
Abstract
The nowadays hyper-diverse clade of Actinopterygii (ray-finned bony fishes) is characterized by a long evolutionary history and an extremely rich global fossil record. This work builds upon 170 years of research on the fossil record of this clade in Greece. The taxonomy and spatiotemporal distribution of the ray-finned fish record of Greece are critically revisited and placed in an updated systematic and stratigraphic framework, while some new fossil data and interpretations are also provided. Greece hosts diverse ray-finned fish assemblages, which range in age from Lower Jurassic to Quaternary. Most known assemblages are of Miocene–Pliocene age and of marine affinities. A minimum of 32 families, followed by at least 34 genera and 22 species, have been recognized in Greece. From originally two named genera and seven species, only two fossil species, established on Greek material, are accepted as valid. Additional taxonomic diversity is anticipated, pending detailed investigations. From a taxonomic perspective, previous knowledge lies on preliminary or authoritative assessments of fossils, with many decades-old treatments needing revision. Little is known about Mesozoic–early Cenozoic occurrences or freshwater assemblages. Given the proven potential of the Greek fossil record, this chapter stresses the need for additional exploration and the establishment of permanent, curated collections of fossil fishes in Greek institutions. Directions for future research are discussed.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Actinopterygii
- Neopterygii
- Teleostei
- Marine environment
- Brackish and freshwater environment
- Neogene
- Miocene
- Pliocene
1 Introduction
A continuously thickening count of over 33,000 living species, already surpassing that of all other vertebrate species combined, renders Actinopterygii (ray-finned bony fishes) the most diverse clade of vertebrate animals (Nelson et al. 2016; Eschmeyer and Fong 2019). This vast taxonomic richness is coupled with an astounding array of anatomical, feeding, reproductive, locomotory, and other biological specializations, which enabled ray-fins to conquer most aquatic niches on the planet (from abyssal plains to hypersaline volcanic lakes), and to even glide, or crawl, outside the aquatic medium (Helfman et al. 2009; Nelson et al. 2016; Froese and Pauly 2019).
Actinopterygian origins date back to at least the late Silurian (~425 Ma), as inferred by the age of the first fossils of their sarcopterygian sister group (Zhu et al. 2009). The oldest definitive actinopterygian fossils are dated in the Early and Middle Devonian (Giles et al. 2015; Lu et al. 2016). Similarly, all major living actinopterygian lineages (Cladistia; Chondrostei; Neopterygii) are widely thought to have diverged in the Paleozoic (Near et al. 2012; Broughton et al. 2013; Sallan 2014; Betancur-R et al. 2015; Friedman 2015), but their first accepted occurrences are in early and middle Mesozoic rocks (Olsen 1984; Hilton and Forey 2009; Giles et al. 2017). The current taxonomic dominance of actinopterygians relative to other aquatic vertebrates was foreshadowed by their impressive Triassic radiation(s), in the aftermath of the catastrophic Permian-Triassic extinction (~251 Ma; Friedman and Sallan 2012; Romano et al. 2016), and the appearance of Telostei in the Middle Triassic (total group: Tintori et al. 2015; Arratia 2017). The Cretaceous–Paleogene Extinction (K/Pg) led to the complete eradication or decimation of several long-lasting Mesozoic actinopterygian lineages, especially in marine waters (Friedman 2009; Friedman and Sallan 2012; Guinot and Cavin 2016). Paleogene and younger radiations in the newly emptied ecological spaces marked the beginning of a new era in the evolutionary history of the clade, in which Teleostei, largely represented by members of the ostariophysan (mostly in freshwaters) and acanthomorph (spiny-rayed fishes) super-radiations, are established as dominant vertebrate components of global aquatic niches (Alfaro et al. 2009, 2018; Sibert and Norris 2015).
The long evolutionary history of ray-finned fishes is evidenced by a particularly rich fossil record. Stratified aquatic deposits can yield large numbers of articulated, yet flattened individuals (for some examples from Greece, see Gaudant 2001, 2004). Laminated rocks, corresponding to rapid deposition and low energy and low bottom oxygenation environments, can sometimes allow the preservation of very delicate structures, such as gut contents, embryos, or even soft tissues and pigments (e.g., Viohl 1990; Vinther 2015; Argyriou et al. 2016; Maxwell et al. 2017), allowing for painting a more vivid picture of past ecosystems and biotic interactions within them, while providing important insights into the evolution of key anatomical structures and biological specializations. Three-dimensional anatomical structure is only rarely preserved in fossil actinopterygians, but such fossils can allow the study of structures that are lost in flattened individuals (e.g., braincases, sensory endocasts, gill skeletons), and are of tremendous value to paleontologists, especially in light of modern fossil imaging and investigation techniques (Friedman et al. 2015; Argyriou et al. 2018). Equally important biological and geological information often derives from studies of identifiable ichthyoliths (isolated bones, scales and teeth, Bellas et al. 1998; Argyriou et al. 2015; Vasileiadou et al. 2017) and otoliths, (e.g., Agiadi et al. 2013, 2019; Agiadi this volume). Due to their exquisite fossil record, actinopterygians constitute an excellent means for studying the evolution of aquatic life on the planet, and its response to major biotic crises and extinctions (Friedman and Sallan 2012; Guinot and Cavin 2016; Romano et al. 2016). At the same time, the largely ectothermic traits, in conjunction with the strict environmental and habitat preferences of most ray-fins (e.g., Helfman et al. 2009), establish their fossil record as a powerful tool for making paleoenvironmental and paleobiogeographical inferences (Gaudant 2002; Landini and Sorbini 2005a; Argyriou et al. 2015; Carnevale et al. 2018), often through the scope of isotope geochemistry (e.g., Otero et al. 2011).
Historically, the study of fossil actinopterygians has been associated with several challenges, chief of which is the sheer richness of actinopterygian biodiversity and fossil record, which encompasses numerous groups of tens or hundreds of millions of years of divergent evolutionary histories (Friedman and Sallan 2012; Near et al. 2012; Broughton et al. 2013; Betancur-R et al. 2015; Friedman 2015; Nelson et al. 2016). As a result, commonly used synapomorphies for many clades of ray-fins—mostly teleosts—are established on modern taxa alone, and their validity has not been questioned by large-scale phylogenetic analyses (see, e.g., Wiley and Johnson 2010). Placing early diverging fossil taxa without closely related modern representatives on the tree of life is an extremely difficult task and constitutes a frontier in current paleoichthyology (Sallan 2014; Friedman 2015; Giles et al. 2017; Argyriou et al. 2018). Even when modern-looking forms are examined, their study usually requires consultation of extensive comparative materials in collections. Often limited anatomical information coming from two-dimensionally flattened fossils, as well as prevalent issues related to structural homology—mostly encountered when examining deeply nested forms (Schultze 2008)—pose additional obstacles for deciphering the fossil record and history of the clade.
Fossil actinopterygians form an extremely important, but historically overlooked, component of Greek vertebrate paleontological research. This synthesis aims at bringing together available information on the rich record of body fossils of the clade in Greece and critically assessing historical attributions in terms of validity and reliability—at least according to the opinion of the author, and based on a critical review of the literature. By placing all relevant occurrences in an up-to-date systematic, geographic, and stratigraphic context, it is anticipated that this chapter will fuel future paleo-ichthyological research in Greece.
2 Historical Overview and Collection Information
Actinopterygians from the Miocene of Kymi, Euboea, were the first to attract scientific attention on fishes from Greece, during the 1800s (de Waldheim 1838, 1850; Valenciennes 1861; Gaudry 1862; Unger 1867), but these early authoritative—rather than synapomorphy-based—taxonomic assessments are in need of revision, especially since they are not accompanied by adequate figures of the material. Fossil cyprinodontids were discovered in the Messinian of Crete by Raulin (1861) and were later revisited by Woodward (1901). In the same work, Woodward updated the taxonomic status of fossil percomorphs from Kymi, first described by de Waldheim (1838, 1850), on the basis of material in the collections of what is nowadays the Natural History Museum of London (but see Centropomidae section below). After a four-decade hiatus, Weiler described a fossil clupeid from the Miocene of Serres Basin (Weiler 1943).
The second half of the twentieth century saw an increase in interest in the study of Greek ray-fin fossil record. Kottek described some fragmentary leptolepid remains from the Toarcian (Early Jurassic) of Leukada, which derive from a since unexplored, and allegedly rich fossil fish bonebed near the village of Anavryssada (Kottek 1964). This represents the stratigraphically oldest occurrence of fossil ray-fins in Greece known to date. Melentis was the first Greek paleontologist to publish on fossil actinopterygians (Melentis 1967a, b). Unfortunately, his accounts of a †pycnodontid from the Cenomanian (Late Cretaceous) of Lindos, Rhodos, should be treated with extreme caution (see †Pycnodontidae section below). Important, but largely overlooked and preliminary taxonomic lists and figures of Late Cretaceous–Paleogene actinopterygians from Eurytania, have been provided in a stratigraphic monograph by Koch and Nikolaus (1969).
Symeonidis and Bachmayer set the stage for later work on the diverse Neogene marine ichthyofaunas of Crete (Symeonidis 1969; Bachmayer and Symeonidis 1978; Bachmayer et al. 1984). Their taxonomic results in the case of the so-called †Mene psarianosi (= †Alectis psarianosi) and their newly erected syngnathiform species have been since revised (Bannikov 1987; Bürgin 1994; Gaudant 2001, 2004). Müller and Strauch (1994) also provided a short account of a fossil assemblage from the Pliocene of Prassas, Crete, which was subsequently revised (Gaudant 2001). Another interesting assemblage, containing both articulated skeletons, ichthyoliths and otoliths from the Messinian of Karpathos, was presented by Bellas et al. (Bellas et al. 1998). The bulk of research on Neogene marine actinopterygians was led by Gaudant between the late 1970s and the early 2010s. These works expanded on the alpha-taxonomic status and paleoenvironmental implications of fossils from Aegina (Gaudant et al. 2010b), Attiki (Gaudant and Symeonidis 1995), Crete (Gaudant 1979, 1980, 2001, 2004, 2014b; Gaudant et al. 1994, 1997a) and Gavdos Island (Gaudant et al. 2005, 2006), highlighted the richness and potential of the Miocene–Pliocene actinopterygian archive of Greece, and also contributed data for downstream analyses of Mediterranean actinopterygian diversity changes during the latter part of the Cenozoic (Gaudant 2002; Landini and Sorbini 2005a). Yet, detailed descriptions of Neogene actinopterygians from Greece and their systematic treatment are still lacking for the most part.
Few treatments of the Greek record of ray-fins have been published in the past two decades. Careful anatomical and systematic work on Tortonian (Přikryl and Carnevale 2017) and Pliocene (Sorbini and Tyler 2004) actinopterygians from Crete revealed the presence of new fossil species, indicating that actinopterygian diversity and taxonomy in the Neogene of Crete, and conceivably elsewhere in Greece, remain underappreciated. The study of Maastrichtian actinopterygians from Gavdos Island represents the first recent attempt to investigate the pre-Cenozoic fossil record of actinopterygians of the country (Cavin et al. 2012). The first investigation of microscopic ichthyoliths from Greek freshwater deposits (early Miocene, Lesvos Island) was only published a few years ago (Vasileiadou et al. 2017). Furthermore, a recently completed MSc. thesis has shed light on the late Miocene freshwater–brackish clupeids of Aidonochori, Serres Basin, Northern Greece (Kevrekidis 2016). Finally, a new study dealing with the Maastrichtian–Paleogene actinopterygian assemblages from Eurytania has been made available while this work was in press (Argyriou and Davesne 2021).
Over 170 years of research have contributed significantly toward creating a framework for the temporal, stratigraphic, and geographical distribution of fossil ray-fins in Greece, while also providing taxonomic assessments of relevant assemblages. At the same time, past research efforts have been sporadic and/or have largely concluded at a preliminary level, without tapping the potential informativeness of this rich record for a variety of subjects. These include the evolutionary histories and paleobiogeography of Eastern Mediterranean marine and freshwater faunas, or the study of major events, such as the K/Pg Extinction and the Messinian Salinity Crisis. Unlike late Cenozoic mammals (see relevant chapters in this volume), research on the actinopterygian record of Greece has not received enough scientific attention and is still in its infancy (but see otoliths in Agiadi this volume). Detailed descriptions of Greek fossils are largely wanting, and older taxonomic assessments are in dire need of revision, posing obstacles for further analytical work. This is further exacerbated by the lack of organized collections of fossil fishes and recent comparative material in Greek institutions. A rudimentary-only collection of Cretaceous–Pliocene fossil fishes is housed at the AMPG (organized by the author and S. Roussiakis). Additionally, numerous teleostean skeletons from several Neogene localities of Crete (mainly Bregmaceros and Spratelloides from the Heraklion Basin) form part of the collections of the Natural History Museum of Crete. By contrast, rich collections from the Cretan Neogene are housed in the Natural History Museum of Vienna (Gaudant 2001, 2004). Some few findings from the Miocene of Kymi, Euboea (Gaudry 1862; T.A. personal observations and communication with Giorgio Carnevale) and the Zanclean of Aegina Island (Gaudant et al. 2010b) are housed in the MNHN, Paris, while additional fossils from the Miocene of Crete and possibly Kymi are housed in the Natural History Museum of London (Woodward 1901).
3 Phylogenetic Relationships
Actinopterygians are widely accepted as a monophyletic group, despite owing their name to their rayed fins, which are also present in sarcopterygians (Friedman 2015). Combinations of hard-tissue synapomorphies [(i) absence of jugal canal; (ii) presence of median aortic canal in the occiput; (iii) absence of endoskeletal basal plates of dorsal and anal fins; (iv) narrow interorbital septum; (v) lateral cranial canal; (vi) posteriorly expanded supratemporals; (vii) acrodin caps on teeth; (viii) fenestrated metapterygoidean portion of palatoquadrate; (ix) perforate propterygium embraced by fin rays; (x) presence of ganoine] are employed to distinguish Paleozoic actinopterygians from other osteichthyes, though, naturally, distinguishing lines dim when examining deeply nested taxa (Friedman 2015; Giles et al. 2015; Lu et al. 2016).
The relationships among actinopterygians—neopterygians and teleosts in particular—had been a matter of controversy (e.g., see Patterson 1973 vs. Grande 2010, for the status of Holostei; Patterson and Rosen 1977 vs. Arratia 1997, for identifying the deepest branch of crown group Teleostei; or Davesne et al. 2016 for a summary of the complicated history of acanthomorph interrelationships) before the establishment of reliable molecular systematic methodologies (Near et al. 2012, 2013; Betancur-R et al. 2015; Hughes et al. 2018). More recent morphology-based investigations are slowly bridging the gap between molecular and morphological phylogenetic hypotheses (e.g., Grande 2010; Davesne et al. 2016). It is now accepted that extant Actinopterygii are distributed across three major lineages of inferred Paleozoic origins: the basally diverging Cladistia (ropefish and bichirs); Chondrostei (sturgeons and paddlefish); and Neopterygii, with the latter including the nowadays depauperate Holostei (gars and bowfin) and the overabundant Teleostei (Near et al. 2012; Betancur-R et al. 2015, 2017; Nelson et al. 2016; Hughes et al. 2018). The relationships of the major groups of ray-finned fishes found in Greece are shown in Fig. 1.

Fossil record and time-calibrated phylogenetic relationships of major groups of Greek fossil fishes (skeletal record only). Only occurrences considered as valid herein are taken into account. Occurrences marked in darker gray boxes correspond to better constrained ages (usually biostratigraphically). White occurrence boxes correspond to broadly or tentatively assigned ages. Phylogenetic placement and time calibrations for extant groups follow Betancur-R et al. 2017. Placement of fossil taxa is as follows: †Pycnodontidae are established as non-teleosteomorphs; †Leptolepididae and †Ichthyodectoidei (†Ichthyodectiformes) sensu Arratia 2017; †Dercetidae and †Enchodontidae as closely related to modern Aulopifomes sensu Davis and Fielitz 2010, although an agnostic placement of fossil taxa relative to extant ones is preferred here. Results from Argyriou and Davesne (2021) were not incorporated in the figures, or the text, as this work became available after the preparation of this chapter
Teleostei account for ~99% of living actinopterygian species and are strongly supported as monophyletic by both morphological and molecular data (Near et al. 2012; Betancur-R et al. 2013, 2015; Broughton et al. 2013; Arratia 2017; López-Arbarello and Sferco 2018). Numerous hard-tissue synapomorphies support teleost monophyly, including: (i) orbital region of skull narrower than postorbital; (ii) presence of one suborbital bone between the posterodorsal margin of the infraorbital series and the opercular apparatus; (iii) presence of two supramaxillae; (iv) lower jaw articulation located ventral to the posterior margin of the orbit; (v) articular fused with angular and retroarticular; (vi) notch on ascending margin of dentary; (vii) coronoid process formed by angular and dentary; (viii) lateral ridge on dentary separating dental and splenial portions of lower jaw; (ix) long epineural processes; (x) presence of 20–26 principal rays; (xi) presence of an enlarged dorsal scute preceding caudal fin (sensu Arratia 2017).
Apart from one problematic occurrence of a Cenomanian †pycnodontid in Rhodos Island (Melentis 1967a, b), the Greek actinopterygian archive is exclusively populated by teleosts (Appendix). †Leptolepid remains from the Toarcian of Leukada (Kottek 1964) represent the only ascertained stem teleost occurrence in Greece while all remaining occurrences correspond to members of crown group lineages. Reliably documented Mesozoic actinopterygians from Greece have been ascribed to extinct families (e.g., †Dercetidae, †Enchodontidae; see Koch and Nikolaus 1969; Cavin et al. 2012), whereas Cenozoic fossils have been predictably included in extant taxa (families, genera, and in some cases to the species-level; see Appendix) on the basis of broad morphological and meristic similarities. The phylogenetic affinities of Greek actinopterygian fossils have not yet been explored, to the exclusion of Enchodus cf. dirus from the Maastrichtian of Gavdos Island (Cavin et al. 2012).
4 Distribution in Greece and Nature of Deposits
As noted above, ray-finned fishes, and teleosts in particular, are expected to have gradually expanded their occupation of most aquatic ecological niches on the planet, especially after the Paleozoic (Friedman and Sallan 2012). They are thus expected to be present in sedimentary rocks that were deposited in a large spectrum of paleoenvironments, ranging from offshore and deep marine waters, to fluvial, or lacustrine freshwaters. However, freshwater deposits (and included fossils) are more difficult to be preserved in the geological record, due to their relative ephemerality and patchiness. It is thus unsurprising that the vast majority of fossil ray-fins in Greece come from marine deposits. To paint a more complete picture of the Greek fossil record, locality information mined from gray literature (e.g., conference, abstracts field guides, and unpublished theses), stratigraphic and sedimentological works from the 1970s to the 2010s, or deriving from personal observations of the author and communication with colleagues is also taken into account. A brief summary of fossiliferous localities, ages, and paleoenvironments is given below, and more extensively in the Appendix; these lists might not be exhaustive.
Marine Occurrences
At least 33 indisputably marine, or likely marine localities have been recognized in Greece, although not all have been subjected to proper collecting and study (Fig. 2). The marine actinopterygian record of Greece begins with some poorly preserved †leptolepids from the Toarcian of the Ionian Unit, Leukada Island (Kottek 1964). Other published Mesozoic occurrences are restricted to the Cenomanian of Lindos, Rhodos Island (Melentis 1967a, b), and the terminal Maastrichtian of Pindos Unit, in Eurytania (Koch and Nikolaus 1969; Argyriou and Davesne 2021) and Gavdos Island (Cavin et al. 2012). Unpublished teleost remains are also present in Late Cretaceous exposures of Pindos Unit in Northern and central Peloponnese (T.A. personal observations and personal communication with E. Koskeridou, G. Theodorou and K. Trikolas).

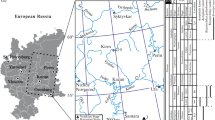
Map of the occurrences of fossil ray-finned fishes from Greece. See Appendix for further information. Marine occurrences : 1, Northern Milos (Pliocene–Pleistocene); 2, near Frangokastello (middle–late Pliocene); 3, Rouphas (Piacenzian, late Pliocene); 4, Stavromenos (Piacenzian); 5, Amnissos (Piacenzian); 6, Voutes (Piacenzian); 7, Prassas (Piacenzian); 8, Gournes (Piacenzian); 9, Aghia Marina (Piacenzian?,); 10, Aghios Thomas (Zanclean); 11, Faros, Aegina (Zanclean?, early Pliocene?); 12, Temple of Aphrodite, Aegina (Zanclean?, early Pliocene?); 13, Kalamaki (Pliocene); 14, Akropotamos (Messinian); 15, ‘Unit b’, Pigadia Basin (Messinian); 16, Tsangaraki (late Messinian); 17, Mires (late Messinian); 18, unknown locality near Rethymnon (Messinian?); 19, Psalida (Tortonian-early Messinian); 20, Metochia (Tortonian–Messinian); 21, Karave (Tortonian); 22, Aghia Paraskevi Chapel (Tortonian); 23, 400 m from Aghia Paraskevi Chapel (Tortonian); 24, type strata of Makrylia Fm. (Tortonian), 25, Keri (Tortonian); 26, unknown locality in Kefallonia (late Miocene?); 27, Aghioi Theodoroi (Paleocene); 28, Gavdos (Pindos unit; late Maastrichtian); 29, near Megalopoli (Maastrichtian); 30, Aghia Trias-Domnianoi Road (Maastrichtian); 31, near Karpenisi (late Maastrichtian); 32, Lindos (Cenomanian?); 33, Anavryssada (Toarcian). Brackish and potentially brackish occurrences: 34, Vatera (Pliocene–Pleistocene); 35, Voukolies (Messinian); 36, Kalergiana (Messinian); 37, Neo Khorio (Messinian); 38, Kymi (early Miocene?). Freshwater occurrences: 39, Marathousa 1 (middle Pleistocene); 40, Florina–Ptolemais–Servia Basin (Messinian–Zanclean); 41, Ano Metochi (Messinian); 42, Thymiana (Burdigalian-Langhian); 43, Lapsarna (Burdigalian); 44, Aidonochori. Uncertain environment: 45, ‘Unit e’, Pigadia Basin (middle Pleistocene), 46, Gryllos-Mesovouni (Pliocene-Pleistocene); 47, lignite mine in Strymonas valley (Pliocene). See Appendix for more information. Image exported from Google Earth Pro © 2019, map data from US Dept. of State Geographer, SIO, NOAA, U.S. Navy, NGA, GEBCO, image from Landsat/Copernicus. Scale bar equals 80 km, North faces upward
The Cenozoic record of Greece is very rich, but is largely skewed toward the late Miocene and Pliocene. The oldest reported Cenozoic occurrences come from flysch deposits of the Pindos Unit in Eurytania and are dated as Paleocene–early Eocene (Koch and Nikolaus 1969; Argyriou and Davesne 2021). Rich Tortonian marine assemblages can be found in the vicinity of Ierapetra and possibly Heraklion, Crete (Symeonidis 1969; Gaudant et al. 1997a; Gaudant 2004), and Gavdos Island (Gaudant et al. 2005), while a fish-bearing, bituminous horizon in Keri, Zakynthos has been identified, but not yet studied (Dermitzakis 1979; T.A. personal observations). Messinian fossiliferous deposits predating the Messinian Salinity Crisis are widespread in Crete, Gavdos (Gaudant 1980; Gaudant et al. 1997b, 2006) and Karpathos (Bellas et al. 1998), but only one poorly studied horizon is known in continental Greece, in Acropotamos, Kavala (Dermitzakis et al. 1986). Teleost teeth broadly ascribed to the late Miocene are also present in Kefalonia (pers. obs. on AMPG material; S. Roussiakis pers. comm.). The most notable early Pliocene occurrences come from the diatomitic horizons of Aegina Island, but despite their potential informativeness regarding the state of the Mediterranean in the immediate aftermath of the MSC, they remain understudied (Gaudant et al. 2010b; Argyriou and Theodorou 2011). The late Pliocene of Heraklion Basin, Crete, hosts exceptionally preserved and diverse ichthyofaunas (Gaudant et al. 1994; Gaudant 2001). Additional Pliocene occurrences have been reported in Rethymno, Chania, and Attiki regions (Gaudant and Symeonidis 1995; Gaudant 2001; Sorbini and Tyler 2004). Late Pliocene and Pleistocene fossiliferous horizons situated in Milos and Rhodos wait to be sampled and studied (Cornée et al. 2006; Calvo et al. 2012). Finally, wooden crates containing what appear to be unstudied marine teleost fossils from the Neogene are hosted in the collections of the AMPG (T.A. personal observations). Some of these fossils are wrapped in newspapers that treat the Russo-Japanese war of 1904–1905 as current news, but no locality or collection information has been retrieved so far.
Freshwater and Brackish Occurrences
Approximately six freshwater, and five brackish or potentially brackish occurrences have been reported. Burdigalian ichthyoliths from Lapsarna, Lesvos (Vasileiadou et al. 2017), and the early Miocene of SE Chios (Reichenbacher and Böhme 2004) constitute the oldest known freshwater fish assemblages in Greece. Ray-fin remains have been reported from the Messinian of Metochi, Serres (Georgalis et al. 2017); the Messinian–Zanclean of Florina-Ptolemais-Servia (Hordijk and de Bruijn 2009); and the Pleistocene of Marathousa, Megalopolis (Panagopoulou et al. 2018), but have not yet been studied.
The understudied assemblages from Kymi, Euboea, are tentatively dated as early Miocene, and tentatively interpreted as brackish, based on the supposed euryhaline affinities of recognized taxa (de Waldheim 1838, 1850; Gaudry 1862; Unger 1867; Woodward 1901; Kottis et al. 2002). Occurrences of †Aphanius crassicaudus in gypsiferous evaporitic deposits in Neo Khorio, Voukolies, and Kallergiana, Crete, have been identified as brackish and concurrent with the peak of the Messinian Salinity Crisis and the supposed desiccation of the Mediterranean in the late Messinian (Freuenthal 1969; Gaudant 1979, 2014b). However, there is evidence of primarily marine clupeids in the same beds, complicating this narrative (Gaudant 1980). The Pliocene clupeids from Serres Basin (Weiler 1943), and unidentified Plio-Pleistocene ray-fins from Vatera, Lesbos (Drinia et al. 2002), might also correspond to brackish, or freshwater deposits. New information on fresh or brackish waters of Greece is coming from the ongoing study of fossil clupeids from Aidonochori, Serres Basin (Kevrekidis 2016). Another possible addition to this list is the middle Pleistocene of Pigadia Basin in Karpathos Island. There, a vertebra possibly belonging to an ostariophysan—a group commonly found in freshwaters—has been found (Bellas et al. 1998).
5 Systematic Paleontology
Due to the taxonomic and phylogenetic breadth covered in this chapter, this section is organized in monophyletic (when possible) families. Synapomorphies for each family are not given, as this lies beyond the scope of this work. Instead, the reader is referred to Wiley and Johnson (2010), Nelson et al. (2016), and Betancur-R et al. (2017) for basic information on morphological synapomorphies and redirection to specialized literature. Taxonomic ranks superior to the genus level are unnamed, since they are obsolete in a modern phylogenetic context. Information on naming authorities and type genera and species is largely based on Wiley and Johnson (2010) and Eschmeyer and Fong (2019). The order of families and classification are based on the scheme proposed by Betancur-R et al. (2017), with fossil families being included according to their widely accepted position (see caption of Fig. 1). References regarding naming authorities, which are not also cited in the running text are not given in the reference list due to space constrains. Throughout the text and figures, extinct taxa are preceded by the dagger symbol ‘†’, following Patterson and Rosen (1977). Fossils that were not properly described and/or figured, or the geological context of which is problematic are only listed in the Appendix.
-
Actinopterygii sensu Goodrich, 1930
-
Neopterygii Regan, 1923
-
†Pycnodontiformes Berg, 1937
-
†Pycnodontidae Agassiz, 1833
Type Genus
†Pycnodus Agassiz, 1833.
-
† Coelodus Heckel 1854
Type Species
†Coelodus saturnus Heckel, 1854.
Nomenclatural and Taxonomical History
Initially described as †Coelodus muensteri (Melentis 1967a, b; Fig. 3a), but the specific attribution was subsequently challenged and removed (Schultz et al. 1997).

Distribution
In Greece, this taxon is known only from the Cenomanian of Lindos, Rhodos (Melentis 1967a, b). Outside Greece, this taxon has been reported from Europe and the Americas, but only its European occurrences have been revised. The latter include the Upper Jurassic–Cenomanian of UK and continental Europe, including the Balkans as well (Schultz et al. 1997).
Comments
This is a problematic occurrence, as the original locality and lithological context are conjecturally established. Prior to its study (Melentis 1967a, b), the †pycnodontid in question was part of the private collection of a cleric, who had in turn bought it from a fossil vendor, as evidenced by the transaction receipt that accompanies the specimen in the collections of the AMPG (S. Roussiakis pers. comm.; Fig. 3b). The fossiliferous facies that might have produced this fossil have not been since identified and explored for additional vertebrate remains. Therefore, the possibility that this specimen was not originally found in Rhodos, or even Greece, cannot be fully ruled out.
-
Teleostei Müller, 1846 (sensu Arratia 2017 )
-
†Leptolepididae Pictet, 1851
Type Genus
†Leptolepis Agassiz, 1832.
Nomenclatural and Taxonomical History
†Leptolepididae has a complicated taxonomic history and has been previously treated as a grade-like assemblage of primitive teleosts (Nybelin 1974; Arratia 1999, 2017; Arratia and Hikuroa 2010; Konwert and Stumpf 2017).
Distribution
In Greece, this taxon is known from the Toarcian of the Ionian Unit, and particular from Aghios Ilias, Anavryssada, Lefkada Island (Kottek 1964). Outside Greece, †Leptolepididae s.l. appeared in the Early Jurassic of Western and Central Europe and seem to have had achieved a cosmopolitan distribution by the Middle Jurassic (Arratia and Hikuroa 2010; Konwert and Stumpf 2017).
Comments
A partial maxilla and associated dentalosplenial of a †leptolepidid-like actinopterygian have been found in the Toarcian of Aghios Ilias (Anavryssada exposure), Lefkada, and have been identified as †Leptolepis cf. coryphaenoides (Kottek 1964). The material resembles †Leptolepis, but lacks any synapomorphies of the genus (Arratia 2017; Konwert and Stumpf 2017). This occurrence corresponds to the oldest record of actinopterygians in Greece, but should be treated as cf. †Leptolepididae indet. until more complete material becomes available.
-
† Ichthyodectiformes Bardack and Springle, 1969
-
†Ichthyodectoidei Romer, 1966
Distribution
In Greece, †Ichthyodectoidei have only been reported from the Maastrichtian (Late Cretaceous) of the Pindos Unit, Gavdos Island (Cavin et al. 2012). Outside Greece, †Ichthyodectiformes are found in Bathonian (Middle Jurassic)–Maastrichtian (Late Cretaceous) marine, and, rarely, in brackish and freshwater deposits of all continents (Arratia et al. 2004; Cavin et al. 2013).
Comments
This occurrence is based on a partially preserved postcranium from the Maastrichtian of Gavdos, which exhibits aspects of the caudal part of the vertebral column and associated dorsal, anal and caudal fins (Cavin et al. 2012). Similarities of the Gavdos specimen with †Saurodon elongatus from the Maastrichtian of Nardó, Italy, have been noted, but the specimen was left in open nomenclature due to its incomplete preservation (Cavin et al. 2012).
-
Clupeiformes Bleeker, 1859
-
Clupeidae Cuvier, 1817
-
Alosa Linck, 790
Type Species
Alosa alosa (Linnaeus, 1758).
Included Taxa
†Alosa crassa (Sauvage, 1873); Alosa tanaica (Grim, 1901); †Alosa elongata Agassiz, 1843.
Nomenclatural and Taxonomical History
†Alosa crassa is often included in the extant genus Sardina Antipa 1904, as †Sardina crassa, or †Sardina cf. crassa (Dermitzakis et al. 1986; Gaudant 2004, 2014b; Gaudant et al. 2005). The original generic attribution Alosa is herein maintained, following Grande (1985) and Carnevale (2007).
Distribution
In Greece, marine occurrences of †Alosa cf. crassa, or Alossa sp. are known from the Tortonian of the Makrilia Formation, in the vicinity of Ierapetra, Crete, the Messinian and Pliocene of Crete and the Tortonian–Messinian of Gavdos Island. Alosa elongata has been signaled in the Pliocene of Heraklion basin, in Crete. A. tanaica (first described as Caspialoassa nordmanni is known from the Pliocene of Strymonas valley in Serres (see Appendix for detailed information). Outside Greece, fossils affiliated with †Alosa crassa are common in the Meditteranean Neogene, with records in Algeria, Spain, Italy, and Cyprus (Sauvage 1873; Arambourg 1925, 1927; Gaudant et al. 2000, 2015; Landini and Sorbini 2005a; Carnevale 2007; Gaudant 2008a, b). Also, †Alosa elongata is known from the late Miocene of Algeria, Spain, Italy (Arambourg 1927; Landini and Sorbini 2005a; Gaudant 2008b; Gaudant et al. 2008, 2010a). Fossils attributed to A. tanaica have been reported from the Paratethyan Miocene (Paucǎ 1931; Rückert-Ülkümen 1994; Schwarzhans et al. 2017.
Comments
Most of the attributions to Alosa sp. refer to isolated scales, or bones of larger sized clupeids. The alosin from the Pliocene of Serres Basin, was originally attributed to Caspialosa nordmanni (Antipa 1904; Weiler 1943), which is considered a junior synonym of the Black Sea species A. tanaica (Grim 1901). This species is often mentioned as A. nordmanni (Schwarzhans et al. 2017) and, adding to the confusion, some of the contained fossils have been moved to †Alosa pinarhisariensis (Rückert-Ülkümen 1994). The specific attribution of the Serres alosin is ambiguous at best. However, future studies on newly discovered fossils from the late Miocene of Aidonochori, Serres Basin, might help shed some light on the complicated systematics of freshwater–brackish clupeids from the area (Kevrekidis 2016).
-
Sardina Antipa, 1904
Type Species
Sardina pilchardus (Walbaum, 1792). Only included species.
Distribution
In Greece, this taxon is known from the Pliocene of Heraklion Basin and Rethymno, Crete (see Appendix). Outside Greece, Sardina fossils are known from the Pliocene and Pleistocene of Italy (Landini and Sorbini 2005a). S. pilchardus is present in the Western Atlantic, the Mediterranean, and the Black and Azov seas (Froese and Pauly 2019).
Comments
Many of the Sardina sp., or Sardina? sp. occurrences in Greece correspond to poorly preserved, and/or disarticulated material, and might show taxonomic overlap with Alosa spp.
-
Spratelloides Bleeker, 1851
Type Species
Clupea argyrotaeniata (Bleeker, 1849) [= junior synonym of Spratelloides gracilis (Temminck and Schlegel, 1846)].
Included Taxa
S. gracilis (Temminck and Schlegel, 1846).
Distribution
In Greece, this taxon is known from the Tortonian of the Makrilia Formation, vicinity of Ierapetra, Crete, the Zanclean of Aegina Island, the Pliocene of Attiki, the Pliocene of Heraklion Basin and Rethymno, in Crete (Appendix and Fig. 4). Outside Greece, fossils affiliated with S. gracilis are known from the late Miocene, Pliocene and early Pleistocene of Algeria, Spain, and Italy (Arambourg 1927; Gaudant 2002, 2008b; Landini and Sorbini 2005a; Carnevale et al. 2006; Gaudant et al. 2015). Nowadays, S. gracilis is present in tropical waters of the Indo-Pacific Realm (Froese and Pauly 2019).

Spratelloides cf. gracilis (Clupeidae) from the Zanclean diatomitic marls near Mesagros, Aegina Island (AMPG collection). Scale bar equals 1 cm
-
Cypriniformes Bleeker, 1859
-
Cyprinidae Cuvier, 1817
Type Genus
Cyprinus Linnaeus, 1758.
-
† Palaeoleuciscus Obrhelová, 1967
Type Species
†Palaeoleuciscus chartaceus (Laube, 1905).
Distribution
In Greece, this taxon is known only from the early Miocene of Lapsarna Locality, Lesvos Island, Greece (Vasileiadou et al. 2017; Appendix). Outside Greece, its oldest fossils come from the Oligocene of Germany and Switzerland (Cavender 1991; Böhme and Ilg 2003). Τhere numerous records of tooth and skeletal remains of the genus in the Miocene of Europe and Anatolia as well (e.g., Cavender 1991; Böhme and Ilg 2003; Rückert-Ülkümen and Yiğitbaş 2007; Böhme 2010; Gaudant 2014a, 2015).
-
Barbus Cuvier and Cloquet, 1816
Type Species
Barbus barbus (Linnaeus, 1758).
Distribution
In Greece, this taxon is only known from the early Miocene of Lapsarna Locality, Lesvos Island, Greece (Vasileiadou et al. 2017; Appendix). Outside Greece, Barbus-like (?Barbus sp.) pharyngeal teeth first appear in the Oligocene of Afroarabia (Otero and Gayet 2001). Barbus remains are quite common in Neogene and younger freshwater deposits in Europe, Africa, the Arabian subcontinent, and Anatolia (Cavender 1991; Stewart 2001, 2003; Böhme and Ilg 2003; Přikryl et al. 2016b; Stewart and Murray 2017). The distribution of modern Barbus s.s. species ranges from Europe to the Caspian Sea, Anatolia, and Tigris and Euphrates drainages, with approx. 13 species found in Greek freshwaters (Froese and Pauly 2019).
Comments
It is widely accepted that modern and fossil species lumped under the genus Barbus correspond to different lineages, rendering the genus paraphyletic (Berrebi and Tsigenopoulos 2003; Kottelat and Freyhof 2007). Barbus s.l. is used to indicate uncertainty relative to the placement of fossils relative to modern lineages, whereas Barbus s.s. indicates affinities with the European and Anatolian subgenus Barbus. Isolated pharyngeal teeth or bones are difficult to attribute to specific lineages, yet similarities of the Lapsarna Barbus with Barbus s.s. have been noted (Vasileiadou et al. 2017).
-
Cyprinion Heckel, 1843
Type Species
Cyprinion macrostomus Heckel, 1843.
Distribution
In Greece, pharyngeal bones and teeth of Cyprinion have been found in the early Miocene of Lapsarna Locality, Lesvos Island, Greece (Vasileiadou et al. 2017; Appendix). Outside Greece, pharyngeal teeth putatively attributed to the genus have also been found in the early Miocene of Turkey (Böhme and Ilg 2003). Modern representatives of Cyprinion have a distribution that ranges from the Arabian subcontinent to Myanmar (Froese and Pauly 2019).
Comments
According to Vasileiadou et al. (2017), the Lapsarna material likely represents a new species, which has not yet been described formally.
-
Aulopiformes Rosen, 1973
-
†Dercetidae Woodward, 1901
Type Species
†Dercetis scutatus (Agassiz, 1834).
Distribution
In Greece, fossils of †Dercetidae are so far known from the Late Maastrichtian of Pindos Unit, in the vicinity of Karpenisi, Eurytania (Koch and Nikolaus 1969). Outside Greece, †dercetids are known from the Late Cretaceous–Paleocene of Europe, the Middle East, North and Central Africa, and South America (summarized in da Silva and Gallo 2011, 2016; Vernygora et al. 2018).
Comments
The †dercetids found in the vicinity of Karpenisi have been tentatively ascribed to either the aulopiform †Rhynchodercetis or, erroneously, the stem teleost †Belonostomus (Koch and Nikolaus 1969). Only one †dercetid-like elongate lower jaw, bearing numerous minute teeth, has been figured, but this material has not been described or treated systematically (Koch and Nikolaus 1969, Table 33:3), and bears no apomorphies to support any attribution below the Family-level. No repository information has been provided for these fossils.
-
†Enchodontidae Woodward, 1901
Type Genus
†Enchodus Agassiz, 1835.
-
† Enchodus Agassiz, 1835
Type Species
†Enchodus lewesiensis (Mantell, 1822).
Distribution
In Greece, fossils of †Enchodus cf. dirus are known from the late Maastrichtian, Pindos Unit, Gavdos Island (Cavin et al. 2012). Additional †enchodontid remains, possibly referable to †Enchodus, have been collected from Maastrichtian deposits of the Pindos Unit, exposed on the road from Aghia Triada to Domniani, near Karpenisi, Eurytania (Koch and Nikolaus 1969). Outside Greece, the oldest disarticulated remains referred to †Enchodus sp. have been collected in the Early Cretaceous of Spain (Kriwet 2003), while articulated remains are known from the Albian–Cenomanian of Mexico (Fielitz and González-Rodríguez 2010). This genus achieved a cosmopolitan distribution during the Late Cretaceous, with occurrences in North and South America, England and western continental Europe, Africa, India, and the Middle East (summarized in Goody 1969; Chalifa 1996; Cavin et al. 2012; da Silva and Gallo 2016; Holloway et al. 2017).
Comments
A relatively well-preserved skull of †Enchodus, also exhibiting features of the palate and jaws, was found in the late Maastrichtian deposits of Gavdos Island and was attributed to a juvenile individual (Cavin et al. 2012). The specimen was included in a phylogenetic analysis alongside other †enchodontids, and was resolved as sister taxon to †Enchodus dirus (Cavin et al. 2012), which, in turn, is found in several Late Cretaceous localities of the Western Interior Seaway, North America (Goody 1969). The Gavdos and North American specimens seem to only differ in size and ornamentation, and their seemingly close relationship supports complex biogeographical scenarios, involving the ability of, at least some, Late Cretaceous †enchodontids to cross oceanic basins (Cavin et al. 2012).
-
Paralepididae Gill, 1872
Type Genus
Paralepis Cuvier, 1817.
-
Paralepis Cuvier, 1817
Type Species
Paralepis coregonoides Risso, 1820.
Included Taxa
†Paralepis albyi (Sauvage, 1870).
Distribution
In Greece, this taxon is known from the Messinian of Gavdos (Metochia: †P. albyi; Gaudant et al. 2006). Outside Greece, †P. albyi is present in the late Miocene of Spain and Italy (Arambourg 1925; Gaudant 2002; Carnevale 2007). Two species of the deep-water genus Paralepis are nowadays present in the Mediterranean (Froese and Pauly 2019).
-
Stomiatiformes sensu Rosen, 1973
-
Gonostomatidae Gill, 1893
Type Genus
Cyclothone Goode and Bean, 1883.
-
Cyclothone Goode and Bean, 1883
Type Species
Cyclothone acclinidens Garman, 1899.
Included Taxa
†Cyclothone gaudanti Přikryl and Carnevale, 2017.
Remarks
The modern species Cyclothone braueri, Cyclothone microdon, and Cyclothone pygmaea have been reported from the Italian Pliocene and Pleistocene (Landini and Sorbini 2005a; Přikryl and Carnevale 2017). The genus has a cosmopolitan distribution in modern deep marine waters (Froese and Pauly 2019).
-
★†Cyclothone gaudanti Přikryl and Carnevale, 2017
Nomenclatural and Taxonomical History
Cyclothone sp. in Gaudant 2004 (initial identification); Cyclothone gaudanti Přikryl and Carnevale, 2017 (new species).
Type Material
NHMW 1999z0042/0020, nearly complete, articulated individual.
Type Locality
Locality 400 m to the North of Aghia Paraskevi Chapel, vicinity of Ierapetra, Crete, Greece, Makrilia Formation.
Distribution
This species is only known from the Tortonian of Makrilia Formation, in the vicinity of Ierapetra, Crete (Aghia Paraskevi Chapel Locality, and from a locality 400 m to the North of Aghia Paraskevi Chapel; Gaudant 2004, Přikryl and Carnevale 2017). These localities constitute the whole range of this taxon.
Remarks
Cyclothone gaudanti demonstrates affinities with the small-sized and lightly pigmented species of the genus, such as Cyclothone alba, C. braueri, and Cyclothone signata (Přikryl and Carnevale 2017). Based on its affinities and the depositional environment of Makrilia Formation, it is estimated that this species possibly occupied the upper mesopelagic layer (Přikryl and Carnevale 2017).
-
Sternoptychidae Duméril, 1806
Type Genus
Sternoptyx Hermann, 1781.
-
Maurolicus Cocco, 1838
Type Species
Maurolicus muelleri (Gmelin, 1789). Only included species.
Distribution
In Greece, this taxon is known with the species M. muelleri from the Tortonian of the Makrilia Formation, in the vicinity of Ierapetra, Crete (Gaudant 2004), the Messinian of Gavdos (Gaudant et al. 2006), and the Messinian of Mires (Gaudant et al. 1997a). Furthermore, Maurolicus cf. M. muelleri has been reported from the Pliocene of Heraklion as well (Gaudant 2001; Appendix). Outside Greece, M. muelleri has a continuous presence in the Mediterranean Realm (including Algeria, Italy and Spain) since the late Miocene (Gaudant 2002, 2008b; Landini and Sorbini 2005a; Gaudant et al. 2008, 2010a), and is characterized by a cosmopolitan distribution in modern bathypelagic waters (Froese and Pauly 2019).
-
Argyropelecus Cocco, 1829
Type Species
Argyropelecus hemigymnus Cocco, 1829.
Distribution
In Greece, the only record of Argyropelecus sp. comes from the Pliocene (Zanclean) of Aegina Island (Argyriou and Theodorou 2011), and is figured here for the first time (Fig. 5). Outside Greece, the fossil record of the genus dates back to the Oligocene of Romania, Poland, and the Caucasus (summarized in Carnevale 2003).

Argyropelecus sp. (Sternoptychidae), from the Zanclean diatomitic marls near Mesagros, Aegina Island (AMPG collection). This specimen is figured here for the first time. Scale bar equals 1 cm
Remarks
So far, only two species have been recognized on the basis of body fossils in the Mediterranean region: the late Miocene †A. logearti in Algeria (Arambourg 1927; Carnevale 2003), and the extant A. hemigymnus in the Pliocene and Pleistocene of Italy (Carnevale 2003, 2007; Landini and Sorbini 2005a).
-
Myctophiformes Regan, 1911
-
Myctophidae Gill, 1893
Type Genus
Myctophum Rafinesque, 1810a.
-
Myctophum Rafinesque, 1810a
Type Species
Myctophum punctatum Rafinesque, 1810b.
Included Taxa
†“Myctophum” licatae Sauvage, 1870; †“Myctophum” dorsale Sauvage, 1870.
Distribution
In Greece, myctophid body fossils are known from the Tortonian of Makrilia Formation, vicinity of Ierapetra (Gaudant 2004), whereas occurrences identified to the species level are known from the Tortonian–Messinian of Messara, Heraklion (Psalidha, Mires: †“Myctophum” licatae, †“Myctophum” dorsale; Tsangaraki: †“Myctophum” licatae; Gaudant et al. 1997a), and the Tortonian (Metochia, Karave: †“Myctophum” licatae) and Messinian (Metochia: †M. licatae, †M. dorsale; Fig. 5) of Gavdos (Gaudant et al. 2005, 2006). A single myctophid tooth was also recovered from the Messinian strata of ‘Unit b’ of Pigadia Basin in Karpathos (Bellas et al. 1998). Outside Greece, these two Miocene species have been recognized in several Mediterranean localities in Algeria, Spain, Italy, and Cyprus (Arambourg 1925, 1927; Gaudant 2002, 2008a, b; Merle et al. 2002; Gaudant et al. 2010a). Several extant species of Myctophum have been reported from Pliocene and Pleistocene localities of the Mediterranean (Landini and Sorbini 2005a). Nowadays, Myctophum species have a cosmopolitan bathypelagic distribution (Froese and Pauly 2019).
Remarks
Greek body fossils of Myctophidae were lumped by Gaudant in the extant genus Myctophum, following his taxonomic reappraisal of material previously studied by Arambourg (1925), from the late Miocene of Licata, Sicily (Gaudant and Ambroise 1999). However, Gaudant’s taxonomical scheme relied heavily on putatively overlapping ratios of body measurements and the overall presence or absence of photophores, and did not take important anatomical characters of synapomorphical value into proper account (see e.g., Paxton 1972; Carnevale 2007; Denton 2013). Detailed considerations of fossil myctophids from e.g., the Serravalian of Italy revealed the simultaneous presence of multiple genera—but not Myctophum—in the Mediterranean Neogene (Carnevale 2007), signaling the necessity for future revision of fossil myctophid diversity in the entirety of the Mediterranean Basin, including Greece. Given the scope of the present work, I refrain from reappraising Greek myctophid fossils and retain Gaudant’s classification in inverted commas (also employed by Gaudant in his later works). New material from Metochia C, Gavdos, collected by the author (e.g., Fig. 6), preserves delicate anatomical structures that can facilitate relevant undertakings in the future.

Myctophidae indet. From the Messinian of Metochia C, Gavdos Island (AMPG collection; photograph provided by JD Carrillo). Scale bar equals 1 cm
-
Gadiformes sensu Endo, 2002
-
Gadidae Rafinesque, 1810b
Type Genus
Gadus Linnaeus, 1758.
-
Micromesistius Gill, 1863
Type Species
Micromesistius poutassou (Risso, 1827).
Distribution
In Greece , the only fossil occurrence of Micromesistius sp. comes from the Pliocene Amnissos, Heraklion Basin (Gaudant 2001). Outside Greece, this taxon is known from the Pliocene to Pleistocene of Italy (Landini and Sorbini 2005a). The genus is nowadays present in temperate Atlantic and Pacific waters, including the Mediterranean (Froese and Pauly 2019).
-
Moridae Goode and Bean, 1896
Type Genus
Mora Risso, 1827.
-
Gadella Lowe, 1843
Type Species
Gadella gracilis Lowe, 1843.
Distribution
In Greece, occurrences of Gadella are known from the Pliocene of Attiki (Gaudant and Symeonidis 1995) and Heraklion Basin (Gaudant 2001). Outside Greece, this taxon is known from the Pliocene of Italy (Landini and Sorbini 2005a). Gadella is still present in tropical to subtropical deep waters around the world, including the Mediterranean (Froese and Pauly 2019).
-
Bregmacerotidae Gill, 1872
Type Genus
Bregmaceros Thompson, 1840.
-
Bregmaceros Thompson, 1840
Type Species
Bregmaceros mcclellandi Thompson, 1840.
-
†Bregmaceros albyi (Sauvage, 1880)
Distribution
This is one of the most widely distributed taxa in Greece, with occurrences from the Tortonian of Makrilia Formation, vicinity of Ierapetra, Crete (Symeonidis 1969; Bürgin 1994; Gaudant 2004), the Tortonian and Messinian of Metochia, Gavdos (Gaudant et al. 2005, 2006), the Tortonian–early Messinian of Messara, Heraklion, Crete (Gaudant et al. 1997a), the Messinian of Karpathos (Bellas et al. 1998), the Pliocene (cf. Zanclean) of Aegina Island (Argyriou and Theodorou 2011; Fig. 7), the Pliocene of Heraklion (Gaudant et al. 1994; Müller and Strauch 1994; Gaudant 2001) and the Piacenzian of Rethymno (Symeonidis and Schultz 1973; see Appendix for more information). Outside Greece, body fossils of †Bregmaceros albyi are known from the Miocene of Algeria, Italy, Malta, and Spain, and from the Pliocene of Italy and, possibly (as †B. cf. albyi), Cyprus (Gaudant 2002; Landini and Sorbini 2005a; Přikryl et al. 2016a; and references therein). The species is also present in the early Pleistocene of Italy (Landini and Sorbini 2005a).

Bregmaceros cf. albyi (Bregmacerotidae), from the Zanclean diatomitic marls near Mesagros, Aegina Island. This specimen is figured here for the first time (AMPG collection). Scale bars equal 1 cm
-
Acanthomorpha Rosen, 1973
-
Atheriniformes sensu Dyer and Chernoff, 1996
-
Atherinidae Risso, 1827
Type Genus
Atherina Linnaeus, 1758.
Distribution
In Greece, atherinids are known only from the Pliocene of Attiki (Gaudant and Symeonidis 1995). Outside Greece, atherinids are known to have been present in Tethys since at least the Eocene of Monte Bolca, Italy (Carnevale et al. 2014), but their more recent fossil record in the area is largely composed of otoliths (Landini and Sorbini 2005a). Body fossils of atherinids are known from the Miocene of the Paratethys (Schwarzhans et al. 2017) and California (Jordan and Gilbert 1919).
-
Beloniformes Berg, 1937
-
Belonidae Bonaparte, 1832
Type Genus
Belone Cuvier, 1817.
-
Belone Cuvier, 1817
Type Species
Belone belone (Linnaeus, 1760).
Distribution
In Greece, this taxon is only known from the Pliocene of Heraklion Basin (Gaudant et al. 1994; Gaudant 2001). Outside Greece, fossils of this taxon are more common, coming from the Oligocene of the Caucasus (Smirnov 1936), the Miocene of the USA (de Sant’Anna et al. 2013), and the Miocene to Pleistocene of Italy (Arambourg 1925; Landini and Sorbini 2005a). The genus is nowadays present in the Atlantic Ocean, the Mediterranean, and the Black Sea (Froese and Pauly 2019).
-
Scomberesox Lacepède, 1803
Type Species
Scomberesox saurus (Walbaum, 1792).
Distribution
In Greece, the only possible fossils of this genus come from the Pliocene of Heraklion Basin (Gaudant 2001). Outside Greece, fossils of Scomberesox are known from the Miocene of USA, Algeria, and Italy, and the Pleistocene of Italy (Jordan and Gilbert 1919; Gaudant 2002, 2008b; Landini and Sorbini 2005a). Nowadays, the genus has a circumglobal distribution, which also includes the Mediterranean (Froese and Pauly 2019).
-
Blenniiformes sensu Springer, 1993
-
Blenniidae Rafinesque, 1810a
Type Genus
Blennius Linnaeus, 1758.
-
Salaria Forskål, 1775
Type Species
Not designated.
Included Taxa
Salaria pavo (Risso, 1810).
Nomenclatural and Taxonomical History
Initially ascribed by Gaudant et al. (1994) to the genus Blennius, as Blennius cf. B. pavo (Risso 1810). This species is now included in the genus Salaria, as S. pavo (see Eschmeyer and Fong 2019).
Distribution
In Greece, this taxon is known only from the Pliocene of Heraklion Basin (Gaudant et al. 1994). So far no additional body-fossil records exist for this taxon. Nowadays, S. pavo is present in shallow marine to brackish waters along the Western Atlantic, Mediterranean, and Black Sea coasts (Froese and Pauly 2019).
-
Perciformes sensu Betancur-R et al. 2017
-
Serranidae Swainson, 1839
Type Genus
Serranus Cuvier, 1817.
-
Serranus Cuvier, 1817
Type Species
Serranus cabrilla (Linnaeus, 1758).
Distribution
In Greece , this taxon is certainly known from the Pliocene of Heraklion Basin (Gaudant 2001), while the presence of the family has been signaled in the Pliocene of Aegina Island as well (Gaudant et al. 2010b). Outside Greece, Serranus fossils are known from the Miocene of Algeria, and the Plio-Pleistocene of Sardinia (Abbazzi et al. 2004; Gaudant 2008b). Serranus has a modern cosmopolitan, Tropical–Temperate, marine distribution (Froese and Pauly 2019).
-
Cyprinodontiformes sensu Parenti, 1981
-
Cyprinodontidae Gill, 1865
Type Genus
Cyprinodon Lacepède, 1803.
-
Aphanius Nardo, 1827
Type Species
Aphanius nanus Nardo, 1827.
Included Taxa
†A. crassicaudus Agassiz, 1839.
Nomenclatural and Taxonomical History
Initially included in the genus †Lebias (Agassiz, 1833–1843), as †L. crassicaudus (Raulin 1861). Woodward erected the genus Pachylebias to accommodate pachyostotic cyprinodontiform material from the Messinian of Italy and Crete (Woodward 1901). †Pachylebias was later found to be a junior synonym of the extant genus Aphanius (Gaudant 1979).
Distribution
In Greece, this taxon is known from the Messinian of Chania, Crete (Raulin 1861; Woodward 1901; Freuenthal 1969; Gaudant 1979, 2014b). Outside Greece, Aphanius is an essential component of Messinian oligohaline fish assemblages of the Mediterranean, including occurrences in Algeria, Spain, Morocco, and Italy (Gaudant 2002; Carnevale et al. 2018).
-
Scombriformes sensu Betancur-R et al. 2017
-
Scombridae Rafinesque, 1815
Type Genus
Scomber Linnaeus, 1758.
-
Scomber Linnaeus, 1758
Type Species
Scomber scombrus Linnaeus, 1758 (= S. scomber L., 1758).
Distribution
In Greece, this genus is only known from the Pliocene of Heraklion Basin (Gaudant 2001). Outside Greece, the genus is known from the Oligocene of central Europe and the Caucasus, and the Neogene of Russia, the United States, Japan, Algeria, Italy, Croatia, and Serbia (Arambourg 1927; Yabumoto and Uyeno 1994; Landini and Sorbini 2005a; Micklich and Hildenbrandt 2010; Gregorová 2011; Nazarkin and Bannikov 2014). Modern representatives of the genus are widespread in tropical to temperate waters (Froese and Pauly 2019).
-
Scomberomorus Lacepède, 1801
Type Species
Scomberomorus regalis (Bloch, 1793) (= S. plumierii Lacepède, 1801).
Distribution
In Greece, this genus is only known from the Pliocene of Heraklion Basin (Gaudant 2001). Outside Greece, the genus appears in the Paleocene of Turkmenistan (Monsch and Bannikov 2012). It is also known the Paleogene of Kazakhstan, Georgia, the United States, Germany, and the Neogene of Japan, Poland, and the United States (Schultz 1979; Yabumoto and Uyeno 1994; Weems 1999; Micklich and Hildenbrandt 2010; Monsch and Bannikov 2012; Weems et al. 2017). The genus is found today in tropical-subtropical waters of the Atlantic, Indian, and Pacific oceans, and the Western Mediterranean (Froese and Pauly 2019).
-
Trichiuridae Rafinesque, 1810a
Type Genus
Trichiurus Linnaeus, 1758.
-
Lepidopus Goüan, 1770
Type Species
Lepidopus caudatus Euphrasen, 1788.
Included Taxa
†L. albyi Sauvage, 1870.
Distribution
In Greece, this genus is known from the Tortonian of Makrilia Formation, in the vicinity of Ierapetra, Crete (Symeonidis 1969; Gaudant 2004), the Tortonian and Messinian of Gavdos (Gaudant et al. 2005), the Messinian of Messara (as †Lepidopus albyi in the text, but Lepidopus sp. in the respective figure caption; Gaudant et al. 1997a), and the Pliocene of Heraklion Basin and Rethymno (Gaudant et al. 1994; Gaudant 2001; see Appendix for further information). Outside Greece, †Lepidopus albyi is known from the late Miocene of Algeria, Spain, Italy, Cyprus, and Pliocene of Italy (Arambourg 1925, 1927; Gaudant 2002; Merle et al. 2002; Landini and Sorbini 2005a). A second fossil species, †Lepidopus proargenteus, is known from the late Miocene of Algeria and Italy (Arambourg 1927; Gaudant 2008b; Gaudant et al. 2010a). Remains affiliated with the living L. caudatus have been reported from the Pliocene of Italy (Landini and Sorbini 2005a).
Remarks
Symeonidis tentatively ascribed fragmentary remains of a juvenile Lepidopus from the Tortonian of Crete to the Oligocene Paratethyan species †Anenchelum glaricianum (his †Lepidopus cf. glaricianus, Symeonidis 1969). Gaudant revised this attribution to Lepidopus sp. (Gaudant 2004).
-
Gobiiformes Günther, 1880
-
Gobiidae Cuvier, 1817
Type Genus
Gobius Linnaeus, 1758.
-
Gobius Linnaeus, 1758
Type Species
Gobius niger Linnaeus, 1758.
Distribution
In Greece, unidentified records of this genus are reported from the Pliocene of Heraklion Basin (Gaudant et al. 1994) and Attiki (Gaudant and Symeonidis 1995). Outside Greece, Gobius skeletons first appear in the early Miocene of the Czech Republic (Reichenbacher et al. 2018), but are found in a wide array of Neogene fossil sites in North Africa, Europe, and Asia Minor (Arambourg 1927; Gaudant 2002; Böhme and Ilg 2003; Reichenbacher et al. 2018). Gobius has a circumglobal modern distribution (Froese and Pauly 2019).
Comments
Gobioids are among the largest clades of vertebrates (Nelson et al. 2016). In many cases, included genera and species are diagnosed on the basis of very delicate anatomical features and soft tissue structures, which are not often visible/preserved in fossils (Reichenbacher et al. 2018). Thus, Greek gobiid fossils only loosely affiliate with Gobius (Gaudant et al. 1994; Gaudant and Symeonidis 1995), and might actually encompass other genera as well. It should be noted here that poorly documented occurrences of Gobiidae in the Miocene of Kymi have been ascribed to the genus Periophthalmus (Unger 1867; Appendix), but, due to lack of descriptive or pictorial documentation, these are not further treated here.
-
Carangiformes sensu Betancur-R et al. 2017
-
Carangidae Rafinesque, 1815
Type Genus
Caranx Lacepède, 1801.
-
Caranx Lacepède, 1801
Type Species
Caranx hippos (Linnaeus, 1766).
Distribution
In Greece, this genus is only known from the Pliocene of Heraklion Basin (Gaudant 2001). Outside Greece, this taxon is known from the Miocene of Algeria, Italy (Arambourg 1925, 1927), but also from the Paratethys (Schwarzhans et al. 2017).
-
Alectis Rafinesque, 1815
Type Species
Alectis ciliaris (Bloch, 1787).
-
★†Alectis psarianosi (Symeonidis, 1969)
Nomenclatural and Taxonomical History
This species , based on a single specimen from the Tortonian of Crete, was formerly included in the genus Mene (Menidae), as †Mene psarianosi (Symeonidis 1969). It was later reattributed to the carangid genus Alectis, but the species name was retained (Bannikov 1984, 1987; Gaudant 2004).
Type Material
AMPG 185/1969, complete, laterally compressed individual (Fig. 8).

The type specimen (AMPG 185/1969) of †Alectis psarianosi (Carangidae) from the Tortonian of Aghia Paraskevi Chapel, vicinity of Ierapetra, Crete. Scale bars equal 1 cm
Type Locality
Tortonian of Makrilia Formation, vicinity of Ierapetra, Crete (Aghia Paraskevi Chapel Locality; Symeonidis 1969; Gaudant 2004).
Distribution
Known only from the type locality.
Remarks
Bannikov (1987) dispelled the original, poorly supported attribution of this unique fossil to Menidae by Symeonidis (1969), after recognizing typical carangid features, including a high rhomboidal body shape ; a well-developed fronto-occipital crest; and the relative length of fin rays in its median fins. Gaudant (2004) re-examined the specimen, providing its first detailed anatomical description and confirming Bannikov’s assessment, but without attempting additional comparisons. According to Bannikov (1987), this fossil species differs from modern species in having “somewhat shorter anterior rays of the soft unpaired fins.” Whether this difference is sufficient to maintain †Alectis psarianosi as a valid fossil species remains to be investigated in the future , given the fact that the fossil corresponds to a somewhat small, possibly immature, individual.
-
Trachurus Rafinesque, 1810a
Type Species
Trachurus trachurus (Linnaeus, 1758) (= T. saurus Rafinesque, 1810b).
Distribution
In Greece, occurrence of this genus comes from the Tortonian of Makrilia Formation, in the vicinity of Ierapetra, Crete (Gaudant 2004), the Pliocene of Attiki (Gaudant and Symeonidis 1995), and the Pliocene of Heraklion (Gaudant 2001; see Appendix for further information). Outside Greece, fossils more or less securely attributed to Trachurus are known from the Miocene of Algeria and Spain, the Miocene and Pliocene of Italy and the Pliocene of Cyprus (Arambourg 1927; Landini and Sorbini 2005a; Gaudant 2008a, b; Gaudant et al. 2010a; Gaudant and Courme 2014). Three species of Trachurus are native to modern Mediterranean pelagic habitats (Froese and Pauly 2019).
-
Spariformes sensu Akazaki, 1962
-
Sparidae Rafinesque, 1810a
Type Genus
Sparus Linnaeus, 1758.
-
Boops Cuvier, 1814
Type Species
Boops boops (Linnaeus, 1758).
Distribution
In Greece, this genus is known from the Pliocene of Heraklion Basin with various occurrences (Gaudant et al. 1994; Gaudant 2001). Outside Greece, Boops fossils have been found in the Miocene of Algeria and the Miocene and Pliocene of Italy (Arambourg 1927; Gaudant 2002). Nowadays, Boops is represented by two species, one of which, B. boops, occurs in the Mediterranean (Froese and Pauly 2019).
Comments
Gaudant et al. (1994) initially ascribed the Amnissos boopsins to Boops cf. B. boops . In a subsequent work, Gaudant (2001) noted differences in meristic features of the dorsal and caudal fins of the Boops from Amnissos, from both the Miocene species †B. roulei (Arambourg 1927), and the extant B. boops, leaving the Cretan fossils in open nomenclature, and implying that they might correspond to a new species. This might also be true for the remaining Heraklion Basin boopsins. As evidenced by isolated teeth, additional sparids were likely present in the Messinian of Karpathos (Bellas et al. 1998).
-
Centriscidae Bonaparte, 1831
Type Genus
Centriscus Linnaeus, 1758.
-
Aeoliscus Jordan and Starks, 1902
Type Species
Aeoliscus strigatus (Günther, 1861).
Nomenclatural and Taxonomical History
Centriscids from the Pliocene of Amnissos were initially thought to correspond to a new fossil species (†Amphisile cretensis Bachmayer et al. 1984). Gaudant re-examined these specimens and tentatively ascribed these fossils to the extant genus Amphisile (Cuvier 1817), as Amphisile cf. strigata (Gaudant 2001). Amphihisile strigata is, however, a synonym of Aeoliscus. strigatus (Günther 1861. In a subsequent work, Gaudant corrected the taxonomic designation of the Cretan material to Aeoliscus cf. strigatus (Gaudant and Courme 2014).
Distribution
In Greece, this taxon is known from the Pliocene of Heraklion Basin, (Bachmayer et al. 1984; Gaudant 2001). Outside Greece, Aeoliscus. strigatus is also known from the late Miocene and Pliocene of Italy and the Pliocene of Cyprus (Gaudant 2002; Landini and Sorbini 2005a; Gaudant and Courme 2014). This species is nowadays encountered in tropical Indo-Pacific waters (Froese and Pauly 2019).
Remarks
Given the invalidity of the genus Amphisile (see above), the occurrence of Amphisile sp. in the Messinian of Gavdos (Gaudant et al. 2006) is hereby treated conservatively as Centriscidae indet.
-
Syngnathidae Rafinesque, 1810a
Type Genus
Syngnathus Linnaeus, 1758.
-
Syngnathus Linnaeus, 1758
Type Species
Syngnathus acus Linnaeus, 1758.
Included Taxa
†Syngnathus albyi Sauvage, 1870.
Nomenclatural and Taxonomical History
Two fossil species of Syngnathus have been erected on the basis of material from the Neogene of Crete: the late Miocene †S. kaehsbaueri Bachmayer and Symeonidis (1978) from Aghia Paraskevi Chapel, Makrilia Formation, in the vicinity of Ierapetra, and the Pliocene ★†S. heraklionis Bachmayer et al., 1984 from Amnissos, near Heraklion. Reinvestigations of the type material of both species found no anatomical differences from the Miocene species †S. albyi (Sauvage 1870), establishing the Cretan species as invalid and referred their type material to †Syngnathus cf. †S. albyi (Gaudant 2001, 2004).
Distribution
In Greece, occurrences of this taxon are known from the Tortonian of Makrilia Formation, near Ierapetra, Crete (Bachmayer and Symeonidis 1978; Gaudant 2004), the Tortonian and Messinian of Metochia, Gavdos (Gaudant et al. 2005, 2006), the Messinian of Messara, Crete (Gaudant et al. 1997a), and the Pliocene of Heraklion, Crete (Bachmayer et al. 1984; Gaudant et al. 1994; Gaudant 2001). Remains of short-snouted Syngnathus cf. S. acus were also found in the Pliocene (cf. Zanclean) of Aegina (Argyriou and Theodorou 2011, Fig. 9). Outside Greece, the fossil species †S. albyi is common in “middle” and late Miocene and possibly Pliocene faunas of Algeria, Spain, Italy, and Cyprus (Arambourg 1925, 1927; Gaudant et al. 2000, 2010a; Gaudant 2002, 2008b; Merle et al. 2002; Landini and Sorbini 2005a; Carnevale 2007), whereas records of this species in the Miocene of Paratethys are considered dubious (Schwarzhans et al. 2017). The modern species S. acus and S. phlegon have been reported from the late Pliocene and Pleistocene of Italy (Landini and Sorbini 2005a).

Syngnathus cf. acus (Syngnathidae), from the Zanclean diatomitic marls near Mesagros, Aegina Island (AMPG collection). This specimen is figured here for the first time. Scale bar equals 1 cm
-
Dactylopteridae Gill, 1861
Type Genus
Dactylopterus Lacepède, 1801.
-
Dactylopterus Lacepède, 1801
Type Species
Dactylopterus volitans (Linnaeus, 1758).
Distribution
In Greece, this taxon is only known from the Pliocene of Heraklion Basin. Outside Greece, it is known from the Pliocene of Italy and Cyprus (Landini and Sorbini 2005a; Gaudant and Courme 2014). One species survives in the Atlantic and the Mediterranean (Froese and Pauly 2019).
-
Pleuronectiformes Bleeker, 1859
-
Bothidae Smitt, 1892
Type Genus
Bothus Rafinesque, 1810a.
-
Arnoglossus Bleeker, 1862
Type Species
Pleuronectes arnoglossus (Bloch and Schneider, 1801) (junior synonym of Arnoglossus laterna Walbaum, 1792).
Distribution
In Greece, this taxon is only known from the Tortonian of Makrilia Formation, in the vicinity of Ierapetra, Crete (Gaudant 2004). Outside Greece, Arnoglossus is reported in the Miocene of Algeria and Spain, and from the Miocene, Pliocene, and Pleistocene of Italy (Landini and Sorbini 2005a; Gaudant 2008a, b; Gaudant et al. 2010a). Arnoglossus has a cosmopolitan modern distribution (Froese and Pauly 2019).
-
Centropomidae sensu Greenwood, 1976
Type Genus
Not established.
-
Latinae indet.
Nomenclatural and Taxonomical History
This fossil was initially employed to establish the genus †Allocotus by de Waldheim (1838). In a following publication, de Waldheim, erected a new genus and species, †Platacanthus ubinoi, to accommodate the same individual (de Waldheim 1850). The Musée de la Société Impériale des Naturalistes, Moscow, was listed as repository of this specimen (Fig. 9a). Woodward reassigned †P. ubinoi to the modern genus Morone (his Labrax), utilizing additional material from the Tertiary of Kymi, Greece, deposited in what is nowadays the Natural History Museum, UK (Woodward 1901), which, however, might correspond to a different taxon. Given the unknown location and status of the type material for these taxa, and their almost complete disappearance from the literature (but see Bannikov 1993), I suggest that †Allocotus should be treated as invalid and †Morone ubinoi as nomen dubium. Adding to the confusion, additional percomorph taxa have been signaled in the Miocene of Kymi, but all these old attributions are doubtful and poorly documented (Valenciennes 1861; Gaudry 1862; Unger 1867; Appendix). The holotype of one of these, known as †Smerdis isabellae Gaudry 1862, has been synonymized by MNHN, Paris, staff with Woodward’s †Morone (= Labrax) ubinoi in the early 1900s (T.A. personal observations).
Distribution
In Greece, this taxon is known (possibly) from the early Miocene of an unknown locality near Kymi, Euboea. Outside Greece, Lates-like Centropomidae were present in Eocene–Neogene freshwater, brackish and sometimes marine deposits in Europe and Afroarabia (Murray and Attia 2004; Otero 2004; Argyriou et al. 2015). A total of fourteen species (Eschmeyer and Fong 2019) survive in African fresh–estuarine waters, and Indo-Pacific coastal environments (Froese and Pauly 2019).
Comments
The original description of this taxon does not contain clear information about its geographical or lithological provenance, other than that the fossil comes from some calcareous, oolithic horizon in Greece, which was thought to resemble the Solnhofen limestone (de Waldheim 1850). This specimen was not found by the author himself, but was sent to him by a certain Dr. Ubino who found, or purchased it during his travels through Greece (de Waldheim 1838). Woodward traces the provenance of the material in British collections to the freshwater deposits of Kymi, Greece. It is thus logical to hypothesize that this specimen derives from the classical Miocene plant and fish-bearing horizons of Kymi (e.g., Unger 1867; Kottis et al. 2002), yet this needs to be corroborated by future sampling. Although the drawings of the specimen are rather uninformative (de Waldheim 1838, 1850), the figured individual shows a preopercle bearing a horizontal anteroventral arm, which, in turn, forms a robust posterior spinous process, and three robust ventral processes (Fig. 10a). The possible resemblance of this animal to the percichthyid †Cyclopoma (= Cyclopome), or the centropomid Lates was noted by de Waldheim (1838). The imaged preopercle differs clearly from that of the preopercle of the Eocene genus †Cyclopoma (Agassiz 1833–1843). The shape and orientation of these processes resemble instead the preopercles of Latinae (Latidae in Otero 2004; now included within Centropomidae; see Greenwood 1976, Betancur-R et al. 2017; Fig. 10b), leaving little room for doubt that the illustrated skull is a Lates-like centropomid. Despite this profound resemblance, and unlike Latinae (Otero 2004), the posterior margin of the dorsal limb of the Kymi specimen is figured as non-serrate, but this feature might have been poorly preserved, or illustrated. Additional similarities between the illustrated Kymi specimen and Lates-like animals (Otero 2004) can be found in the acute ventral and anterior processes of the ectopterygoid, or what appears to be a broad palatine toothplate, in the anterior portion of the skull, and in what appears to be a single, posterodorsal process of the opercle. Woodward, possibly based on material from Kymi housed in British collections, mentioned that †Morone (= Labrax) ubinoi lacks a posterior spinous process of the preopercle, and also exhibits two feeble spines on the posterior margin of the opercle (Woodward 1901). It is thus possible that Woodward lumped two different taxa, one being a moronid and the other being de Waldheim’s centropomid, into one. This highlights the need to restudy Woodward’s material. This notion is reinforced by personal observations on Gaudry’s material, which was used to establish the species †Smerdis isabellae and is now housed at the MNHN, Paris. The latter would require a detailed study to clarify its taxonomic status , which lies beyond the context of this work.

(a) Specimen used to establish †Allocotus sp. (de Waldheim 1838) and subsequently †Platacanthus ubinoi (de Waldheim 1850), but hereby reassigned to Centropomidae indet.; modified from de Waldheim 1838, scale not provided in original work. (b) Preopercle of Lates cf. niloticus from the Mio-Pliocene of Sahabi, Libya (collected and attributed by the author; the specimen belongs to the collections of the University of Benghazi, Libya), for comparison with the illustrated preopercle of (a). Scale bar equals 1 cm
-
Labridae Cuvier, 1817
Type Genus
Labrus Linnaeus, 1758.
Distribution
In Greece, this group is known from the Pliocene of Heraklion Basin (Gaudant 2001). Ichthyoliths from the Pliocene of Aghia Marina, Aegina Island, have been tentatively attributed to the sparid †Sargus cf. oweni (Leivaditis 1974), which was in turn found to be a junior synonym of the labrid †Trigonodon jugleri (Schultz and Bellwood 2004). This might correspond to another occurrence of Labridae in Greece, but it is so far poorly substantiated. Labrids appear in the Eocene of Italy (Bannikov and Carnevale 2010). Mostly disarticulated (e.g., oral or pharyngeal teeth), but also articulated remains are common in Neogene deposits around the world (e.g., Algeria, Italy, France, Portugal, Poland, Austria, Nigeria, the United States, and New Zealand; summarized in Bellwood et al. 2019).
-
Acanthuriformes sensu Betancur-R et al. 2017
-
Acanthuridae Bleeker, 1859
Type Genus
Acanthurus Forskål, 1775.
Distribution
In Greece, this taxon is known from the Tortonian of Makrilia Formation, in the vicinity of Ierapetra, Crete (Gaudant 2004), and the Tortonian of Gavdos (Gaudant et al. 2005). Outside Greece, articulated acanthurid fossils are known from the Paleogene of Europe (e.g., Italy, Switzerland, Russia) (Tyler and Bannikov 2000; Tyler and Micklich 2011), while their Miocene occurrences (e.g., Austria and Croatia) usually correspond to isolated teeth (Schultz 2003; Tripalo et al. 2016). Nowadays, the family is absent from the Mediterranean, but is otherwise characterized by circumtropical distribution (Froese and Pauly 2019).
Comments
The Gavdos specimen is probably a juvenile (Gaudant et al. 2005).
-
Tetraodontiformes Berg, 1937
-
Monacanthidae Nardo, 1842
Type Genus
Monacanthus Oken, 1817.
-
† Frigocanthus Sorbini and Tyler, 2004
Type Species
†Frigocanthus stroppanobili Sorbini and Tyler, 2004.
Included Taxa
†Frigocanthus margaritatus Sorbini and Tyler, 2004.
Distribution
In Greece, †F. margaritatus is known from the Pliocene of Heraklion Basin (Gaudant 2001; Sorbini and Tyler 2004), whereas †F. stroppanobili from the middle-late Pliocene of Chania , Crete (Sorbini and Tyler 2004). Outside Greece, this taxon is known from the Pliocene and Pleistocene of Italy (Sorbini and Tyler 2004).
-
Caproiformes sensu Rosen, 1984
-
Caproidae Lowe, 1843
Type Genus
Capros Lacepède, 1802.
-
Capros Lacepède, 1802
Type Species
Capros (= Zeus) aper (Linnaeus, 1758).
Distribution
In Greece, this genus is known only from the Tortonian of Makrilia Formation, in the vicinity of Ierapetra (Gaudant 2004). Outside Greece, fossil species of the genus have been recognized in the Oligocene of the Carpathians, the Caucasus, Germany, and France (Baciu et al. 2005b; Přikryl et al. 2016c), and the late Miocene of Italy and Algeria (Baciu et al. 2005a). The living species C. aper is known from the Pliocene–Pleistocene of Italy (Landini and Sorbini 2005a).
6 Conclusions and Avenues for Future Research
Over a century and a half of exploration and sporadic scientific research on the actinopterygian fossil record of Greece has led to the discovery of numerous fossil sites, and laid the foundations for further work on the regional fossil record of the clade. Actinopterygian fossils in Greece have a patchy record that spans over 180 million years, from the Toarcian (Lower Jurassic) to the Quaternary, with the vast majority of known occurrences concentrated in the late Neogene (late Miocene–Pliocene) and being of clear marine affinities (summarized in the Appendix). Little information is available for older, or freshwater and brackish localities. The pre-Neogene fossil record of Greece merits further scientific attention and exploration, especially given the huge record of sedimentary rocks and complex geological history of Greece. Deposits predating the Jurassic have not yet, to the author’s knowledge, been investigated extensively for potential paleoichthyological content (excluding conodonts), and like in the case of chondrichthyans (Argyriou et al. 2017), actinopterygian fossils are anticipated in Paleozoic–early Mesozoic deposits.
Historically, alpine sedimentary formations of Greece (e.g., the “External Hellenides”), which record a major part of the Mesozoic and early Cenozoic, have been considered to be of principally tectonostratigraphic importance to the geological and paleontological community, and have remained outside the interest of vertebrate paleontologists. Yet, preliminary investigations have indicated the yet untapped paleoichthyological potential of such deposits. The presumably rich Toarcian bonebeds of Ionian Unit exposures in Lefkada have been totally neglected since their first discovery (Kottek 1964), despite their potential informativeness on early teleost diversification. Moreover, understudied fish assemblages from the late Cretaceous–Paleogene of Pindos Unit can shed light on the evolution and biogeography of Tethyan faunas (Koch and Nikolaus 1969; Cavin et al. 2012; Argyriou and Davesne 2021; pers. obs. on the K/Pg of Eurytania, Northern–Central Peloponnese and Gavdos) during this critical time interval of global marine restructuring, which is otherwise poorly documented in the global fossil record. Further research should be directed toward freshwater and brackish faunas, which are so far known from Miocene or younger deposits (e.g., Miocene of Kymi Euboea: de Waldheim 1838, 1850; Unger 1867; Miocene of Chios: Reichenbacher and Böhme 2004), and their insights into the evolution of modern European freshwater ichthyofaunas and their biogeographical and paleohydrological inferences.
Late Miocene–Pliocene fossiliferous localities and horizons, especially the ones in Crete and Gavdos, host diverse actinopterygian assemblages of often exquisite preservation quality (Gaudant et al. 1994, 1997a, 2005, 2006; Gaudant 2001, 2004). Numerous families and genera have already been recognized in these assemblages, largely on the basis of broad morphological and meristic similarities with late Miocene taxa from Italy and Algeria (e.g., Arambourg 1925, 1927; Gaudant 2008b). Previous works, however, have treated these assemblages from an alpha-taxonomic point of view, and provided some limited information on their paleoenvironmental significance. Additional taxonomic diversity, including new species, remains to be identified in these sites, pending detailed anatomical descriptions (Přikryl and Carnevale 2017), and is a prerequisite for further systematic and biogeographic work. The importance of following this line of research cannot be overstressed, especially when taking into account the scarcity of information for this part of (Neo)Tethys, in comparison to what is now the Western and Central Mediterranean (e.g., Arambourg 1925, 1927; Landini and Sorbini 2005a; Carnevale 2007; Gaudant 2008a, b). Research on Greek fossils is anticipated to unveil further clues for tracing the gradual emergence of the modern endemic and Atlantic hallmark of Mediterranean marine faunas from the tropical, Indopacific-like faunas of the Miocene, and the role of major events, such as the Messinian Salinity Crisis and Pleistocene cooling(s) in this process (Gaudant et al. 1994, 1997a, 2005, 2006; Gaudant 2001, 2002, 2004; Landini and Sorbini 2005a, b).
This work collates available information on actinopterygian fossils and their spatiotemporal distribution in Greece, as well as their potential significance. It is hoped to form a stimulating basis for further paleoichthyological research in the country. As a concluding remark, properly curated paleoichthyological collections of Greek fish fossils are crucial for invigorating paleoichthyology in the country, and, given funding and staff limitations, Greek institutions and museums are encouraged to work toward this goal.
References
Abbazzi L, Angelone C, Arca M, Barisone G, Bedetti C, Delfino M, Kotsakis T, Marcolini F, Palombo MR, Pavia M, Piras P, Rook L, Torre D, Tuveri C, Valli AMF, Wilkens B (2004) Plio-Pleistocene fossil vertebrates of Monte Tuttavista (Orosei, Eastern Sardinia, Italy), an overview. Riv Ital Paleontol Stratigr 110:681–706
Agiadi K (this volume) The fossil otolith record of fishes (Vertebrata: Teleostei) in Greece. In: Vlachos E (ed) The fossil vertebrates of Greece vol. 1 – Basal vertebrates, amphibians, reptiles, afrotherians, glires, and primates. Springer – Nature Publishing Group, Cham
Agiadi K, Koskeridou E, Triantaphyllou M, Girone A, Karakitsios V (2013) Fish otoliths from the Pliocene Heraklion Basin (Crete Island, Eastern Mediterranean). Geobios 46:461–472
Agiadi K, Vasileiou G, Koskeridou E, Moissette P, Cornée J-J (2019) Coastal fish otoliths from the early Pleistocene of Rhodes (eastern Mediterranean). Geobios 55:1–15
Alfaro ME, Santini F, Brock C, Alamillo H, Dornburg A, Rabosky DL, Carnevale G, Harmon LJ (2009) Nine exceptional radiations plus high turnover explain species diversity in jawed vertebrates. Proc Natl Acad Sci 106:13410–13414
Alfaro ME, Faircloth BC, Harrington RC, Sorenson L, Friedman M, Thacker CE, Oliveros CH, Černý D, Near TJ (2018) Explosive diversification of marine fishes at the Cretaceous–Palaeogene boundary. Nat Ecol Evol 2:688–696
Antipa G (1904) Die Clupeinen des westlichen Teiles des Schwarzen Meeres und der Donaumündungen. Anz Akad Wiss Wien 41:299–303
Arambourg C (1925) Révision des poissons fossiles de Licata (Sicile). Masson, Paris
Arambourg C (1927) Les poissons fossiles d’Oran. Matériaux pour la Carte géologique d’Algérie. Jules Careonel, Alger
Argyriou T, Davesne D (2021) Offshore marine actinopterygian assemblages from the Maastrichtian–Paleogene of the Pindos Unit in Eurytania, Greece. PeerJ 9:e10676
Argyriou T, Theodorou G (2011) New findings from the Pliocene (Zanclean) ichthyofauna of Aegina island, Greece. Paper presented at the European Association of Vertebrate Paleontologists 9th annual meeting, Heraklion, 2011
Argyriou T, Cook TD, Muftah AM, Pavlakis P, Boaz NT, Murray AM (2015) A fish assemblage from an early Miocene horizon from Jabal Zaltan, Libya. J Afr Earth Sci 102:86–101
Argyriou T, Clauss M, Maxwell EE, Furrer H, Sánchez-Villagra MR (2016) Exceptional preservation reveals gastrointestinal anatomy and evolution in early actinopterygian fishes. Sci Rep 6:18758
Argyriou T, Romano C, Carrillo-Briceño JD, Brosse M, Hofmann R (2017) The oldest record of gnathostome fossils from Greece: Chondrichthyes from the Lopingian of Hydra Island. Palaeontol Electron 20.1.8A:1–9
Argyriou T, Giles S, Friedman M, Romano C, Kogan I, Sánchez-Villagra MR (2018) Internal cranial anatomy of early Triassic species of †Saurichthys (Actinopterygii: †Saurichthyiformes): implications for the phylogenetic placement of †saurichthyiforms. BMC Evol Biol 18:161
Arratia G (1999) The monophyly of Teleostei and stem-group teleosts. Consensus and disagreements. In: Arratia G, Schultze HP (eds) Mesozoic fishes 2 – systematics and fossil record. Dr. Friedrich Pfeil, Munich, pp 265–334
Arratia G (1997) Basal teleosts and teleostean phylogeny. Palaeo Ichthyol 7:5–168
Arratia G (2017) New Triassic teleosts (Actinopterygii, Teleosteomorpha) from northern Italy and their phylogenetic relationships among the most basal teleosts. J Vertebr Paleontol 37:e1312690
Arratia G, Hikuroa DCH (2010) Jurassic fishes from the Latady Group, Antarctic Peninsula, and the oldest teleosts from Antarctica. J Vertebr Paleontol 30:1331–1342
Arratia G, Scasso RA, Kiessling W (2004) Late Jurassic fishes from Longing Gap, Antarctic Peninsula. J Vertebr Paleontol 24:41–55
Bachmayer F, Symeonidis N (1978) Eine fossile Seenadel (Syngnathidae) aus dem Obermiozän von Ierapetra, Insel Kreta (Griechenland). Ann Naturhist Mus Wien 81:121–127
Bachmayer F, Kähsbauer P, Symeonidis N (1984) Weitere fossile Seenadelreste aus dem Obermiozän der Insel Kreta (Griechenland). Ann Naturhist Mus Wien 86A:1–6
Baciu D-S, Bannikov AF, Santini F (2005a) A new species of Caproidae (Acanthomorpha, Teleostei) from the Messinian (upper Miocene) of Oran (Algeria). Geodiversitas 27:381–390
Baciu D-S, Bannikov AF, Tyler JC (2005b) Revision of the fossil fishes of the family Caproidae (Acanthomorpha) Miscelanea Paleontologica nr. 8. Studi e ricerche sui giacimenti terziari di Bolca XI, Verona, pp 7–74
Bannikov AF (1984) The new subfamily of carangid fishes. Studi e ricerche sui giacimenti terziari di Bolca 4:319–321
Bannikov AF (1987) The phylogenetic relationships of the carangid fishes of the subfamily Caranginae. Paleontol J 21:44–54
Bannikov AF (1993) A new species of bass (Teleostie, Moronidae) from the Sarmatian of Moldova. Paleontol J 27:54–62
Bannikov AF, Carnevale G (2010) Bellwoodilabrus landinii n. gen., n. sp., a new genus and species of labrid fish (Teleostei, Perciformes) from the Eocene of Monte Bolca. Geodiversitas 32:201–220
Bellas SM, Kussius K, Kommerell JK, Kriwet J (1998) lntegrated biostratigraphical approach of the Neogene Pigadia basin of Karpathos lsland (Dodecanes Group, Greece) - lmplications on the depositional palaeoenvironment based on calcareous nannofossils, ostracodes and facies development data with special references to Messinian fish assemblages. Berliner geowissenschaftliche Abhandlungen Reihe E 28:129–156
Bellwood DR, Schultz O, Siqueira AC, Cowman PF (2019) A review of the fossil record of the Labridae. Ann Naturhist Mus Wien 121:125–194
Berrebi P, Tsigenopoulos CS (2003) Phylogenetic organization of the genus Barbus sensu stricto: a review based on data obtained using molecular markers. In: The freshwater fishes of Europe 5. AULA, Wiebelsheim, pp 11–22
Betancur-R R, Broughton RE, Wiley EO, Carpenter K, López JA, Li C, Holcroft NI, Arcila D, Sanciangco M, Cureton Ii JC, Zhang F, Buser T, Campbell MA, Ballesteros JA, Roa-Varon A, Willis S, Borden WC, Rowley T, Reneau PC, Hough DJ, Lu G, Grande T, Arratia G, Ortí G (2013) The tree of life and a new classification of bony fishes. PLoS Curr 5:ecurrents.tol.53ba26640df0ccaee75bb165c8c26288
Betancur-R R, Ortí G, Pyron RA (2015) Fossil-based comparative analyses reveal ancient marine ancestry erased by extinction in ray-finned fishes. Ecol Lett 18:441–450
Betancur-R R, Wiley EO, Arratia G, Acero A, Bailly N, Miya M, Lecointre G, Ortí G (2017) Phylogenetic classification of bony fishes. BMC Evol Biol 17:162
Böhme M (2010) Ectothermic vertebrates (Actinopterygii, Allocaudata, Urodela, Anura, Crocodylia, Squamata) from the Miocene of Sandelzhausen (Germany, Bavaria) and their implications for environment reconstruction and palaeoclimate. Paläontol Z 84:3–41
Böhme M, Ilg A (2003) fosFARbase, Vol. 2019
Broughton RE, Betancur-R R, Li C, Arratia G, Ortí G (2013) Multi-locus phylogenetic analysis reveals the pattern and tempo of bony fish evolution. PLoS Curr 5:ecurrents.tol.2ca8041495ffafd0c92756e75247483e
Bürgin T (1994) Echte Knochenfische (Actinopterygii; Teleostei) aus dem Oberen Miozän der Insel Kreta (Griechenland): Vorläufige Resultate. Eclog Geol Helv 87:1007–1022
Calvo JP, Triantaphyllou MV, Regueiro M, Stamatakis MG (2012) Alternating diatomaceous and volcaniclastic deposits in Milos Island, Greece. A contribution to the upper Pliocene–lower Pleistocene stratigraphy of the Aegean Sea. Palaeogeogr Palaeoclimatol Palaeoecol 321–322:24–40
Carnevale G (2003) Redescription and phylogenetic relationships of Argyropelecus logearti (Teleostei: Stomiiformes: Sternoptychidae), with a brief review of fossil Argyropelecus. Riv Ital Paleontol Stratigr 109:63–76
Carnevale G (2007) Fossil fishes from the Serravallian (Middle Miocene) of Torricella Peligna, Italy. Paleontographica Ital 91:1–67
Carnevale G, Landini W, Sarti G (2006) Mare versus Lago-mare: marine fishes and the Mediterranean environment at the end of the Messinian Salinity Crisis. J Geol Soc 163:75–80
Carnevale G, Bannikov AF, Marramà G, Tyler JC, Zorzin R (2014) The Pesciara-Monte Postale Fossil-Lagerstätte: 2. Fishes and other vertebrates. Rend Soc Paleontol Ital 4:37–63
Carnevale G, Dela Pierre F, Natalicchio M, Landini W (2018) Fossil marine fishes and the ‘Lago Mare’; event: Has the Mediterranean ever transformed into a brackish lake? Newsl Stratigr 51:57–72
Cavender TM (1991) The fossil record of the Cyprinidae. In: Winfield IJ, Nelson JS (eds) Cyprinid Fishes: systematics, biology and exploitation. Springer Netherlands, Dordrecht, pp 34–54
Cavin L, Alexopoulos A, Piuz A (2012) Late Cretaceous (Maastrichtian) ray-finned fishes from the island of Gavdos, southern Greece, with comments on the evolutionary history of the aulopiform teleost Enchodus. Bull Soc Géol Fr 183:561–572
Cavin L, Forey PL, Giersch S (2013) Osteology of Eubiodectes libanicus (Pictet & Humbert, 1866) and some other ichthyodectiformes (Teleostei): phylogenetic implications. J Syst Palaeontol 11:115–177
Chalifa Y (1996) A new species of Enchodus (Aulopiformes: Enchodontidae) from the Northern Negev, Israel, with comments on the evolutionary trends in the Enchodontoidei. In: Arratia G, Viohl G (eds) Mesozoic fishes - systematics and paleoecology. Dr. Friedrich Pfeil, München, pp 349–367
Cornée J-J, Moissette P, Joannin S, Suc J-P, Quillévéré F, Krijgsman W, Hilgen F, Koskeridou E, Münch P, Lécuyer C, Desvignes P (2006) Tectonic and climatic controls on coastal sedimentation: the Late Pliocene–Middle Pleistocene of northeastern Rhodes, Greece. Sediment Geol 187:159–181
da Silva HMA, Gallo V (2011) Taxonomic review and phylogenetic analysis of Enchodontoidei (Teleostei: Aulopiformes). An Acad Bras Cienc 83:483–511
da Silva HMA, Gallo V (2016) Distributional patterns of enchodontoid fishes in the Late Cretaceous. Cretac Res 65:223–231
Davesne D, Gallut C, Barriel V, Janvier P, Lecointre G, Otero O (2016) The phylogenetic intrarelationships of spiny-rayed fishes (Acanthomorpha, Teleostei, Actinopterygii): Fossil Taxa increase the congruence of morphology with molecular data. Front Ecol Evol 4:129
Davis MP, Fielitz C (2010) Estimating divergence times of lizardfishes and their allies (Euteleostei: Aulopiformes) and the timing of deep-sea adaptations. Mol Phylogenet Evol 57:1194–1208
de Sant’Anna VB, Collette BB, Godfrey SJ (2013) †Belone countermani, a new Miocene needlefish (Belonidae) from the St. Marys Formation of Calvert Cliffs, Maryland. Proc Biol Soc Wash 126:137–150
de Waldheim GF (1838) Recherches sur les ossemens fossiles de la Russie II. Semen, Moscow
de Waldheim GF (1850) Sur une poisson fossile de la Grèce. Bull Socimperiale des naturalistes Moscou 23:285–288
Denton JSS (2013) Lanternfish (Teleostei, Myctophiformes, Myctophidae) body fossils from the Modelo Formation (upper Miocene) of Los Angeles County. Calif J Vertebr Paleontol 33:786–793
Dermitzakis MD (1979) The Neogene of Zakynthos. In: Symeonidis N, Papanikolaou D, Dermitzakis MD (eds) Field guide to the Neogene of Megara – Pelopponisos – Zakynthos. Department Geology & Paleontology, University of Athens, Athens, pp 45–67
Dermitzakis MD, Georgiades-Dikeoulia E, Velitzelos E (1986) Ecostratigraphic observations on the Messinian deposits of Akropotamos area (Kavala, N. Greece). Ann Géol Pays Ηéllen 33:367–376
Drinia H, Dermitzakis MD, Kouli K, Tsourou T (2002) Sedimentary facies analysis and paleoenvironmental interpretation of Vatera Formation, Lesvos Island, Greece. Ann Géol Pays Ηéllen 39:15–35
Eschmeyer WN, Fong JD (2019) Species by family/subfamily in the Catalog of Fishes, vol 2016. California Academy of Sciences, San Francisco
Fielitz C, González-Rodríguez KA (2010) A new species of Enchodus (Aulopiformes: Enchodontidae) from the Cretaceous (Albian to Cenomanian) of Zimapán, Hidalgo, México. J Vertebr Paleontol 30:1343–1351
Freuenthal T (1969) Stratigraphy of Neogene deposits in the Khania Province, Crete, with special reference to Foraminifera of the family Planorbulinidae and the genus Heterostegina. Utrecht Mircropaleontol Bull 1:1–208
Friedman M (2009) Ecomorphological selectivity among marine teleost fishes during the end-Cretaceous extinction. Proc Natl Acad Sci 106:5218–5223
Friedman M (2015) The early evolution of ray-finned fishes. Palaeontology 58:213–228
Friedman M, Sallan LC (2012) Five hundred million years of extinction and recovery: a phanerozoic survey of large-scale diversity patterns in fishes. Palaeontology 55:707–742
Friedman M, Beckett HT, Close RA, Johanson Z (2015) The English chalk and London clay: two remarkable British bony fish Lagerstätten. Geol Soc Lond, Spec Publ 430:165
Froese R, Pauly D (2019) FishBase, Vol. 08/2019. World Wide Web electronic publication
Gaudant J (1979) «Pachylebias crassicaudus» (Agassiz) (Poisson téléostéen, cyprinodontiforme), un constituant majeur de l’ichthyofaune du Messinien continental du bassin méditerranéen. Geobios 12:47–73
Gaudant J (1980) Sur la présence d’Alosa crassa SAUVAGE (Poissons téléostéens, Clupeidae) dans les gypses messiniens de Crète occidentale. Proc K Ned Akad Wet B 83:263–268
Gaudant J (2001) Amnissos: un gisement clé pour la connaissance de l’ichthyofaune du Pliocène supérieur de Crète. Ann Naturhist Mus Wien 102A:131–187
Gaudant J (2002) La crise messinienne et ses effets sur l’ichthyofaune néogène de la Méditerranée: le témoignage des squelettes en connexion de poissons téléostéens. Geodiversitas 24:691–710
Gaudant J (2004) Additions à l’ichthyofaune tortonienne du bassin de Ierapetra (Crète orientale, Grèce). Ann Naturhist Mus Wien 105:257–285
Gaudant J (2008a) L’ichthyofaune messinienne du basin de Sorbas (Almería, Espagne) et ses rapports avec l’environment sédimentaire. Rev Esp Paleont 23:211–223
Gaudant J (2008b) Paléobiodiversité et paléoenvironnements: l’exemple des gisements de poissons téléostéens du Messinien préévaporitique d’Oran et du bassin du Chélif (Algérie). Geodiversitas 30:141–163
Gaudant J (2014a) Articulated skeletons of Palaeoleuciscus aff. oeningensis (Agassiz) (Teleostean fish, Cyprinidae) in the lower Miocene of Frankfurt on the Main (Germany), with a note on cyprinid pharyngeal teeth found in the lower Miocene of the Mainz basin. Palaeobiodiv Palaeoenviron 94:481–486
Gaudant J (2014b) Fish mass mortality in the Upper Miocene laminated gypsum of western Crete (Hania Province, Greece). Palaeodiversity 7:39–45
Gaudant J (2015) Re-examination of the upper Miocene freshwater fish fauna from Höwenegg (Hegau, Germany). Swiss J Palaeontol 134:117–127
Gaudant J, Ambroise D (1999) Réexamen critique des Myctophidae Messiniens de Licata (Sicile, Italie): conséquences taxonomiques. Cybium 23:131–145
Gaudant J, Courme M-D (2014) A new Pliocene marine fish fauna from north-western Cyprus, with a micropalaeontological account. Neues Jahrb Geol Palaontol Abh 272:205–212
Gaudant J, Symeonidis N (1995) Découverte d’une frayère à Spratelloides (poissons téléostéens) dans le Pliocène moyen de Kalamaki (Attique, Grèce). Ann Géol Pays Ηéllen 36:331–349
Gaudant J, Delrieau B, Dermitzakis MD, Symeonidis NK (1994) Découverte d’une ichthyofaune marine dans les diatomites du Pliocène supérieur (Plaisancien) des environs d’Héraklion (Crète centrale, Grèce). C R Acad Sci Paris 319:589–596
Gaudant J, Fourtanier E, Lauriat-Rage A, Tsagaris S, Vénec-Peyré M-T, Zorn I (1997a) Découverte d’une ichthyofaune marine dans le Messinien préévaporitique de la Messara (Crète centrale, Grèce): interprétation paléoécologique. Géol Méditerr 24:175–195
Gaudant J, Saint-Martin J-P, Bessedik M, Mansour B, Moissette P, Rouchy J-M (1997b) Découverte d’une frayère de poissons téléosteens dans des diatomites messiniennes du Djebel Murdjadjo (environs d’Oran, Algérie). J Afr Earth Sci 24:511–529
Gaudant J, Barker MJ, Courme M-D, di Stephano A, Martill DM, Vénec-Peyré M-T, Zorn I, Panayides I (2000) Alassa: a new fossil fish fauna from the middle Miocene Serravallian of Cyprus. Paper presented at the 3rd, International conference on the geology of the eastern Mediterranean, Nicosia, Cyprus, 2000
Gaudant J, Tsaparas N, Antonarakou A, Drinia H, Dermitzakis MD (2005) The Tortonian fish fauna of Gavdos Island (Greece). C R Palevol 4:687–695
Gaudant J, Tsaparas N, Antonarakou A, Drinia H, Saint-Martin S, Dermitzakis MD (2006) A new marine fish fauna from the pre-evaporitic Messinian of Gavdos Island (Greece). C R Palevol 5:795–802
Gaudant J, Cavallo O, Courme-Rault M-D, Fornaciari E, Lauriat-Rage A (2008) Paléontologie du gisement de poissons fossiles du Messinien préévaporitique de Roddi, près d’Alba (Piémont, Italie). Riv Piemont Storia Nat 29:3–60
Gaudant J, Courme-Rault M-D, Fornaciari E, Fourtanier E (2010a) The Upper Miocene fossil fish locality of Pecetto di Valenza (Piedmont, Italy): a multidisciplinary approach. Boll Soc Paleontol Ital 49:203–225
Gaudant J, Courme-Rault M-D, Saint-Martin S (2010b) On the fossil fishes, diatoms, and foraminifera from Zanclean (Lower Pliocene) diatomitic sediments of Aegina Island (Greece): a stratigraphical and palaeoenvironmental study. Palaeodiversity 3:141–149
Gaudant J, Soria MJ, Fierro I, Saint Martin S (2015) The Upper Miocene fish fauna of the Fortuna Basin, as observed in the surroundings of Albanilla (Murcia Province, Spain). Span J Palaeontol 30:67–78
Gaudry A (1862) Animaux fossiles et géologie de l’Attique d’apres les recherches faites en 1855–56 et en 1860 sous les auspices de l’Academie des sciences par Albert Gaudry. Savy, F, Paris
Georgalis GL, Villa A, Delfino M (2017) Fossil lizards and snakes from Ano Metochi – a diverse squamate fauna from the latest Miocene of northern Greece. Hist Biol 29:730–742
Giles S, Coates MI, Garwood RJ, Brazeau MD, Atwood R, Johanson Z, Friedman M (2015) Endoskeletal structure in Cheirolepis (Osteichthyes, Actinopterygii), an early ray-finned fish. Palaeontology 58:849–870
Giles S, Xu G-H, Near TJ, Friedman M (2017) Early members of ‘living fossil’ lineage imply later origin of modern ray-finned fishes. Nature 549:265–268
Gmelin JF (1789) Pisces; Caroli a Linné. Systema naturae per regna tria naturae: secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis, Lipsiae
Goody PC (1969) The relationships of certain Upper Cretaceous teleosts with special reference to myctophoids. Bull Br Mus (Nat Hist) Geol Ser Suppl 7:1–255
Grande L (1985) Recent and fossil clupeomorph fishes with materials for revision of the subgroups of clupeoids. Bull Am Mus Nat Hist 181:231–372
Grande L (2010) An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy: the resurrection of Holostei. American Society of Ichthyologists and Herpetologists Special Publication 6. Copeia Suppl 10:871
Greenwood PH (1976) A review of the family Centropomidae (Pisces, Perciformes). Bull Br Mus (Nat Hist) Zool 29:1–81
Gregorová R (2011) Fossil fish fauna (Teleostei, Selachii) from the Dynów marlstone (Rupelian, NP 23) of the Menilitic Formation at the locality of Litenčice (Czech Republic). Acta Mus Moraviae – Sci Geol 2:3–33
Grim O (1901) The herrings of the Sea of Azov. Vestnik Rybopromysi St. Petersburg 16
Guinot G, Cavin L (2016) ‘Fish’ (Actinopterygii and Elasmobranchii) diversification patterns through deep time. Biol Rev 91:950–981
Helfman GS, Collette B, Facey DE, Bowen BW (2009) The diversity of fishes. Wiley-Blackwell, Chichester
Hilton EJ, Forey PL (2009) Redescription of †Chondrosteus acipenseroides Egerton, 1858 (Acipenseriformes, †Chondrosteidae) from the Lower Lias of Lyme Regis (Dorset, England), with comments on the early evolution of sturgeons and paddlefishes. J Syst Palaeontol 7:427–453
Holloway WL, Claeson KM, Sallam HM, El-Sayed S, Kora M, Sertich JJW, O’Connor PM (2017) A new species of the neopterygian fish Enchodus from the Duwi Formation, Campanian, Late Cretaceous, Western Desert, central Egypt. Acta Palaeontol Pol 62:603–611
Hordijk K, de Bruijn H (2009) The succession of rodent faunas from the Mio/Pliocene lacustrine deposits of the Florina-Ptolemais-Servia Basin (Greece). Hell J Geosci 44:21–103
Hughes LC, Ortí G, Huang Y, Sun Y, Baldwin CC, Thompson AW, Arcila D, Betancur-R R, Li C, Becker L, Bellora N, Zhao X, Li X, Wang M, Fang C, Xie B, Zhou Z, Huang H, Chen S, Venkatesh B, Shi Q (2018) Comprehensive phylogeny of ray-finned fishes (Actinopterygii) based on transcriptomic and genomic data. Proc Natl Acad Sci 115:6249–6254
Jordan DS, Gilbert JZ (1919) Fossil fishes of the (Miocene) Monterey Formations of Southern Californi; pp. 13–60, Fossil fishes of Southern California. Stanford University Press
Kamperis EG (1987) Geological and petrogeological study of Northwest Peloponnese. School of mining & metallurgical engineering. National Technical University of Athens, Athens
Kevrekidis C (2016) Taxonomic study of selected Clupeidae (herrings) from the Miocene of the Mediterranean and Paratethys: comparative meristics and morphometry: In Faculty of geosciences, M.Sc. Thesis, Ludwig-Maximilians-Universität München, Munich
Koch KE, Nikolaus HJ (1969) Zur Geologie des Ostpindos – Flyschbeckens und seiner Umrandung. Institute for geology and subsurface research, Athens
Konwert M, Stumpf S (2017) Exceptionally preserved Leptolepidae (Actinopterygii, Teleostei) from the late early Jurassic Fossil-Lagerstätten of Grimmen and Dobbertin (Mecklenburg-Western Pomerania, Germany). Zootaxa 4243:249–296
Kottek A (1964) Fischreste aus dem griechischen Lias. Ann Géol Pays Ηéllen 39:175–181
Kottelat M, Freyhof J (2007) Handbook of European freshwater fishes. Publications Kottelat, Cornol and Berlin
Kottis T, Ioakim C, Velitzelos E, Kvaček Z, Denk T, Velitzelos D (2002) Field trip guidebook Evia Island. Neogene sedimantary basin of Kymi/Aliveri, Athens
Kriwet J (2003) Lancetfish teeth (Neoteleostei, Alepisauroidei) from the Early Cretaceous of Alcaine, NE Spain. Lethaia 36:323–331
Landini W, Sorbini C (2005a) Evolutionary dynamics in the fish faunas of the Mediterranean basin during the Plio-Pleistocene. Quat Int 140–141:64–89
Landini W, Sorbini C (2005b) Evolutionary trends in the Plio-Pleistocene ichthyofauna of the Mediterranean Basin: nature, timing and magnitude of the extinction events. Quat Int 131:101–107
Leivaditis GI (1974) Geological and geomorphological observations on Aegina Island: In Department of Geology, PhD thesis, National and Kapodistrian University of Athens
López-Arbarello A, Sferco E (2018) Neopterygian phylogeny: the merger assay. R Soc Open Sci 5:172337
Lu J, Giles S, Friedman M, den Blaauwen JL, Zhu M (2016) The oldest actinopterygian highlights the cryptic early history of the hyperdiverse ray-finned fishes. Curr Biol 26:1602–1608
Maxwell EE, Argyriou T, Stockar R, Furrer H (2017) Evolutionary implications of the ontogeny and reproductive biology of Saurichthyidae (Actinopterygii). Zitteliana 91:62
Melentis JK (1967a) Studien über fossile Vertebraten Griechenlands. 21. Über Coelodus münsteri AG. (Pisces) aus dem Cenoman von Griechenland. Ann Géol Pays Ηéllen 18:456–462
Melentis JK (1967b) Über Coelodus münsteri AG. (Pisces) aus dem Cenoman von Griechenland. Praktika Akadimias Athinon 42:115–123
Merle D, Lauriat-Rage A, Gaudant J, Pestrea S, Courme-Rault M-D, Zorn I, Blanc-Valleron M-M, Rouchy J-M, Orszag-Sperber F, Krijgsman W (2002) Les paléopeuplements marins du Messinien pré-évaporitique de Pissouri (Chypre, Méditerranée orientale): aspects paléoécologiques précédant la crise de salinité messinienne. Geodiversitas 24:669–689
Micklich N, Hildenbrandt HL (2010) Emergency excavation in the Grube (Unterfeld) Frauenweiler clay pit (Oligocene Rupelian; Baden-Wurttemberg, S Germany): new records and palaeoenvironmental information. Kaupia 17:3–21
Monsch K, Bannikov AF (2012) New taxonomic synopses and revision of the scombroid fishes (Scombroidei, Perciformes), including billfishes, from the Cenozoic of territories of the former USSR. Earth Environ Sci Trans R Soc Edinb 102:253–300
Müller AH, Strauch F (1994) Eine pliozäne Fischfauna von Prassas bei Heraklion, Kreta. Münster Forsch Geol Palaeontol 76:243–253
Murray AM, Attia YS (2004) A new species of lates (Teleostei: Perciformes) from the Lower Oligocene of Egypt. J Vertebr Paleontol 24:299–308
Nazarkin MV, Bannikov AF (2014) Fossil mackerel (Actinopterygii: Scombridae: Scomber) from the Neogene of South-Western Sakhalin, Russia. Zoosyst Rossica 23:158–163
Near TJ, Eytan RI, Dornburg A, Kuhn KL, Moore JA, Davis MP, Wainwright PC, Friedman M, Smith WL (2012) Resolution of ray-finned fish phylogeny and timing of diversification. Proc Natl Acad Sci 109:13698–13703
Near TJ, Dornburg A, Eytan RI, Keck BP, Smith WL, Kuhn KL, Moore JA, Price SA, Burbrink FT, Friedman M, Wainwright PC (2013) Phylogeny and tempo of diversification in the superradiation of spiny-rayed fishes. Proc Natl Acad Sci 110:12738–12743
Nelson JS, Grande TC, Wilson MVH (2016) Fishes of the world. Wiley, Hoboken, p i-xli + 707
Nybelin O (1974) A Revision of the Leptolepid fishes. Acta Regiae Societatis Scientiarum et Litterarum Gothoburgensis Zoologica 9:1–202
Offshore marine actinopterygian assemblages from the Maastrichtian–Paleogene of the Pindos Unit in Eurytania, Greece
Olsen PE (1984) The skull and pectoral girdle of the parasemionotid fish Watsonulus eugnathoides from the Early Triassic Sakamena Group of Madagascar, with comments on the relationships of the holostean fishes. J Vertebr Paleontol 4:481–499
Otero O (2004) Anatomy, systematics and phylogeny of both recent and fossil latid fishes (Teleostei, Perciformes, Latidae). Zool J Linnean Soc 141:81–133
Otero O, Gayet M (2001) Palaeoichthyofaunas from the Lower Oligocene and Miocene of the Arabian Plate: palaeoecological and palaeobiogeographical implications. Palaeogeogr Palaeoclimatol Palaeoecol 165:141–169
Otero O, Lécuyer C, Fourel F, Martineau F, Mackaye HT, Vignaud P, Brunet A (2011) δ18O of fishes indicates a Messinian increasing of the aridity in Central Africa. Geology 39:435–438
Panagopoulou E, Tourloukis V, Thompson N, Konidaris G, Athanassiou A, Giusti D, Tsartsidou G, Karkanas P, Harvati K (2018) The lower palaeolithic site of Marathousa 1, Megalopolis, Greece: overview of the evidence. Quat Int 497:33–46
Patterson C (1973) Interrelationships of holosteans. In: Greenwood PH, Miles RS, Patterson C (eds) Interrelationships of fishes. Academic Press, London, pp 233–305
Patterson C, Rosen DE (1977) Review of ichthyodectiform and other Mesozoic teleost fishes and the theory and practice of classifying fossils. Bull Am Mus Nat Hist 158:81–172
Paucǎ M (1931) Alosa nordmanni Antipa aus dem Pontien von Targu-Jiu. Bull Sec Sci Acad Roum 16:106–107
Paxton JR (1972) Osteology and relationships of the lanternfishes (Family Myctophidae). Bull Nat Hist Mus Los Angeles Cty, Sci 13:1–81
Přikryl T, Carnevale G (2017) Miocene bristlemouths (Teleostei: Stomiiformes: Gonostomatidae) from the Makrilia Formation, Ierapetra, Crete. C R Palevol 16:266–277
Přikryl T, Brzobohatý R, Gregorová R (2016a) Diversity and distribution of fossil codlets (Teleostei, Gadiformes, Bregmacerotidae): review and commentary. Palaeobiodiv Palaeoenviron 96:13–39
Přikryl T, Gómez de Soler B, Campeny G, Oms O, Roubach S, Blain H-A, Agustí J (2016b) Fish fauna of the Camp dels Ninots locality (Pliocene; Caldes de Malavella, province of Girona, Spain) – first results with notes on palaeoecology and taphonomy. Hist Biol 28:347–357
Přikryl T, Kania I, Krzemiński W (2016c) Synopsis of fossil fish fauna from the Hermanowa locality (Rupelian; Central Paratethys; Poland): current state of knowledge. Swiss J Geosci 109:429–443
Raulin V (1861) Description physique et naturelle de l’isle de Crète. Actes Soc Linnéenne Bord 23:321–383
Reichenbacher B, Böhme M (2004) Fish remains (otoliths, teeth) from the Miocene of Chios (Greece) – new palaeobiogeographic and palaeoclimatic results. Paper presented at the 5th International Symposium on Eastern Mediterranean Geology, Thessaloniki, Greece, 2004
Reichenbacher B, Gregorová R, Holcová K, Šanda R, Vukić J, Přikryl T (2018) Discovery of the oldest Gobius (Teleostei, Gobiiformes) from a marine ecosystem of early Miocene age. J Syst Palaeontol 16:493–513
Risso A (1810) Ichthyologie de Nice, ou histoire naturelle des poissons du Département des Alpes Maritimes. F. Schoell, Paris
Romano C, Koot MB, Kogan I, Brayard A, Minikh AV, Brinkmann W, Bucher H, Kriwet J (2016) Permian–Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution. Biol Rev 91:106–147
Rückert-Ülkümen N (1994) Zur systematischen Stellung einiger Clupeidae aus Thrakien, Türkei. Mitt Bayer Staatssammlung Paläontol Hist Geol 34
Rückert-Ülkümen N, Yiğitbaş E (2007) Pharyngeal teeth, lateral ethmoids, and jaw teeth of fishes and additional fossils from the late Miocene (late Khersonian/early Maeotian) of Eastern Paratethys (Yalova, near İstanbul, Turkey). Turk J Earth Sci 16:211–224
Sallan LC (2014) Major issues in the origins of ray-finned fish (Actinopterygii) biodiversity. Biol Rev 89:950–971
Sauvage MH-E (1873) Memoire sur la faune ichthyologigue de la periode Tertiare et plus specialement sur les poissons fossiles d’Oran et de Licata. Ann Sci Geol 4:1–272
Schultz O (1979) Supplementary notes on elasmobranch and teleost fish remains from the Korytnica Clays (Middle Miocene; Holy Cross Mountains, Central Poland). Acta Palaeontol Pol 29:287–293
Schultz O (2003) The Middle Miocene Fish Fauna (excl. otolithes) from Mühlbach am Manhartsberg and Grund near Hollabrunn, Lower Austria. Ann Naturhist Mus Wien 104A:185–193
Schultz O, Bellwood DR (2004) Trigonodon oweni and Asima jugleri are different parts of the same species Trigonodon jugleri, a Chiseltooth Wrasse from the lower and middle Miocene in Central Europe (Osteichthyes, Labridae, Trigonodontinae). Ann Naturhist Mus Wien 105A:287–305
Schultz O, Paunović M, von Summesberger H (1997) Der Nachweis von Coelodus (Osteichthyes, Pycnodontidae) im Turonien (Oberkreide) von Gams bei Hieflau, Steiermark, Österreich, und aus der Oberkreide von Kroatien und Italien. Ann Naturhist Mus Wien 98:73–141
Schultze HP (2008) Nomenclature and homologization of cranial bones in actinopterygians. In: Arratia G, Schultze HP, Wilson MVH (eds) Mesozoic fishes 4. Dr. Friedrich Pfeil, München, pp 23–48
Schwarzhans W, Carnevale G, Bannikov AF, Japundžić S, Bradić K (2017) Otoliths in situ from Sarmatian (Middle Miocene) fishes of the Paratethys. Part I: Atherina suchovi Switchenska, 1973. Swiss J Palaeontol 136:7–17
Sibert EC, Norris RD (2015) New age of fishes initiated by the Cretaceous−Paleogene mass extinction. Proc Natl Acad Sci 112:8537–8542
Smirnov VP (1936) Fishes of the North Caucasian Oligocene. Chernorechenskii District (in Russian). Travaux scientifiques de l’Université de l’Usbekistan 1:1–96
Sorbini C, Tyler JC (2004) Review of the fossil file fishes of the family Monacanthidae (Tetraodontiformes), Pliocene and Pleistocene of Europe, with a new genus, Frigocanthus, and two new species related to the recent Aluterus. Boll Mus Civico Storia Nat Verona 28:41–76
Stewart KM (2001) The freshwater fish of Neogene Africa (Miocene-Pleistocene): systematics and biogeography. Fish Fish 2:177–230
Stewart KM (2003) Fossil fish remains from the Pliocene Kanapoi Site, Kenya. In: Harris JM, Leakey MG (eds) Geology and vertebrate paleontology of the Early Pliocene site of Kanapoi, Northern Kenya. Natural History Museum of Los Angeles County, Los Angeles, pp 21–38
Stewart KM, Murray AM (2017) Biogeographic implications of fossil fishes from the Awash River, Ethiopia. J Vertebr Paleontol 37:e1269115
Symeonidis N (1969) Fossile Fische aus der Gegend von Ierapetra (Kreta). Ann Géol Pays Héllén 21:501–530
Symeonidis N, Schultz O (1973) Bemerkungen zur neogenen Fischfauna Kretas und Beschreibung zweier Fundstellen mit miozänen Mollusken, Echiniiden etc., Insel Kreta, Griechenland. Ann Naturhist Mus Wien 77:141–147
Tintori A, Sun Z, Ni P, Lombardo C, Jiang D, Motani R (2015) Oldest stem Teleostei from the late Ladinian (Middle Triassic) of Southern China. Riv Ital Paleontol Stratigr 121:285–296
Tripalo K, Japundžić S, Sremac J, Bošnjak M (2016) First record of Acanthuridae (surgeonfish) from the Miocene deposits of the Medvednica Mt. Geol Croat 69:201–204
Tyler JC, Bannikov AF (2000) A new species of the surgeon fish genus Tauichthys from the Eocene of Monte Bolca, Italy (Perciformes, Acanthuridae). Boll Mus Civico Storia Nat Verona 24:29–36
Tyler JC, Micklich NR (2011) A new genus and species of surgeon fish (Perciformes, Acanthuridae) from the Oligocene of Kanton Glarus, Switzerland. Swiss J Palaeontol 130:203–216
Unger F (1867) Die fossile Flora von Kumi auf der Insel Euboea. Denkschriften K Akad Wiss Math Naturwiss Kl 27:27–87
Valenciennes A (1861) Rapport sur les collections des espèces de Mammifères déterminées par leurs nombreux ossements fossiles rectieillis par M. Albert Gaudry, à Pikermi, près d’Athènes, pendant son voyage en Attique. C R Hebd Séances Acad Sci 52:1295–1300
Vasileiadou K, Böhme M, Neubauer TA, Georgalis GL, Syrides GE, Papadopoulou L, Zouros N (2017) Early Miocene gastropod and ectothermic vertebrate remains from the Lesvos Petrified Forest (Greece). Palaeontol Z 91:541–564
Vernygora O, Murray AM, Luque J, Ruge MLP, Fonseca MEP (2018) A new Cretaceous dercetid fish (Neoteleostei: Aulopiformes) from the Turonian of Colombia. J Syst Palaeontol 16:1057–1071
Vinther J (2015) A guide to the field of palaeo colour. BioEssays 37:643–656
Viohl G (1990) Piscivorous fishes of the Solnhofen lithographic limestone. In: Boucot AJ (ed) Evolutionary paleobiology of behavior and coevolution. Elsevier, Amsterdam, pp 287–303
Weems RE (1999) Part 4. Actinopterygian fishes from the Fisher/Sullivan site. In: Weems RE, Grimsley GJ (eds) Early Eocene vertebrates and plants from the Fisher/Sullivan site (Nanjemoy Formation) Stafford County, Virginia. Commonwealth of Virginia Department of Mines, Minerals and Energy Division of Mineral Resources, Charlottesville, pp 53–99
Weems RE, Edwards LE, Landacre B (2017) Geology and biostratigraphy of the Potomac River cliffs at Stratford Hall, Westmoreland County, Virginia. Field Guid 47:125–152
Weiler W (1943) Über einen pliozänen Fischrest aus Mazedonien. Z Deut Geol Ges 95:210–213
Wiley EO, Johnson GD (2010) A teleost classification based on monophyletic groups. In: Nelson JS, Schultze HP, Wilson MVH (eds) Origin and phylogenetic interrelationships of Teleosts. Dr. Friedrich Pfeil, Munich, pp 123–182
Woodward AS (1901) Catalogue of the fossil fishes in the British Museum (National History). Part IV. Containing the actinopterygian Teleostomi of the suborders Isospondyli (in part), Ostariophysi, Apodes, Percesoces, Hemibranchii, Acanthopterygii, and Anacanthini. British Museum (Natural History), London
Yabumoto Y, Uyeno T (1994) Late Mesozoic and Cenozoic fish faunas of Japan. Island Arc 3:255–269
Zhu M, Zhao W, Jia L, Lu J, Qiao T, Qu Q (2009) The oldest articulated osteichthyan reveals mosaic gnathostome characters. Nature 458:469–474
Acknowledgments
I am indebted to my former advisors and mentors G. Theodorou and P. Pavlakis (both NKUA) for supporting and encouraging me early to work on the fossil fishes of Greece, and beyond. S. Roussiakis (NKUA) is thanked for providing collection assistance and scientific discussion, and K. Agiadi (NKUA) is thanked for useful and stimulating discussions on the particulars of this chapter. Several colleagues are thanked for helping the author (re)assess some of the information presented here on the field. Listed in alphabetic order are: K. Agiadi (NKUA); A. Alexopoulos (NKUA); J. Carrillo-Briceño (Univ. Zurich); L. Cavin (MNHN, Geneva); D. Davesne (Oxford Univ.); I. Dimitriou; C. Fassoulas (NHM, Crete); A. Filipidi (Univ. Utrecht); C. Giamali (NKUA); G. Iliopoulos (Univ. Patras); E. Koskeridou (NKUA); M. Louvari (NKUA); S. Roussiakis (NKUA); G. Theodorou (NKUA); K Trikolas. Giorgio Carnevale (Univ. Torino), Charalampos Kevrekidis (LMU Munich) and C. Romano (Univ. Zurich) are thanked for their insightful reviews, which improved the quality of this work. This research was funded by a Swiss National Science Foundation early postdoctoral mobility grant (P2ZHP3_184216) to the author.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Appendix
Appendix
Tables with a detail list of the occurrences of ray-finned fishes in Greece.
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2022 The Author(s)
About this chapter

Cite this chapter
Argyriou, T. (2022). The Fossil Record of Ray-Finned Fishes (Actinopterygii) in Greece. In: Vlachos, E. (eds) Fossil Vertebrates of Greece Vol. 1. Springer, Cham. https://doi.org/10.1007/978-3-030-68398-6_4
Download citation
DOI: https://doi.org/10.1007/978-3-030-68398-6_4
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-68397-9
Online ISBN: 978-3-030-68398-6
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)