metric dimensional scaling analysis (NMDS) of macroinvertebrates from study sites in the Macun Lakes region during the study period. Data represent means ± SE of sampling years for each site. Stress = 0.17 for the compiled data. ACRO Acrophylax zerberus, AGAB Agabus bipustulatus, BAET Baetis alpinus, CHIR Chironomidae, CREN Crenobia alpine, DICR Dicranota sp., DICT Dictyogenus sp., DINO Dinocras sp., DIVE Others, DRUS Drusus melanchaetus, EMPE Empedidae, HELO Helophorus glacialis, HYDR Hydracarina, HYFO Hydroporus foveolatus, HYNI Hydroporus nivalis, LEUC Leuctra spp., LIMN Limnephilus coenosus, LIPE Limnius perrisi, NEMO Nemoura sp., OLIG Oligochaeta, PERL Perlodes sp., PROS Prosimulium sp., PROT Protonemura sp., PSEU Pseudopsilopteryx zimmeri, RHIT Rhithrogena loyolaea, RHYP Rhypholophus sp., SIMU Simulium sp., THAU Thaumaleidae
The NMDS analysis of chironomids clearly differentiated north basin from south basin sites along axis-2 (Fig. 5; PERMANOVA, p < 0.05). Immez outlet (site 9) was grouped with north basin sites, whereas Zeznina (site 10) fell to the right of but with some overlap with north basin sites. Sites within the north or south basin tended to separate along NMDS axis-1. Here, upper sites in the south basin were situated more to the right of the plot than lower sites, suggesting a potential longitudinal shift in assemblage structure. A similar pattern was observed in the north basin with upper sites also situated more towards the right of the plot than lower sites. The most common chironomids in the south basin included Pseudodiamesa nivosa, Diamesa zernyi gr./cinerella gr., Pseudodiamesa sp., and Parorthocladius nudipennis. Common chironomids at Zeznina (site 10) included Tvetenia bavarica/calvescens and Diamesa latitarsis gr., whereas Corynoneura sp., Diamesa bertrami and Pseudokiefferiella parva dominated the chironomid assemblages at north basin sites (Fig. 5).

Scatterplot of an NMDS analysis of chironomid species from study sites in the Macun Lakes region during the study period. Data represent means ± SE of sampling years for each site. Stress = 0.21 for the compiled data. CORY Corynoneura sp., COSC Corynoneura scutellata gr., DIBE Diamesa bertrami, DILA Diamesa latitarsis gr., DIZE Diamesa zernyi, DIZC Diamesa zernyi gr./cinerella gr., EUMF Eukiefferiella minor/fittkaui, HEMA Heterotrissocladius marcidus, MIIN Micropsectra insignilobus-type, PAAU Paratanytarsus cf. austriacus, PANU Parorthocladius nudipennis, PAST Parametriocnemus stylatus, PSBR Pseudodiamesa branickii, PSEU Pseudodiamesa sp., PSNI Pseudodiamesa nivosa, PSPA Pseudokiefferiella parva, ORFR Orthocladius (M.) frigidus, ORJU Orthocladiinae (juveniles), RHEF Rheocricotopus effuses, TORE Tokunagaia rectangularis gr., TVBC Tvetenia bavarica/calvescens
Mean macroinvertebrate taxa richness was highest at Zeznina (site 10) when compared to that of north basin sites, south basin sites and Immez outlet (site 9) (Fig. 6a; Tukey’s test, p < 0.05). Mean macroinvertebrate abundance was highest at south basin sites when compared to that of the other sites (Fig. 6b; Tukey’s test, p < 0.05). Mean Simpson’s index was lowest at Zeznina when compared to that of the other sites (Fig. 6c; Tukey’s test, p < 0.05), reflecting the higher taxa richness at Zeznina. Temporal patterns in abundance and richness showed high inter-annual variation and no trends over the study period (Fig. S1). There was no correlation between periphyton biomass and macroinvertebrate abundance or taxa richness for any group (r-values ranged from −0.16 to 0.21), indicating that the lower periphyton values post-2010 at all sites had no relation with temporal patterns in macroinvertebrate assemblages.

Taxa richness, abundance and Simpson’s index for macroinvertebrates in the north basin (NB; sites 1–4 combined), south basin (SB; sites 5–8 combined), Immez outlet (IM; site 9) and Zeznina (ZE; site 10) averaged among years during the study period. a–c Data for all macroinvertebrates, d–f data for Chironomidae only. Bars represent means + SE
Thus far, a total of 74 chironomid species (Tanypodinae, Diamesinae, Orthocladiinae, and Tanytarsini) have been collected from surface waters of the Macun Lakes region, with three of these species new to science (see Supplementary Table 1). Chironomid richness was lowest at north basin sites when compared with that for the other sites (Fig. 6d; Tukey’s test, p < 0.05), but usually only by two or three taxa (data from kick-net sampling). Chironomid abundances were notably higher at south basin sites and Immez outlet (on average 185 individuals at both sites) than north basin sites and Zeznina (both on average 55 individuals) (Fig. 6e; Tukey’s test, p < 0.05). Lastly, Simpson’s index for chironomids was lowest for Zeznina (site 10) when compared with those of the other sites (Fig. 6f; Tukey’s test, p < 0.05).
Discussion
Alpine landscapes are being rapidly transformed in response to a warming climate, which is causing the reduction and loss of glaciers globally. The hydrology of alpine fluvial networks has been modified as a result, with alterations in flow regimes, water permanency and physico-chemistry (Brown et al. 2007; Milner et al. 2009; Paillex et al. 2020). The fluvial network of Macun encompasses a number of lakes along the longitudinal flow path, with significant lake-order effects on surface waters (Robinson and Oertli 2009). Further, the stream network comprises two distinct basins, one with significant inputs from rock glacier waters, which cause noticeable differences in physico-chemistry between the two basins. Over the study period, the inputs of rock glaciers diminished, with a strong shift in physico-chemistry being observed around 2010 [summarized in Robinson et al. (2022)]. The reduced rock glacier inputs have also caused homogenization of the physico-chemistry of surface waters in the Macun catchment. The question in this study was whether the temporal changes in physico-chemistry between Macun waters would be reflected in the macroinvertebrate assemblages in the catchment. A previous analysis of diatoms at the study sites showed a clear relationship between the changes in surface water physico-chemistry and changes in diatom assemblage structure (Peszek et al. 2022), especially with respect to pre- and post-2010 data.
Organic resources measured as periphyton biomass displayed basin and site differences associated with the differences in surface water physico-chemistry. For example, water temperatures of north basin sites were generally 3 °C higher than those of south basin sites (see Robinson et al. 2022). Further, site Zeznina had the lowest periphyton biomass values during the study period; this was likely related to the influx of waters from a large rock glacier that resulted in a different physico-chemical signature than that seen in the other basins and sites. The other notable observation was the decrease in periphyton biomass in both basins after 2010 to levels similar to those at site Zeznina, and also following the shift observed in surface water physico-chemistry in the catchment in general. For instance, the major reduction in nitrate levels in the surface waters is likely related to the decrease in periphyton biomass. The question relevant to this study was whether levels of periphyton, as a food resource, limited the abundance of macroinvertebrates. In this regard, no correlations were evident between periphyton biomass and macroinvertebrate abundances or taxa richness over the study period, from which we infer that abundances and richness were not influenced by periphyton levels at the study sites. Periphyton comprises mostly diatoms in alpine waters, and thus is a high-quality food resource for macroinvertebrates inhabiting alpine streams (Lamberti and Moore 1984; Jacobsen and Dangles 2017). The temporal changes in diatom assemblages in the catchment pre- and post-2010 apparently had no major influence on macroinvertebrate abundances.
Macroinvertebrate taxa richness was generally low in the catchment (less than eight taxa on average, with chironomids grouped together), but was notably higher at site Zeznina, at 11 taxa on average. Zeznina is the outflow stream of the catchment and is larger (comprising waters from both basins plus substantial rock glacier inputs) and more heterogeneous in habitat conditions than the other sites (CTR, CJ and BL-C, personal observations). In contrast, chironomid richness ranged from five to seven taxa on average among sites in the catchment, and was lowest in the north basin (mean = 5 taxa). Alpine streams are known to be inhabited by a lower number of taxa than lower elevation streams (Jacobsen and Dangles 2017), and the Macun streams fit this general observation. Also, Simpson’s index was lowest for site Zeznina (ca. 50% lower than for the other sites), suggesting that it had a more evenly distributed macroinvertebrate assemblage structure than the other sites. Macroinvertebrate abundances were highest in the south basin (150 more individuals collected per kick sample than at other sites), mostly due to large numbers of chironomids. Chironomid abundances were lowest in the north basin and Zeznina (30% of the abundances noted for the south basin and the Immez outlet). The assemblage evenness of chironomids was similar across sites (mean Simpson’s index = 0.4–0.5), and suggested multiple species were relatively abundant at the sites. As noted earlier, no obvious temporal trends were observed in the metrics above, indicating that finer resolution data (or a longer time series) may be needed to answer the study question.
The composition of the macroinvertebrate assemblages displayed basin and site differences. North basin site assemblages differed from those of south basin sites as well as those of the Immez outlet (site 9) and Zeznina (site 10). With respect to the NMDS, site Zeznina clearly differed from all the other sites (located in the upper left corner of the scatterplot), whereas Immez outlet differed from the other sites mainly along axis-1. Zeznina was clearly more species rich than the other sites; there, discharge was greater due to rock glacier inputs, and it was three-fold steeper (> 3° gradient, fast-flowing waters) than the other sites (< 1° gradient, slow-flowing waters), which made the system more physically dynamic. The data further indicate that the spatial differences in surface water physico-chemistry and associated organic resources (differences in primary producers between basins) likely play a role in the distribution and abundance of macroinvertebrates between basins and among sites. For example, particulate organic matter as well as total phosphorus differed between basins (higher in the north basin than south basin), suggesting a stronger glacial legacy effect in the south basin, where inputs of rock glaciers are still evident (see Del Siro et al. 2023). It is notable that filter-feeding simuliids (Prosimulium sp.) had high abundances in north basin streams but were essentially absent from south basin streams, reflecting the differences in particulate organic matter between these basins. The glacial signature in water physico-chemistry was also reflected in the differences in diatom assemblages between basins in the catchment (Peszek et al. 2022). Similarly, it is well known that macroinvertebrate assemblages differ among stream types in glacial fluvial networks (Hieber et al. 2005; Brown et al. 2007).
Macroinvertebrate assemblages differed spatially among sites in each basin, supporting a lake-order effect on streams, as suggested by Robinson et al. (2012). This spatial pattern was observed in macroinvertebrate assemblages overall, as well as for the chironomids in particular. The juxtaposition of lakes and streams in the Macun drainage network provides a functionally heterogeneous environment that clearly affects the biotic properties along fluvial flow paths. Lake-order effects, influencing flow and temperature regimes as well as nutrient dynamics of outflow streams, have been documented previously (Kling et al. 2000; Epstein et al. 2013). The large Macun lakes are functionally dynamic, with major seasonal and diel fluctuations in physico-chemistry and metabolism (Robinson et al. 2022) that extend to outlet streams along the fluvial network. However, rock glacial inputs still maintain the glacial chemical signature of streams in the south basin, with the main-stem stream being 3 °C cooler than the main-stem stream in the north basin, which clearly influences macroinvertebrate assemblages between basins. These data indicate a hierarchical habitat filtering process in the catchment with surface water physico-chemistry effects (basin differences) followed by lake-order effects (longitudinal patterns) on the spatial distribution of macroinvertebrates (and diatoms) (Robinson et al. 2012; Peszek et al. 2022).
The number of chironomid species has increased substantially over the period of study with the sampling of more types of freshwater habitats. Up until now, 68 morphospecies and six genera have been documented for the catchment. Sampling has included sweep-netting of different lakes (Oertli et al. 2008) and kick-netting of the 10 long-term stream sites (Lods-Crozet et al. 2012; Robinson et al. 2016) and tributaries (periodically over the years) (Ruegg and Robinson 2004), Malaise trapping of adults in some years, and 24-h drift sampling for midge exuviae at selected streams (e.g., rock glacier streams, main-stem channel in both basins) in the last 2 years. The current list includes three species new to science (Moubayed-Breil and Lods-Crozet 2018, 2021, 2023). Bryocolous and hygropetric habitats seem to represent the most favorable aquatic microhabitats for the larval populations. These new species, which belong to the subfamily Orthocladiinae, are members of a crenophilous community (Lindegaard 1995; Lencioni et al. 2000). Chironomids are known to be common in alpine aquatic habitats; for example, Lencioni et al. (2000) documented 33 morphospecies in alpine streams in northern Italy, and Füreder et al. (2006) noted over 60 chironomids among various high-elevation lakes scattered throughout the European Alps. Many alpine chironomids are endemic and have physiological adaptations (e.g., resistance to or tolerance of freezing) to the relatively harsh conditions found in high-elevation aquatic systems (e.g., pro-glacial streams) (Lencioni 2004). Our data indicate that chironomids have been understudied in alpine environments and that more effort should be put into describing their biodiversity, including the use of current methods involving environmental DNA and DNA sequencing of species. The presence of the three new species in this high alpine catchment highlights the importance of some cold glacial enclaves, considered as hotspots of endemism, in the preservation and persistence of autochthonous and sensitive alpine relic species. It is noteworthy that other high alpine environments in the northern and southern Alps are home to some other newly described species (Stur and Spies 2011; Rossaro et al. 2017; Moubayed-Breil and Lods-Crozet 2016, 2018, 2023). Such species are considered relevant from a biogeographical perspective, and their loss would be indicative of environmental change in alpine landscapes (e.g., Lencioni et al. 2021).
The endemism, taxonomic richness and abundance of chironomids in alpine aquatic habitats suggest they may be good sentinels of environmental change that is relative to macroinvertebrates in general. The NMDS results showed a clear separation between the chironomid assemblages in the north and south basin, as well as a clear longitudinal change within each basin. Importantly, these patterns were more distinct than those observed with the macroinvertebrate data, where chironomids were lumped together in the analysis; this indicates that higher resolution macroinvertebrate data provide a clearer picture of response in freshwater ecosystems. The most common species in the south basin included the chironomids Pseudodiamesa nivosa, Pseudodiamesa sp., Parorthocladius nudipennis and Diamesa zernyi gr./cinerella gr., all of which are found in cold alpine streams/lakes (Lods-Crozet et al. 2001). As discussed above, the south basin streams retain a glacial water signature and are on average 3 °C colder than north basin streams. Further, the larvae of Pseudodiamesa spp. are carnivores (Serra-Tosio 1976) in freshwaters, and perhaps fill a predatory niche in alpine streams (predatory stoneflies were present in low numbers at some sites). Corynoneura sp., Diamesa bertrami and Pseudokiefferiella parva dominated the chironomid assemblages at north basin sites. Diamesa spp. are well-known cold water specialists (Lencioni 2004), whereas Corynoneura sp. are typically found in ponds/lakes, and the slow-flowing waters of north basin streams likely allow for its presence. Further, the orthoclads, which are known to be found in warmer waters and usually at a longitudinal sequence downstream of the Diamesinae, were mainly found in north basin sites. The presence of chironomids Tvetenia bavarica/calvescens and Diamesa latitarsis at Zeznina further indicate that the rock glacier water inputs influence the physico-chemistry at this site. Lastly, no clear temporal trends were evident in the long-term records of chironomids for any sampling site, which supports the general idea of a lake mitigation effect on the fluvial network in Macun.
To summarize, alpine surface waters are sensitive to climate-related changes in environmental conditions, especially those associated with the ongoing glacial recession occurring globally. Our results showed significant differences in the physico-chemistry of waters between basins in the alpine cirque landscape of the Macun Lakes region that partially reflected the influence of rock glacier inputs in one basin. The physico-chemical data revealed temporal changes as well, especially when data from before and after 2010 were compared, as well as a general homogenization of surface waters over the study period. Periphyton biomass differed between basins prior to 2010, but decreased across sites post-2010 concomitant with the changes in physico-chemistry, which diminished biomass differences between basins. In contrast, macroinvertebrates (as well as chironomids, in particular) showed no temporal trends over the study period, although basin differences occurred and persisted over this period. These data suggest that the juxtaposition of lakes and running waters, and the influence of lakes on outlet streams, may buffer macroinvertebrate assemblages to subtle temporal changes in water physico-chemistry and food resources. Lakes in the Macun catchment are metabolically active (see Robinson et al. 2022) and likely play a role in the ecological response of interconnected streams to environmental changes, such as climate-related ones. We suggest that additional efforts should be made to compare temporal patterns in alpine fluvial networks with and without interconnected lakes to test this supposition.
Data Availability
All of the data are available from the senior author uponrequest.
References
Allan JD (2004) Landscapes and riverscapes: the influence of land use on stream ecosystems. Annu Rev Ecol Evol Syst 35:257–284
Barsch D (1969) Studien und Blockgletschern in Macun, Unterengadin. Z Geomorphol Ann Geomorphol 8:11–30
Brown LE, Hannah DM, Milner AM (2007) Vulnerability of alpine stream biodiversity to shrinking glaciers and snowpacks. Glob Change Biol 13:958–966
Brown LE, Khamis K, Wilkes M, Blaen P, Brittain JE, Carrivik JL, Fell S, Friberg N, Gislason GM, Hainie S, Hannah DM, James WHM, Lencioni V, Olafsson JS, Robinson CT, Saltveit SJ, Thompson C, Milner AM (2017) Functional diversity and community assembly of river invertebrates show globally consistent responses to decreasing glacier cover. Nature Ecol Evol. https://doi.org/10.1038/s41559-017-0426-x
Cauvy-Fraunié S, Andino P, Espinosa R, Calvez R, Jacobsen D, Dangles O (2016) Ecological responses to experimental glacier-runoff reduction in alpine rivers. Nature Comm 7:12025
Chanut PCM, Drost A, Siebers AR, Paillex A, Robinson CT (2022) Flow intermittency affects structural and functional properties of macroinvertebrate communities in alpine streams. Freshw Biol 68:212–228. https://doi.org/10.1111/fwb.14018
Del Siro C, Scapozza C, Perga ME, Lambiel C (2023) Investigating the origin of solutes in rock glacier springs in the Swiss Alps: a conceptual model. Front Earth Sci 11:1056305. https://doi.org/10.3389/feart.2023.1056305
Derungs D, Tischhauser M (2017) Blockgletscher Macun—Dritte Folgemessung. Bachelor thesis, Fachhochschule Nordwestschweiz Hochschule für Architektur Bau und Geomatik Institut Vermessung und Geoinformation, 61 p
Ekrem T (2004) Immature stages of European Tanytarsus species. I. The eminulus-, gregarius-, lugens- and mendax species groups (Diptera, Chironomidae). Mitt Mus Naturkd Berl 51:97–146
Epstein DM, Neilson BT, Goodman KJ, Stevens DK, Wurtsbaugh WA (2013) A modeling approach for assessing the effect of multiple alpine lakes in sequence on nutrient transport. Aquat Sci 75:199–212
Fehr M, Reich R (2015) Blockgletscher Macun—Zweite Folgemessung. Bachelor’'s thesis, Fachhochschule Nordwestschweiz Hochschule für Architektur Bau und Geomatik Institut Vermessung und Geoinformation, 8 p
Ferrington LC, Sæther OA (2011) A revision of the genera Pseudosmittia Edwards, 1932, Allocladius Kieffer, 1913, and Hydrosmittia gen. n. (Diptera: Chironomidae, Orthocladiinae). Zootaxa 2849:1–314
Finn DS, Räsänen K, Robinson CT (2010) Physical and biological changes to lengthening stream gradient following a decade of rapid glacial recession. Glob Change Biol 16:3314–3326
Füreder L, Schütz C, Burger R, Wallinger M (2000) Seasonal abundance and community structure of chironomidae in two contrasting high alpine streams. Verh Int Verein Theor Angew Limnol 27:1597–1601
Füreder L, Ettinger R, Boggero A, Thaler B, Thies H (2006) Macroinvertebrate diversity in alpine lakes: effects of altitude and catchment properties. Hydrobiologia 562:123–144. https://doi.org/10.1007/s10750-005-1808-7
Hieber M, Robinson CT, Uehlinger U, Ward JV (2005) A comparison of benthic macroinvertebrate assemblages among different types of alpine streams. Freshw Biol 50:2087–2100
Ilyashuk B, Ilyashuk E, Makarchenko EA, Heiri O (2010) Midges of the genus Pseudodiamesa Goetghebuer (Diptera, Chironomidae): current knowledge and palaeoecological perspective. J Paleolimnol 44:667–676
IPCC (2023) AR6 synthesis report climate change 2023: longer report. Intergovernmental Panel on Climate Change https://www.ipcc.ch/report/ar6/ syr/
Jacobsen D, Dangles D (2017) Ecology of high altitude waters. Oxford University Press, Oxford, p 284
Jacobsen D, Milner AM, Brown LE, Dangles D (2012) Biodiversity under threat in glacier-fed river systems. Nat Clim Chang 2:361
Jacobsen D, Cauvy-Fraunié S, Andino P, Espinosa R, Cueva D, Dangles D (2014) Variations in glacial runoff change longitudinal distribution patterns of macroinvertebrates in an Ecuadorian glacier-fed stream: implications for the effects of global warming? Freshw Biol 59:2038–2050
Kling GW, Kipphut GW, Miller MM, O’Brien JO (2000) Integration of lakes and streams in a landscape perspective: the importance of material processing on spatial patterns and temporal coherence. Freshw Biol 43:477–549
Lamberti GA, Moore JW (1984) Aquatic insects as primary consumers. In: Resh VH, Rosenberg DM (eds) The ecology of aquatic insects. Praeger, New York, pp 164–195
Langton PH, Bitusik P, Mitterova J (2013) A contribution towards a revision of West Palaearctic Procladius Skuse (Diptera: Chironomidae). Chironomus Newsl Chironomidae Res 26:41–44
Langton PH (1991) A key to pupal exuviae of West Palaearctic Chironomidae. Langton PH, 5 Kylebeg Avenue, Mountsandel, Cleraine, County Londonderry, Northern Ireland
Lehmann J (1972) Revision der europäischen Arten (Puppen und Imagines) der Gattung Eukiefferiella Thienemann (Diptera: Chironomidae). Beitr Ent 22:347–405
Lencioni V (2004) Survival strategies of freshwater insects in cold environments. J Limnol 63:45–55
Lencioni V, Rossaro B (2005) Microdistribution of chironomids (Diptera: Chironomidae) in Alpine streams: an auto-ecological perspective. Hydrobiologia 533:61–76
Lencioni V, Maiolini B, Rossaro B (2000) The kryal and rhithral chironomid community in the Carè Alto system (Italian Central-Eastern Alps). Verh Int Verein Limnol 27:1–5
Lencioni V, Marziali L, Rossaro B (2012) Chironomids as bioindicators of enviromental quality in mountain springs. Freshw Sci 31:525–541
Lencioni V, Jousson O, Guella G, Bernabo P (2015) Cold adaptive potential of chironomids overwintering in a glacial stream. Physiol Entomol 40:43–53
Lencioni V, Franceschini A, Paoli F, Debiasi D (2021) Structural and functional changes in the macroinvertebrate community in alpine stream networks fed by shrinking glaciers. Fundam App Limnol 194:237–258
Lindegaard C (1995) Chironomidae (Diptera) of European cold springs and factors influencing their distribution. J Kansas Entomol Soc 68:108–131
Lods-Crozet B, Lencioni V, Olafsson JS, Snook D, Velle G, Brittain JE, Castella E, Rossaro B (2001) Chironomid (Diptera: Chironomidae) communities in six European glacier-fed streams. Freshw Biol 46:1791–1809
Lods-Crozet B, Oertli B, Robinson CT (2012) Long-term patterns of chironomid assemblages in a high elevation stream/lake network (Switzerland)—implications for global change. Fauna Norv 31:71–85
Milner AM, Brown LE, Hannah DM (2009) Hydroecological response of river systems to shrinking glaciers. Hydrol Process 23:62–77. https://doi.org/10.1002/hyp.7197
Milner AM, Khamis K, Battin TJ, Brittain JE, Barrand NE, Füreder LC, Fraunié S, Gíslason GM, Jacobsen D, Hannah DM, Hodson AJ, Hood E, Lencioni V, Ólafsson JS, Robinson CT, Tranter M, Brown LE (2017) Glacier shrinkage driving global changes in downstream systems. Proc Natl Academy Sci 114:9770–9778
Montagna M, Urbanelli S, Rossaro B (2016) The species of the genus Diamesa (Diptera, Chironomidae) known to occur in the Italian Alps and Apennines. Zootaxa 4193:317–331. https://doi.org/10.11646/zootaxa.4193.2.7
Moubayed-Breil J, Lods-Crozet B (2016) Heleniella helvetica sp. n., a cold stenothermic species inhabiting the upper Rhône catchment in central Switzerland (Diptera, Chironomidae, Orthocladiinae). J Entomol Acaral Res 48:339–344
Moubayed-Breil J, Lods-Crozet B (2018) On the genus Chaetocladius s. str. Kieffer, 1911 from Switzerland with descriptions of five new relic species occurring in glacial alpine springs and streams (Diptera, Chironomidae). Alpine Entomol 2:15–34
Moubayed-Breil J, Lods-Crozet B (2021) Pseudorthocladius immezensis sp. n., a new relict species inhabiting the Macun high-alpine stream, Swiss Alps (Diptera, Chironomidae). J Chironomidae Res 34:13–20
Moubayed-Breil J, Lods-Crozet B (2023) Limnophyes knispelae sp. n. and L. sartorii sp. n., two new crenophilous species from the Swiss Alps (Chironomidae, Orthocladiinae). J Chironomidae Res 37:48–58
Oertli B, Indermuehle N, Angelibert S, Hinden H, Stoll A (2008) Macroinvertebrate assemblages in 25 high alpine ponds of the Swiss National park (Cirque of Macun) in relation to environmental variables. Hydrobiologia 597:29–41. https://doi.org/10.1007/s10750-007-9218-7
Paillex A, Siebers AR, Ebi C, Mesman J, Robinson CT (2020) High stream intermittency in an alpine fluvial network: Val Roseg, Switzerland. Limnol Oceanogr 65:557–568
Peszek L, Kawecka B, Robinson CT (2022) Long-term response of diatoms in high-elevation streams influenced by rock glaciers. Ecol Ind 144:109515. https://doi.org/10.1016/j.ecolind.2022.109515
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Robinson CT, Matthaei S (2007) Hydrological heterogeneity of an alpine stream/lake network in Switzerland. Hydrol Process 21:3146–3154
Robinson CT, Oertli B (2009) Long-term biomonitoring of alpine waters in the Swiss National Park. Eco Mont 1:23–34
Robinson CT, Hieber M, Wenzelides V, Lods-Crozet B (2007) Macroinvertebrate assemblages of a high elevation stream/lake network with an emphasis on the Chironomidae. Fundam Appl Limnol 169:25–36
Robinson CT, Thompson C, Lods-Crozet B, Alther R (2016) Chironomidae diversity in high elevation streams in the Swiss Alps. Fundam Appl Limnol 188:201–213
Robinson CT, Jolidon C, Consoli G, Bloem S, Ebi C (2022) Temporal dynamics in the physico-chemistry of a high-alpine stream network in the Swiss National park. Eco Mont 14:22–34
Robinson CT, Kawecka B, Spaak P (2012) Limnologie der Macun-Seen (Schweizerischer Nationalpark). In: Schanz F, Scheurer T, Steiner B (eds) Ergebnisse aus 70 Jahren Gewässerforschung im Schweizerischen Nationalpark. Nat Park Forsch, pp 71–80
Rossaro B, Magoga G, Montagna M (2017) Revision of the genus Chaetocladius Kieffer (Diptera, Chironomidae), 1st note: description of four new species from Italy. J Entomol Acarological Res 49:36–47
Rüegg J, Robinson CT (2004) Comparison of macroinvertebrate assemblages of permanent and temporary streams in an Alpine flood plain, Switzerland. Fundam Appl Limnol 161:489–510
Sæther OA (1990) A review of the genus Limnophyes Eaton from the Holarctic and Afrotropical regions (Diptera: Chironomidae, Orthocladiinae). Ent Scand Suppl 35:1–135
Sæther OA (1995) Metriocnemus van der Wulp: seven new species, revision of species, and new records (Diptera: Chironomidae). Ann Limnol 31:35–64
Schmid PE (1993) A key to the larval chironomidae and their instars from Austrian Danube region streams and rivers. Part 1. Diamesinae, Prodiamesinae and Orthocladiinae. Wasser Abwasser Suppl 3(93):1–513
Serra-Tosio B (1976) Chironomides des Alpes: l genre Pseudodiamesa (Diptera, Chironomidae). Trav Sci Parc Natl De La Vanoise 7:117–138
Stur E, Ekrem T (2006) A revision of West Palaearctic species of the Micropsectra atrofasciata species group (Diptera: Chironomidae). Zool J Linn Soc 146:165–225
Stur E, Spies M (2011) Description of Chaetocladius longivirgatus sp.n., with a review of C. suecicus (Kieffer) (Diptera, Chironomidae). Zootaxa 2762:37–48
Tachet H, Richoux P, Bournaud M, Usseglio-Polatera P (2010) Invertébrés d’eau douce: systématique, biologie, écologie, 2nd ed. CNRS Editions, Paris
Thienemann A (1936) Alpine chironomiden (Ergebnisse von Untersuchungen in der Gegend von Garmisch-Partenkirchen, Oberbayern). Arch Hydrobiol 30:167–262
Tockner K, Malard F, Burgherr P, Robinson CT, Uehlinger U, Zah R, Ward JV (1997) Physico–chemical characterization of channel types in a glacial floodplain ecosystem (Val Roseg, Switzerland). Arch Hydrobiol 140:433–463
Wiederholm T (1983) Chironomidae of the Holarctic region. keys and diagnoses. part 1. Larvae Entomologica Scandinavica Suppl 19:1–449
Wiederholm T (1986) Chironomidae of the Holarctic region. Keys and diagnoses. Part 2. Pupae Entomol Scand Suppl 28:1–482
Wiederholm T (1989) Chironomidae of the Holarctic region. Keys and diagnoses. Part 3. Adult males. Entomol Scand Suppl 34:1–532
Wilkes MA, Carrivick JL, Castella E, Ilg C, Cauvy-Fraunié S, Fell SC, Füreder L, Huss M, James W, Lencioni V, Robinson CT, Brown LE (2023) Glacier retreat reorganizes river habitats leaving refugia for alpine invertebrate biodiversity poorly protected. Nat Ecol Evol 7:841–851
Zar JH (1984) Biostatistical analysis, 2nd edn. Prentice Hall, Englewood Cliffs, NJ
Acknowledgements
The study began in 2001 after annexation of the Macun Lakes area to the Swiss National Park. The authors graciously thank the Swiss National Park for logistical support of the annual expeditions over the last 21 years. We owe a debt of gratitude to the chemical lab (Aua lab) at Eawag for analyzing the water samples each year. We thank S. Blaser and P. Rodriguez for field assistance and lab processing of samples. We are very grateful to J. Moubayed-Breil for the description of three chironomids new to science. We thank V. Lubini for the examination and identification of Trichoptera collected in Macun. All data are available on request from the senior author or the data repository of the Swiss National Park. Lastly, we thank two anonymous reviewers for constructive comments that improved this paper.
Funding
Open Access funding provided by Lib4RI – Library for the Research Institutes within the ETH Domain: Eawag, Empa, PSI & WSL.
Author information
Authors and Affiliations
Contributions
CTR conceived study, collected samples, summarized data and wrote the paper, CJ assisted in the field and processed samples, BL-C conceived the study, analyzed samples, summarized the data, and wrote the paper.
Corresponding author
Ethics declarations
Conflict of interest
There are no conflicts of interest regarding this paper.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Robinson, C.T., Jolidon, C. & Lods-Crozet, B. Long-term patterns of stream macroinvertebrates in an alpine cirque landscape. Aquat Sci 86, 91 (2024). https://doi.org/10.1007/s00027-024-01106-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00027-024-01106-6