
Abstract
Successful stand regeneration is one of the keystone elements of sustainable forest management. It ensures that ecosystems submitted to stand-replacing disturbances return to a forested state so that they can maintain the provision of wood fiber, biodiversity, carbon sequestration, and other ecosystem services. This chapter describes how plantation forestry, including tree breeding, and novel tools, such as genomic selection, can support the sustainable management of boreal forests in the face of climate change by, among other benefits, reducing management pressure on natural forests and favoring ecosystem restoration.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


1 Introduction
Natural disturbances, such as wildfire, insect outbreaks, and windthrow, and anthropogenic disturbances, such as harvesting, are common in the boreal biome (Brandt et al., 2013; Shorohova et al., 2009). These events modify stand structure and affect the availability of environmental resources. Canopy removal increases light levels in the understory, modifying the microenvironment, plant community, and tree regeneration. These changes can have cascading effects on the capacity of forests to sustain their provision of ecosystem services. Vegetation can rapidly colonize the disturbed areas and prevent regeneration of the desired tree species or forest composition. For example, in some black spruce (Picea mariana (Mill.) BSP) stands of eastern Canada, the effects of harvesting on light levels and water table depth can trigger the growth of bryophyte communities; this shift favors paludification (Fenton & Bergeron, 2006), which in turn reduces forest productivity (Leroy et al., 2016). Similarly, site encroachment by ericaceous species such as Kalmia angustifolia L. or Empetrum hermaphroditum Hagerup can lead to a significant decline in soil fertility and conifer growth rates on some forest sites (Mallik, 2003).
Because successful stand regeneration can mitigate these effects, it is one of the keystone elements of sustainable forest management. In Canada, for example, regeneration success is used to monitor changes in conditions relevant to sustainable forest management under the Montreal Process (NRC 2020). It is also mandatory in Norway, Finland, and Sweden.

Successful regeneration, both from natural propagules and plantation practices, ensures that ecosystems submitted to stand-replacing disturbances return to a forested state so that they can maintain wood fiber supply, biodiversity, carbon sequestration, wildlife habitat, spiritual values, social values, and other ecosystem services (Fig. 14.1).
Boreal ecosystems are dominated by conifer species having long-lived aerial seed banks, reproduction from vegetative growth (e.g., the formation of layers around trees), established dense understory seedling banks, or a combination of these characteristics (Thiffault et al., 2015). Depending on jurisdictions and years, natural regeneration can thus play a substantial role in forest renewal. For example, in Sweden, about 24% of the area harvested between 1999 and 2019 regenerated naturally, although showing a declining trend (SFA, 2020). In Canada, the average was about 44% between 2000 and 2020 (NRC, 2020). The remaining areas are regenerated with plantations, which can take various forms. They include intensively managed, even-aged forest areas planted with one or two species at a regular spacing. The primary objective of these regenerated areas is wood production (Fig. 14.2a, b), although there is also a consideration of environmental and societal values in most boreal jurisdictions. In contrast, less intensively managed plantations, resembling natural forests at stand maturity, have the main purpose of ecosystem restoration, the protection of soil and water values, and/or support of socioeconomic objectives (Fig. 14.2c). Globally, intensively managed and other planted forests cover about 291 million ha, an area that represents 7% of the world’s forests (FAO, 2020).

Examples of various plantations, showing a an intensively managed white spruce (Picea glauca (Moench) Voss.) plantation in eastern Canada, b a mature Norway spruce (Picea abies (L.) Karst.) plantation in Sweden, c an extensively managed white spruce plantation in eastern Canada containing natural balsam fir (Abies balsamea (L.) Mill.) regeneration, and d a Norway spruce plantation in Sweden that was submitted to cleaning with the retention of naturally regenerated Scots pine (Pinus sylvestris L.). Photo credits a, c Nelson Thiffault, b, d Karin Hjelm
The capability of forest plantations to fulfill their role relies on interdependent decisions and actions. In most forestry contexts in the boreal zone, this means selecting appropriate genotypes and seedling size, managing the soil and humus to create appropriate microsites, controlling competing vegetation, managing stand density, and, in some cases, increasing nutrient availability. Tree breeding, silviculture, and their interactions drive the production of ecosystem services from planted forests (e.g., Burdon et al., 2017). This chapter describes the actual and potential role that plantation silviculture, tree breeding, and novel tools such as genomic selection can play in supporting the sustainable management of boreal forests in the face of climate change. First, we summarize some of the fundamentals of plantation silviculture and show how various treatments support sustainable forest management objectives. Then, we explore the role of tree breeding and genomic tools in assisting forest management. Third, we provide examples illustrating the role plantation forestry plays in maintaining various ecosystem services from boreal stands in the context of global change. Finally, we identify some issues and challenges facing plantation forestry in the context of sustainable forest management.
2 Plantation Establishment and Silviculture
When a forest stand is harvested, the energy previously captured by the canopy now reaches the understory and the soil (Fig. 14.3); this exposure increases soil and air temperature and the evaporative demand of the air. For newly planted seedlings, a higher soil temperature can positively affect root growth and the uptake of water and nutrients and reduce the risk of frosts. On the other hand, the increased evaporative demand and higher air temperatures can increase the risk of drought. Although soil water availability increases when trees are removed and more precipitation reaches the ground, colonizing vegetation competes for water in the rooting zone. Because the root functioning of newly planted seedlings is often poor (Grossnickle, 2005), this may further increase the impact of drought. Moreover, rapid changes in temperature between day and night can cause frost damage. Many nutrients are removed by harvesting the standing trees, although changes in the energy balance can heighten some nutrient cycle processes such as nitrification (Jerabkova et al., 2011).

Some environmental factors influenced by harvesting or natural disturbances that determine the establishment success of newly planted seedlings in the boreal forest. These factors can be manipulated using silvicultural treatments, such as mechanical site preparation, vegetation management, and fertilization, so that plantations can support sustainable forest management objectives. Tree-breeding programs can also select genotypes best adapted to sustain specific environmental conditions
In this context, plantation success in supporting sustainable forest management objectives relies on the proper selection and use of stock type and silviculture treatments (Rubilar et al., 2018). These decisions ensure that the planted seedlings have access to sufficient environmental resources from the time of planting until maturity so that survival is maximized and growth rates meet the silviculture and management objectives.
In general, seedling stock types vary in the size of the shoot/above ground biomass and the morphology of their root system. The choice of stock type size is generally based on the competing environment in which the seedlings are to be outplanted, as initial seedling size influences their inherent growth and capacity to compete for environmental resources (Jobidon et al., 2003). Larger seedlings are generally preferred on sites dominated, or that have the potential to be dominated, by fast-growing, light-demanding species. Smaller seedlings are ideal for sites where competition for light is low, as generally found at high latitudes where competing species consist mainly of shrubs, mosses, and lichens (Bell et al., 2011).
Most harvesting treatments alter vegetation dynamics; species well adapted to the new environmental conditions establish rapidly, occupy the site, and compete with planted seedlings for resources (Fig. 14.3). Vegetation management aims to direct the evolution of the forest succession to achieve a range of management objectives. The use of mechanical, chemical, biological treatments, or a combination thereof, applied during the various stages of early stand development can improve planted tree growth, vigor, resistance to damage from insects, survival, nutrient status, crown length and width, and stand volume growth (Wiensczyk et al., 2011). In most cases, vegetation management treatments are carried out to increase the wood production of desired species; however, vegetation management enables achieving other objectives, such as restoring declining species or diversifying stand composition and structure.
Mechanical site preparation is used to improve site factors and reduce seedling stress following planting (Fig. 14.3), leading to positive effects on seedling survival and growth (Sikström et al., 2020). In boreal contexts, mechanical site preparation increases the availability of site resources by reducing competition from other species colonizing the regeneration area and improves factors such as microclimate, nutrient mineralization, soil temperature, and soil water availability (e.g., Johansson et al., 2013). Site-specific characteristics and the management context influence the intensity of mechanical site preparation treatments and the impact severity of these treatments on the forest floor and the soil (Löf et al., 2012). For example, the applied treatment can consist of disturbing (locally) the organic layers through the use of motor manual equipment, mounding to create elevated planting spots, disk trenching to create linear rows of furrows and berms, soil inverting to produce planting spots with the mineral soil lying above an inverted humus layer, harrowing to completely mix the organic layers and incorporate them into the underlying soil, or blading, which completely removes the organic layer over large areas of soil.
Stand density (the number of stems growing per unit of space) influences productivity at the tree and stand levels. The size of individual trees is largest at low density because trees are exposed to low levels of intraspecific competition. At higher tree densities, volume production at the stand level is maximized because site occupancy is optimized (Groot & Cortini, 2016). Density management thus offers the opportunity to manipulate resource allocation to best fit the sustainable forest management objectives being pursued. In plantations, stand density is managed at the establishment phase by prescribing the planting distance between the seedlings. Thinning or cleaning treatments can later be used, either at the precommercial or commercial stage of stand development, to maintain or reduce stand density and select crop trees (Fig. 14.2d; Pelletier & Pitt, 2008). Thinning operations reduce competition between crop trees; hence, they improve the growth of the remaining stems. Although increased volume and radial growth rates generally lead to decreased wood density (Jaakkola et al., 2005), these effects can be nonsignificant (Franceschini et al., 2018; Vincent et al., 2011). The pruning of dead or living branches can also be used to increase wood quality and value (Mäkinen et al., 2014).
The availability of soil nitrogen is one of the major growth-limiting factors in boreal forests (Tamm, 1991). Fertilizers can be applied at planting, in the later stages of stand rotation, or at several points in time to promote plantation growth and achieve sustainable forest management objectives. Using fertilizers at planting can promote the rapid establishment and high initial growth of trees; for example, positive effects of amendments have been documented when used in combination with site preparation (Thiffault & Jobidon, 2006) or with nutrient irrigation (Johansson et al., 2012). The fertilization of mature stands, for its part, is seen as one of the most economically important measures to increase wood production. By adding nitrogen to middle-aged or older stands, usually a few years after the last thinning, tree growth can be increased significantly (Jacobson & Pettersson, 2010).
3 Tree Breeding and Genomic Selection
Millions of seedlings are planted each year in the boreal forest, with more than 450 million and 350 million seedlings planted annually in Canada and Sweden, respectively. In most northern countries, seedling material is improved for growth, or at least comes from known origins, to ensure its quality and adaptation to specific planting environments. This practice enhances plantation success and timely restocking so that ecosystem services can be fulfilled as quickly as possible after harvesting (Fig. 14.1). The use of improved planting material also protects investments and guarantees a future fiber supply of sufficient quantity and quality (Jansson et al., 2017).
Tree-breeding programs for boreal conifers have been established in many countries in the northern hemisphere to deliver improved seedlings for reforestation purposes (see Mullin et al., 2011 for an extensive review). The first tree breeding efforts comparing the growth of seed sources from different geographic origins go back to the early twentieth century. Structured tree-breeding programs for many commercial spruce (Picea spp.) and pine (Pinus spp.) species were initiated in the 1950 and 1960s by systematically sampling the genetic base (Fig. 14.4a). Hence, seeds and grafts from plus trees (particularly well-growing trees in natural stands) were collected from the species’ full distribution and planted in common garden experiments. These provenance studies determined the genetic variation within species to identify the best-growing seed sources and to study the genetic response to the environment (e.g., Li et al., 1997; Rehfeldt et al., 1999). These studies also established the foundation for crossing the best-performing individuals, leading to the beginning of a breeding population (Fig. 14.5).

a A 50-year-old mature white spruce (Picea glauca) provenance trial established at the Petawawa Research Forest in Ontario, Canada; and b a 16-year-old genetic trial of controlled crosses of Norway spruce (P. abies), an introduced species to eastern Canada for which several breeding programs are maintained. Trees in a and b were pruned to facilitate access for recurrent measurements of growth and for easier wood quality assessments. Pruning is also common practice in plantations for increasing wood quality. This treatment leads to fewer and smaller knots and hence stronger wood from the first log. Photo credits a Isabelle Duchesne, b Patrick Lenz

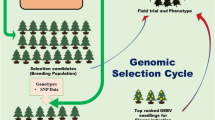
Schematic illustration of the tree-breeding process, in which plus trees are identified to set the groundwork for crossing the best-performing individuals and, hence, establish a breeding population. Genomic selection (GS) can shorten the breeding cycle by decades. The genetic gain of a desired parameter (e.g., height) is thus dependent on the genetic control and the selection differential s, which is the difference between the general mean µ and the mean of the selected subpopulation
Genetic trials of provenances or crosses follow distinct experimental designs that control within-site variation, determine genetic effects, and rank individual trees, families of crosses or their parents on the basis of their genetic merit (Fig. 14.4b). Measuring traits of interest in these experiments allows estimates of the different genetic parameters, e.g., their heritability, that determine the genetic gain expected through selection. For instance, height growth is typically between 20%–30% genetically controlled (e.g., Gamal El-Dien et al., 2015; Hong et al., 2014; Lenz et al., 2020a), whereas wood quality is under even stronger genetic control. In some cases, more than half of the observed variation in wood density and fiber dimension is attributed to genetics (e.g., Chen et al., 2014; Ivkovich et al., 2002). Adverse correlations between desired traits, such as growth and wood quality, require multitrait selection approaches to prevent wood quality degradation in planting stocks with enhanced growth (Hong et al., 2014; Lenz et al., 2020b).
Conventional tree breeding employs a recurrent cycle of evaluation, selection, and crossing of the best individuals, which are then re-evaluated (Fig. 14.5). Traditionally, selected individuals are multiplied and grafted into clone banks for next-generation crosses and into seed orchards for seed production (White et al., 2007). For economic reasons, most seeds used for mass seedling production originate from open-pollinated seed orchards where only the maternal genetic value is well controlled. Other multiplication methods rely on sowing seeds and planting seedlings from controlled crosses or growing seedlings into hedges to produce rooted cuttings. Seedlings from cuttings or emblings obtained through somatic embryogenesis are significantly more expensive than standard material (Chamberland et al., 2020). Nevertheless, these clonal reproduction methods allow for the full control of the genetic makeup and, thus, maximize genetic gain (Park et al., 2016).
Over the last decade, genomic selection has been tested in forest tree breeding, with several proof-of-concept studies being published for boreal conifers (e.g., Beaulieu et al., 2014; Gamal El-Dien et al., 2015). Genomic selection relies on linking the genomic marker profiles of trees to their phenotypes. Once the models are calibrated, predictions are made only on the basis of marker profiles, which can already be obtained at the seedling stage or from embryonic tissue; this avoids the imperative establishment of field tests (Park et al., 2016). Hence, the evaluation time is reduced to a minimum, and completing a selection cycle lasts only 10–15 years until improved seedlings are available (Lenz et al., 2020a) (Fig. 14.5). Genetic gain is enhanced further when vegetative reproduction is used for valuable genotypes. Genomic selection also facilitates the screening for expensive phenotypes, such as resistance, and quality traits in breeding populations. Models can be calibrated for a representative subset of a breeding program, and predictions can be made for other genotyped trees in the same population.
4 Benefits of Plantation Forestry in Sustainable Management
Plantations can provide high yields and offer the opportunity to select for species, genotypes, stand density, and spatial arrangements. They therefore play an important role in augmenting, maintaining, and restoring forest productivity in boreal landscapes and improving the provision of other ecosystem services (Freer-Smith et al., 2019).
4.1 Increasing Wood Production
Plantations increase wood production per unit of area relative to natural forests because they make better use of the space by the desired species, and they are based on genetically improved material. Overall, fiber production of desired quality can be tripled in a plantation compared with that obtained from unmanaged natural stands (Paquette & Messier, 2010). The amount of gain in each breeding cycle depends on the genetic control of growth traits and on the selection intensity. Despite the low to moderate heritability of growth, substantial gains have been achieved in one to two selection cycles in northern conifer breeding programs. For example, Isaac-Renton et al. (2020) reported almost 30% volume gain in first-generation top-crosses of Douglas fir (Pseudotsuga menziesii (Mirb.) Franco) in western Canada. For Norway spruce in Sweden, Liziniewicz et al. (2018) reported volume gains of nearly 30% in realized gain trials in second-cycle seed orchards. Multivarietal forestry offers volume gains greater than 50% by multiplying top-performing clones (Weng et al., 2008). At the landscape level, and when integrated within a functional zoning approach, the enhanced productivity of plantations can reduce the management pressure on natural forests without affecting wood production within the forest management unit (Messier et al., 2003). Enhanced growth also leads to shorter rotations, thus reducing the duration that trees are exposed to biotic and abiotic risk factors, including those related to climate change.
4.2 Adapting Forests to Future Conditions with Trait Selection and Assisted Gene Flow
In addition to growth and stem form, tree breeding can screen for other traits adapted to novel climate or market conditions. More frequent and severe drought events and late and early frosts in the more northern regions will negatively affect tree regeneration at boreal latitudes (Boucher et al., 2020). Plantation forestry, using adapted planting stock, can help maintain forest productivity in these challenging conditions. Breeding and modern genomic tools make it possible to accelerate the selection for various breeding goals (Fig. 14.5), including improved resilience, adaptation, and resistance to climate extremes and more frequent biotic stresses, or particular wood attributes to respond to emerging markets. In recent years, several studies have coupled dendroecology with genetics and genomics to forecast the adaptive potential of populations on the basis of past growth responses extracted from tree-ring data (e.g., Montwé et al., 2018). Moreover, new classes of phenotypes have improved our understanding of the genetic underpinning of adaptation (Housset et al., 2018). Growth resilience and recovery after drought stress are under significant genetic control and can thus be used as breeding criteria for adapting seedling material to future climate conditions; thus, this selection of seedling material can support sustainable forest management (Depardieu et al., 2020). Breeding also provides opportunities to enhance the resistance of conifers to biotic stressors, such as insects (e.g., Lenz et al., 2020b).
4.3 Restoring and Maintaining Natural Species, Closed-Forest Landscapes, and Ecosystem Functions
Although forest plantations can be established with the objective of producing a maximum of wood fiber in the shortest period, forest plantations in the boreal zone are frequently used to compensate for deficient natural regeneration and maintain a closed-crown forest cover. For example, in northeastern Canada, black spruce seedlings are planted alone or in combination with eastern larch (Larix laricina (Du Roi) K. Koch) or other species after wildfires to limit the expansion of lichen woodlands (e.g., Thiffault & Hébert, 2017). Plantations are also used as a tool to address biodiversity issues under the paradigm of ecosystem-based management (Paquette & Messier, 2010). For instance, plantation forestry is used to regenerate species that reproduce through serotinous cones and that rarely reproduce without wildfire (Bouchard, 2008). Forest plantations are also established to restore or maintain certain declining species through enrichment planting (e.g., Neves Silva et al., 2019). White spruce is an example of a species native to boreal eastern Canada that is sensitive to environmental conditions and suffers from needle chlorosis and defoliation, a phenomenon that can potentially be accentuated by climate change in regions characterized by low base cation availability in the soil (Ouimet et al., 2013). Furthermore, natural regeneration of white spruce is impeded by harvesting effects on stand structure. In this context, enrichment planting (Fig. 14.2c) increases the proportion of white spruce in the landscape to restore the historical forest composition (Delmaire et al., 2020). Plantations can also serve to restore wildlife habitats; for example, ecosystems subjected to heavy browsing pressure from large ungulates may experience regeneration failure of palatable tree species (Beguin et al., 2016).
4.4 Capturing and Storing Carbon and Supporting the Bioeconomy
Because of their high productivity, plantations are frequently identified as a means of sequestering atmospheric carbon for mitigating climate change (Waring et al., 2020). This practice is of particular interest in afforestation contexts, i.e., establishing trees on areas previously deprived of a forest cover (e.g., Ouimet et al., 2007). Plantations can, however, show lower net primary production than naturally regenerated forests, resulting in a lower carbon stock (Liao et al., 2010). In the boreal biome, the benefits of using plantations for carbon sequestration can thus be realized only under specific conditions. For example, assuming that the albedo effect is taken into account (Bernier et al., 2011), a positive carbon sink can be observed after the reforestation of forest heaths, which results from cascades of natural or anthropogenic disturbance (Gaboury et al., 2009). Moreover, although site preparation and the planting of boreal stands prone to paludification can result in losses of soil carbon, plantations on such sites should be beneficial because of the increased carbon storage in tree biomass (Lavoie et al., 2005). For example, increasing site preparation intensity on mineral soils can significantly increase carbon stock in the forest ecosystem in the long-term (Mjöfors et al., 2017). The net effect of mechanical site preparation on carbon stock remains, however, dependent upon the initial humus content and the site-specific soil characteristics.
5 Risks and Challenges
Forests play a critical role in addressing many of the largest global challenges. These challenges include mitigating climate change, conserving biodiversity, and providing a variety of ecosystem services, including nutrient cycling, air and water purification, carbon sequestration and storage, and wildlife habitat. Forests also have social and spiritual benefits and are key to important cultural activities. Although plantations can support the delivery of these services, they are also associated with silvicultural regimes that have the greatest potential for the artificialization of natural forests (Barrette et al., 2014).
Silvicultural treatments necessary for establishing successful plantations can have undesired effects on ecosystems and result in unforeseen impacts on the silvicultural regimes themselves. For example, although site preparation improves the establishment of planted seedlings, the increased area of disturbed soil from this treatment favors the establishment of naturally regenerated seedlings (e.g., Johansson et al., 2013). Whereas natural regeneration can complement or replace planted seedlings if mortality occurs, this regeneration can increase the need for precommercial thinning and other silviculture investments. There are also concerns that mechanical site preparation can negatively affect long-term productivity by depleting soil nutrients through rapid decomposition and leaching. Although tree growth appears to persist many decades after treatment (e.g., Hjelm et al., 2019), long-term research is necessary to fully evaluate the legacy effects of mechanical site preparation on site productivity. Similarly, fertilization is one of the most questioned silvicultural measures. Whereas it is effective at increasing tree growth and stemwood production, fertilization may adversely affect the forest ecosystem through, for example, negative impacts on soil-solution chemistry. Fertilization usually modifies plant composition relative to natural succession because of the increased availability of nutrients and reduced light penetration through a denser tree canopy (Hedwall et al., 2010); these effects may remain after felling and regeneration (Strengbom & Nordin, 2008) and could potentially impact sites at a millennial time scale (Dupouey et al., 2002).
The breeding process involved in producing successful plant material for plantation forestry also raises issues and challenges. For example, the extended time frame that breeders must foresee is one of the greatest challenges for decision-making and the selection of the optimal traits. Genomic selection can provide part of the solution. Nonetheless, 10 years of breeding cycle added to 20 years until commercial thinning of improved plantations remains a long time horizon during which environmental or market conditions may likely change with potentially significant social and economic impacts and the risk that the selected genomes will not be adapted to unforeseen changes in biotic and abiotic disturbances. Currently, genomic selection models are not transferable among breeding populations, as they largely trace pedigree and, to a minor extent, marker-trait associations (Lenz et al., 2017). There is hence a continued need for developing appropriate genotyping and statistical methods.
The social acceptability of plantation forestry is ambiguous (e.g., Wyatt et al., 2011). Although plantation forestry is perceived positively in certain circumstances, it is often associated with industrial practices, monocultures, the use of chemicals, a deterioration of water quality, negative effects on biodiversity, fragmentation of the forest matrix, and other landscape-scale impacts (Paquette & Messier, 2010). The use of improved planting material through breeding programs and genomic selection (Fig. 14.5) may wrongly be associated with the use of genetically modified organisms (GMOs).
6 Conclusions and Perspectives
Overall, the use of plantations in the sustainable forest management of the boreal forest undoubtedly raises significant issues related to the scale, localization, and spatial arrangement of plantations, the key attributes and resilience of natural forests, social acceptability, and the productivity and profitability of plantations, particularly in the context of ecosystem-based management (Barrette et al., 2014) (Fig. 14.6). Whereas the role of plantations in supporting sustainable forest management in the boreal forest is undeniable, their use should thus consider the risks associated with their implementation. These risks are increasing as biotic (e.g., native or exotic pests) and abiotic (e.g., drought) hazards expand because of global change, while concurrent economic and social pressures evolve constantly.

Adapted with permission from Barrette et al. (2014)
Some issues related to plantation silviculture in the context of ecosystem-based management in the boreal forest.
Plantation silviculture is compatible with and can support ecosystem-based management objectives (e.g., Barrette et al., 2019). When necessary, adaptive approaches can be applied at the stand level, such as establishing mixed- or multispecies plantations, maintaining biological legacies prior to establishing plantations, preserving patches of natural forest during site preparation, and favoring rare fruit-bearing tree species during cleaning treatments (Barrette et al., 2014). Maintaining or restoring the highest possible degree of naturalness within the forest matrix could address the complex issues associated with plantations at the landscape level (Tittler et al., 2012).
References
Barrette, M., Leblanc, M., Thiffault, N., et al. (2014). Issues and solutions for intensive plantation silviculture in a context of ecosystem management. The Forestry Chronicle, 90(6), 748–762. https://doi.org/10.5558/tfc2014-147.
Barrette, M., Thiffault, N., Tremblay, J. P., et al. (2019). Balsam fir stands of Northeastern North America are resilient to spruce plantation. Forest Ecology and Management, 450, 117504. https://doi.org/10.1016/j.foreco.2019.117504.
Beaulieu, J., Doerksen, T. K., MacKay, J., et al. (2014). Genomic selection accuracies within and between environments and small breeding groups in white spruce. BMC Genomics, 15(1), 1048. https://doi.org/10.1186/1471-2164-15-1048.
Beguin, J., Tremblay, J. P., Thiffault, N., et al. (2016). Management of forest regeneration in boreal and temperate deer–forest systems: Challenges, guidelines, and research gaps. Ecosphere, 7(10), e01488. https://doi.org/10.1002/ecs2.1488.
Bell, F. W., Thiffault, N., Szuba, K., et al. (2011). Synthesis of silviculture options, costs, and consequences of alternative vegetation management practices relevant to boreal and temperate conifer forests: Introduction. The Forestry Chronicle, 87(2), 155–160. https://doi.org/10.5558/tfc2011-005.
Bernier, P. Y., Desjardins, R. L., Karimi-Zindashty, Y., et al. (2011). Boreal lichen woodlands: A possible negative feedback to climate change in eastern North America. Agricultural and Forest Meteorology, 151(4), 521–528. https://doi.org/10.1016/j.agrformet.2010.12.013.
Bouchard, M. (2008). La sylviculture dans un contexte d’aménagement écosystémique en forêt boréale et en forêt mixte. In Gauthier S, Vaillancourt, M. A., Leduc, A., De Grandpré, L., Kneeshaw, D., Morin, H., Drapeau, P., & Bergeron, Y., (Eds.) Aménagement écosystémique en forêt boréale. Presses de l’Université du Québec (pp 335–359).
Boucher, D., Gauthier, S., Thiffault, N., et al. (2020). How climate change might affect tree regeneration following fire at northern latitudes: A review. New Forests, 51, 543–571. https://doi.org/10.1007/s11056-019-09745-6.
Brandt, J. P., Flannigan, M. D., Maynard, D. G., et al. (2013). An introduction to Canada’s boreal zone: Ecosystem processes, health, sustainability, and environmental issues. Environmental Reviews, 21(4), 207–226. https://doi.org/10.1139/er-2013-0040.
Burdon, R. D., Li, Y., Suontama, M., et al. (2017). Genotype × site × silviculture interactions in radiata pine: Knowledge, working hypotheses and pointers for research. New Zealand Journal of Forest Science, 47, 6. https://doi.org/10.1186/s40490-017-0087-1.
Chamberland, V., Robichaud, F., Perron, M., et al. (2020). Conventional versus genomic selection for white spruce improvement: A comparison of costs and benefits of plantations on Quebec public lands. Tree Genetics & Genomes, 16(1), 17. https://doi.org/10.1007/s11295-019-1409-7.
Chen, Z. Q., Gil, M. R. G., Karlsson, B., et al. (2014). Inheritance of growth and solid wood quality traits in a large Norway spruce population tested at two locations in southern Sweden. Tree Genetics & Genomes, 10, 1291–1303. https://doi.org/10.1007/s11295-014-0761-x.
Delmaire, M., Thiffault, N., Thiffault, E., et al. (2020). White spruce enrichment planting in boreal mixedwoods as influenced by localized site preparation: 11-year update. The Forestry Chronicle, 96(1), 27–35. https://doi.org/10.5558/tfc2020-005.
Depardieu, C., Girardin, M. P., Nadeau, S., et al. (2020). Adaptive genetic variation to drought in a widely distributed conifer suggests a potential for increasing forest resilience in a drying climate. New Phytologist, 227(2), 427–439. https://doi.org/10.1111/nph.16551.
Dupouey, J. L., Dambrine, E., Laffite, J. D., et al. (2002). Irreversible impact of past land use on forest soils and biodiversity. Ecology, 83(11), 2978–2984. https://doi.org/10.1890/0012-9658(2002)083[2978:Iioplu]2.0.Co;2.
Food and Agriculture Organization of the United Nations (FAO). (2020). The state of the world’s forests. Forests, biodiversity and people. Rome: Food and Agriculture Organization of the United Nations.
Fenton, N. J., & Bergeron, Y. (2006). Facilitative succession in a boreal bryophyte community driven by changes in available moisture and light. Journal of Vegetation Science, 17(1), 65–76. https://doi.org/10.1111/j.1654-1103.2006.tb02424.x.
Franceschini, T., Gauthray-Guyénet, V., Schneider, R., et al. (2018). Effect of thinning on the relationship between mean ring density and climate in black spruce (Picea mariana (Mill.) B.S.P.). Forestry, 91(3), 366–381. https://doi.org/10.1093/forestry/cpx040.
Freer-Smith, P., Muys, B., Bozzano, M., et al. (2019). Plantation forests in Europe: challenges and opportunities. From Science to Policy 9. (p. 52), Joensuu: European Forest Institute.
Gaboury, S., Boucher, J. F., Villeneuve, C., et al. (2009). Estimating the net carbon balance of boreal open woodland afforestation: A case-study in Québec’s closed-crown boreal forest. Forest Ecology and Management, 257(2), 483–494. https://doi.org/10.1016/j.foreco.2008.09.037.
Gamal El-Dien, O., Ratcliffe, B., Klápště, J., et al. (2015). Prediction accuracies for growth and wood attributes of interior spruce in space using genotyping-by-sequencing. BMC Genomics, 16(1), 370. https://doi.org/10.1186/s12864-015-1597-y.
Groot, A., & Cortini, F. (2016). Effects of initial planting density on tree and stand development of planted black spruce up to age 30. The Forestry Chronicle, 92(2), 200–210. https://doi.org/10.5558/tfc2016-039.
Grossnickle, S. C. (2005). Importance of root growth in overcoming planting stress. New Forests, 30(2–3), 273–294. https://doi.org/10.1007/s11056-004-8303-2.
Hedwall, P. O., Nordin, A., Brunet, J., et al. (2010). Compositional changes of forest-floor vegetation in young stands of Norway spruce as an effect of repeated fertilisation. Forest Ecology and Management, 259(12), 2418–2425. https://doi.org/10.1016/j.foreco.2010.03.018.
Hjelm, K., Nilsson, U., Johansson, U., et al. (2019). Effects of mechanical site preparation and slash removal on long-term productivity of conifer plantations in Sweden. Canadian Journal of Forest Research, 49(10), 1311–1319. https://doi.org/10.1139/cjfr-2019-0081.
Hong, Z., Fries, A., & Wu, H. X. (2014). High negative genetic correlations between growth traits and wood properties suggest incorporating multiple traits selection including economic weights for the future Scots pine breeding programs. Annals of Forest Science, 71, 463–472. https://doi.org/10.1007/s13595-014-0359-3.
Housset, J. M., Nadeau, S., Isabel, N., et al. (2018). Tree rings provide a new class of phenotypes for genetic associations that foster insights into adaptation of conifers to climate change. New Phytologist, 218(2), 630–645. https://doi.org/10.1111/nph.14968.
Isaac-Renton, M., Stoehr, M., Bealle Statland, C., et al. (2020). Tree breeding and silviculture: Douglas-fir volume gains with minimal wood quality loss under variable planting densities. Forest Ecology and Management, 465, 118094. https://doi.org/10.1016/j.foreco.2020.118094.
Ivkovich, M., Namkoong, G., & Koshy, M. (2002). Genetic variation in wood properties of interior spruce. II. Tracheid characteristics. Canadian Journal of Forest Research, 32(12), 2128–2139. https://doi.org/10.1139/x02-139.
Jaakkola, T., Mäkinen, H., & Saranpää, P. (2005). Wood density in Norway spruce: Changes with thinning intensity and tree age. Canadian Journal of Forest Research, 35(7), 1767–1778. https://doi.org/10.1139/x05-118.
Jacobson, S., & Pettersson, F. (2010). An assessment of different fertilization regimes in three boreal coniferous stands. Silva Fennica , 44(5), 815–827. https://doi.org/10.14214/sf.123.
Jansson, G., Hansen, J. K., Haapanen, M., et al. (2017). The genetic and economic gains from forest tree breeding programmes in Scandinavia and Finland. Scandinavian Journal of Forest Research, 32(4), 273–286. https://doi.org/10.1080/02827581.2016.1242770.
Jerabkova, L., Prescott, C. E., Titus, B. D., et al. (2011). A meta-analysis of the effects of clearcut and variable-retention harvesting on soil nitrogen fluxes in boreal and temperate forests. Canadian Journal of Forest Research, 41(9), 1852–1870. https://doi.org/10.1139/x11-087.
Jobidon, R., Roy, V., & Cyr, G. (2003). Net effect of competing vegetation on selected environmental conditions and performance of four spruce seedling stock sizes after eight years in Québec (Canada). Annals of Forest Science, 60(7), 691–699. https://doi.org/10.1051/forest:2003063.
Johansson, K., Nilsson, U., & Örlander, G. (2013). A comparison of long-term effects of scarification methods on the establishment of Norway spruce. Forestry, 86(1), 91–98. https://doi.org/10.1093/forestry/cps062.
Johansson, K., Langvall, O., & Bergh, J. (2012). Optimization of environmental factors affecting initial growth of Norway spruce seedlings. Silva Fennica, 46(1), 64. https://doi.org/10.14214/sf.64.
Lavoie, M., Paré, D., & Bergeron, Y. (2005). Impact of global change and forest management on carbon sequestration in northern forested peatlands. Environmental Reviews, 13(4), 199–240. https://doi.org/10.1139/A05-014.
Lenz, P. R. N., Beaulieu, J., Mansfield, S. D., et al. (2017). Factors affecting the accuracy of genomic selection for growth and wood quality traits in an advanced-breeding population of black spruce (Picea mariana). BMC Genomics, 18(1), 335. https://doi.org/10.1186/s12864-017-3715-5.
Lenz, P. R. N., Nadeau, S., Mottet, M. J., et al. (2019). Multi-trait genomic selection for weevil resistance, growth, and wood quality in Norway spruce. Evolutionary Applications, 13(1), 76–94. https://doi.org/10.1111/eva.12823.
Lenz, P. R. N., Nadeau, S., Azaiez, A., et al. (2020a). Genomic prediction for hastening and improving efficiency of forward selection in conifer polycross mating designs: An example from white spruce. Heredity, 124(4), 562–578. https://doi.org/10.1038/s41437-019-0290-3.
Lenz, P. R. N., Nadeau, S., Mottet, M.-J., et al. (2020b). Multi-trait genomic selection for weevil resistance, growth, and wood quality in Norway spruce. Evolutionary Applications, 13(1), 76–94. https://doi.org/10.1111/eva.12823.
Leroy, C., Leduc, A., Thiffault, N., et al. (2016). Forest productivity after careful logging and fire in black spruce stands of the Canadian Clay Belt. Canadian Journal of Forest Research, 46(6), 783–793. https://doi.org/10.1139/cjfr-2015-0484.
Li, P., Beaulieu, J., & Bousquet, J. (1997). Genetic structure and patterns of genetic variation among populations in eastern white spruce (Picea glauca). Canadian Journal of Forest Research, 27(2), 189–198. https://doi.org/10.1139/x96-159.
Liao, C., Luo, Y., Fang, C., et al. (2010). Ecosystem carbon stock influenced by plantation practice: Implications for planting forests as a measure of climate change mitigation. PLoS ONE, 5, e10867. https://doi.org/10.1371/journal.pone.0010867.
Liziniewicz, M., Berlin, M., & Karlsson, B. (2018). Early assessments are reliable indicators for future volume production in Norway spruce (Picea abies L. Karst) genetic field trials. Forest Ecology and Management, 411, 75–81. https://doi.org/10.1016/j.foreco.2018.01.015.
Löf, M., Dey, D. C., Navarro, R. M., et al. (2012). Mechanical site preparation for forest restoration. New Forests, 43(5–6), 825–848. https://doi.org/10.1007/s11056-012-9332-x.
Mäkinen, H., Verkasalo, E., & Tuimala, A. (2014). Effects of pruning in Norway spruce on tree growth and grading of sawn boards in Finland. Forestry, 87(3), 417–424. https://doi.org/10.1093/forestry/cpt062.
Mallik, A. U. (2003). Conifer regeneration problems in boreal and temperate forests with ericaceous understory: Role of disturbance, seedbed limitation, and keystone species change. Critical Reviews in Plant Sciences, 22, 341–366. https://doi.org/10.1080/713610860.
Messier, C., Bigué, B., & Bernier, L. (2003). Using fast-growing plantations to promote forest ecosystem protection in Canada. Unasylva, 54(214/215), 59–63.
Mjöfors, K., Strömgren, M., Nohrstedt, H. Ö., et al. (2017). Indications that site preparation increases forest ecosystem carbon stocks in the long term. Scandinavian Journal of Forest Research, 32(8), 717–725. https://doi.org/10.1080/02827581.2017.1293152.
Montwé, D., Isaac-Renton, M., Hamann, A., et al. (2018). Cold adaptation recorded in tree rings highlights risks associated with climate change and assisted migration. Nature Communications, 9(1), 1574. https://doi.org/10.1038/s41467-018-04039-5.
Mullin, T. J., Andersson, B., Bastien, J., et al. (2011). Economic importance, breeding objectives and achievements. In C. Plomion, J. Bousquet, & C. Kole (Eds.), Genetics, genomics and breeding of conifers (pp. 40–127). Edenbridge Science Publishers & CRC Press.
Neves Silva, L., Freer-Smith, P., & Madsen, P. (2019). Production, restoration, mitigation: A new generation of plantations. New Forests, 50, 153–168. https://doi.org/10.1007/s11056-018-9644-6.
Nijnik, M., Slee, B., & Nijnik, A. (2014). Biomass production: Impacts on other ecosystem services. In Pelkonen, P., Mustonen, M., Asikainen, A., Egnell, G., Kant, P., Leduc, S., & Pettenella, D. (Eds.) Forest bioenergy for Europe, what science can tell us (Vol. 4, pp. 82–89). Joensuu: European Forest Institute.
Natural Resources Canada (NRC). (2020). The state of Canada’s forests. Annual report 2019. (p. 80), Ottawa: Natural Resources Canada, Canadian Forest Service.
Ouimet, R., Tremblay, S., Périé, C., et al. (2007). Ecosystem carbon accumulation following fallow farmland afforestation with red pine in southern Quebec. Canadian Journal of Forest Research, 37(6), 1118–1133. https://doi.org/10.1139/x06-297.
Ouimet, R., Moore, J. D., Duchesne, L., et al. (2013). Etiology of a recent white spruce decline: Role of potassium deficiency, past disturbances, and climate change. Canadian Journal of Forest Research, 43(1), 66–77. https://doi.org/10.1139/cjfr-2012-0344.
Paquette, A., & Messier, C. (2010). The role of plantations in managing the world’s forests in the Anthropocene. Frontiers in Ecology and the Environment, 8(1), 27–34. https://doi.org/10.1890/080116.
Park, Y. S., Beaulieu, J., & Bousquet, J. (2016). Multi-varietal forestry integrating genomic selection and somatic embryogenesis. In Park, Y. S., Bonga, J. M., & Moon, H. K. (Eds.) Vegetative propagation of forest trees (pp. 302–322). National Institute of Forest Science, Seoul.
Pelletier, G., & Pitt, D. G. (2008). Silvicultural responses of two spruce plantations to midrotation commercial thinning in New Brunswick. Canadian Journal of Forest Research, 38(4), 851–867. https://doi.org/10.1139/x07-173.
Rehfeldt, G. E., Ying, C. C., Spittlehouse, D. L., et al. (1999). Genetic responses to climate in Pinus contorta: Niche breadth, climate change, and reforestation. Ecological Monographs, 69(3), 375–407. https://doi.org/10.1890/0012-9615(1999)069[0375:GRTCIP]2.0.CO;2.
Rubilar, R. A., Lee Allen, H., Fox, T. R., et al. (2018). Advances in silviculture of intensively managed plantations. Current Forestry Reports, 4(1), 23–34. https://doi.org/10.1007/s40725-018-0072-9.
Shorohova, E., Kuuluvainen, T., Kangur, A., et al. (2009). Natural stand structures, disturbance regimes and successional dynamics in the Eurasian boreal forests: A review with special reference to Russian studies. Annals of Forest Science, 66(2), 201. https://doi.org/10.1051/forest/2008083.
Sikström, U., Hjelm, K., Holt Hanssen, K., et al. (2020). Influence of mechanical site preparation on regeneration success of planted conifers in clearcuts in Fennoscandia – a review. Silva Fennica, 54(2), 10172. https://doi.org/10.14214/sf.10172.
Strengbom, J., & Nordin, A. (2008). Commercial forest fertilization causes long-term residual effects in ground vegetation of boreal forests. Forest Ecology and Management, 256(12), 2175–2181. https://doi.org/10.1016/j.foreco.2008.08.009.
Swedish Forest Agency (SFA). (2020). Quality of the regrowth. Jönköping: Swedish Forest Agency.
Tamm, C. O. (1991). Nitrogen in terrestrial ecosystems. In Questions of productivity, vegetational changes, and ecosystem stability. New York: Springer.
Thiffault, N., & Hébert, F. (2017). Mechanical site preparation and nurse-plant facilitation for the restoration of subarctic forest ecosystems. Canadian Journal of Forest Research, 47(7), 926–934. https://doi.org/10.1139/cjfr-2016-0448.
Thiffault, N., & Jobidon, R. (2006). How to shift unproductive Kalmia angustifolia - Rhododendron groenlandicum heath to productive conifer plantation. Canadian Journal of Forest Research, 36(10), 2364–2376. https://doi.org/10.1139/x06-090.
Thiffault, N., Coll, L., & Jacobs, D. F. (2015). Natural regeneration after harvesting. In Peh, K. S. -H., R. T., Corlett, & Y. Bergeron (Eds.). Routledge handbook of forest ecology, (pp. 371–384). Routledge.
Tittler, R., Messier, C., & Fall, A. (2012). Concentrating anthropogenic disturbance to balance ecological and economic values: Applications to forest management. Ecological Applications, 22(4), 1268–1277. https://doi.org/10.1890/11-1680.1.
Vincent, M., Krause, C., & Koubaa, A. (2011). Variation in black spruce (Picea mariana (Mill.) BSP) wood quality after thinning. Annals of Forest Science, 68(6), 1115–1125. https://doi.org/10.1007/s13595-011-0127-6.
Waring, B., Neumann, M., Prentice, I. C., et al. (2020). Forests and decarbonization – roles of natural and planted forests. Frontiers in Forests and Global Change, 3, 58. https://doi.org/10.3389/ffgc.2020.00058.
Weng, Y., Park, Y., Krasowski, M., et al. (2008). Partitioning of genetic variance and selection efficiency for alternative vegetative deployment strategies for white spruce in Eastern Canada. Tree Genetics & Genomes, 4(4), 809–819. https://doi.org/10.1007/s11295-008-0154-0.
White, T. L., Neale, D. B., & Adams, W. T. (2007). Forest genetics. CABI Publishing.
Wiensczyk, A., Swift, K., Morneault, A. E., et al. (2011). An overview of the efficacy of vegetation management alternatives for conifer regeneration in boreal forests. The Forestry Chronicle, 87(2), 175–200. https://doi.org/10.5558/tfc2011-007.
Wyatt, S., Rousseau, M. H., Nadeau, S., et al. (2011). Social concerns, risk and the acceptability of forest vegetation management alternatives: Insights for managers. The Forestry Chronicle, 87(2), 274–289. https://doi.org/10.5558/tfc2011-014.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Thiffault, N., Lenz, P.R.N., Hjelm, K. (2023). Plantation Forestry, Tree Breeding, and Novel Tools to Support the Sustainable Management of Boreal Forests. In: Girona, M.M., Morin, H., Gauthier, S., Bergeron, Y. (eds) Boreal Forests in the Face of Climate Change. Advances in Global Change Research, vol 74. Springer, Cham. https://doi.org/10.1007/978-3-031-15988-6_14
Download citation
DOI: https://doi.org/10.1007/978-3-031-15988-6_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-15987-9
Online ISBN: 978-3-031-15988-6
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)