
Abstract
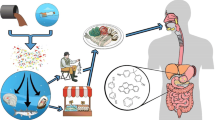
Seafood is an important food source, and this chapter addresses the food safety concerns related to plastic particles in different seafood. Here we focus on those species which are commonly consumed by humans, such as bivalves, gastropods, cephalopods, echinoderms, crustaceans, and finfish. The objectives of this chapter are to (1) outline the major sources, fate, and transport dynamics of microplastics in marine ecosystems, (2) provide a critical assessment and synthesis of microplastics in seafood taxa commonly consumed by humans, (3) discuss the implications of microplastics with regard to human health risk assessments, and (4) suggest future research priorities and recommendations for assessing microplastics in marine ecosystems in the context of global food security and ocean and human health.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
5.1 Introduction
Seafood is an important food source – with fisheries and aquaculture production predicted to increase by about 17.5% from 171 million tonnes in 2016 to approximately 201 million tonnes in 2030 (FAO 2018). It is a necessity that these marine-based foods are carefully managed and are safe for human consumption. Food security is defined by the Food and Agriculture Organization (FAO) as “a situation that exists when all people, at all times, have physical, social and economic access to sufficient, safe and nutritious food that meets their dietary needs and food preferences for an active and healthy life” (FAO 2017). In this chapter, the food safety concerns related to plastic particles in seafood will be addressed.
Global annual production of plastics is estimated to be approximately 300 million tonnes (Galloway 2015) and is still increasing steadily. Most plastic polymers are resistant to complete degradation and pose a potential risk to both human and environmental health. Of particular concern are microplastics, which are defined as particles <5 mm (GESAMP 2019) and which are the focus of this chapter.
Microplastics occur in different shapes and sizes and are formed from different polymers as well as additives, which reflects the diversity of sources and emissions to the environment (Rochman et al. 2019). The dominant microplastic polymers which are detected in the marine environment include polyethylene (PE), polypropylene (PP), polystyrene (PS), polyethylene terephthalate (PET), polyamide, and polyvinylchloride (PVC) (Hantoro et al. 2019). Primary microplastics are manufactured intentionally for a range of commercial uses (e.g., microbeads) whereas secondary microplastics originate from parent material such as textiles and discarded plastic items and are either generated through the use of plastic products or fragmentation following their loss to different environmental compartments. Plastic debris can enter the ocean from ships and fishing gear, as well as from atmospheric deposition, river transport, stormwater, sewage effluents, etc. (Browne et al. 2011; Napper and Thompson 2016; Lebreton et al. 2017; Allen et al. 2019). Plastics and microplastics have been identified in the oceans, from coastal zones to offshore areas, such as oceanic gyres (Eriksen et al. 2014; Jiang et al. 2020;), as well as in remote areas including the Arctic (e.g., Cózar et al. 2017). The ubiquitous nature of plastics in the ocean is an obvious concern for marine ecosystems and their inhabitants. In particular, the progression from macro- to microplastics at sea is a result of physical erosion and UV action and increases the bioavailability of smaller-sized particles to a wide array of marine organisms (Browne et al. 2008; Wright et al. 2013). Plastics have long been reported associated with marine organisms, from the first study of plastic ingestion by fish (Carpenter et al. 1972) to mariculture sites where boring worms facilitate the generation of microplastics from polystyrene buoys (Jang et al. 2018). Many investigations have been conducted to further understand the interaction between marine organisms and microplastics with several studies focusing on microplastic uptake, ingestion, exposure , and metabolic dynamics (Roch et al. 2020). Fibers are routinely identified as the most common microplastic type reported in fish, accounting for 58–87% of the plastic morphologies observed (Walkinshaw et al. 2020). Fragments, films, and fibers are also frequently found in fish and shellfish while microplastics in the forms of spheres are less common. The physical impacts of microplastic ingestion on marine organisms can include oxidative stress, inflammation, and potentially starvation, while less is known regarding the chemical effects of ingestion. The bioavailability and potential toxicity of microplastics are size dependent, with smaller particles able to penetrate further into an organism (Browne et al. 2008), with the potential of the release of associated co-contaminants (Bakir et al. 2016; Batel et al. 2016). It is widely accepted that ingestion is the main route of microplastics uptake in marine biota; however, it has recently also been demonstrated that surface scavenging appears to be an alternative route, as demonstrated in mussels (Kolandhasamy et al. 2018). Microplastics can also be taken up through respiration via the gills (Watts et al. 2016; Franzellitti et al. 2019) and have additionally been demonstrated to be maternally transferred to eggs in zebra fish (Pitt et al. 2018).
Plastics in aquatic environments have been shown to affect an organism’s health (such as behavioral changes and reduced growth rates); however, there is limited information on the effects of microplastics in seafood on human health. Lusher et al. (2017a) reported that more than 220 species of marine organisms including zooplankton, bivalves, crustaceans, fish, marine mammals, sea turtles, and seabirds had been shown to have ingested plastics, and more recently this number of species has increased to 690 (Carbery et al. 2018). Here we focus on those species which are commonly consumed by humans, such as bivalves, echinoderms, gastropods, cephalopods, crustaceans, and finfish. The specific objectives of this chapter are to (1) outline the major sources, fate, and transport dynamics of microplastics in marine ecosystems, (2) provide a critical assessment and synthesis of microplastics in seafood taxa commonly consumed by humans, (3) discuss the implications of microplastics with regard to human health risk assessments, and (4) suggest future research priorities and recommendations for assessing microplastics in marine ecosystems in the context of global food security and ocean and human health.
5.2 Fate and Transport of Microplastics in Marine Ecosystems.
The fate and transport of microplastics in the context of physical and biological oceanography has recently been reviewed by van Sebille et al. (2020), as well as by Thushari and Senevirathna (2020) with older reviews and critical papers developed by Andrady (2011), Wright et al. (2013), Galloway et al. (2017), and Wieczorek et al. (2019). The ocean can be both a source and sink for microplastics (Allen et al. 2020), and important themes within the cycling and degradation of microplastic particles (Weinstein et al. 2016) include the importance of transport from land via rivers (Lebreton et al. 2017; Hurley et al. 2018), the role of seafloor ocean circulation patterns as a driver of microplastic hotspots (Kane et al. 2020), and the concept of marine snow which has been identified as an important mechanism for transporting microplastic particles from the water column to the sediment (Porter et al. 2018). Moreover, fishing gear and other sources of macroplastics can degrade into microplastics via biological, chemical , and physical processes (Davidson 2012). Although settling of microplastic particles to the ocean floor is well-documented, recent research has shown that episodic events such as flooding (Hurley et al. 2018) and typhoons (Wang et al. 2019) are important drivers regarding the distribution and abundance of microplastics in coastal marine ecosystems.
5.3 Microplastic in Bivalves
Bivalves are by far the most investigated seafood species (Smith et al. 2018; Walkinshaw et al. 2020). Much of the investigations were performed for the purpose of uptake of microplastics from the environment, as filtering puts bivalves at an increased risk of microplastic intake from the water column (Li et al. 2019). Early investigations focused on blue mussels (Mytilus spp.), with wild and market brought samples presenting contamination levels of up to 7.2 microplastics per gram (Abidli et al. 2019; Bråte et al. 2018; Cho et al. 2019; De Witte et al. 2014; Li et al. 2015, 2016; Renzi et al. 2018a; van Cauwenberghe and Janssen 2014; van Cauwenberghe et al. 2015; Vandermeersch et al. 2015). Other bivalve species which have been investigated for microplastic uptake include clams (Venerupis philippinarum), oysters (Crassostrea gigas), and scallops (Patinopecten yessoensis) (Abidli et al. 2019; Cho et al. 2019, 2020; Davidson and Dudas 2016; Li et al. 2015; Rochman et al. 2015; van Cauwenberghe and Janssen 2014).
Microplastic fibers are often the most dominant morphology reported in bivalves. For example, fibers accounted for 80% of microplastics in mussels (Mytilus edulis, Perna viridis) from China (Qu et al. 2018), 90% of microplastics in Manila clams (V. philippinarum) from British Columbia (Davidson and Dudas 2016), and 99% in razor clams (Siliqua patula) and Pacific oyster (Crassostrea gigas) from Oregon, USA (Baechler et al. 2020a). One of the hypotheses behind the observed high abundance of this type of microplastic is that fibers are likely harder to remove from digestive tracts. Ward et al. (2019) reported that larger spheres are rejected at higher numbers (98%) than smaller spheres (10–30%). Fragments were most common in blue mussels and Pacific oyster from the French Atlantic coast (Phuong et al. 2018) as well as those from Korea, where EPS fragments likely originated from the high abundance of aquaculture facilities in the region (Cho et al. 2020). De Witte et al. (2014) reported that there was a high prevalence of fibers in blue mussels collected from quaysides related to fishing activities.
Microplastics in bivalves are likely dependent on several factors including, but not limited to, culture conditions and contamination levels in the environment, depuration procedures, filtration capabilities, as well as the tissues targeted for investigation. Some investigations, in distinct parts of the world, have found that bivalves sampled from highly contaminated areas or within the vicinity of urban sources of microplastics contained higher numbers of microplastics (Bråte et al. 2018; Qu et al. 2018; Cho et al. 2020). However, conversely, some investigations have reported no difference in microplastic exposure in bivalves related to sources (Covernton et al. 2019; Phuong et al. 2018).
There have been some reported differences between the occurrence of microplastics in market purchased (80%) and wild-caught individual bivalves (40%) (Ding et al. 2018). Similarly, farmed mussels displayed higher concentrations of microplastics than wild mussels (75 items and 34 items per mussel, respectively) (Mathalon and Hill 2014), although no difference was observed for wild and cultured Manila clams (Venerupis philippinarum) in British Columbia (Davidson and Dudas 2016). The use of depurations procedures appears to reduce the number of microplastics identified in bivalve species (van Cauwenberghe and Janssen 2014 – Mytilus edulis and Crassostrea gigas; Birnstiel et al. 2019 – Perna perna), which would hold significance in the preparation of mussels for consumption. The seasonality of sampling could also play a role in observed microplastic concentrations in marine biota. A significant seasonal variation was observed during summer for oyster samples which contained more microplastics; however, this trend was not detected for razor clams (Baechler et al. 2020c).
Particle selection by bivalves, related to size and morphology, will influence which particles are internalized both pre- and post-ingestion (Ward et al. 2018). Gut retention times, which are known to vary between bivalve species and the age of individuals, have shown, in general, that as particle size decreases, accumulation increases (Browne et al. 2008; Ward and Kach 2009; Ward et al. 2019). Much of the work performed on bivalves is based on the sampling and processing of whole organisms, with no differentiation between and among tissue types; this makes it impossible to determine whether microplastics were internalized by individuals, had migrated from gills and guts to visceral tissue, or were in the process of being egested (e.g., as pseudofeces). Kolandhasamy et al. (2018) reported that microplastic fibers can accumulate on the foot and mantle of blue mussels.
Consequences of microplastic intake/uptake by bivalves indicate that microplastics can directly affect bivalve physiology but also indirectly change the structure of their habitats, impairing food resources and facilitate the efficient transfer of organic pollutants (Zhang et al. 2019a). Other observed implications include negative effects on filtration activity (Green et al. 2019; Xu et al. 2017), feeding behavior (Wegner et al. 2012), and reproduction (Sussarellu et al. 2016; Gardon et al. 2018). It is important to highlight that effects are mostly studied using uniform particles, mostly spheres so these may not be truly representative of environmentally relevant microplastic exposure regimes (see Gomes et al. 2021, Chap. 7, this volume).
5.4 Microplastics in Echinoderms
Sea urchins and sea cucumbers are the main echinoderms consumed as food item, and few studies have been conducted on the abundance of microplastics in these marine organisms. Of the heart urchins (Brissopsis lyrifera) analyzed, 40% were found to contain microplastics in their soft tissue, primarily in the form of flakes (90%, the remaining 10% as fibers). In most cases the number of particles present was 1/individual (Bour et al. 2018). It is noteworthy that this study was conducted for an ecological assessment of the influence of habitat, feeding mode, and trophic level on microplastic abundance in benthic and epibenthic organism and that this species is not commonly consumed. Feng et al. (2020) reported a higher prevalence of microplastics (in 90% of the individuals) in four species of sea urchins (Strongylocentrotus intermedius, Temnopleurus hardwickii, Temnopleurus reevesii, and Hemicentrotus pulcherrimus) harvested from 12 sites along the northern China coast. The average abundance of microplastics (predominantly as fibers) in soft tissue from sea urchins from all sites was 5 particles/individual (1.1 particles/g), considerably higher than reported in heart urchins from the Oslofjord, Norway (Bour et al. 2018). Higher detection rates and abundances were found in sea urchins from Dalian, China (Feng et al. 2020). The tissue of relevance in urchins with regards to seafood safety is the gonads, and while whole soft tissue of heart urchins was analyzed for microplastics (Bour et al. 2018), the abundance in urchins from the Yellow Sea was assessed in gonads, coelomic fluid, and the gut. Gonads and coelomic fluid contained significantly lower number of particles/individual than the gut in all four species of urchin; however, this difference was not evident when normalized to wet weight in three of the species, and it only remained significantly lower in S. intermedius (Feng et al. 2020).
Microplastic ingestion has been reported in several species of sea cucumber including Holothuria grisea, Cucumaria frondosa, Holothuria floridana, Thyonella gemmata (Graham and Thompson 2009), Holothuria tubulosa (Renzi et al. 2018b), Holothuria mexicana, Actinopyga agassizi (Plee and Pomory 2020), and Apostichopus japonicus (Mohsen et al. 2019). Sea cucumbers are commonly eaten in Asia, and farming is widespread to meet consumer demand. The body wall of sea cucumbers is typically eaten raw in Japan and boiled, pickled, or salted in China, and the internal organs (gonads, respiratory trees, and intestines) are also edible (Kiew and Don 2011). Iwalaye et al. (2020) reported microplastic particles in the intestines, coelomic fluid, and respiratory trees of the Holothuria cinerascens and that uptake was both via the feeding tentacles and the respiratory trees. The most abundant microplastics found in farmed sea cucumbers (Apostichopus japonicus) from eight locations in the Bohai Sea and Yellow Sea in China were cellophane microfibers (Mohsen et al. 2019).
5.5 Microplastics in Gastropods
Limited research has been carried out on microplastics in marine gastropods. Xu et al. (2020) analyzed nine species of gastropods from shores in Hong Kong for microplastics, with the highest abundance found in sea snails (Batillaria multiformis, 5.4 ± 1.2 particles/g wet weight) and the lowest observed in Chameleon nerite snails (Nerita chamaeleon, 1.50 ± 0.2 particles/g wet weight). The common periwinkle (Littorina littorea) sampled from four different locations in Galway Bay, Ireland, contained between 0.6 and 2.8 microplastics/g wet weight of soft tissue, and commercial common periwinkles, intended for human consumption, contained on average 2.2 microplastic s/g wet weight soft tissue (Doyle et al. 2019). Most of the microplastics (97%) recorded in periwinkles were fibers. Similarly, fibers accounted for more than half of the total microplastics present in the girdled horn shell (Cerithidea cingulata), whereas film was the most abundant microplastic (approximately 44%) in Thais mutabilis from the Persian Gulf region. The mean number of total microplastics was 13 and 20 particles/g wet soft tissue weight for C. cingulata and T. mutabilis, respectively (Naji et al. 2018). Lower levels of microplastic contamination were reported in periwinkles (Littorina spp.) from two sites on the eastern coast of Thailand with an average of 0.17 particles/g wet weight and 0.23 particles/g wet weight, with no contamination observed in periwinkles from Bangasaen, the third site investigated (Thushari et al. 2017).
5.6 Microplastics in Cephalopods
Cephalopods are the seafood phylum which have received the least focus with regard to microplastic contamination. Oliveira et al. (2020) investigated the levels of microplastics in the stomach, caecum/intestine, and digestive gland of cuttlefish (Sepia officinalis); however, tissue relevant for consumer exposure was not included in this study. Microplastic contamination in Indian squid (Uroteuthis duvaucelii) was found in 18% of the individuals examined, with an average of 0.2 microplastic particles/individual and 0.008 microplastic particles/g wet weight of edible tissue (Peng et al. 2020).
5.7 Microplastics in Crustaceans
Most biota-based studies have examined microplastics in the organisms’ gut, which is not generally an organ consumed directly by humans. However, shellfish including crustaceans and mollusks are an exception since these are frequently eaten either whole or with their gut removed. The risk of ingesting microplastics from other tissues, such as muscle, depends on the ability to cross the intestinal barrier and subsequent accumulation (Zeytin et al. 2020).
To date, most literature on crustaceans which are commonly harvested for human consumption has focused on wild individuals, rather than those that are farmed. Investigations generally have not focused on crustaceans in the context of seafood safety but rather from an environmental contaminant perspective. For example, there have been numerous investigations into langoustine, Nephrops norvegicus, which are also commercially exploited. N. norvegicus, sampled from the Clyde Sea area, were shown to contain more microplastic fibers in their gut than individuals from the North Sea and Minch where only a small percentage of individuals contained microplastic, predominantly as single-strand fibers (Welden and Cowie 2016). Other commercially relevant species, such as spinous spider crabs (Maja squinado), shrimps, and prawns, have been observed to contain microplastics (Welden et al. 2018; Devriese et al. 2015; Zhang et al. 2019b; Cau et al. 2019).
Many crustaceans are harvested from coastal environments, which may be close to sources of microplastic contamination, including the influence of terrestrial plastic sources. As an example, shrimp (A. antennatus) from the Mediterranean had an average occurrence of microplastics equal to 39.2%; however, those in close vicinity to urban areas had 100% presence of microplastics (Carreras-Colom et al. 2018). The same % occurrence trend was observed between remote populations (<40%) of N. norvegicus compared to those sampled near Glasgow (84%) in the Clyde Sea (Welden and Cowie 2016).
Additionally, no spatial pattern was observed in a similar study of N. norvegicus in Irish waters (Hara et al. 2020). Both N. norvegicus and Aristeus antennatus were investigated in the Mediterranean Sea from depths between 270 and 660 meters (Cau et al. 2019). The authors reported a significant difference in the size and composition of microplastics identified between the two species and suggested that the nonselective feeding strategy of N. norvegicus likely led to a higher degree of exposure to microplastics and hence a higher measured abundance. Nonselective feeding is an example of direct uptake of microplastics from the environment. Organisms can also ingest microplastics which have been internalized by prey species, a concept commonly referred to as trophic transfer. Laboratory studies on this topic performed with shore crabs (Carcinus maenas) fed mussels which had been exposed to microplastics showed that polystyrene microspheres could accumulate in the foregut of the crabs (Watts et al. 2015).
Fibers and fragments are the most often reported particle type in crustaceans sampled from the wild, with fiber bundles reported across many species (Welden and Cowie 2016; Cau et al. 2019; McGoran et al. 2020). In most studies, stomachs were often the target organ of microplastics investigations, but other tissues are starting to be considered further, as these may have relevance for human exposure, especially when stomachs are removed prior to cooking and consumption. As an example, microplastics have been found in different tissues of wild-caught Portunus gracilimanus and P. trituberculatus (Zhang et al. 2019b).
5.8 Microplastics in Finfish
Evaluating microplastic occurrence and abundance in finfish is fundamental to understanding how plastics and their associated chemical compounds affect and potentially impact wild fisheries that are relied upon by humans as an important source of food and nutrition (Rochman et al. 2015; Barboza et al. 2018; FAO 2020; Lusher and Welden 2020). The topic of microplastics in the marine environment, including information on finfish, has been reviewed by several authors (Andrady 2011; Cole et al. 2011; Hidalgo-Ruz et al. 2012; Wright et al. 2013; Gall and Thompson 2015; Galloway et al. 2017; Baechler et al. 2020a, b; Thushari and Senevirathna 2020; Wang et al. 2020; Walkinshaw et al. 2020). Microplastics exposure in finfish is largely a result of plastics being mistaken for natural prey items, via ingestion of contaminated prey items or by passive uptake through gills (Lusher et al. 2016; Watts et al. 2015; Nelms et al. 2018; Roch et al. 2020). Trophic transfer of microplastics may also expose predaceous fish to microplastics (Farrell and Nelson 2013; Setälä et al. 2014; Lusher et al. 2016; Baechler et al. 2020a), and microplastics have frequently been detected in finfish gastrointestinal tracts (e.g., Lusher et al. 2017b,). The methodological challenges with identifying particles within fillet muscle tissue have limited the number of published studies thus far, although they have been identified albeit at extremely low concentrations (Zeytin et al. 2020).
Many species of edible demersal, pelagic, and reef fish, sampled from across the globe, have been found to contain microplastics (e.g., Bellas et al. 2016; Rummel et al. 2016; Bråte et al. 2016; Lusher et al. 2013; Tanaka and Takada 2016; Rochman et al. 2015; Neves et al. 2015; Critchell and Hoogenboom 2018; Abbasi et al. 2018; Su et al. 2018). The percentages of different fish species which have been found to contain microplastics in their gut vary greatly: 0.9% Peruvian anchovy, 2.8% Atlantic cod, 8.8% Atlantic herring, 9.4% Skipjack tuna, 24.5% Jack and Horse mackerel, 23.3% Pacific chub mackerel, 23.4% Yellowfin tuna, and 76.6% Japanese anchovy (Neves et al. 2015; Lusher et al. 2013; Güven et al. 2017; Ogonowski et al. 2017; Rummel et al. 2016; Hermsen et al. 2018; Rochman et al. 2015; Choy and Drazen 2013; Markic et al. 2018; Bråte et al. 2016; Liboiron et al. 2016; Tanaka and Takada 2016). Several studies have examined the microplastic particle prevalence in fish with different feeding ecology (Foekema et al. 2013; Lusher et al. 2013). Lusher et al. (2013) did not find any significant difference between the abundance of plastic ingested by pelagic and demersal fish. Of the 24 fish species examined from the Beibu Gulf, one of the world’s largest fishing grounds, in the South China Sea, 12 species contained microplastics (Koongolla et al. 2020). The abundance of microplastics varied from 0.027 to 1 item per individual, and most was present in fish stomach (57.7%) and less in intestines and gills (34.6% and 7.7%, respectively). Nine of the 11 fish species sampled from Zhoushan fishing grounds in the East China Sea were found to contain microplastics, with 23 different polymer types identified, and the highest number of items was 8 in a single individual (Zhang et al. 2019a). It is challenging to compare all the studies listed above, as many different methods have been utilized by researchers to determine the presence or absence of microplastics across these species. Some trends in the methods used have previously been described, with visually searching the most common method (Lusher et al. 2017b); however, the lack of standards and incomplete reporting of data, and quality control procedures have also been highlighted (Hermsen et al. 2018). Differences in sampling and analytical methods may lead to different values being observed and are important to consider when evaluating trends across regions, ecosystem types, and species.
The microplastic content of wild fish has been more widely studied than aquaculture species. A recent review of microplastics in seafood found that data were lacking for four of the ten most cultured aquatic food species, namely, grass carp, whiteleg shrimp, bighead carp, and catla (Walkinshaw et al. 2020).
5.9 Co-contaminants Associated with Microplastics in Seafood
The role of marine microplastics as vectors for major ocean pollutants was recently reviewed by Ziccardi et al. (2016), Santillo et al. (2017), and Amelia et al. (2021). Plastics are inherently complex in size, morphology, and polymer composition and may contain a range of additives, including plasticizers, stabilizers, pigments, fillers, and flame retardants which may leach out into the environment including air, water, and food, and in general, microplastics are now considered to represent a suite of co-contaminants (Rochman et al. 2019). More than 50% of plastics are associated with hazardous monomers, additives, and chemical byproducts (Lithner et al. 2011). Plastics have been shown to accumulate various organic and inorganic co-contaminants from the surrounding water column (Rochman et al. 2013, 2015). The high surface area to volume ratio of small particles and hydrophobic nature facilitate the sorption of chemicals on the plastic surface, forming a complex mixture of contaminants available to marine organisms (Rochman et al. 2013). Laboratory studies have demonstrated that continuous exposure to contaminated plastics can lead to the accumulation of plastic-associated co-contaminants in fish (Rochman et al. 2013; Wardrop et al. 2016).
Both field and modeling studies suggest that transfer of environmental pollutants through microplastics are negligible compared to other routes of uptake (Gouin et al. 2011; Bakir et al. 2016; Espinosa et al. 2018; Koelmans et al. 2016; Ziccardi et al. 2016; Lohmann 2017; Smith et al. 2018). Nonetheless, caution is warranted as many of the chemicals sorbed onto microplastics are known to be potent toxicants to humans and marine biota, triggering adverse effects such as endocrine disruption, neurological disorders, and reduced reproductive success (GESAMP 2016). An example of this is the investigation by Barboza et al. (2020a) who reported significantly higher concentrations of bisphenols in fish with microplastics compared to individuals with no microplastics. However, none of the fish species investigated (European seabass Dicentrarchus labrax, Atlantic horse mackerel Trachurus trachurus, and Atlantic chub mackerel Scomber colias) contained bisphenol A levels which would lead to an exceedance of the Tolerable daily Intake established by the European Food Safety Authority (EFSA) (Barboza et al. 2020a).
5.10 Microplastic Uptake and Toxicity in Humans
The uptake of microplastics is dependent on size, morphology, solubility, and surface charge and chemistry. Microplastics <130 μm in diameter can potentially translocate into human tissue (EFSA 2016), and particles sized 1.5 μm and below can penetrate capillaries (Yoo et al. 2011). Proposed mechanisms for uptake of microplastics include endocytotic and paracellular transfer across epithelial tissues (Wright and Kelly 2017). It is estimated that 90% of ingested microplastics are excreted from the body (EFSA 2016); however, the remaining microplastics may be detrimental to human health, and further research is required to develop a more comprehensive understanding regarding public health aspects of microplastic pollution.
Oxidative stress and subsequent inflammation are both thought to be the main mechanisms of particle toxicity (Feng et al. 2016). Other potential biological responses to microplastic exposure include genotoxicity, apoptosis, and necrosis, which could ultimately lead to tissue damage, fibrosis, and carcinogenesis (Wright and Kelly 2017). The extent of potential adverse effects is dependent on particle size, and nanoparticles have been found to generate more reactive oxygen species (ROS) than larger particles and are more likely to be translocated (Stone et al. 2007). Consequently, potential health effects of microplastics largely depend on particle characteristics, and it is envisaged that nanoplastics are likely more deleterious than microplastics (Feng et al. 2016).
5.11 Consequences of Microplastics in Marine Animals
More than 690 marine species from different trophic levels have been reported to contain plastic debris; however, the transfer of microplastics and associated co-contaminants, from seafood to humans, and the implications for seafood safety have received limited attention to date (Carbery et al. 2018; Lusher et al. 2017a; Walkinshaw et al. 2020). Most studies conducted have considered the environmental rather than the potential human health impacts of micro- and nanoplastics. Effects of micro- and nanoplastic exposure reported in marine organisms include reduced growth, impacted energy metabolism, feeding behavior, and locomotion, effects on the immune system, and hormonal regulation, physiological stress, oxidative stress, inflammation, aberrant development, cell death, general toxicity, and altered lipid metabolism (Kögel et al. 2019). In humans, it is evidenced that consumers may be exposed to microplastics from seafood consumption; however, the risks remain unclear (Smith et al. 2018; VKM 2019).
Shellfish and small fish that are consumed whole are the seafoods which are likely to give the highest exposure risk since the gastrointestinal tract, which generally contains the highest microplastic concentrations, are consumed (van Raamsdonk et al. 2020). In contrast most fish species are filleted, and most crustaceans have their digestive tracts removed before consumption, thereby reducing microplastic exposure . Similarly, bivalves, shellfish, and other lower trophically positioned marine organisms are probably the most important seafood source of dietary exposure to microplastics (Walkinshaw et al. 2020). It has been estimated that the average European shellfish consumer may ingest up to 11,000 microplastics per year based on levels in mussels and oysters (van Cauwenberghe and Janssen 2014). A systematic review and meta-analysis of microplastic contamination in seafood reported a maximum annual consumption of 55,000 microplastic particles, with mollusks from Asia being the most heavily contaminated (Danopoulos et al. 2020).
The presence of several types of microplastics in human stool from different countries has been reported, with PP and PET being the most abundant types (Schwabl et al. 2019), indicating human exposure. However, there is currently no indisputable evidence on the effects of microplastics on human health (Toussaint et al. 2019). Potential health impacts can result directly such as tissue damage but also indirectly from environmental contaminants associated with microplastics or associated microorganisms (Oberbeckmann et al. 2015).
While the focus of the scientific literature has primarily been on human exposure to microplastics from seafood consumption, much less data is available on the occurrence of microplastics in other food groups, so their relative contribution is unknown which is important from a risk assessment perspective (Wright and Kelly 2017). Data on microplastics in foods (Kwon et al. 2020) other than seafood include sugar, salt, honey, and drinking water and beer (Karbalaei et al. 2018), whereas there are significant data gaps for plant- and terrestrial animal-derived foods (van Raamsdonk et al. 2020). To date there are also limited data on microplastic levels in freshwater fish (Collard et al. 2019) and terrestrial foods (e.g., vegetables, poultry). In addition to food and drinking water, inhalation is a potential route of exposure, and atmospheric fallout is thus also an important source of microplastic exposure (Dris et al. 2016; Allen et al. 2019). Catarino et al. (2018) concluded that the potential for microplastic ingestion from shellfish consumption was minimal, especially when compared to general air exposure from household dust (123–4620 particles/year/capita and 13,731–68,415 particles/year/capita from food versus dust, respectively). Similarly, Rist et al. (2018) highlighted that food and beverages likely only constitute a minor exposure pathway to human microplastic exposure . Based on the current knowledge on microplastics in seafood, there is no evidence that food safety is compromised (Gamarro et al. 2020).
The extent to which microplastics present in foods contribute to human exposure is not well understood, especially as studies evaluating microplastics and associated chemical exposure to humans are not consistent (Rist et al. 2018; Barboza et al. 2020a, b). Human exposure estimates in the USA to microplastics in food (seafood , sugars, salts, honey), drinking water, alcohol, and air found that inhalation was the main route of exposure for adults whereas drinking water was the main source for children (Cox et al. 2019). However, this study did not include major food groups such as meats, grains, and vegetables due to a lack of empirical data.
5.12 Challenges and Priorities in Marine Microplastic Research
Risk characterization including information on the particle size-dependent toxico-kinetics and dynamics of microplastics is needed to calculate evidence-based guidance or tolerable weekly intakes to support realistic human health and exposure risk assessments. Discrepancies exist in the sampling, extraction, identification, and quantification of microplastics (Collard et al. 2019), and there is a need for harmonization of current procedures (Hartmann et al. 2019; Cowger et al. 2020). An effective risk assessment of the human health effects of microplastics requires reliable human exposure data which is currently limited (Toussaint et al. 2019). Knowledge gaps regarding the uptake and potential human health effects of microplastic exposure have been highlighted (EFSA 2016; Wright and Kelly 2017; van Raamsdonk et al. 2020).
Importantly, there is currently still a lack of harmonized and proven methods for microplastics which can compromise the level to which microplastic contamination in seafood species (and other foods) can be compared. Some recommendations have been presented which focus on the methods that – thus far – have proven efficient at isolating microplastics from biota tissues (e.g., Dehaut et al. 2019; Lusher et al. 2020; Ribeiro et al. 2020). The field of microplastic research has been moving very rapidly, with several advancements in methods emerging in parallel. It is therefore of great urgency to coordinate an effort to compare the field and laboratory-based methods to one another to determine the level of comparability and overall effectiveness. This is easier said than done. Currently, laboratory comparisons have been limited to scientific approaches conducted by individual research groups (e.g., Catarino et al. 2017; Karlsson et al. 2017; Thiele et al. 2019; Yu et al. 2019; Jaafar et al. 2020; Ribeiro et al. 2020), rather than between different institutions. Some interlaboratory efforts have been made, but these have generally focused on clean water samples, rather than complex matrices such as seafood material and biological tissues (e.g., ongoing EU-JRC and SCCWRP intercalibration exercises). Similarly, there are different reporting criteria that have been applied to the study of microplastics in biota, and the quantification of the microplastics is not standardized which presents some important challenges to this subdiscipline of environmental chemistry. Different measurement units are often used (e.g., numbers per weight or per individual) highlighting the need for harmonization and standardization.
Moving forward, methods will need to be adopted that are truly reproducible and that can be validated and compared using standard reference materials. This requires that validation and feasibility assessments are undertaken while also supporting initiatives that promote scientific discovery and method development. There are several methods that are promising, and utilizing these novel approaches will allow for the development of more robust and comparable methods across different sectors/regions within the sphere of microplastic research. Unfortunately, several methods are focused on the larger fraction of microplastics (e.g., >100 μm), and method development is still required for accurately detecting smaller microplastics (<20 μm) and nanoplastics (<1 μm). Methods that focus on smaller fractions are needed to support risk characterization and exposure assessments in marine biota and humans.
5.13 Future Recommendations and Conclusions
Microplastics are ubiquitously found in seafood , and the importance of standardized and harmonized methods for the effective biomonitoring of farmed and wild seafood species including bivalves and finfish has become evident (Lusher et al. 2017b; Hartmann et al. 2019; Ribeiro et al. 2020). Lower trophically positioned organisms may be at the highest risk of contamination from microplastics, and currently there is insufficient evidence to conduct realistic and meaningful human health risk assessments. Moreover, several seafood species from wild fisheries and aquaculture are not well studied in the context of global food security including commonly consumed taxa (Lusher et al. 2017a; Walkinshaw et al. 2020). Microplastic pollution and exposure to plastics and their associated co-contaminants via seafood consumption will likely serve as effective themes to help link the IOC-UNESCO’s Decade of Ocean Science for Sustainable Development (2021–2030) with the UN Decade of Action on Nutrition (2016–2025) and to gather critical stakeholders and develop important sustainable development strategies to support ocean and human health. In conclusion, the effects of microplastics on food security are still largely unknown, and further research and robust biomonitoring efforts on seafood are required to elucidate potential impacts.
References
Abbasi S, Soltani N, Keshavarzi B, Moore F, Turner A, Hassanaghaei M (2018) Microplastics in different tissues of fish and prawn from the Musa Estuary, Persian Gulf. Chemosphere 205:80–87
Abidli S, Lahbib Y, El Menif NT (2019) Microplastics in commercial molluscs from the lagoon of Bizerte (Northern Tunisia). Mar Pollut Bull 142:243–252
Allen S, Allen D, Phoenix VR (2019) Atmospheric transport and deposition of microplastics in a remote mountain catchment. Nat Geosci 12:339–344
Allen S, Allen D, Moss K, Le Roux G, Phoenix VR, Sonke JE (2020) Examination of the ocean as a source for atmospheric microplastics. PLoS One 15(5):e0232746. https://doi.org/10.1371/journal.pone.0232746
Amelia TSM, Khalik WMAWM, Ong MC, Shao YT, Pan H-J, Bhubalan K (2021) Marine microplastics as vectors of major ocean pollutants and its hazards to the marine ecosystem and humans. Prog Earth Planet Sci 8:12
Andrady AL (2011) Microplastics in the marine environment. Mar Pollut Bull 62:1596–1605
Baechler BR, Granek EF, Hunter MV, Conn KE (2020a) Microplastic concentrations in two Oregon bivalve species: spatial, temporal, and species variability. Limnol Oceanogr Lett 5:54–65
Baechler BR, Stienbarger CD, Horn DA, Joseph J, Taylor AR, Granek EF, Brander SM (2020b) Microplastic occurrence and effects in commercially harvested North American finfish and shellfish: current knowledge and future directions. Limnol Oceanogr Lett 5:113–136
Baechler BR, Granek EF, Mazzone SJ, Nielsen-Pincus M, Brander SM (2020c) Microplastic exposure by razor clam in recreational harvester-consumers along a sparsely populated coastline. Front Mar Sci 7:980
Bakir A, O’Connor IA, Rowland SJ, Hendriks AJ, Thompson RC (2016) Relative importance of microplastics as a pathway for the transfer of hydrophobic organic chemicals to marine life. Environ Pollut 219:56–65. https://doi.org/10.1016/j.envpol.2016.09.046
Barboza LGA, Vethaak AD, Lavorante BRBO, Lundebye A-K, Guilhermino L (2018) Marine microplastic debris: an emerging issue for food security, food safety and human health. Mar Pollut Bull 133:336–348
Barboza LGA, Cunha SC, Monteiro C, Fernandes JO, Guilhermino L (2020a) Bisphenol A and its analogs in muscle and liver of fish from the North East Atlantic Ocean in relation to microplastic contamination. Exposure and risk to human consumers. J Hazard Mater 393:122419
Barboza L, Lopes C, Oliveira P, Bessa F, Otero V, Henriques B, Raimundo J, Caetano M, Vale C, Guilhermino L (2020b) Microplastics in wild fish from North East Atlantic Ocean and its potential for causing neurotoxic effects, lipid oxidative damage, and human health risks associated with ingestion exposure. Sci Tot Environ 717:134625
Batel A, Linti F, Scherer M, Erdinger L, Braunbeck T (2016) Transfer of benzo[a]pyrene from microplastics to Artemia nauplii and further to zebrafish via a trophic food web experiment: CYP1A induction and visual tracking of persistent organic pollutants. Environ Toxicol Chem 35(7):1656–1666
Bellas J, Martínez-Armental J, Martínez-Cámara A, Besada V, Martínez-Gómez C (2016) Ingestion of microplastics by demersal fish from the Spanish Atlantic and Mediterranean coasts. Mar Pollut Bull 109:55–60. https://doi.org/10.1016/j.marpolbul.2016.06.026
Birnstiel S, Soares-Gomes A, da Gama BA (2019) Depuration reduces microplastic content in wild and farmed mussels. Mar Pollut Bull 140:241–247
Bour A, Avio CG, Gorbi S, Regoli F, Hylland K (2018) Presence of microplastics in benthic and epibenthic organisms: influence of habitat, feeding mode and trophic level. Environ Pollut 243:1217–1225. https://doi.org/10.1016/j.envpol.2018.09.115
Bråte ILN, Eidsvoll DP, Steindal CC, Thomas KV (2016) Plastic ingestion by Atlantic cod (Gadus morhua) from the Norwegian coast. Mar Pollut Bull 112:105–110. https://doi.org/10.1016/j.marpolbul.2016.08.034
Bråte ILN, Hurley R, Iversen K, Beyer J, Thomas KV, Steindal CC, Green NW, Olsen M, Lusher A (2018) Mytilus spp. as sentinels for monitoring microplastic pollution in Norwegian coastal waters: a qualitative and quantitative study. Environ Pollut 243:383–393
Browne MA, Dissanayake A, Galloway TS, Lowe DM, Thompson RC (2008) Ingested microscopic plastic translocates to the circulatory system of the mussel, Mytilus edulis (L). Environ Sci Technol 42:5026–5031
Browne M, Crump P, Niven SJ, Teuten E, Tonkin A, Galloway T, Thompson R (2011) Accumulation of microplastic on shorelines worldwide: sources and sinks. Environ Sci Technol 45. https://doi.org/10.1021/es201811s
Carbery M, O'Connor W, Palanisami T (2018) Trophic transfer of microplastics and mixed contaminants in the marine food web and implications for human health. Environ Int 115:4. https://doi.org/10.1016/j.envint.2018.03.007
Carpenter EJ, Anderson SJ, Harvey GR, Miklas HP, Peck BB (1972) Polystyrene spherules in coastal waters. Science 178(4062):749–750. https://doi.org/10.1126/science.178.4062.749
Carreras-Colom E, Constenla M, Soler-Membrives A, Cartes JE, Baeza M, Padrós F, Carrassón M (2018) Spatial occurrence and effects of microplastic ingestion on the deep-water shrimp Aristeus antennatus. Mar Pollut Bull 133:44–52
Catarino AI, Thompson R, Sanderson W, Henry TB (2017) Development and optimization of a standard method for extraction of microplastics in mussels by enzyme digestion of soft tissues. Environ Toxicol Chem 36(4):947–951
Catarino AI, Macchia V, Sanderson WG, Thompson RC, Henry TB (2018) Low levels of microplastics (MP) in wild mussels indicate that MP ingestion by humans is minimal compared to exposure via household fibres fallout during a meal. Environ Polllut 237:675–684
Cau A, Avio CG, Dessì C, Follesa MC, Moccia D, Regoli F, Pusceddu A (2019) Microplastics in the crustaceans Nephrops norvegicus and Aristeus antennatus: flagship species for deep-sea environments? Environ Polllut 255:113107
Cho Y, Shim WJ, Jang M, Han GM, Hong SH (2019) Abundance and characteristics of microplastics in market bivalves from South Korea. Environ Pollut 245:1107–1116
Cho Y, Shim WJ, Jang M, Han GM, Hong SH (2020) Nationwide monitoring of microplastics in bivalves from the coastal environment of Korea. Environ Pollut 270:116175
Choy CA, Drazen JC (2013) Plastic for dinner? Observations of frequent debris ingestion by pelagic predatory fishes from the central North Pacific. Mar Ecol Prog Ser 485:155–163
Cole M, Lindeque P, Halsband C, Galloway SC (2011) Microplastics as contaminants in the marine environment: a review. Mar Pollut Bull 62:2588–2597
Collard F, Gasperi J, Gabrielsen GW, Tassin B (2019) Particle ingestion by wild freshwater fish: a critical review. Environ Sci Technol 53:12974–12988. https://doi.org/10.1021/acs.est.9b03083
Covernton GA, Collicutt B, Gurney-Smith HJ, Pearce CM, Dower JF, Ross PS, Dudas SE (2019) Microplastics in bivalves and their habitat in relation to shellfish aquaculture proximity in coastal British Columbia, Canada. Aquac Environ Interact 11:357–374. https://doi.org/10.3354/aei00316
Cowger W, Booth AM, Hamilton BM, Thaysen C, Primpke S, Munno K, Lusher AL, Dehaut A, Vaz VP, Liboiron M, Devriese LI, Hermabessiere L, Rochman C, Athey SN, Lynch JM, De Frond H, Gray A, Jones OAH, Brander B, Steele C, Moore S, Sanchez A, Nel H (2020) Reporting guidelines to increase the reproducibility and comparability of research on microplastics. Appl Spectrosc 74:1066–1077. https://doi.org/10.1177/0003702820930292
Cox KD, Covernton GA, Davies HL, Dower JF, Juanes F, Dudas SE (2019) Human consumption of microplastics. Environ Sci Technol 53:7068. https://doi.org/10.1021/acs.est.9b01517
Cózar A, Martí E, Duarte CM, García-de-Lomas J, van Sebille E, Ballatore TJ, Eguíluz VM, González-Gordillo JI, Pedrotti ML, Echevarría F, Troublè R, Irigoien X (2017) The Arctic Ocean as a dead end for floating plastics in the North Atlantic branch of the thermohaline circulation. Sci Adv 3(4):e1600582
Critchell K, Hoogenboom MO (2018) Effects of microplastic exposure on the body condition and behaviour of planktivorous reef fish (Acanthochromis polyacanthus). PLoS One 13(3):e0193308. https://doi.org/10.1371/journal.pone.0193308
Danopoulos E, Jenner LC, Twiddy M, Rotchell JM (2020) Microplastic contamination of seafood intended for human consumption: a systematic review and meta-analysis. Environ H Persp 126002-1. https://doi.org/10.1289/EHP7171
Davidson TM (2012) Boring crustaceans damage polystyrene floats under docks polluting marine waters with microplastic. Mar Pollut Bull 64(9):1821–1828
Davidson K, Dudas SE (2016) Microplastic ingestion by wild and cultured Manila clams (Venerupis philippinarum) from Baynes Sound, British Columbia. Arch Environ Contam Toxicol 71(2):147–156
De Witte B, Devriese L, Bekaert K, Hoffman S, Vandermeersch G, Cooreman K, Robbens J (2014) Quality assessment of the blue mussel (Mytilus edulis): comparison between commercial and wild types. Mar Pollut Bull 85(1):146–155
Dehaut A, Hermabessiere L, Duflos G (2019) Current frontiers and recommendations for the study of microplastics in seafood. Trends Anal Chem 116:346–359
Devriese LI, Van der Meulen MD, Maes T, Bekaert K, Paul-Pont I, Frère L, Vethaak AD (2015) Microplastic contamination in brown shrimp (Crangon crangon, Linnaeus 1758) from coastal waters of the southern North Sea and channel area. Mar Pollut Bull 98(1–2):179–187
Ding JF, Li JX, Sun CJ, He CF, Jiang FH, Gao FL, Zheng L (2018) Separation and identification of microplastics in digestive system of bivalves. Chin J Anal Chem 46(5):690–697
Doyle D, Gammell M, Frias J, Griffin G, Nash R (2019) Low levels of microplastics recorded from the common periwinkle, Littorina littorea on the west coast of Ireland. Mar Pollut Bull 149:110645. https://doi.org/10.1016/j.marpolbul.2019.110645
Dris R, Gasperi J, Saad M, Mirande C, Tassin B (2016) Synthetic fibres in atmospheric fallout: a source of microplastics in the environment? Mar Pollut Bull 104:290–293. https://doi.org/10.1016/j.marpolbul.2016.01.006
EFSA (2016) Presence of microplastics and nanoplastics in food, with particular focus on seafood. Panel on contaminants in the food chain. EFSA J 14:e04501. https://doi.org/10.2903/j.efsa.2016.4501
Eriksen M, Lebreton LC, Carson HS, Thiel M, Moore CJ, Borerro JC, Galgani F, Ryan PG, Reisser J (2014) Plastic pollution in the world’s oceans: more than 5 trillion plastic pieces weighing over 250,000 tons afloat at sea. PLoS One 9(12):e111913
Espinosa C, García Beltrán JM, Esteban MA, Cuesta A (2018) In vitro effects of virgin microplastics on fish head-kidney leucocyte activities. Environ Pollut 235:30–38. https://doi.org/10.1016/j.envpol.2017.12.054
FAO (2017) The state of food security and nutrition in the world 2017. Building resilience for peace and food security. FAO, Rome. https://doi.org/10.1080/15226514.2012.751351
FAO (2018) The state of world fisheries and aquaculture meeting the sustainable development goals. Food and Agriculture Organization of the United Nations. https://doi.org/10.1364/OE.17.003331
FAO (2020) The state of world fisheries and aquaculture 2020. Sustainability in action. Rome. doi: https://doi.org/10.4060/ca9229en
Farrell P, Nelson K (2013) Trophic level transfer of microplastic: Mytilus edulis (L.) to Carcinus maenas (L.). Environ Pollut 177:1–3
Feng S, Gao D, Liao F, Zhou F, Wang X (2016) The health effects of ambient PM2.5 and potential mechanisms. Ecotoxicol Environ Saf 128:67–74. https://doi.org/10.1016/j.ecoenv.2016.01.030
Feng Z, Wang R, Zhang T, Wang J, Huang W, Li J, Xu J, Gao G (2020) Microplastics in specific tissues of wild sea urchins along the coastal areas of northern China. Sci Total Environ 728:138660. https://doi.org/10.1016/j.scitotenv.2020.138660
Foekema EM, De Gruijter C, Mergia MT, van Franeker JA, Murk AJ, Koelmans AA (2013) Plastic in north sea fish. Environ Sci Technol 47(15):8818–8824. https://doi.org/10.1021/es400931b
Franzellitti S, Canesi L, Auguste M, Wathsala RHGR, Fabbri E (2019) Microplastic exposure and effects in aquatic organisms: a physiological perspective. Environ Toxicol Pharmacol 68:37–51
Gall SC, Thompson RC (2015) The impact of debris on marine life. Mar Pollut Bull 92:170–179
Galloway TS (2015) Micro- and nano-plastics and human health. In: Marine Anthropogenic Litter. https://doi.org/10.1007/978-3-319-16510-3_13
Galloway TS, Cole M, Lewis C (2017) Interactions of microplastic debris throughout the marine ecosystem. Nat Ecol Evol 1:1–8
Gamarro EG, Ryder J, Elvevoll EO, Olsen RL (2020) Microplastics in fish and shellfish – a threat to seafood safety? J Aquat Food Product Technol 29:417–425. https://doi.org/10.1080/10498850.2020.1739793
Gardon T, Reisser C, Soyez C, Quillien V, Le Moullac G (2018) Microplastics affect energy balance and gametogenesis in the pearl oyster Pinctada margaritifera. Environ Sci Technol 52(9):5277–5286
GESAMP (2016) Sources, fate and effects of microplastics in the marine environment: part two of a global assessment. In: Kershaw PJ, Rochman CM (eds) Joint Group of Experts on the Scientific Aspects of Marine Environmental Protection, Rep. Stud. GESAMP No. 93. 220 p
GESAMP (2019) Guidelines or the monitoring and assessment of plastic litter and microplastics in the ocean. In: Kershaw PJ, Turra A, Galgani F (eds) IMO/FAO/UNESCO-IOC/UNIDO/WMO/IAEA/UN/UNEP/UNDP/ISA Joint Group of Experts on the Scientific Aspects of Marine Environmental Protection, Rep. Stud. GESAMP No. 99. 130p
Gouin T, Roche N, Lohmann R, Hodges G (2011) A thermodynamic approach for assessing the environmental exposure of chemicals absorbed to microplastic. Environ Sci Technol 45:1466–1472
Graham ER, Thompson JT (2009) Deposit- and suspension-feeding sea cucumbers (Echinodermata) ingest plastic fragments. J Exp Mar Biol Ecol 368:22–29. https://doi.org/10.1016/j.jembe.2008.09.007
Green DS, Colgan TJ, Thompson RC, Carolan JC (2019) Exposure to microplastics reduces attachment strength and alters the haemolymph proteome of blue mussels (Mytilus edulis). Environ Pollut 246:423–434
Güven O, Gökdağ K, Jovanović B, Kıdeyş AE (2017) Microplastic litter composition of the Turkish territorial waters of the Mediterranean Sea, and its occurrence in the gastrointestinal tract of fish. Environ Pollut 223:286–294. https://doi.org/10.1016/j.envpol.2017.01.025
Hantoro I, Löhr AJ, Van Belleghem F, Widianarko B, Ragas A (2019) Microplastics in coastal areas and seafood: implications for food safety. Food Addit Contam 36(5):674–711
Hara J, Frias J, Nash R (2020) Quantification of microplastic ingestion by the decapod crustacean Nephrops norvegicus from Irish waters. Mar Pollut Bull 152:110905
Hartmann NB, Hüffer T, Thompson RC, Hassellöv M, Verschoor A, Daugaard AE, Rist S, Karlsson T, Brennholt N, Cole M, Herrling MP, Hess MC, Ivleva NP, Lusher AL, Wagner M (2019) Are we speaking the same language? Recommendations for a definition and categorization framework for plastic debris. Environ Sci Technol 53(3):1039–1047
Hermsen E, Mintenig SM, Besseling E, Koelmans AA (2018) Quality criteria for the analysis of microplastic in biota samples: a critical review. Environ Sci Technol 52(18):10230–10240
Hidalgo-Ruz V, Gutow L, Thompson RC, Thiel M (2012) Microplastics in the marine environment: a review of the methods used for identification and quantification. Environ Sci Technol 46:3060–3075
Hurley R, Woodward J, Rothwell JJ (2018) Microplastic contamination of river beds significantly reduced by catchment-wide flooding. Nat Geosci 11:251–257. https://doi.org/10.1038/s41561-018-0080-1
Iwalaye OA, Moodley GK, Robertson-Andersson DV (2020) The possible routes of microplastics uptake in sea cucumber Holothuria cinerascens (Brandt, 1835). Environ Pollut 264:114644. https://doi.org/10.1016/j.envpol.2020.114644
Jaafar N, Musa SM, Azfaralariff A, Mohamed M, Yusoff AH, Lazim AM (2020) Improving the efficiency of post-digestion method in extracting microplastics from gastrointestinal tract and gills of fish. Chemosphere 260:127649
Jang M, Shim WJ, Han GM, Song YK, Hong SH (2018) Formation of microplastics by polychaetes (Marphysa sanguinea) inhabiting expanded polystyrene marine debris. Mar Pollut Bull 131:365–369
Jiang Y, Yang F, Zhao Y, Wang J (2020) Greenland Sea Gyre increases microplastic pollution in the surface waters of the Nordic Seas. Sci Total Environ 712:136484
Kane IA, Clare MA, Miramontes E, Wogelius R, Rothwell JJ, Garreau P, Pohl F (2020) Seafloor microplastic hotspots controlled by deep-sea circulation. Science 368:1140–1145
Karbalaei S, Hanachi P, Walker TR, Cole M (2018) Occurrence, sources, human health impacts and mitigation of microplastic pollution. Environ Sci Pollut Res Int 25:36046–36063. https://doi.org/10.1007/s11356-018-3508-7
Karlsson TM, Vethaak AD, Almroth BC, Ariese F, van Velzen M, Hassellöv M, Leslie HA (2017) Screening for microplastics in sediment, water, marine invertebrates and fish: method development and microplastic accumulation. Mar Pollut Bull 122(1–2):403–408
Kiew PL, Don MM (2011) Jewel of the seabed: sea cucumbers as nutritional and drug candidates. Int J Food Sci Nutr 63(5):616–636. https://doi.org/10.3109/09637486.2011.641944
Koelmans AA, Bakir A, Burton GA, Janssen CR (2016) Microplastic as a vector for chemicals in the aquatic environment: critical review and model-supported reinterpretation of empirical studies. Environ Sci Technol 50(7):3315–3326. https://doi.org/10.1021/acs.est.5b06069
Kögel T, Bjorøy Ø, Toto B, Bienfait AM, Sanden M (2019) Micro- and nanoplastic toxicity on aquatic life: determining factors. Sci Total Environ 709:136050. https://doi.org/10.1016/j.scitotenv.2019.136050
Kolandhasamy P, Su L, Li J, Qu X, Jabeen K, Shi H (2018) Adherence of microplastics to soft tissue of mussels: a novel way to uptake microplastics beyond ingestion. Sci Total Environ 610-611:635–640. https://doi.org/10.1016/j.scitotenv.2017.08.053
Koongolla JB, Lin L, Pan Y-F, Yang C-P, Sun D-R, Liu S, Xu X-R, Maharana D, Huang J-S, Li H-X (2020) Occurrence of microplastics in gastrointestinal tracts and gills of fish from Beibu Gulf, South China Sea. Environ Pollut 258:113734. https://doi.org/10.1016/j.envpol.2019.113734
Kwon J-H, Kim J-W, Pham TD, Tarafdar A, Hong S, Chun S-H, Lee S-H, Kang D-Y, Kim J-Y, Kim S-B, Jung J (2020) Microplastics in food: a review on analytical methods and challenges. Int J Environ Res Public Health 17(18):6710. https://doi.org/10.3390/ijerph17186710
Lebreton L, van der Zwet J, Damsteeg J, Slat B, Andrady A, Reisser J (2017) River plastic emissions to the world’s oceans. Nat Commun 8:15611. https://doi.org/10.1038/ncomms15611
Li J, Yang D, Li L, Jabeen K, Shi H (2015) Microplastics in commercial bivalves from China. Environ Pollut 207:190–195
Li J, Qu X, Su L, Zhang W, Yang D, Kolandhasamy P, Shi H (2016) Microplastics in mussels along the coastal waters of China. Environ Pollut 214:177–184
Li J, Lusher AL, Rotchell JM, Deudero S, Turra A, Bråte ILN, Sun C, Hossain MS, Li Q, Kolandhasamy P, Shi H (2019) Using mussel as a global bioindicator of coastal microplastic pollution. Environ Pollut 244:522–533
Liboiron M, Liboiron F, Wells E, Richárd N, Zahara A, Mather C, Bradshaw H, Murichi J (2016) Low plastic ingestion rate in Atlantic cod (Gadus morhua) from Newfoundland destined for human consumption collected through citizen science methods. Mar Pollut Bull 113(1–2):428–437. https://doi.org/10.1016/j.marpolbul.2016.10.043
Lithner D, Larsson A, Dave G (2011) Environmental and health hazard ranking and assessement of plastic polymers based on chemical composition. Sci Total Environ 409:3309–3324
Lohmann R (2017) Microplastics are not important for the cycling and bioaccumulation of organic pollutants in the oceans-but should microplastics be considered POPs themselves? Integr Environ Assess Manag 13(3):460–465. https://doi.org/10.1002/ieam.1914
Lusher A, Welden NAC (2020) Microplastic impact in fisheries and aquaculture. In: Rocha-Santos T et al (eds) Handbook of microplastics in the environment. Springer, Cham
Lusher AL, McHugh M, Thompson RC (2013) Occurrence of microplastics in the gastrointestinal tract of pelagic and demersal fish from the English Channel. Mar Pollut Bull 67:94–99. https://doi.org/10.1016/j.marpolbul.2012.11.028
Lusher AL, O'Donnell C, Officer R, O'Connor I (2016) Microplastic interactions with North Atlantic mesopelagic fish. ICES J Mar Sci 73:1214–1225
Lusher A, Hollman P, Mendoza-Hill J (2017a) Microplastics in fisheries and aquaculture – status of knowledge on their occurrence and implications for aquatic organisms and food safety. FAO Fisheries and Aquaculture Technical Paper, 615
Lusher AL, Welden NA, Sobral P, Cole M (2017b) Sampling, isolating and identifying microplastics ingested by fish and invertebrates. Anal Methods 9(9):1346–1360
Lusher AL, Munno K, Hermabessiere L, Carr S (2020) Isolation and extraction of microplastics from environmental samples: an evaluation of practical approaches and recommendations for further harmonization. Appl Spectrosc 74(9):1049–1065
Markic A, Niemand C, Bridson JH, Mazouni-Gaertner N, Gaertner JC, Eriksen M, Bowen M (2018) Double trouble in the South Pacific subtropical gyre: increased plastic ingestion by fish in the oceanic accumulation zone. Mar Pollut Bull 136:547–564. https://doi.org/10.1016/j.marpolbul.2018.09.031
Mathalon A, Hill P (2014) Microplastic fibers in the intertidal ecosystem surrounding Halifax Harbor, Nova Scotia. Mar Pollut Bull 81(1):69–79
McGoran AR, Clark PF, Smith BD, Morritt D (2020) High prevalence of plastic ingestion by Eriocheir sinensis and Carcinus maenas (Crustacea: Decapoda: Brachyura) in the Thames estuary. Environ Pollut 265:114972
Mohsen M, Wang Q, Zhang L, Sun L, Lin C, Yang H (2019) Microplastic ingestion by the farmed sea cucumber Apostichopus japonicus in China. Environ Pollut 245:1071–1078. https://doi.org/10.1016/j.envpol.2018.11.083
Naji A, Nuri M, Vethaak AD (2018) Microplastics contamination in molluscs from the northern part of the Persian Gulf. Environ Pollut 235:113–120. https://doi.org/10.1016/j.envpol.2017.12.046
Napper IE, Thompson RC (2016) Release of synthetic microplastic plasticfibres fromdomestic washing machines: effects of fabric type and washing conditions. Mar Pollut Bull 112:39–45. https://doi.org/10.1016/j.marpolbul.2016.09.025
Nelms SE, Galloway TS, Godley BJ, Jarvis DS, Lindeque PK (2018) Investigating microplastic trophic transfer in marine top predators. Environ Pollut 238:999–1007. https://doi.org/10.1016/j.envpol.2018.02.016
Neves D, Sobral P, Ferreira JL, Pereira T (2015) Ingestion of microplastics by commercial fish off the Portuguese coast. Mar Pollut Bull 101(1):119–126. https://doi.org/10.1016/j.marpolbul.2015.11.008
Oberbeckmann SL, Martin GJ, Labrenz M (2015) Marine microplastic-associated biofilms – a review. Environ Chem 12(5):551–562. https://doi.org/10.1071/EN15069
Ogonowski M, Wenman V, Danielsson S Gorokhova E (2017) Ingested microplastic is not correlated to HOC concentrations in Baltic Sea herring. In: Proceedings of the 15th international conference on environmental science and technology, Rhodes, Greece, 31 August–2 September
Oliveira AR, Sardinha-Silva A, Andrews PLR, Green D, Cooke GM, Hall S, Blackburn K, Sykes AV (2020) Microplastics presence in cultured and wild-caught cuttlefish, Sepia officinalis. Mar Pollut Bull 160:111553. https://doi.org/10.1016/j.marpolbul.2020.111553
Peng L, Fu D, Qi H, Lan CQ, Yu H, Ge C (2020) Micro- and nano-plastics in marine environment: source, distribution and threats—a review. Sci Total Environ 698:134254. https://doi.org/10.1016/j.scitotenv.2019.134254
Phuong NN, Poirier L, Pham QT, Lagarde F, Zalouk-Vergnoux A (2018) Factors influencing the microplastic contamination of bivalves from the French Atlantic coast: location, season and/or mode of life? Mar Pollut Bull 129(2):664–674
Pitt JA, Trevisan R, Massarsky A, Kozal JS, Levin ED, Di Giulio RT (2018) Maternal transfer of nanoplastics to offspring in zebrafish (Danio rerio): a case study with nanopolystyrene. Sci Total Environ 643:324–334. https://doi.org/10.1016/j.scitotenv.2018.06.186
Plee TA, Pomory CM (2020) Microplastics in sandy environments in the Florida keys and the panhandle of Florida, and the ingestion by sea cucumbers (Echinodermata: Holothuroidea) and sand dollars (Echinodermata: Echinoidea). Mar Pollut Bull 158:111437. https://doi.org/10.1016/j.marpolbul.2020.111437
Porter A, Lyons BP, Galloway TS, Lewis C (2018) Role of marine snows in microplastic fate and bioavailability. Environ Sci Technol 52:7111–7119
Qu X, Su L, Li H, Liang M, Shi H (2018) Assessing the relationship between the abundance and properties of microplastics in water and in mussels. Sci Total Environ 621:679–686
Renzi M, Guerranti C, Blašković A (2018a) Microplastic contents from maricultured and natural mussels. Mar Pollut Bull 131:248–251
Renzi M, Blašković A, Bernardi G, Russo GF (2018b) Plastic litter transfer from sediments towards marine trophic webs: a case study on holothurians. Mar Pollut Bull 135:376–385. https://doi.org/10.1016/j.marpolbul.2018.07.038
Ribeiro F, Okoffo ED, O'Brien JW, Fraissinet-Tachet S, O'Brien S, Gallen M, Samanipour S, Kaserzon S, Mueller JF, Galloway T, Thomas KV (2020) Quantitative analysis of selected plastics in high-commercial-value Australian seafood by pyrolysis gas chromatography mass spectrometry. Environ Sci Technol 54:9408–9417
Rist S, Almroth BC, Hartmann NB, Karlsson TM (2018) A critical perspective on early communications concerning human health aspects of microplastics. Sci Total Environ 626:720–726
Roch S, Friedrich C, Brinker A (2020) Uptake routes of microplastics in fishes: practical and theoretical approaches to test existing theories. Sci Rep 10:3896
Rochman CM, Hoh E, Kurobe T, The SJ (2013) Ingested plastic transfers hazardous chemicals to fish and induces hepatic stress. Sci Rep 3:3263. https://doi.org/10.1038/srep03263
Rochman CM, Tahir A, Williams SL, Baxa DV, Lam R, Miller JT, The F-C, Werorilangi S, Teh SJ (2015) Anthropogenic debris in seafood: plastic debris and fibers from textiles in fish and bivalves sold for human consumption. Sci Rep 5:14340
Rochman CM, Brookson C, Bikker J, Djuric N, Earn A, Bucci K, Athey S, Huntington A, McIlwraith H, Munno K, De Frond H, Kolomijeca A, Erdle L, Grbic J, Bayoumi M, Borrelle SB, Wu T, Santoro S, Werbowski LM, Zhu X, Giles RK, Hamilton BM, Thaysen C, Kaura A, Klasios N, Ead L, Kim J, Sherlock C, Ho A, Hung C (2019) Rethinking microplastics as a diverse contaminant suite. Environ Toxicol Chem 38:703–711
Rummel CD, Löder MG, Fricke NF, Lang T, Griebeler EM, Janke M, Gerdts G (2016) Plastic ingestion by pelagic and demersal fish from the North Sea and Baltic Sea. Mar Pollut Bull 102(1):134–141. https://doi.org/10.1016/j.marpolbul.2015.11.043
Santillo D, Miller K, Johnston P (2017) Microplastics as contaminants in commercially important seafood species. Integr Environ Assess Manag 13:516–521
Schwabl P, Koppel S, Konigshofer P, Bucsics T, Trauner M, Reiberger T, Liebmann B (2019) Detection of various microplastics in human stool: a prospective case series. Ann Intern Med. https://doi.org/10.7326/M19-0618
Setälä O, Fleming-Lehtinen V, Lehtiniemi M (2014) Ingestion and transfer of microplastics in the planktonic food web. Environ Pollut 185:77–83
Smith M, Love DC, Rochman CM, Neff RA (2018) Microplastics in seafood and the implications for human health. Curr Environ Health Rep 5:375. https://doi.org/10.1007/s40572-018-0206-z
Stone V, Johnston H, Clift MJD (2007) Air pollution, ultrafine and nanoparticle toxicology: cellular and molecular interactions. IEEE Trans Nanobioscience 6(4):331–340. https://doi.org/10.1109/TNB.2007.909005
Su L, Deng H, Li B, Chen Q, Pettigrove V, Wu C, Shi H (2018) The occurrence of microplastic in specific organs in commercially caught fishes from coast and estuary area of East China. J Hazard Mater. https://doi.org/10.1016/j.jhazmat.2018.11.024
Sussarellu R, Suquet M, Thomas Y, Lambert C, Fabioux C, Pernet ME, Le Goïc N, Quillien V, Mingant C, Epelboin Y, Corporeau C, Guyomarch J, Robbens J, Paul-Pont I, Soudant P, Huvet A (2016) Oyster reproduction is affected by exposure to polystyrene microplastics. Proc Natl Acad Sci U S A 113(9):2430–2435. https://doi.org/10.1073/pnas.1519019113
Tanaka K, Takada H (2016) Microplastic fragments and microbeads in digestive tracts of planktivorous fish from urban coastal waters. Sci Rep 6:34351. https://doi.org/10.1038/srep34351
Thiele CJ, Hudson MD, Russell AE (2019) Evaluation of existing methods to extract microplastics from bivalve tissue: adapted KOH digestion protocol improves filtration at single-digit pore size. Mar Pollut Bull 142:384–393
Thushari G, Senevirathna J (2020) Plastic pollution in the marine environment. Heliyon 6(8):e04709. https://doi.org/10.1016/j.heliyon.2020.e04709
Thushari GGN, Senevirathna JDM, Yakupitiyage A, Chavanich S (2017) Effects of microplastics on sessile invertebrates in the eastern coast of Thailand: an approach to coastal zone conservation. Mar Pollut Bull 124:349–355. https://doi.org/10.1016/j.marpolbul.2017.06.010
Toussaint B, Raffael B, Angers-Loustau A, Gilliland D, Kestens V, Petrillo M, Rio-Echevarria IM, Van den Eede G (2019) Review of micro- and nanoplastic contamination in the food chain. Food Addit Contam A 36:639. https://doi.org/10.1080/19440049.2019.1583381
van Cauwenberghe L, Janssen CR (2014) Microplastics in bivalves cultured for human consumption. Environ Pollut 193:65–70
van Cauwenberghe L, Claessens M, Vandegehuchte MB, Janssen CR (2015) Microplastics are taken up by mussels (Mytilus edulis) and lugworms (Arenicola marina) living in natural habitats. Environ Pollut 199:10–17
van Raamsdonk LWD, van der Zande M, Koelmans AA, Hoogenboom RLAP, Peters RJB, Groot MJ, Peijnenburg ACM, Weesepoel YJA (2020) Current insights into monitoring, bioaccumulation, and potential health effects of microplastics present in the food chain. Foods. https://doi.org/10.3390/foods9010072
van Sebille E, Aliani S, Law KL, Maximenko N, Alsina JM, Bagaev A, Bergmann M, Chapron B, Chubarenko I, Cózar A, Delandmeter P, Egger M, Fox-Kemper B, Garaba SP, Goddijn-Murphy L, Hardesty BD, Hoffman MJ, Isobe A, Jongedijk CE, Kaandorp MLA, Khatmullina L, Koelmans AA, Kukulka T, Laufkötter C, Lebreton L, Lobelle D, Maes C, Martinez-Vicente V, Morales Maqueda MA, Poulain-Zarcos M, Rodríguez E, Ryan PG, Shanks AL, Shim WJ, Suaria G, Thiel M, van den Bremer TS, Wichmann D (2020) The physical oceanography of the transport of floating marine debris. Environ Res Lett 15:023003. https://doi.org/10.1088/1748-9326/ab6d7d
Vandermeersch G, Van Cauwenberghe L, Janssen CR, Marques A, Granby K, Fait G, Kotterman MJ, Diogène J, Bekaert K, Robbens J, Devriese L (2015) A critical view on microplastic quantification in aquatic organisms. Environ Res 143:46–55. https://doi.org/10.1016/j.envres.2015.07.016
VKM (2019) Microplastics; occurrence, levels and implications for environment and human health related to food. Scientific opinion of the Scientific Steering Committee of the Norwegian Scientific Committee for Food and Environment (No. 2019:16). Norwegian Scientific Committee for Food and Environment (VKM), Oslo, Norway
Walkinshaw C, Lindeque PK, Thompson R, Tolhurst T, Cole M (2020) Microplastics and seafood: lower trophic organisms at highest risk of contamination. Ecotoxicol. Environ Saf 190. https://doi.org/10.1016/j.ecoenv.2019.110066
Wang J, Lu L, Wang M, Jiang T, Liu X, Ru S (2019) Typhoons increase the abundance of microplastics in the marine environment and cultured organisms: a case study in Sanggou Bay, China. Sci Total Environ 667:1–8. https://doi.org/10.1016/j.scitotenv.2019.02.367
Wang W, Ge J, Yu X (2020) Bioavailability and toxicity of microplastics to fish species: a review. Ecotoxicol Environ Saf 189:109913. https://doi.org/10.1016/j.ecoenv.2019.109913
Ward JE, Kach DJ (2009) Marine aggregates facilitate ingestion of nanoparticles by suspension-feeding bivalves. Mar Environ Res 68(3):137–142
Ward JE, Rosa M, Shumway SE (2018) Capture, ingestion, and egestion of microplastics by suspension-feeding bivalves: a 40-year history. Mar Microplast Pollut Contr 2(1):39–49. https://doi.org/10.1139/anc-2018-0027@anc-mmpc.issue1
Ward JE, Zhao S, Holohan BA, Mladinich KM, Griffin TW, Wozniak J, Shumway SE (2019) Selective ingestion and egestion of plastic particles by the blue mussel (Mytilus edulis) and eastern oyster (Crassostrea virginica): implications for using bivalves as bioindicators of microplastic pollution. Environ Sci Technol 53(15):8776–8784
Wardrop P, Shimeta J, Nugegoda D, Morrison PD, Miranda A, Tang M, Clarke BO (2016) Chemical pollutants sorbed to ingested microbeads from personal care products accumulate in fish. Environ Sci Technol 50:4037–4044. https://doi.org/10.1021/acs.est.5b06280
Watts AJR, Urbina MA, Corr S, Lewis C, Galloway T (2015) Ingestion of plastic microfibers by the crab Carcinus maenas and its effect on food consumption and energy balance. Environ Sci Technol 49(24):14597–14604. https://doi.org/10.1021/acs.est.5b04026
Watts AJR, Urbina MA, Goodhead R, Moger J, Lewis C, Galloway TS (2016) Effect of microplastic on the gills of the shore crab Carcinus maenas. Environ Sci Technol 50:5364–5369. https://doi.org/10.1021/acs.est.6b01187
Wegner A, Besseling E, Foekema EM, Kamermans P, Koelmans AA (2012) Effects of nanopolystyrene on the feeding behavior of the blue mussel (Mytilus edulis L.). Environ Toxicol Chem 31(11):2490–2497
Weinstein JE, Crocker BK, Gray AD (2016) From macroplastic to microplastic: degradation of high-density polyethylene, polypropylene, and polystyrene in a salt marsh habitat. Environ Toxicol Chem 35:1632–1640. https://doi.org/10.1002/etc.3432
Welden NA, Cowie PR (2016) Environment and gut morphology influence microplastic retention in langoustine, Nephrops norvegicus. Environ Pollut 214:859–865
Welden NA, Abylkhani B, Howarth LM (2018) The effects of trophic transfer and environmental factors on microplastic uptake by plaice, Pleuronectes plastessa, and spider crab, Maja squinado. Environ Pollut 239:351–358
Wieczorek AM, Croot PL, Lombard F, Sheahan JN, Doyle TK (2019) Microplastic ingestion by gelatinous zooplankton may lower efficiency of the biological pump. Environ Sci Technol 53(9):5387–53955. https://doi.org/10.1021/acs.est.8b07174
Wright SL, Kelly FJ (2017) Plastic and human health: a micro issue? Environ Sci Technol 51(12):6634–6647. https://doi.org/10.1021/acs.est.7b00423
Wright SL, Thompson RC, Galloway TS (2013) The physical impacts of microplastics on marine organisms: a review. Environ Pollut 178:483–492
Xu XY, Lee WT, Chan AKY, Lo HS, Shin PKS, Cheung SG (2017) Microplastic ingestion reduces energy intake in the clam Atactodea striata. Mar Pollut Bull 124(2):798–802
Xu X, Wong CY, Tam NFY, Lo H-S, Cheung S-G (2020) Microplastics in invertebrates on soft shores in Hong Kong: influence of habitat, taxa and feeding mode. Sci Tot Environ 715:13699. https doi.org/10.1016/j.scitotenv.2020.136999
Yoo JW, Doshi N, Mitragotri S (2011) Adaptive micro and nanoparticles: temporal control over carrier properties to facilitate drug delivery. Adv Drug Deliv Rev 63(14–15):1247–1256
Yu Z, Peng B, Liu LY, Wong CS, Zeng EY (2019) Development and validation of an efficient method for processing microplastics in biota samples. Environ Toxicol Chem 38(7):1400–1408
Zeytin S, Wagner G, Mackay-Roberts N, Gerdts G, Schuirmann E, Klockmann S, Slater M (2020) Quantifying microplastic translocation from feed to the fillet in European sea bass Dicentrarchus labrax. Mar Pollut Bull 156:111210
Zhang F, Man YB, Mo WY, Man KY, Wong MH (2019a) Direct and indirect effects of microplastics on bivalves, with a focus on edible species: A mini-review. Crit Rev Environ Sci Technol 1–35. https://doi.org/10.1080/10643389.2019.1700752
Zhang F, Wang X, Xu J, Zhu L, Peng G, Xu P, Li D (2019b) Food-web transfer of microplastics between wild caught fish and crustaceans in East China Sea. Mar Pollut Bull 146:173–182. https://doi.org/10.1016/j.marpolbul.2019.05.061
Ziccardi LM, Edgington A, Hentz K, Kulacki KJ, Driscoll SK (2016) Microplastics as vectors for bioaccumulation of hydrophobic organic chemicals in the marine environment: a state-of-the-science review. Environ Toxicol Chem 35:1667–1676
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2022 The Author(s)
About this chapter

Cite this chapter
Lundebye, AK., Lusher, A.L., Bank, M.S. (2022). Marine Microplastics and Seafood: Implications for Food Security. In: Bank, M.S. (eds) Microplastic in the Environment: Pattern and Process. Environmental Contamination Remediation and Management. Springer, Cham. https://doi.org/10.1007/978-3-030-78627-4_5
Download citation
DOI: https://doi.org/10.1007/978-3-030-78627-4_5
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-78626-7
Online ISBN: 978-3-030-78627-4
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)