
Abstract
Fossil footprints are very useful palaeontological tools. Their features can help to identify their makers and also to infer biological as well as behavioural information. Nearly all the hominin tracks discovered so far are attributed to species of the genus Homo. The only exception is represented by the trackways found in the late 1970s at Laetoli, which are thought to have been made by three Australopithecus afarensis individuals about 3.66 million years ago. We have unearthed and described the footprints of two more individuals at Laetoli, who were moving on the same surface, in the same direction, and probably in the same timespan as the three found in the 1970s, apparently all belonging to a single herd of bipedal hominins walking from south to north. The estimated stature of one of the new individuals (about 1.65 m) exceeds those previously published for Au. afarensis. This evidence supports the existence of marked morphological variation within the species. Considering the bipedal footprints found at Laetoli as a whole, we can hypothesize that the tallest individual may have been the dominant male, the others smaller females and juveniles. Thus, considerable differences may have existed between sexes in these human ancestors, similar to modern gorillas.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
Forty Years of Research at Laetoli
Fossil skeletal elements can offer plenty of data on different aspects of human evolution and palaeobiology. However, the amount of information we can get about our ancestors can increase significantly through the study of fossil footprints. In fact, these ephemeral traces of life in the geological past can provide key palaeobiological insights on anatomy, locomotion biomechanics, body size, social behaviours, palaeoenvironments, and even reproductive strategies of extinct hominins (Falkingham et al. 2018). Unfortunately, due to the extremely peculiar taphonomic conditions that can lead to their preservation, fossil footprints are very rare. The hominin ichnological record is particularly poor (Bennett and Morse 2014), especially when compared to that of other vertebrate groups like dinosaurs. Nearly all the hominin tracks discovered so far are attributed to species of the genus Homo, with the outstanding exception of the record from Laetoli (Tanzania), dated to 3.66 million years ago (Ma).
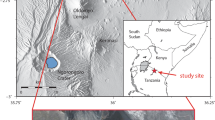
Laetoli is one of the most important palaeoanthropological sites in the world. It is located in northern Tanzania (Fig. 8.1) at the southern margin of the Serengeti plains, within the Ngorongoro Conservation Area (NCA), in which several other world-famous palaeoanthropological localities like Olduvai Gorge, Lake Ndutu, and Nasera Rock, are found.

Geographical location and site map; (a) location of Tanzania; (b) location of Laetoli in northern Tanzania; (c) plan view of Laetoli Locality 8 (Sites G and S); (d) plan view of the four test-pits excavated at Laetoli Site S. Dashed lines indicate uncertain contours. Hominin tracks in orange, equid tracks in dark green, rhinoceros track in red, giraffe tracks in light brown, and bird tracks in blue. (Modified from Masao et al. 2016)
The Laetoli stratigraphic sequence is composed of Plio-Pleistocene volcano-sedimentary deposits divided into five main lithological units, the Laetolil Beds, Ndolanya Beds, Olgol Lavas/Naibadad Beds, Olpiro Beds, and Ngaloba Beds, from bottom to top (Hay 1987). At the base, the Laetolil Beds make up most of the sedimentary sequence, with a thickness of more than 120 m (Ditchfield and Harrison 2011). They probably formed from tephra erupted from the extinct Sadiman volcano, located about 20 km to the east of Laetoli (Hay 1987; Mollel et al. 2011), although this hypothesis is questioned by some authors (Zaitsev et al. 2011, 2015). The Laetolil Beds are divided into two units, namely, the Lower and Upper Laetolil Beds (4.36–3.85 Ma and 3.85–3.63 Ma, respectively) (Hay 1987; Deino 2011). The latter consist of a series of aeolian and fall-out tuffs (Hay 1987) and are well known for their abundant palaeontological content (Harrison and Kweka 2011).
The palaeoanthropological relevance of the Laetoli area and of the Upper Laetolil Beds in particular is well known since the mid-1930s (Reck and Kohl-Larsen 1936; Kohl-Larsen 1943), but the site turned the attention of the academia and general public in the 1970s, with the discovery of the holotype and other remains of Australopithecus afarensis (Leakey et al. 1976; Johanson et al. 1978), as well as of the earliest bipedal hominin tracks in the world (Leakey and Hay 1979; Leakey and Harris 1987).
Mammal, bird, and insect prints and trails were identified by Mary Leakey and collaborators in 18 sites (labelled from A to R) out of 33 total palaeontological localities in the Laetoli area (Leakey 1987; Harrison and Kweka 2011; Musiba et al. 2008). The so-called Footprint Tuff, which corresponds to the lower part of Tuff 7 in the Upper Laetolil Beds’ stratigraphic sequence, hosts at least ten sublevels in which footprints are found (Hay 1987). Among these, hominin tracks were originally discovered at Site G (Locality 8). A short trackway of humanlike footprints was also unearthed at Site A (Locality 6) but was later attributed to a bear (Tuttle 2008). Site G footprints were referred to three individuals (G1, G2, G3) of different body sizes: the smaller G1 walked side by side on the left of the larger G2, while the intermediate-sized G3 superimposed its feet over those of G2 (Leakey 1981). These trackways are ascribed to Au. afarensis (White and Suwa 1987; Masao et al. 2016), which is the only hominin taxon found to date in the Upper Laetoli Beds (Harrison 2011).
Immediately after the first publication (Leakey and Hay 1979), the scientific and public interest in the Laetoli footprints has spread extraordinarily. Since then, they have been “mentioned in hundreds, if not thousands, of scientific works” (Jungers 2016), and a Google search for Laetoli footprints returns more than 66,000 results at the date of writing this contribution. In the first years after the discovery, the tracks were studied in several papers dealing with interactions between trackmakers (Leakey and Hay 1979; Leakey 1981), foot anatomy and locomotion (Day and Wickens 1980; White 1980; Charteris et al. 1981, 1982; Stern and Susman 1983), and depositional and palaeoenvironmental setting (Leakey and Hay 1979; Leakey 1981). In more recent years, with the development of new technologies and methods, many researchers worked on the key topic of locomotion of the Laetoli trackmakers by means of different approaches. These analyses led to conflicting views, with some authors (e.g. Raichlen et al. 2010; Crompton et al. 2012) inferring that the gait pattern of the Laetoli hominins was similar to that of modern humans, while others (e.g. Meldrum 2004; Schmid 2004; Bennett et al. 2009; Hatala et al. 2016) inferring that it was qualitatively and/or quantitatively different. However, regardless of the methods used, all the above studies are equally negatively affected by the fact that they are focused only on a limited number of G1 tracks. Although most of the G1 trackway is well preserved (unlike the overlapping footprints of G2 and G3), it belongs to the smallest of G individuals, which was very likely a juvenile (see “Laetoli Site S Footprints: Results and Implications”). Moreover, the original tracks are today buried under a protective cover (Feibel et al. 1996), and most of the studies were carried out on casts.
In light of the above, the recent discovery of new human footprints in Laetoli is of crucial importance for the knowledge of the anatomy, palaeobiology, and behaviour of Pliocene hominins.
The Discovery of Laetoli Site S
In 2014, two of us (F.T.M., E.B.I.) were commissioned to carry out a Cultural Heritage Impact Assessment (CHIA) aimed at evaluating the impact of a proposed new field museum in the area of Locality 8, that is, the palaeontological locality in which M. Leakey and co-authors discovered the first human tracks in the 1970s. According to Tanzania’s Environmental Management Act (United Republic of Tanzania 2004), the CHIA is part of the Environmental and Sociological Impact Assessment (ESIA), which is a mandatory evaluation process expected to address the impact of a certain development project (e.g. infrastructure construction) on the environment, landscape, and social context (Ichumbaki and Mjema 2018). In particular, the CHIA is focused on the possible impacts on cultural heritage, both extinct (e.g. archaeological and palaeontological record) and extant (e.g. ethno-anthropological context).
Specific objectives of the CHIA were:
-
1.
Salvaging as much of the threatened heritage as possible through surface collection and excavation
-
2.
Preliminary analysis of the archaeological and palaeontological material rescued
-
3.
Packing the material and presenting it to Ngorongoro Conservation Area Authority (NCAA) for curation and storage
-
4.
Proposing mitigation measures including immediate conservation of special features encountered in the fieldwork process and proposing an appropriate monitoring schedule
The CHIA assignment was accomplished through two main fieldwork seasons. During the first season (June 21–30, 2014), the team of archaeologists, cartographers, conservators, and skilled workers aimed at obtaining an overall picture of the cultural heritage features in the area impacted by the project. In particular, the team surveyed a wide area within 500 m radius from the Site G trackways, i.e. the core area of the proposed museum project. The second season (September 13 to October 22, 2014) focused on the area of maximum impact, i.e. the surroundings of Site G. Sixty-two 2 × 2 m test-pits (each corresponding to about 2% of the total surface) were randomly positioned within a grid and carefully excavated down to the Footprint Tuff and sometimes deeper. If necessary, in case of particularly significant finds (see below), some pits were enlarged compared to the standard of 2 × 2 m.
About 150 m to the south of Site G, the team unearthed 14 hominin tracks associated with abundant tracks of other vertebrates. Footprints were found in three test-pits, respectively labelled L8, M9, and TP2 from north to south. The original square shape of L8 was modified soon after the discovery of the first bipedal tracks in order to follow the trail, thus obtaining a quite irregular shape of this test-pit (southern side, 2 m; western oblique side, 4 m). M9 was excavated some 14 m to the SSE of L8 and kept the standard size of 2 × 2 m. Following the putative alignment of the trackway, a third smaller test-pit, TP2 (1 × 1.2 m), was excavated at some 8 m to the SSE of M9. Finally, test-pit M10 (2 × 3 m) was excavated about 15 m to the east of M9 to assess the occurrence of other interesting tracks (Fig. 8.1).
Once the presence of the new tracks has been ascertained and with the aim of characterizing the printed surface with a multidisciplinary approach, the Tanzanian-Italian research group was established, pivoting on a collaboration already started for years in Olduvai Gorge (e.g. Cherin et al. 2016). The new team reopened the four test-pits in September 2015. Fourteen hominin tracks in different preservation states always associated with tracks of other vertebrates were unearthed in test-pits L8, M9, and TP2 (Masao et al. 2016). All these prints are clearly referable to a single trail, with an estimated total length of 32 m and trending SSE to NNW. Following the code used for the other footprint sites in Laetoli (Leakey 1981; Leakey 1987; Harrison and Kweka 2011), the new site was identified as Site S, and the new tracks are attributed to individual S1 (footprint numbers S1-1–7 in L8, S1-1–4 in M9, and S1-1–2 in TP2) (Fig. 8.2). An additional track referable to a second individual (S2) was found in the SW corner of TP2. Conversely, only non-hominin footprints were recorded in M10 (Masao et al. 2016).

Test-pit L8. Photo (left) and shaded 3D photogrammetric model (right) of the southern part of the printed surface, with close-ups (bottom) of the four footprints
Survey of Laetoli Site S: A Case Study for Photogrammetry Application in Extreme Environments
Modern developments in computing power, rendering software, and hardware availability allowed a rapid and widespread diffusion of photogrammetry techniques in Earth Sciences and other disciplines. The majority of these techniques are based on Structure from Motion (SfM) algorithms (e.g. Luhmann et al. 2013; Mallison and Wings 2014; James et al. 2017). Among others, SfM techniques have been used in recent years to study river systems (e.g. Marteau et al. 2017), landslide dynamics and volumes (e.g. Stumpf et al. 2015), cliff morphology (e.g. Warrick et al. 2017), active fault structure and dynamics (e.g. Johnson et al. 2014), geobody architecture of depositional systems (e.g. Mancini et al. 2019), as well as ichnological contexts with human tracks (e.g. Rüther et al. 2012; Bennett et al. 2016; Bustos et al. 2018; Helm et al. 2018; Zimmer et al. 2018; Romano et al. 2019). SfM algorithms allow obtaining three-dimensional models from a series of overlapping pictures taken from different camera positions. The obtained high-resolution models can be easily shared between researchers and can be used for detailed qualitative descriptions and accurate quantitative analyses at the sub-mm-scale (Mallison and Wings 2014). However, in order to get affordable data from SfM, field data must be supported by accurate in situ topographic measurements.
The photogrammetric survey of the new footprint Site S was carried out in an extreme environmental context, characterized by unfavourable climatic conditions, need for light equipment, and little time available. Therefore, we had to set up a working procedure that, despite these problems, could lead to good results in terms of accuracy and precision and could also serve as a reference for other scientific activities in similar contexts (Menconero et al. 2019).
The Laetoli area is located over a wide plateau at about 1700 m above sea level, to the west of the volcanic complex of Sadiman (2870 m), Lemagrut (3135 m), and Oldeani (3200 m), and north to the Lake Eyasi basin. The plateau is characterized by a tabular or slightly corrugated morphology. In some areas, the landscape is more articulated due to the presence of valleys, gorges, and gullies originated by the action of wind and small streams, whose erosional energy is very intense during the dry season (May–October) and rainy season (November–May), respectively. The current vegetation cover is primarily determined by topography, soil composition, and climate (Anderson 2008) but is also influenced by natural and anthropic fires, as well as by the grazing activity of the extremely abundant wild herbivore mammals and domestic livestock (cattle, sheep, and goats) bred by local tribes with nomadic/semi-nomadic pastoral economy (Holdo et al. 2009). The vegetation mainly includes thorny thickets and dry bushland, consisting of shrubby and arboreal deciduous species of the genera Vachellia, Senegalia, and Commiphora, associated with several forms of grasses (e.g. Sporoboro, Digitaria, Themeda, Aristida, Brachiaria, Cenchrus) (Herlocker and Dirschl 1972; Andrews and Bamford 2008). The presence of numerous and densely distributed thorny plants can cause numerous problems during research activities. The wild animal community of the Laetoli area is still abundant and diverse, thanks to the low human demographic density, the presence of impenetrable thorny xerophilous shrublands, and the protection measures by the NCAA. Among reptiles, several snakes – such as the black mamba (Dendroaspis polylepis), green mamba (D. angusticeps), Egyptian cobra (Naja haje), spitting cobra (N. nigricollis), and puff adder (Bitis arietans) – can be potentially very dangerous to humans. The same goes for some large-sized mammals, like the African bush elephant (Loxodonta africana), spotted hyena (Crocuta crocuta), leopard (Panthera pardus), and African lion (Panthera leo). With regard to scientific activities, the low demographic density makes very hard to get consumer goods like food, water, and materials, which have to be bought in larger villages, such as Karatu, about 4-hour drive from Laetoli. As for the hygienic and sanitary conditions, thanks to the high average altitude and predominantly dry climate of the plateau, the whole Laetoli area is less affected by tropical pathologies that are common in the nearby low-altitude areas. However, especially in wet areas close to perennial small rivers, there are small populations of hematophagous dipterans like the mosquitos Anophele (vector of Plasmodium, responsible for malaria) and Aedes (carrier of various viruses responsible for serious diseases), as well as some horseflies (Tabanidae) and blackflies (Simuliidae), which can cause painful bites and severe skin irritations.
The above environmental conditions (climate, vegetation, fauna) are added to logistical complications (short time available, problems related to natural lighting, lack of electricity, long car trips along rough trails) in making extremely difficult the fieldwork in Laetoli (and similar contexts). Under these conditions, clear goals for the fieldwork are necessary. In our case, the work at Site S was aimed at obtaining 3D models of the new tracks for documentation and morphometric analysis. We chose the SfM photogrammetry technique, thanks to its technical advantages (relatively short time of data acquisition and processing, light and handy equipment, reduced costs) and excellent results in terms of resolution (Westoby et al. 2012).
Each test-pit (L8, M9, TP2, and M10) was entirely surveyed at lower resolution (Fig. 8.3), and then detailed 3D models of some inner portions (single tracks or groups of close prints) were acquired (Fig. 8.4). Targets of the control point system were immediately positioned after excavation. We placed four perimeter targets at the corner of each test-pit and four inner targets around each subarea surveyed in detail (14 in L8, 10 in M9, 14 in TP2, and 14 in M10). For the measurement of the control points, we used a measuring tape and a water level, which are lighter and easier to handle than a total station theodolite. We selected these low-tech tools also considering (1) measuring only four points for each test-pit to scale the general 3D models and (2) aligning the detailed 3D models of the single footprints to the general models using the coordinates of the inner targets. For the perimeter target measurements, we placed two rods equipped with a spherical level on successive pairs of targets, and we marked points at the same height on the rods for each pair by using the water level device. The vertical distance between these points and the targets, as well as their mutual distance, were recorded. Repeating this process for all pairs of targets, the relative plan position and the height of the control points were determined respectively by trilateration and levelling. A preliminary accuracy check was carried out by means of trilateration graphic rules in plan and with the method of successive levelling for heights. By assigning a z-coordinate to the first control point, all subsequent coordinates were derived from addition and subtraction of heights between two successive points. The check was performed by computing the sum of all height differences and by verifying that the obtained value was close to zero. Finally, the error obtained in each test-pit was distributed to every z-coordinate of the points, in order to minimize it.

Test-pit L8; (a) eidotype; (b) shaded model; (c) textured model; (d) textured and shaded model; (e) drawing; (f) density of the point cloud by determining the number of nearest neighbours in a sphere with 0.5 cm radius

Three footprints from test-pit L8; (a) textured model; (b) textured and shaded model; (c) shaded model; (e) density of the point cloud by determining the number of nearest neighbours in a sphere with 0.5 cm radius
Photographic acquisition was performed with a DSLR camera, sometimes fixed on a 3-m-long telescopic rod for photographic shots from the top downwards. With regard to scene lighting, since we had no possibility to control light intensity and direction, we tried to reduce shadows by shooting especially during the central hours of the day (i.e. with subvertical sun rays). However, this was not always possible due to the excavation schedule and little time available. Therefore, we had to address the problem of high-contrast shadows in post-processing.
The texture resolution control of 3D models, namely, the Ground Sampling Distance (GSD), can be performed a priori using geometric formulas. The calculation is based on the principle of similar triangles, which are found in the geometry of the shooting. The variables are the size of the sensor (Sw) and the focal length of the camera (Fl), the size in pixel of the images (Iw), and the distance (H). The triangle with the base Sw and height Fl is similar to the triangle which has the base Gw (width of the image on the ground) and height H; consequently, the two triangles have proportional respective sides (Sw: Gw = Fl: H). The GSD is the ratio between Gw and Iw multiplied by 100 (GSD = Gw/Iw × 100). Connecting the proportion with the formula of GSD, the final formula GSD = (Sw × H × 100): (Fl × Iw) is obtained. Among the variables, the one that can be easily managed is the distance H, since all the others depend on the photographic equipment available (Menconero et al. 2019).
It is impossible to know a priori the density of the point cloud coming from a photogrammetric process. As for the Laetoli footprints, the goal was to obtain a texture resolution less than 0.1 cm/px. This was achieved by choosing suitable shooting distance both for the whole test-pits and individual footprints. More than 2000 photos were taken in three working days, for a total of about 50 GB. Especially when working in remote areas, it is important not to economize on shots and possibly select them a posteriori.
Data processing started with checking topographic measurements in plan and height, which is preliminary to the definition of the control point coordinates. The trilateration method was used to obtain x, y coordinates of the control points in plan. For each test-pit, six measurements were taken at the same height: the length of the four sides of the perimeter and the length of the two diagonals. Redundant measurements were used to compute the errors. In addition to a preliminary graphical control by CAD software, we used an automatic calculation software to adjust a new set of x, y coordinates and heights of the control points by least squares technique. The residues of adjustments never exceeded 10 mm, which is fully acceptable considering the size of the test-pits. We used the adjusted x, y, z coordinates of the control points to scale and locate in the 3D space the SfM models. A check on point cloud density was also carried out by a software for 3D point cloud and mesh processing and analysis. The average density found in the Laetoli point clouds is around 20 points/cm3 for the test-pits and 1500 points/cm3 for the detailed footprints (Masao et al. 2016; Menconero et al. 2019).
The 3D models obtained by SfM were also used in the morphometric analysis of the hominin tracks. We used a contouring and modelling software that transforms x, y, z data into maps. The x, y, z-format files were imported into the software and transformed into grid files. The software uses randomly spaced x, y, z data to create regularly spaced grids composed of nodes with x, y, z coordinates. The Triangulation with Linear Interpolation gridding method was applied, because it works better with data that are evenly distributed over the grid area. This method creates network of triangles with no edge intersection starting from data points and computes new values along the edges. The grid spacing was set at 1 mm. Standard morphometric measurements (footprint length, footprint max width, footprint heel width, angle of gait, step length, and stride length) were taken from contour maps and compared with those taken manually both on the original tracks during the fieldwork and on 1:1 scale sketches of the test-pits, hand-drawn on transparent plastic sheets (Masao et al. 2016).
Laetoli Site S Footprints: Results and Implications
The detailed analysis of the new bipedal footprints at Site S started trying to frame this outstanding finding into the stratigraphic context of the Upper Laetolil Beds. A detailed sequence analysis of the excavation profiles at Site S and extended geological observations in the whole Laetoli area were performed. In particular, we tried to reconstruct the stratigraphic relationships between the footprint-bearing units of Site S and Site G, using both field observation and literature descriptions of the sequence outcropping in the original site.
The Laetoli Footprint Tuff is part of Tuff 7 together with the overlying Augite Biotite Tuff and can be divided into a lower and an upper unit. These can be respectively subdivided into 14 and 4 sublevels. Tracks are found on eight sublevels within the lower unit and two within the upper one (Leakey and Hay 1979; Hay and Leakey 1982; Hay 1987). In particular, hominin tracks at Site G are located on the top of horizon B, namely, on sublevel 14 of the Footprint Tuff lower unit (Hay and Leakey 1982; White and Suwa 1987). Though with some local differences presumably due to lateral variability, we found that the Site S sequence corresponded quite well with the original description of the Footprint Tuff stratigraphy provided by Hay (1987). In particular, we observed that the Site S tracks were printed on the top of the lower subunit of the Footprint Tuff, corresponding to the aforementioned horizon B (Masao et al. 2016). Consequently, our data indicate with reasonable confidence that the footprints of S1 and S2 lie on the same stratigraphic position as those at Site G. Considering that (1) Tuff 7 includes a sequence of several sublevels originated by distinct volcanic eruptions close in time, and that its overall deposition time is estimated in weeks (Hay and Leakey 1982; Hay 1987), (2) trackways from Site G and Site S show almost the same orientation, and (3) all trackmakers were moving approximately at the same moderate speed (see below), we hypothesized that the tracks from the two sites were left by a homogeneous group of hominins walking together on the same palaeosurface (Masao et al. 2016).
The overall morphology of the S1 tracks fits those from Site G and is particularly similar to the prints of G2, namely, the larger individual (Robbins 1987): the heel is oval shaped and is pressed deeply into the ground; the medial side of the arch is higher than the lateral one; the ball region is oriented at an angle of about 75° with respect to the longitudinal axis of the foot and is delimited anteriorly by a transversal ridge, formed when the toes gripped the wet volcanic ash and pushed it posteriorly; the adducted hallux extends more anteriorly than the other toes in all visible footprints; unfortunately, no clear distinction among the other toes is visible. The only preserved track of S2 is abnormally widened in the anterior part, probably due to a lateral slipping of the foot before the toe-off and/or to taphonomic factors related to the fragmentation of the Footprint Tuff.
Stride length was used to estimate the walking speed of the Laetoli trackmakers. Mean values of about 0.44–0.9 m/s were obtained, depending on the computing method (Alexander 1976; Dingwall et al. 2013). The average length of the S1 tracks is 261 mm (range 245–274 mm). Lower average values were measured for the three individuals at Site G: 180 mm for G1, 225 mm for G2, and 209 mm for G3 (Leakey 1981; Tuttle 1987), although a study of some G footprint casts based on digital methods (Bennett et al. 2016) suggested higher values for G1 (193 mm) and G3 (228 mm). Stature was computed first with Tuttle’s (1987) approach, which is based on the ratio between foot length and stature in modern humans (foot length in Homo sapiens is generally about 14–16% of stature). We also estimated stature using the two methods published by Dingwall et al. (2013). The first is based on regressions of stature by footprint length in modern Daasanach people from Lake Turkana (Kenya); the second – which we considered more reliable because it is not influenced by modern human data – is based on the foot/stature ratio known for Au. afarensis. Similarly, we estimated the body mass of the trackmakers by means of the regression equation that relates footprint area to body mass in H. sapiens, as well as of the equation based on the ratio between foot length and body mass in Au. afarensis (Dingwall et al. 2013). All the above data were also measured/calculated for G individuals, using a 3D model of a first-generation cast of the southern portion of the Site G trackways.
Our results showed that no matter which method is employed to estimate stature and body mass, S1 and S2 were taller and had a larger body mass than the G individuals (S1, 161–168 cm/41.3–48.1 kg; S2, 142–149 cm/36.5–42.4 kg; G1, 111–116 cm/28.5–33.1 kg; G2, 139–145 cm/35.6–41.4 kg; G3, 129–135 cm/33.1–38.5 kg) (Masao et al. 2016). These results extended the dimensional range of the Laetoli trackmakers and identified S1 as a large-sized individual, probably a male. The stature of about 165 cm for S1 is remarkable and exceeds those estimated to date for any australopithecine. The stature of S1 falls within the maximum range of modern Homo sapiens and also fits the available Homo erectus sensu lato estimates based on both skeletal remains and footprints. The body mass range estimated for S1 falls within the range of male Au. afarensis (40.2–61.0 kg) (Grabowski et al. 2015).
Our results provided independent evidence for the occurrence of large-sized individuals among hominins as ancient as 3.66 Ma and supported a nonlinear evolutionary trend in hominin body size (Jungers et al. 2016). Moreover, ascribing the S1 tracks to a possible male allowed reconsidering the sex and age of the other Laetoli individuals. According to our body-mass estimations, G1 and G3 fall within the range of putative Au. afarensis females, whereas G2 and S2 span across the upper female and the lower male ranges, which are estimated at 25.5–38.1 and 40.2–61.0 kg, respectively (Grabowski et al. 2015). A possible reconstruction is that the Laetoli individuals are S1, a male; G2 and S2, females; and G1 and G3, smaller females or juvenile individuals.
Both the new composition of the group and the impressive body size difference point to a considerable sexual dimorphism in Au. Afarensis (Fig. 8.5), as hypothesized by many scholars on the basis of skeletal remains (e.g. Johanson and White 1979; Kimbel and White 1988; McHenry 1991; Richmond and Jungers 1995; Lockwood et al. 1996; Plavcan et al. 2005; Harmon 2006; Gordon et al. 2008). In turn, this view supports social organization and reproductive strategies closer to the polygynous gorillas (Harcourt and Stewart 2007) than to other moderately dimorphic species, like the promiscuous chimpanzees or the extant and, possibly, extinct humans (Masao et al. 2016).

Minimum and maximum estimated statures of selected fossil hominins by species and locality over time for the interval 4–1 million years
Laetoli Footprints: Perspectives
The recent discovery of Laetoli Site S footprints, after about 40 years from the pioneering works by Mary Leakey and colleagues at Site G, has achieved a remarkable media coverage and has drawn the attention of the scientific community.
Raichlen and Gordon (2017) used proportional toe depth (i.e. a measure of the difference between toe depth and hindfoot depth in tracks) as a proxy to get information about the locomotor style of the Laetoli trackmakers. They confirmed that the footprints from Site S are overall very similar to those from Site G, thus supporting the hypothesis that bipedal locomotion in Au. afarensis was more similar to modern-human-like extended-limb pattern than to chimpanzee-like bent-knee-bent-hip pattern (Raichlen et al. 2010).
DeSilva et al. (2019) included data on the Laetoli footprints from Sites G and S in their comprehensive review of Plio-Pleistocene hominin foot evolution.
In their very interesting work, Villmoare et al. (2019) inferred data on sexual dimorphism in H. erectus s.l. through the analysis of fossil footprints from Ileret, Kenya (about 1.5 Ma). Their results are in perfect agreement with ours in the recognition of a gorilla-like high level of dimorphism in Au. afarensis from Laetoli. These data are in contrast with those obtained for the Ileret sample, in which footprints show a much lower degree of sexual dimorphism, although slightly higher than that of modern humans. According to the authors, this would suggest that by 1.5 million years ago, at least H. erectus s.l. had transitioned away from polygyny (Villmoare et al. 2019).
Following a completely different line of research, the original contribution by Ichumbaki et al. (2019) addressed the topic of local community’s interpretations of the Laetoli hominin footprints. For the first time, the authors documented narratives of Maasai (i.e. local people living in the Laetoli area) dealing with their perceptions on what the footprints are and to whom they belong. The Maasai people connect Laetoli footprints to the tale of Lakalanga, a strong hero who helped them to win a battle against a neighbouring community. According to the story – which is consolidated into the local community oral tradition – Lakalanga was so big that wherever he walked, he left visible tracks on the ground. Thus, the discovery of the large-sized footprints at Site S has offered a further confirmation to the Maasai that the hero warrior Lakalanga really existed (Ichumbaki et al. 2019).
The aforementioned papers represent a synthetic selection of those in which Site S footprints have been studied/mentioned after their recent description (Masao et al. 2016). However, besides these research contributions, the discovery of Site S calls on the whole international scientific community to question itself on the challenging issue of conservation. Our fieldwork at Laetoli in 2014–2015 showed us the relevance and peculiarities of the site. Besides the good preservation of the footprints and their outstanding scientific significance, we could also verify the aggressiveness of the East African environment on the ichnological record. Through qualitative observations of the conservation status of the Footprint Tuff, we could ascertain how the disruptive action of weathering, flora, and fauna is threatening the footprints even before excavation. This is jeopardizing a unique piece of cultural heritage that is still largely unknown. Similar concerns were highlighted by several authors (Feibel et al. 1996; Getty Conservation Institute 1996; Agnew and Demas 1998) after the assessment of the state of conservation of the Site G footprints, which in the 1990s had been the subject of a project of consolidation, reburial, and protection coordinated by the Getty Conservation Institute and Tanzanian Antiquities Division (Musiba et al. 2012). Further analyses are necessary at Site S to address the crucial issue of the conservation of this invaluable palaeontological heritage. Large portions of the printed surfaces unearthed in the test-pits are already severely threatened by natural agents and could quickly disappear even if unexcavated (Masao et al. 2016). Fractures are bringing to disintegration of part of the Footprint Tuff into small cubic blocks. Roots are displacing the stratigraphic sequence and are opening preferential ways for water to penetrate the substrate and for arthropods to dig burrows in the ground. Therefore, keeping the situation as it is may not be the right way to preserve the site, because unexcavated footprints may be saved from weathering if they are excavated and properly treated. At the same time, we are aware that any excavation without a clear understanding of the physical, chemical, and biological risks to which the Footprint Tuff is exposed should not be undertaken. A modern project aimed at the excavation, conservation, and valorization of the Laetoli tracks must be preceded by a multidisciplinary study of environmental data such as local microclimate, temperature and humidity variations, rainfall, dominant wind, geological and petrographic characteristics of the substrate, interstitial water composition and flow, physical damage due to trespassing of livestock and wild animals, and local vegetation composition and its change. Future plans must include a programme of continuous monitoring of the footprints, especially with the involvement of the local community (Ichumbaki et al. 2019).
Once established a comprehensive plan of conservation and valorization of the Laetoli footprints in close collaboration with all the involved Tanzanian (e.g. Ministry of Natural Resources and Tourism, NCAA) and international (e.g. UNESCO) institutions, new systematic excavations can be carried out. This would allow unearthing the entire S1 and S2 trackways and also opening up the intriguing possibility of discovering tracks of other individuals. This new research would noticeably improve the available dataset on the foot anatomy, locomotor pattern, body size variation, social structure, and behaviour of the Laetoli australopithecines.
References
Agnew, N., & Demas, M. (1998). Preserving the Laetoli footprints. Scientific American, 262, 44–55.
Alexander, R. M. (1976). Estimates of speeds of dinosaurs. Nature, 261, 129–130.
Anderson, T. M. (2008). Plant compositional change over time increase with rainfall in Serengeti grasslands. Oikos, 117, 675–682.
Andrews, P., & Bamford, M. (2008). Past and present vegetation ecology of Laetoli, Tanzania. Journal of Human Evolution, 54, 78–98.
Bennett, M. R., & Morse, S. A. (2014). Human footprints: Fossilised locomotion? New York: Springer.
Bennett, M. R., Harris, J. W., Richmond, B. G., Braun, D. R., Mbua, E., Kiura, P., et al. (2009). Early Hominin foot morphology based on 1.5-million-year-old footprints from Ileret, Kenya. Science, 323, 1197–1201.
Bennett, M. R., Reynolds, S. C., Morse, S. A., & Budka, M. (2016). Laetoli’s lost tracks: 3D generated mean shape and missing footprints. Scientific Reports, 6, 21916.
Bustos, D., Jakeway, J., Urban, T. M., Holliday, V. T., Fenerty, B., Raichlen, D. A., et al. (2018). Footprints preserve terminal Pleistocene hunt? Human-sloth interactions in North America. Science Advances, 4, eaar7621.
Charteris, J., Wall, J. C., & Nottrodt, J. W. (1981). Functional reconstruction of gait from the Pliocene hominid footprints at Laetoli, northern Tanzania. Nature, 290, 496–498.
Charteris, J., Wall, J. C., & Nottrodt, J. W. (1982). Pliocene hominid gait: New interpretations based on available footprint data from Laetoli. American Journal of Physical Anthropology, 58, 133–144.
Cherin, M., Iurino, D. A., Njau, J. K., & Masao, F. T. (2016). New material of hyaenids (Mammalia, Carnivora) from Olduvai Gorge, Tanzania (Early Pleistocene). Bollettino della Società Paleontologica Italiana, 55, 1–9.
Crompton, R. H., Pataky, T. C., Savage, R., D’Août, K., Bennett, M. R., Day, M. H., et al. (2012). Human-like external function of the foot, and fully upright gait, confirmed in the 3.66 million year old Laetoli hominin footprints by topographic statistics, experimental footprint-formation and computer simulation. Journal of the Royal Society Interface, 9, 707–719.
Day, M. H., & Wickens, E. H. (1980). Laetoli Pliocene hominid footprints and bipedalism. Nature, 286, 385–387.
Deino, A. L. (2011).40Ar/39Ar dating of Laetoli, Tanzania. In T. Harrison (Ed.), Paleontology and geology of Laetoli: Human evolution in context (Vol. 1, pp. 77–97). New York: Springer.
DeSilva, J., McNutt, E., Benoit, J., & Zipfel, B. (2019). One small step: A review of Plio-Pleistocene hominin foot evolution. American Journal of Physical Anthropology, 168(S67), 63–140.
Dingwall, H. L., Hatala, K. G., Wunderlich, R. E., & Richmond, B. G. (2013). Hominin stature, body mass, and walking speed estimates based on 1.5 million-year-old fossil footprints at Ileret, Kenya. Journal of Human Evolution, 64, 556–568.
Ditchfield, P., & Harrison, T. (2011). Sedimentology, lithostratigraphy and depositional history of the Laetoli area. In T. Harrison (Ed.), Paleontology and geology of Laetoli: Human evolution in context (Vol. 1, pp. 47–76). New York: Springer.
Falkingham, P. L., Bates, K. T., Avanzini, M., Bennett, M., Bordy, E. M., Breithaupt, B. H., et al. (2018). A standard protocol for documenting modern and fossil ichnological data. Palaeontology, 61, 469–480.
Feibel, C. S., Agnew, N., Latimer, B., Demas, M., Marshall, F., Waane, S. A. C., et al. (1996). The Laetoli hominid footprints – A preliminary report on the conservation and scientific restudy. Evolutionary Anthropology, 5, 149–154.
Getty Conservation Institute. (1996). Laetoli project: Conservation of the hominid trackway site at Laetoli, Tanzania: Report on the 1995 field season. Los Angeles: Getty Conservation Institute.
Gordon, A. D., Green, D. J., & Richmond, B. G. (2008). Strong postcranial size dimorphism in Australopithecus afarensis: Results from two new resampling methods for multivariate data sets with missing data. American Journal of Physical Anthropology, 135, 311–328.
Grabowski, M., Hatala, K. G., Jungers, W. L., & Richmond, B. G. (2015). Body mass estimates of hominin fossils and the evolution of human body size. Journal of Human Evolution, 85, 75–93.
Harcourt, A. H., & Stewart, K. J. (2007). Gorilla society: Conflict, compromise, and cooperation between the sexes. Chicago/London: University of Chicago Press.
Harmon, E. H. (2006). Size and shape variation in Australopithecus afarensis proximal femora. Journal of Human Evolution, 51, 217–227.
Harrison, T. (2011). Hominins from the Upper Laetolil and Upper Ndolanya Beds, Laetoli. In T. Harrison (Ed.), Paleontology and geology of Laetoli: Human evolution in context (Vol. 2, pp. 141–188). New York: Springer.
Harrison, T., & Kweka, A. (2011). Paleontological localities on the Eyasi plateau, including Laetoli. In T. Harrison (Ed.), Paleontology and geology of Laetoli: Human evolution in context (Vol. 1, pp. 17–45). New York: Springer.
Hatala, K. G., Demes, B., & Richmond, B. G. (2016). Laetoli footprints reveal bipedal gait biomechanics different from those of modern humans and chimpanzees. Proceedings of the Royal Society B, 283, 20160235.
Hay, R. L. (1987). Geology of the Laetoli area. In M. D. Leakey & J. M. Harris (Eds.), Laetoli: A Pliocene site in northern Tanzania (pp. 23–47). Oxford: Clarendon.
Hay, R. L., & Leakey, M. D. (1982). The fossil footprints of Laetoli. Scientific American, 246, 50–57.
Helm, C. W., McCrea, R. T., Cawthra, H. C., Lockley, M. G., Cowling, R. M., Marean, C. W., et al. (2018). A new Pleistocene hominin tracksite from the Cape south Coast, South Africa. Scientific Reports, 8, 3772.
Herlocker, D. J., & Dirschl, H. J. (1972). Vegetation of Ngorongoro conservation area, Tanzania. Ottawa: Canadian Wildlife Service.
Holdo, R. M., Sinclair, A. R. E., Dobson, A. P., Metzger, K. L., Bolker, B. M., Ritchie, M. E., et al. (2009). A disease-mediated trophic cascade in the Serengeti and its implications for Ecosystem C. PLoS Biology, 7, e1000210.
Ichumbaki, E. B., & Mjema, E. (2018). The impact of small-scale development projects on archaeological heritage in Africa: The Tanzanian experience. Journal of Conservation and Management of Archaeological Sites, 2, 18–34.
Ichumbaki, E. B., Cherin, M., Masao, F. T., & Moggi Cecchi, J. (2019). Local people’s interpretations of the hominin footprints at Laetoli, Tanzania. Journal of Community Archaeology & Heritage, 6, 122–138.
James, M. R., Robson, S., & Smith, M. W. (2017). 3-D uncertainty-based topographic change detection with structure-from-motion photogrammetry: Precision maps for ground control and directly georeferenced surveys. Earth Surface Processes and Landforms, 42, 1769–1788.
Johanson, D. C., & White, T. D. (1979). A systematic assessment of early African hominids. Science, 203, 321–330.
Johanson, D. C., White, T. D., & Coppens, Y. (1978). A new species of the genus Australopithecus (Primates: Hominidae) from the Pliocene of eastern Africa. Kirtlandia, 28, 1–14.
Johnson, K., Nissen, E., Saripalli, S., Arrowsmith, J. R., McGarey, P., Scharer, K., et al. (2014). Rapid mapping of ultrafine fault zone topography with structure from motion. Geosphere, 10, 969–986.
Jungers, W. L. (2016). These feet were made for walking. eLife, 5, e22886.
Jungers, W. L., Grabowski, M., Hatala, K. G., & Richmond, B. G. (2016). The evolution of body size and shape in the human career. Philosophical Transactions of the Royal Society B, 371, 20150247.
Kimbel, W. H., & White, T. D. (1988). Variation, sexual dimorphism and the taxonomy of Australopithecus. In F. E. Grine (Ed.), Evolutionary history of the “Robust” Australopithecines (pp. 175–192). New York: Aldine de Gruyter.
Kohl-Larsen, L. (1943). Auf den Spuren des Vormenschen. Stuttgart: Strecker und Schröder.
Leakey, M. D. (1981). Tracks and tools. Philosophical Transactions of the Royal Society B, 292, 95–102.
Leakey, M. D. (1987). Introduction. In M. D. Leakey & J. M. Harris (Eds.), Laetoli: A Pliocene site in Northern Tanzania (pp. 1–22). Oxford: Clarendon.
Leakey, M. D., & Harris, J. M. (1987). Laetoli: A Pliocene site in Northern Tanzania. Oxford: Clarendon.
Leakey, M. D., & Hay, R. L. (1979). Pliocene footprints in the Laetolil Beds at Laetoli, northern Tanzania. Nature, 278, 317–323.
Leakey, M. D., Hay, R. L., Curtis, G. H., Drake, R. E., Jackes, M. K., & White, T. D. (1976). Fossil hominids from the Laetolil Beds. Nature, 262, 460–466.
Lockwood, C. A., Richmond, B. G., Jungers, W. L., & Kimbel, W. H. (1996). Randomization procedures and sexual dimorphism in Australopithecus afarensis. Journal of Human Evolution, 31, 537–548.
Luhmann, T., Robson, S., Kyle, S., & Boehm, J. (2013). Close-range photogrammetry and 3D imaging. Berlin/Boston: Walter de Gruyter.
Mallison, H., & Wings, O. (2014). Photogrammetry in paleontology: A practical guide. Journal of Paleontological Techniques, 12, 1–31.
Mancini, A., Capezzuoli, E., Erthal, M., & Swennen, R. (2019). Hierarchical approach to define travertine depositional systems: 3D conceptual morphological model and possible applications. Marine and Petroleum Geology, 103, 549–563.
Marteau, B., Vericat, D., Gibbins, C., Batalls, R. J., & Green, D. R. (2017). Application of structure-from-motion photogrammetry to river restoration. Earth Surface Processes and Landforms, 42, 503–515.
Masao, F. T., Ichumbaki, E. B., Cherin, M., Barili, A., Boschian, G., Iurino, D. A., et al. (2016). New footprints from Laetoli (Tanzania) provide evidence for marked body size variation in early hominins. eLife, 5, e19568.
McHenry, H. M. (1991). Femoral lengths and stature in Plio-Pleistocene hominids. American Journal of Physical Anthropology, 85, 149–158.
Meldrum, D. J. (2004). Fossilized Hawaiian footprints compared with Laetoli hominid footprints. In D. J. Meldrum & C. E. Hilton (Eds.), From biped to strider (pp. 63–83). New York: Springer.
Menconero, S., Barili, A., Boschian, G., Cherin, M., Ichumbaki, E. B., Iurino, D. A., et al. (2019). 3D survey in extreme environment: The case study of Laetoli hominin footprints in Tanzania. In W. Börner & S. Uhlirz (Eds.), Proceedings of the 23rd international conference on cultural heritage and new technologies 2018 (pp. 1–20). Vienna: Museen der Stadt Wien, Stadtarchäologie.
Mollel, G. F., Swisher, C. C., III, Feigenson, M. D., & Carr, M. J. (2011). Petrology, geochemistry and age of Satiman, Lemagurut and Oldeani: Sources of the volcanic deposits of the Laetoli Area. In T. Harrison (Ed.), Paleontology and geology of Laetoli: Human evolution in context (Vol. 1, pp. 99–119). New York: Springer.
Musiba, C. M., Mabula, A., Selvaggio, M., & Magori, C. C. (2008). Pliocene animal trackways at Laetoli: Research and conservation potential. Ichnos, 15, 166–178.
Musiba, C., Mabulla, A., Mutakyahwa, M., Masao, F. T., Runyoro, V., Yul Kim, J., et al. (2012). Tanzania and the outstanding universal value of its paleoanthropology: Approaches at Laetoli and lessons learned. UNESCO World Heritage Series, 33, 115–125.
Plavcan, J. M., Lockwood, C. A., Kimbel, W. H., Lague, M. R., & Harmon, E. H. (2005). Sexual dimorphism in Australopithecus afarensis revisited: How strong is the case for a human-like pattern of dimorphism? Journal of Human Evolution, 48, 313–320.
Raichlen, D. A., & Gordon, A. D. (2017). Interpretation of footprints from Site S confirms human-like bipedal biomechanics in Laetoli hominins. Journal of Human Evolution, 107, 134–138.
Raichlen, D. A., Gordon, A. D., Harcourt-Smith, W. E. H., Foster, A. D., & Haas, W. R. (2010). Laetoli footprints preserve earliest direct evidence of human-like bipedal biomechanics. PLoS One, 5, e9769.
Reck, H., & Kohl-Larsen, L. (1936). Erster Überblick über die jungdiluvialen Tier- und Menschenfunde Dr. Kohl-Larsen’s im nordöstlichen Teil Des Njarasa-Grabens (Ostafrika) und die geologischen Verhältnisse des Fundgebietes. Geologische Rundschau, 27, 401–441.
Richmond, B., & Jungers, W. L. (1995). Size variation and sexual dimorphism in Australopithecus afarensis and living hominoids. Journal of Human Evolution, 29, 229–245.
Robbins, L. M. (1987). Hominid footprints from site G. In M. D. Leakey & J. M. Harris (Eds.), Laetoli: A Pliocene site in Northern Tanzania (pp. 497–502). Oxford: Clarendon.
Romano, M., Citton, P., Salvador, I., Arobba, D., Rellini, I., Firpo, M., et al. (2019). A multidisciplinary approach to a unique palaeolithic human ichnological record from Italy (Bàsura Cave). eLife, 8, e45204.
Rüther, H., Smit, J., & Kamamba, D. (2012). A comparison of close-range photogrammetry to terrestrial laser scanning for heritage documentation. South African Journal of Geology, 1, 1–14.
Schmid, P. (2004). Functional interpretation of the Laetoli footprints. In D. J. Meldrum & C. E. Hilton (Eds.), From biped to strider (pp. 49–62). New York: Springer.
Stern, J. T., & Susman, R. L. (1983). The locomotor anatomy of Australopithecus afarensis. American Journal of Physical Anthropology, 60, 279–317.
Stumpf, A., Malet, J. P., Allemand, P., Pierrot-Deseilligny, M., & Skupinski, G. (2015). Ground-based multi-view photogrammetry for the monitoring of landslide deformation and erosion. Geomorphology, 231, 130–145.
Tuttle, R. H. (1987). Kinesiological inferences and evolutionary implications from Laetoli bipedal trails G-1, G-2/3, and A. In M. D. Leakey & J. M. Harris (Eds.), Laetoli: A Pliocene site in Northern Tanzania (pp. 503–522). Oxford: Clarendon.
Tuttle, R. H. (2008). Footprint clues in hominid evolution and forensics: Lessons and limitations. Ichnos, 15, 158–165.
United Republic of Tanzania. (2004). The environmental management act. Dar es Salaam: Government Printing Press.
Villmoare, B., Hatala, K. G., & Jungers, W. L. (2019). Sexual dimorphism in Homo erectus inferred from 1.5 Ma footprints near Ileret, Kenya. Scientific Reports, 9, 7687.
Warrick, J. A., Ritchie, A. C., Adelman, G., Adelman, K., & Limber, P. W. (2017). New techniques to measure cliff change from historical oblique aerial photographs and structure-from-motion photogrammetry. Journal of Coastal Research, 33, 39–55.
Westoby, M. J., Brasington, J., Glasser, N. F., Hambrey, M. J., & Reynolds, J. M. (2012). ‘Structure-from-motion’ photogrammetry: A low-cost, effective tool for geoscience applications. Geomorphology, 179, 300–314.
White, T. D. (1980). Evolutionary implications of Pliocene hominid footprints. Science, 208, 175–176.
White, T. D., & Suwa, G. (1987). Hominid footprints at Laetoli: Facts and interpretations. American Journal of Physical Anthropology, 72, 485–514.
Zaitsev, A. N., Wenzel, T., Spratt, J., Williams, T. C., Strekopytov, S., Sharygin, V. V., et al. (2011). Was Sadiman volcano a source for the Laetoli footprint tuff? Journal of Human Evolution, 61, 121–124.
Zaitsev, A. N., Spratt, J., Sharygin, V. V., Wenzel, T., Zaitseva, O. A., & Markl, G. (2015). Mineralogy of the Laetolil Footprint Tuff: A comparison with possible volcanic sources from the Crater Highlands and Gregory Rift. Journal of African Earth Sciences, 111, 214–221.
Zimmer, B., Liutkus-Pierce, C., Marshall, S. T., Hatala, K. G., Metallo, A., & Rossi, V. (2018). Using differential structure-from-motion photogrammetry to quantify erosion at the Engare Sero footprint site, Tanzania. Quaternary Science Reviews, 198, 226–241.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2021 The Author(s)
About this chapter

Cite this chapter
Cherin, M. et al. (2021). Frozen in the Ashes. In: Pastoors, A., Lenssen-Erz, T. (eds) Reading Prehistoric Human Tracks. Springer, Cham. https://doi.org/10.1007/978-3-030-60406-6_8
Download citation
DOI: https://doi.org/10.1007/978-3-030-60406-6_8
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-60405-9
Online ISBN: 978-3-030-60406-6
eBook Packages: HistoryHistory (R0)